
Page 71
5
Empirical and Modeling Studies of Marine Reserves
~ enlarge ~
The goals for marine reserves, briefly described in Chapter 2, include conserving biodiversity, improving management of fisheries, and preserving and restoring habitat. These goals derive from societal wishes to preserve areas for the enjoyment of nature, to maintain functioning ecosystems, to establish replenishment zones for overexploited species, and to provide insurance against the uncertainty inherent in managing living natural resources. This chapter introduces the concepts underlying the use of reserves as a management strategy, describes empirical evidence from studies on existing reserves, and reviews various modeling studies.
CONCEPTUAL BASIS
Protect Intact Ecosystems
Current approaches to managing living marine resources typically address each species independently and ignore the spatial heterogeneity of marine systems. Consequently, policies may fail to protect some habitats and species. As a management tool, reserves have both disadvantages and advantages when applied to the diversity of marine species and habitats. The primary disadvantages are
1. establishing a reserve may result in the displacement of some fishers from customary fishing grounds with no impact on others, thus pitting the interests of one community against another;
Page 72
2. closing an area may lead to increased human impacts on open areas;
3. to protect some species effectively, a reserve would have to be so large that it would be infeasible to implement and enforce; and
4. reserves, in general, will not be effective without continued conventional management of the area outside the reserves.
The primary advantages of using a spatial approach are
1. reserves should be relatively simple and inexpensive to enforce once the boundaries are established and recognized;
2. regulations may be tailored to specific habitats within the jurisdiction of regional management authorities (e.g., zoning seagrass beds as off-limits to destructive fishing gears);
3. reserves support conservation of the full range of marine resources, including habitat, biological diversity, and exploited species such as commercial fish stocks;
4. reserves provide unique sites for education and research on marine ecosystems, especially for comparison to areas altered by human activities; and
5. reserves provide “control” areas for determining natural mortality rates for different life-history stages, rates that are critical variables in stock assessment models (Box 5-1).
However, a marine reserve is envisioned to play a role in the ecosystem on a scale larger than its boundaries (Agardy, 1994). Reserves that are intended to fill heritage needs—for instance, to protect endangered species, collapsed habitat, or special features—could also provide protection for other vulnerable species that may support the recovery of areas disturbed by human activities. Examples include reserves for a habitat such as the Oculina Banks coral beds of eastern Florida (Koenig et al., 2000) or reserves intended to protect a specific geological feature such as the Texas Flower Gardens coral reefs (Gittings and Hickerson, 1998).
Preserve and Restore Habitat
Both biological diversity and productivity are fundamentally dependent on habitat, and loss of habitat is the leading cause of declining biodiversity (Wilcove et al., 1998; Wilcove and Wilson, 2000). In concept, reserves can protect and restore habitats that are critical for living marine resources. Structurally complex biological habitats often shelter breeding aggregations, provide nursery habitat, and supply food for adults (Ebeling and Hixon, 1991; Lindholm et al., 1999). Studies of areas in which the structural integrity of the habitat has been lost typically show a clear reduction in biomass and biodiversity (Dayton et al., 1995; Morton, 1996; Watling and Norse, 1998; Lindeman and Snyder, 1999; Koenig et al., 2000).
Page 73
BOX 5-1Case Study for Red SnapperManagement of the red snapper (Lutjanus campechanus) fishery is perhaps the most contentious issue in the Gulf of Mexico because it involves two valuable fisheries in the region. Red snapper stocks have failed to recover from overexploitation. Some have attributed this failure to bycatch of juveniles in shrimp trawls, rather than fishing by directed commercial and recreational fisheries (Goodyear, 1995; Schirripa and Legault, 1999). Red snapper fishers suggest that an essentially unfished stock of fish exists offshore, where it is unavailable to the fishery (Schirripa and Legault, 1999). They have argued that this presumed offshore stock provides sufficient spawning activity and that the onus for recovery rests on shrimpers who need to decrease bycatch of young red snapper. Juvenile bycatch represents as much as 70% of the entire fishing mortality of this species (Nichols et al., 1990; Nichols and Pellegrin, 1992; Schirripa and Legault, 1999) although it comprises only 1% of the total incidental take in the shrimp fishery. However, the shrimp industry finds fault with National Marine Fisheries Service's (NMFS's) bycatch estimates and suggests that NMFS address alternative issues, including large-scale chronic (e.g., global warming) or acute (e.g., harmful algal blooms, hypoxia) events and the effects of the directed fishery on the age structure of the population. A recent peer review of the red snapper stock assessment indicated that little effective management could be accomplished until basic information on red snapper population dynamics was acquired (Stokes, 1997). For example, the level of bycatch reduction required for stock recovery is uncertain without accurate estimates of natural juvenile mortality rates. It is essential to determine the age structure of the red snapper population in order to estimate natural mortality rates. Although a number of analyses of the red snapper data incorporate changing rates of natural mortality, the level of uncertainty surrounding these values is high (Good-year, 1995, 1997; Schirripa and Legault, 1999), with some arguing for low levels, based on the red snapper's longevity (e.g., Camber, 1955; Wilson et al., 1994), and others for higher levels, based on the exclusion of regulatory discards from the samples (McAllister, 1997). The importance of bycatch reduction as a management strategy for red snapper recovery depends on the value used for the natural mortality rate (Stokes, 1997). One way to obtain more accurate estimates of natural mortality would be to establish reserves and determine the age structure in these unfished areas. Such reserves also would promote red snapper recovery by protecting a fraction of the stock from fishing. Hence, reserves could play dual roles in promoting stock recovery and providing better estimates for population parameters used in stock assessment. This could serve as an additional incentive for the fishing industry to work on reserves with managers and scientists to make management more effective. |
Reserves also can serve important functions in habitat restoration projects. Reserves act as controls (undisturbed sites) for evaluating the effectiveness of restoration projects at disturbed sites. Designation of areas as reserves could be used to protect sites so that restoration occurs naturally. There is a distinction
Page 74
between restoration of sites and mitigation that “creates” particular types of habitat where none existed previously. For example, artificial reefs placed on previously unstructured seabed and then protected from fisheries likely do not substitute for protecting natural ecosystems because functional biological communities also depend on the local oceanographic and environmental conditions and long-term interactions among species. Hence, artificial reefs may have little impact on overall productivity of reef fish populations and may in effect increase pressure on populations occurring on natural sites.
One category of habitat that has been severely reduced is coastal wetlands. Significant acreage of wetlands is still being lost each year to agricultural drainage, coastal development, oil spills, sewage, toxic chemicals, nonpoint source pollution, and destruction by introduced species. In California alone, nearly 90% of historical wetlands (once 3 million to 5 million acres, now estimated at 450,000 acres) are now used for agriculture and urban development, representing the greatest percentage loss in the nation. Thousands of acres of wetlands continue to be drained despite legislation (Food Security Act of 1985) that prohibits conversion of wetlands to agricultural use by landowners receiving federal farm payments. Recognition of the need to improve and restore estuarine habitat resulted in the creation of the National Estuarine Research Reserve System (NERRS) under the Coastal Zone Management Act (CZMA) of 1972. However, this program emphasizes developing and providing information to promote informed resource management rather than mandating specific protections (see Chapter 8).
The effects of habitat losses on fish stocks are diverse and ubiquitous, ranging from lower biodiversity in benthic habitats to reduced abundance of larger, economically important species (see Jones, 1992; Dayton et al., 1995; Kaiser, 1998; Pilskaln et al., 1998; Watling and Norse, 1998). Habitat loss from water diversion projects, hydropower dams, agricultural practices, and urbanization is a major factor contributing to declines in anadromous fish stocks (Rosenberg et al., 2000). Declines in shellfish and fish stocks may be caused directly by pollution or indirectly through algal blooms that either are toxic or deplete the water of oxygen (NRC, 2000a, b). More direct, fishing-related effects include destruction of biological structures (including oyster and coral reefs, sponge and bryozoan habitats) through abrasion and repetitive disturbance of mud-bottom communities by fishing trawls, dredges, or anchors, and loss of seagrass habitat from boating activities. Mitigating habitat loss will require water quality management and prohibition of activities that cause long-term declines in habitat quality and ecosystem health.
The movement toward an ecosystem approach to managing marine species gained momentum when habitat became a central theme of the Magnuson-Stevens Fishery Conservation and Management Act (MSFCMA) of 1996. This is embodied primarily in guidelines for applying the precautionary approach to management and recognizing the linkages between the goals of sustainable fish-
Page 75
ery production and conservation. The incorporation of concerns about essential fish habitat established the framework for an ecosystem approach to management, more akin to that developed for conserving terrestrial species (Zacharias et al., 1998).
Defining essential habitat is difficult because most marine and estuarine waters of the United States can be considered habitat essential to managed stocks and thus worthy of some level of protection. Structure and substrate in these habitats may vary from limestone outcroppings, to extensive seagrass beds, to kelp beds, to coral heads. Also important are mobile habitats, such as the extensive drifts of floating Sargassum in the Atlantic. Worldwide, many of these habitats have been declining for reasons ranging from nonpoint source pollution, to direct impact of fishing gear, to global climate change. In particular, loss of spawning habitat affects fish stocks in much the same way as fishing mortality of adult fish (Crowder et al., 2000). Hence, the dual mandates to adopt both precautionary and ecosystem approaches (FAO, 1995; NOAA, 1996a), coupled with that of defining habitat essential to sustain fisheries, make it imperative to evaluate habitat, determine its condition, and estimate its productivity. Life cycle patterns of the majority of economically important marine fish species can be reduced to several general habitat-related patterns (Koenig et al., 2000), some of which are quantifiable through development of abundance indices (Koenig and Coleman, 1998).
Establish Replenishment Zones
A marine reserve has the potential to play an important role in replenishing exploited marine ecosystems through the dispersal of larval or adult fish from the closed areas into regions where fishing is allowed (DeMartini, 1993). The idea that reserves will replenish fish stocks in open areas depleted by fishing is both promising (Carr and Reed, 1993; Quinn et al., 1993; Roberts, 1997a, Allison et al., 1998) and controversial (Coleman and Travis, in review). The promise lies in protecting fish so that they reach larger sizes, produce more offspring, and thereby increase the reproductive potential of a given species. One of the controversies, however, is that closed areas, in the absence of other measures to limit effort, may lead to increased fishing pressure operating outside reserve boundaries, such that the overall biomass of the stock decreases. In this circumstance, the anticipated benefits of the closure would not be realized, and there would be no net gain to the fishery.
Still, the idea of using closed areas as replenishment zones warrants investigation based on the biomass overflow and larval export hypotheses. The biomass overflow (or spillover) hypothesis suggests that the higher densities and greater average sizes of fish within a reserve will favor migration of adult fish into surrounding waters, thus augmenting or replenishing the population outside the reserve. Much of the empirical support for this hypothesis is based on anecdotal
Page 76
references. However, studies in the Philippines and Kenya documented increases in the populations of large adult fish in protected areas and subsequent population enhancement in adjoining regions (McClanahan and Kaunda-Arara, 1996; Russ and Alcala, 1996). Further evaluation in a variety of habitats with different species assemblages is required to determine whether spillover commonly provides significant replenishment for depleted fisheries.
The larval export hypothesis postulates that larvae will disperse out of the reserve and enhance recruitment in the fishing areas. Because reserves are associated with an increase in the density and size of fish and invertebrates, these populations should have higher spawning potentials and hence produce more eggs and larvae (Dugan and Davis, 1993; Bohnsack, 1996, 1998). If larvae disperse from the reserve into areas that provide appropriate nursery conditions, then stock enhancement in the open fishing areas is probable.
One of the better ways to replenish stocks and increase fishery yields is to protect stocks from growth overfishing. This is particularly difficult to do for species whose juveniles are caught incidentally in other fisheries. Designating nursery areas as reserves can protect juvenile fish from bycatch if the species are relatively sedentary during the juvenile stage. In the North Sea, large numbers of juvenile cod are killed annually in groundfish trawls (Horwood et al., 1998). Similarly, along the east coast of the United States, millions of dollars worth of yellowtail flounder (Pleuronectes ferrugineus) die each year as bycatch in trawl fisheries. In the Gulf of Mexico, nearly 70% of newly recruited juvenile red snapper are killed in shrimp trawls (Schirripa and Legault, 1999). If juveniles were protected in reserves, benefits would be delivered to the fishery when juveniles are exported from reserve areas to fishing grounds. Single-species closures used for plaice (Pleuronectes platessa) in the North Sea and mackerel (Scomber scombrus) in southwest England have resulted in increased yields by enhancing juvenile survival, even though neither area is completely closed (Horwood et al., 1998).
Provide Insurance Against Uncertainty
Insurance against the collapse of fish stocks is a primary, but elusive, goal of marine fishery management. However, managing fisheries and other living resources requires provisions for uncertainty in stock assessment (including limitations of the models and data used to estimate fish abundance) and for gaps in our understanding of the relative roles of environmental forces, fishing pressure, and management actions (see Chapter 3). Because uncertainty will always persist, fishery reserves have been proposed as a form of insurance to buffer potential failures of conventional management (Clark, 1996; Lauck, 1996; Lauck et al., 1998; Walters, 1998). In concept, protection of a substantial fraction of the fish stock in reserves will limit exploitation rates and prevent severe overfishing when other management measures fail (Bohnsack, 1998). In the event of collapse outside the reserve, a significant reproductive refuge would still be safe-
Page 77
guarded that should serve as a source of replenishment to the depleted area and could accelerate recovery. Such a refuge would be effective regardless of assessment uncertainties, thus adding robustness to management, although reserves probably would not be sufficient without additional controls (Allison et al., 1998).
Does hedging against uncertainty provide sufficient justification to establish reserves? Is it likely that this form of insurance, with its own uncertainties, will be acceptable or preferable to conservative catch targets and thresholds within the context of conventional fishery management? One limitation of regulating fishing activity based on stock assessment is the inability to anticipate, and then compensate for, natural fluctuations in populations. A natural downward shift in abundance may not be detected until the closure of a fishing season, when it is too late to adjust catch levels for the actual size of the stock. The need for insurance also depends on the success of conventional management in preventing overfishing. Thus, the incentive for using reserves to ensure against stock collapse depends on the characteristics of the fishery and the quality of stock assessments, to the extent that these will determine the relative efficacy and implementation costs of spatial closures versus conventional management for ensuring sustainability. Highly mobile species, for instance, may require such huge spatial closures to achieve even modest levels of stock protection (Walters, 2000) that more conventional approaches would be preferable (e.g., improved assessments, reduced quotas, tightened effort controls). However, there are situations in which conventional management tools are particularly limiting. When assessments are poor or unreliable and not likely to improve, conventional management approaches may carry unacceptably high risks. In these circumstances, spatial closures may be the most practical alternative to provide insurance against stock collapse due to overfishing.
Fishery management is not the only source of uncertainty in complex, poorly understood marine ecosystems. For example, many estuaries, coastal ecosystems, and coral reefs are threatened by a variety of anthropogenic activities that include shipping, dredging, petroleum extraction, shoreline development, wetlands destruction, river and stream diversion, exotic species introductions, and pollution. These direct threats to habitats and ecosystems can indirectly affect fishery yields (Agardy, 2000).
Global climate change and interdecadal climate events such as El Niño, also place stress on coral reefs (Glynn, 1984, 1991; Glynn and D'Croz, 1990; Brown, 1997), further reducing the resiliency of reefs and other marine ecosystems to human impacts. It is now apparent that many marine organisms, especially those restricted to small geographic domains, face the threat of major declines or even extinction (Brander, 1981; Carlton, 1993; Casey and Myers, 1998; Roberts and Hawkins, 1999).
Will MPAs help protect against these stresses? In many cases marine zoning can reduce or offset the effects of shipping, dredging, and petroleum extraction, while more stringent regulation can reduce the effects of development and
Page 78
pollution. However, MPAs cannot protect an area from global warming or other large-scale climatic events (Allison et al., 1998), nor do they provide refuge from invasion by nonindigenous species (Simberloff, 2000). Although marine ecosystems are resilient and survive both short- and long-term climate variability as well as anthropogenic activities, resiliency has its natural limits (NMFS, 1999). Increasing human use, extraction, and modification of marine ecosystems have elevated the risk of long-term damage and provided the impetus to create MPAs as protection against unchecked expansion of human activities in marine ecosystems. In the wake of the catastrophic Santa Barbara oil spill in 1969, Congress passed the National Marine Sanctuary Act for “the purpose of preserving or restoring such areas for their conservation, recreational, ecological or esthetic values.” Most commercial or recreational activities remain unrestricted in the sanctuaries, but the program establishes a framework for increasing the protection of vulnerable marine habitat.
Marine reserves could provide a mechanism for ensuring preservation of habitat areas of particular concern (HAPCs). The HAPC designation, developed by the National Marine Fisheries Service, refers to habitat judged to be particularly important to the long-term productivity of populations of managed species, to be especially vulnerable to degradation, or to represent examples of rare habitat types. Habitats with this designation include the Florida Middle Grounds in the Gulf of Mexico, which contains stony and octocoral habitat; the Oculina Banks in the South Atlantic, composed of thickets of fragile ivory tree coral Oculina varicosa; and a proposed HAPC in the gravel-cobble pavements in Closed Area II on Georges Bank in New England.
Distinguish Natural Variability from Human Impacts
Marine ecosystems are characterized by spatial and temporal variability that occurs at scales of centimeters to hundreds of kilometers and minutes to centuries. Global climate change and natural events that occur episodically over several years or decades, such as El Niño or other regime shifts, can result in long-lasting shifts in ecosystem composition and function. On a more local scale, the introduction of an invasive, nonnative species can affect an entire bay or large regions of coastline. Understanding the influence of human actions on marine systems is critical to evaluating the need for and effectiveness of management actions, but differentiating between natural and anthropogenic events is extremely difficult (NRC, 2000a). Any indicator of change in a system must be compared to a well-defined natural standard, or benchmark, against which the magnitude of the change can be evaluated to determine its cause and significance. Without control areas, such as reserves, that are relatively free from human influence to compare with areas altered by human activities, explaining the sources of variability becomes even more difficult. For example, much of what we know about fish populations is derived from fishery landings and is
Page 79
inherently biased, because estimates of fishing effort do not provide an index of fish abundance, and fishing changes the population dynamics of the target and associated species (NRC, 1998a). There is a significant need for fishery-independent sampling programs that include areas closed to fishing and other activities that disturb fish populations and habitat.
EMPIRICAL EVIDENCE AND INFERENCES FROM MODELS
Changes Within Marine Reserves
The finding that marine reserves improve species' abundance, biomass, and diversity came about almost by accident. Initial experience with reserves came from university research sites in Sumilon Island in the Philippines (Russ and Alcala, 1994), Leigh Marine Reserve in New Zealand (Walls, 1998), and Las Cruces in Chile (Castilla and Durán, 1985). In these research sites, fishing was prohibited to protect experiments from interference and to investigate effects of humans on coastal systems. However, exclusion also resulted in a dramatic transformation of the resident ecological communities. Careful studies at these reserves allowed researchers to gather quantitative evidence that closed areas could be used in marine conservation to restore or maintain a more natural ecosystem structure and to facilitate local recovery of depleted fish stocks (Russ, 1985; Ballantine, 1991; Castilla, 1999).
These initial studies indicated that establishing reserves had diverse effects, including rapid buildup in biomass (numbers and average body size) of previously exploited species and increased species richness (number of species per unit area)—that is, species appeared that had been absent prior to establishment of the reserve.
Effects on Abundance, Body Size, and Biomass
Halpern (in review) recently analyzed 76 studies of reserves covering a wide variety of locations, conditions, and enforcement levels. Most of these studies were conducted in newly formed reserves, so the effects represent early rather than long-term results. Because of the diverse taxa represented and the diverse methods of data presentation, Halpern derived a single aggregate measure to evaluate the results of each study. Many of the studies relied on “snapshot” comparisons of reserves with nearby unprotected sites, so the rate of change could not be determined. In any reserve, the composition and density of species in the area are expected to continue to change over time. However, in 69% of the reports, Halpern found overall increases in the densities of resident animals. Averaged across all of the studies, the density in reserves was almost twofold higher than in the unprotected sites. In general, local density appeared to increase with the length of time an area was closed (Table 5-1).
Page 80
|
Leigh Marine (New Zealand) |
Apo Island (Philippines) |
Saba Marine Park (Caribbean) |
St. Lucia |
|
|
Density |
8 |
8 |
1.4 |
NA |
|
Years closed |
15 |
11 |
4 |
|
|
Biomass |
NA |
8 |
4 |
3 |
|
Years closed |
9 |
11 |
3 |
|
|
Species |
Lobster |
Predatory fish |
5 exploited fish families |
Haliotis kamtscharkana |
|
Reference |
MacDiarmid and Breen, 1992 |
Russ and Alcala, 1996 |
Polunin and Roberts, 1993 (Roberts unpublished data) |
Wallace, 1999 |
NOTES: Density and biomass values represent increases relative to the density of comparable populations in neighboring areas open to fishing. NA = not available.
Page 81
Ruckleshaus and Hays (1998), in analyzing similar pre- and post-closure changes in abundance in five reserves, found that reserves did not result in increased density of every species. Their analysis indicates that changes in community structure after the establishment of a reserve will not necessarily result in the increased density of all species affected by fishing.
In addition to abundance, the average size of individuals of a given species can change significantly within a reserve. In Halpern's aggregate analysis of reserve impacts, 88% of the studies found that the average body size of fish increased. Averaged across all target species and studies, the body size increased by about one-third (Halpern, in review).
Some long-lived species require decades to attain maximum size. Hence average body size within a reserve could continue to increase for many years following protection. Such increases have been observed after 15 years of protection in Egypt's Ras Mohammed Marine Park, where the average body weight of the lunar-tail grouper (Variola louti) increased three-fold in reserves compared to adjacent fished waters (Roberts and Polunin, 1993b). Similar findings were reported for northern abalone (Haliotis kamtschatkana) populations off Vancouver Island (Wallace, 1999); for quahog clams (Mercenaria mercenaria) within a closed area in Rhode Island (Rice et al., 1989); and generally for many fish species in reserves in New Zealand (McCormick and Choat, 1987) and the Mediterranean (Bell, 1983).
Biomass provides a more sensitive measure of reserve effects because it integrates changes in abundance and body size. For example, in and around Belize's Hol Chan Marine Reserve, fish densities increased in only a few families while fish biomass increased significantly after three years of closure (Polunin and Roberts, 1993; Carter and Sedberry, 1997). Biomass of economically important fish species increased two-fold in peripheral parts of the reserve and nine-fold in the center of the reserve compared to fished areas (Roberts and Polunin, 1994). Halpern (in review) found increases in 92% of the studies he reviewed, with the aggregated biomass in reserves on average two and a half times greater than in fished areas.
Effects on Species Diversity
Biodiversity encompasses all levels of organizational complexity, from genetic diversity to species diversity to ecosystem diversity (Chapin et al., 2000). Species diversity is the most commonly monitored aspect of biodiversity used in evaluating conservation areas. The concept of species diversity can be broken down into at least three components: (1) species richness, or the number of species per unit area; (2) species evenness, or the relative abundances of the range of species present; and (3) species assemblages, or the interactions between species (e.g., trophic structure). Changes in the assemblages or diversity
Page 82
of species can serve as key indicators of the impact of a reserve on ecosystem functioning.
In existing reserves, the point diversity of fish (i.e., the number of species per unit area of reef) generally increased when reserve sites were established in heavily exploited regions (e.g., Roberts and Polunin, 1993a; Russ and Alcala, 1996; Wantiez et al., 1997; Watson et al., 1997). Halpern (in review) found that 59% of the studies reported greater species richness in reserves. When averaged across studies, species richness had increased by one-third. In some cases, the apparent increased point diversity may have reflected an increase in abundance and hence an increase in the probability of observation, rather than the reappearance of a species in that area. Reserves may improve the long-term viability of some rare populations, especially for species that require relatively high densities for successful reproduction.
Expecting increases in all species is unrealistic given the complex linkages in ecological communities (Pimm, 1982), especially predator-prey relationships. Thus, species diversity may not be the best indicator of reserve performance. For instance, the density of some species may decline in an area closed to fishing (Ruckelhaus and Hays, 1998). Koenig et al. (2000) found that species diversity in the Oculina Reserve appeared lower when top-level predators were present than it did after removal of large predatory fish. In addition to real decreases in prey abundance from predation by species protected in the reserve, predator avoidance behavior may contribute to the apparent decrease. The effect of such avoidance behavior on assessments of species diversity will be particularly difficult to determine in deepwater habitats where census methods are performed remotely.
Establishing reserves in rocky intertidal habitats in Chile facilitated the recovery of the overexploited tunicate Pyura chilensis and the snail Concholepas concholepas. However, cascading trophic effects from the recovery of the snail resulted in lower overall diversity in this area (Davis, 1995; Castilla, 1999). These reports also documented the reduced abundance of mussels and herbivorous gastropods, and increased cover of algae or barnacles (Davis, 1995; Castilla, 1999). The Chilean studies illustrate how the trophic balance of an ecosystem shifts both with fishing and with subsequent protection, potentially benefiting some species at the expense of others.
Changes in species diversity associated with a closure will depend on the level of exploitation, life-history characteristics of the individual species, and potential for replenishment from surrounding areas. A common goal of reserves is to foster the recovery of species depleted directly or indirectly by intense fishing pressure (Fogarty et al., 2000). Species that are more vulnerable to exploitation and extirpation because of certain life-history characteristics (Table 5-2) are likely to benefit most from protection (Roberts and Hawkins, 1999). For example, the Looe Key National Marine Sanctuary in Florida supports six species of economically valuable fish that are absent from surrounding exploited
Page 83
|
Vulnerability |
||
|
Characteristics |
High |
Low |
|
Population Turnover |
||
|
Longevity |
Long |
Short |
|
Growth rate |
Slow |
Fast |
|
Natural mortality rate |
Low |
High |
|
Production biomass |
Low |
High |
|
Reproduction |
||
|
Reproductive effort |
Low |
High |
|
Reproductive frequency |
Semelparity |
Iteroparity |
|
Age or size at sexual maturity |
Old or large |
Young or small |
|
Sexual dimorphism |
Large differences in size between sexes |
Does not occur |
|
Sex change |
Occurs |
Does not occur |
|
Spawning |
In aggregations at predictable locations |
Not in aggregations |
|
Allee effectsa at reproduction |
Strong |
Weak |
|
Capacity for recovery |
||
|
Regeneration from fragments |
Does not occur |
Occurs |
|
Dispersal |
Short distance |
Long distance |
|
Competitive ability |
Poor |
Good |
|
Colonizing ability |
Poor |
Good |
|
Adult mobility |
Low |
High |
|
Recruitment by larval settlement |
Irregular and/or low level |
Frequent and intense |
|
Allee effects a at settlement |
Strong |
Weak |
|
Range and Distribution |
||
|
Horizontal distribution |
Nearshore |
Offshore |
|
Vertical depth range |
Narrow |
Broad |
|
Geographic range |
Small |
Large |
|
Patchiness of population within range |
High |
Low |
|
Habitat specificity |
High |
Low |
|
Habitat vulnerability to destruction by people |
High |
Low |
|
Commonness and/or Rarity |
Rare |
Abundant |
|
Trophic Level |
Rare |
Low |
aAllee effects occur when a reduction in population density has significant impacts on reproduction.
SOURCE: Roberts and Hawkins, 1999.
Page 84
areas (Clark et al., 1989). In St. Lucia, some species are observed only in protected reserves (Roberts and Hawkins, 1997). However, there is no guarantee that a species will recover in a reserve if it has been eliminated regionally as a consequence of widespread fishing. The recovery of animals in a reserve in Jamaica's Discovery Bay is constrained by the virtual absence, to date, of recruitment by large snappers and groupers. In contrast, population rebounds have been observed for some species of smaller fishes that persisted in the area despite intensive exploitation for more than a century (Watson and Munro, in press). Hence, recovery in a reserve depends on the presence of source populations within the dispersal range of the depleted species and is especially critical for long-lived species (Roberts, in press). Recovery will be more rapid and assured when a reserve is established before exploitation essentially removes the entire breeding stock.
Contribution of Reserves to Replenishing Surrounding Areas
The contribution of reserves to replenishing fish stocks depends on the export of adults or young (larval or juvenile) recruits to fishing areas. Potentially, the more intense the exploitation outside the reserve, the greater is the density difference that will develop between protected and fished populations. The expectation is that the relative contribution of reserves to recruitment will be higher for intensively fished, non-migratory stocks. There are few studies demonstrating the replenishment of fish stocks on fishing grounds via export from reserves (Bustamante and Castilla, 1990; Tegner, 1993; Roberts, 1995). In some cases, this lack of data feeds the skepticism of the fishing community, but for some intensively fished species, models indicate that reserves have a high probability of increasing yields (see Attwood and Bennett, 1994; Murawski et al., 2000). An overview of studies of the effects of reserves on fisheries is presented in Table 5-3.
Replenishment from Spillover of Adult Fish
Direct confirmation of the export of fish from reserves to open fishing grounds is difficult and may require long time series to detect changes due to reserves over the normal fluctuations in the abundance of fish populations. A few mark-recapture studies show that significant numbers of fish tagged within reserves were caught in the adjacent fishing grounds and that fishing efficiency (catch per unit effort) increased outside the reserve boundaries (Attwood and Bennett, 1994; McClanahan and Kaunda-Arara, 1996; Johnson et al., 1999). Because such studies are so logistically difficult and costly, models show greater potential to improve our understanding of how marine reserves function in a regional context (see Gerber et al., in review). However, the conclusions drawn from a modeling effort are limited by the underlying assumptions on which the
Page 85
|
Park |
Source |
Species Abundance |
CPUE |
Species Richness |
Shannon-Weiner Diversity |
Biomass |
Size |
Number of Fish |
Density |
Fishery |
Design |
|
Kisite Marine National Park, Kenya |
27, 28 |
⇑ |
↔ |
↔ |
⇑ |
⇑ |
I/O, A |
||||
|
Cerbere-Banyuls, France |
9 |
⇓ |
⇑ |
↔ |
I/O, A |
||||||
|
Goat Island, New Zealand |
8 |
↔ |
↔ |
I/O, A |
|||||||
|
Five Islands in New Caledonia |
26 |
⇑ |
⇑ |
↔ |
⇑ |
B/A, I/O, A |
|||||
|
Anse Chastanet |
21 |
⇑ |
I/O, A |
||||||||
|
Apo Island, Philippines |
1, 2, 24 |
⇑ |
⇑ |
⇑ |
Temp |
||||||
|
Sumilon Island, Philippines |
1, 2, 24 |
⇑ |
⇑ |
⇑ |
Temp |
||||||
|
Maria Island, Tasmania |
10 |
↔ |
↔ |
BACI |
|||||||
|
Medes Islands |
11 |
⇑ |
↔ |
⇑ |
I/O, A |
||||||
|
Okakari Point to Cape Rodney Marine Reserve, New Zealand |
18 |
⇑ |
I/O, A |
Page 86
|
Barbados Marine Reserve |
20 |
⇑ |
⇑ |
I/O, A |
|||||||
|
De Hoop Marine Reserve, South Africa |
3, 4 |
⇑ |
I/O, A, Model |
||||||||
|
Tsitsikamma Coastal National Park, South Africa |
5, 6 |
I/O, A |
|||||||||
|
Okakari Point Marine Reserve, New Zealand |
18 |
⇑ |
I/O, A |
||||||||
|
Soufriere Marine Management Area, St. Lucia |
12 |
↔ |
Temp |
||||||||
|
Cousin Island Nature Reserve, Seychelles |
14, 15 |
⇑ |
⇑ |
⇑ |
I/O, A, Grad |
||||||
|
Sainte Anne Marine National Park, Seychelles |
15 |
⇑ |
⇑ |
Grad |
|||||||
|
Baie Ternay Marine National Park, Seychelles |
15 |
? a |
? a |
⇑ |
Grad |
||||||
|
Curieuse Marine National Park, Seychelles |
15 |
? a |
? a |
⇑ |
Grad |
Page 87
|
Mobasa Marine National Park, Kenya |
17 |
⇑ |
⇑ |
⇑ |
⇓ |
I/O, A |
|||||
|
Saba Marine Park, Netherlands, Antilles |
19, 23 |
⇑ |
⇑ |
⇑ |
Temp, A |
||||||
|
Hol Chan Marine Reserve, Belize |
19, 23 |
⇑ |
⇑ |
⇑ |
I/O, A |
||||||
|
Ras Mohammed Marine Park, Egypt |
22, 23 |
↔ |
↔ |
I/O, A |
|||||||
|
Exuma Cay Land and Sea Park, Bahamas |
25 |
⇑ |
⇑ |
I/O, A b |
|||||||
|
Mayotte Island, Indian Ocean |
16 |
⇑ |
⇑, ⇓ |
⇑ |
I/O, A |
||||||
|
Carrie-le-Rouet, France |
13 |
⇑ |
⇑ |
I/O, A |
|||||||
|
Looe Key National Marine Sanctuary, Florida |
7 |
⇑ |
⇑ |
Temp |
NOTES: a Unregulated reserve used as a reference site. b Only Nassau grouper (Epinephelus striatus), investigated. A: After reserve formation. B: Before reserve formation. BACI: Before/after control/impact design. I/O: Inside/outside reserve comparative study. Grad: Fishing intensity gradient design. Model: Model used to calculate fishery benefits. Temp: Temporal design, sampling before and after reserve formation. CPUE: Catch-per-unit-effort. ⇑ = increase, ⇓ = decrease, ↔ = no change.
SOURCE CODES: 1 = Alcala, 1988; 2 = Alcala and Russ, 1990; 3 = Bennett and Attwood, 1991; 4 = Bennett and Attwood, 1993; 5 = Buxton, 1993; 6 = Buxton and Smale, 1989; 7 = Clark et al., 1989; 8 = Cole et al., 1990; 9 = Dufour et al., 1995; 10 = Edgar and Barrett, 1997; 11 = Garcia-Rubies and Zabala, 1990; 12 = Goodridge et al., 1997; 13 = Harmelin et al., 1995; 14 = Jennings et al., 1995; 15 = Jennings et al., 1996; 16 = Letourneur, 1996; 17 = McClanahan and KaundaArara, 1996; 18 = McCormick and Choat, 1987; 19 = Polunin and Roberts, 1993; 20 = Rakintin and Kramer, 1996; 21 = Roberts and Hawkins, 1997; 22 = Roberts and Polunin, 1993b; 23 = Roberts and Polunin, 1993a; 24 = Russ and Alcala, 1996; 25 = Sluka et al., 1997; 26 = Wantiez et al., 1997; 27 = Watson et al., 1996; 28 = Watson and Ormond, 1994.
Page 88
model is based, particularly by assumptions about the degree of exchange among the reserves within a network or between reserves and the exploited habitats surrounding them (Palumbi, 2000).
Models have been developed to examine the potential contribution of dispersal from reserves to a fishery. For species with low adult mobility, individual size and population abundance should increase within the reserve, but with minimal spillover to adjacent areas. Indeed, Zeller and Russ (1998) found low rates of movement of the site-attached adult coral trout (Plectropomus leopardus) across reserve boundaries. At the other extreme, high rates of adult mobility between reserves and unprotected areas may drain the reserve population, especially when reserves are small. Fisheries for species with moderate rates of dispersal are predicted to benefit most from the establishment of reserves (Hastings and Botsford, 1999).
Effects of Reserves on Reproductive Capacity
The size of individuals and the abundance of target species frequently increase in reserves, yielding a higher reproductive capacity. Because fecundity (number of eggs produced) increases rapidly with weight in fish, the reproductive output of fish in protected areas should be significantly higher than that in areas where fishing removes a proportion of the larger, older fish. In actuality, fecundity increased two- to three-fold in two species of rockfish in a California reserve (Paddack, 1996) and six-fold for Nassau grouper (Epinephelus striatus) inside the Bahamas Exuma Cays reserve (Sluka et al., 1997).
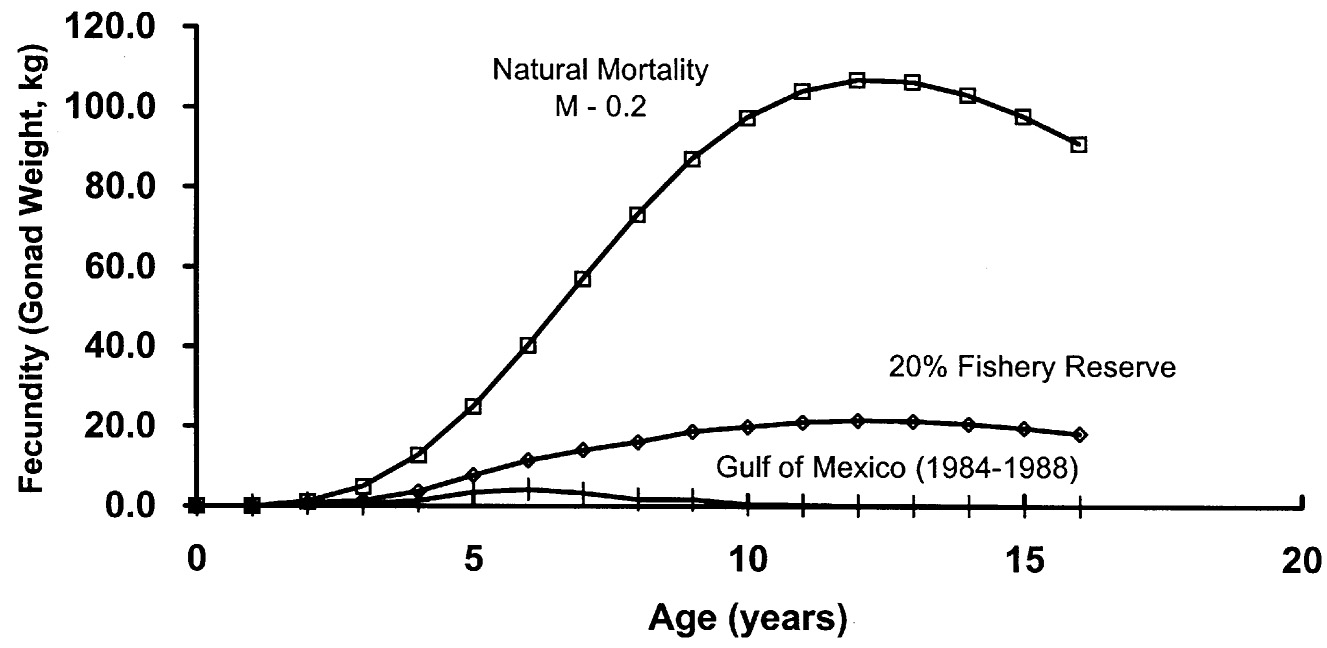
In a simple modeling exercise, Bohnsack (1992) examined egg production by red snapper (Lutjanus campechanus) in the Gulf of Mexico with and without a 20% network of reserves. He estimated that closing 20% of the fishing grounds would increase egg production by approximately twelve-fold over the exploited stock (see Figure 5-1). The increase in reproduction derives from increased abundance of larger, older, more fecund females in the population.
Characteristics of Larval Dispersal
Most marine species exhibit considerable interannual recruitment variability (Dayton and Tegner, 1990). Dispersal distance depends upon time spent in the water column and oceanic transport (Hjort, 1914). The time spent in the water column depends on the time required for larvae to grow to settlement size, which is influenced by temperature and food availability (Bingham, 1992; Maloney et al., 1994). Hjort (1914) proposed the “critical period” for fish larvae as the time of first feeding coupled to successful dispersal to appropriate settling sites. It is during this time that fish must encounter prey in sufficiently high densities to ensure larval growth and survival. The longer they remain small, the longer they will be vulnerable to predation (Houde, 1987, 1997). In many cases, the survival
Page 89
~ enlarge ~
FIGURE 5-1 Estimated red snapper total fecundity (gonad weight) under conditions of natural mortality, average fishing mortality observed in the Gulf of Mexico between 1984 and 1988, and under conditions of a simulated reserve system that protects 20% of the population. The reserve system assumes that 20% of the cohort experiences only natural mortality, while the remaining 80% experiences the above fishing conditions. Used with permission from the author (Bohnsack, 1992).
of entire cohorts depends on weather and oceanographic variability that may either concentrate or disperse prey (Lasker, 1975), providing a biological explanation for the importance of the presettlement period (Jackson and Strathmann, 1981; Maloney et al., 1994). Recent results suggest that it is not possible to develop a generalized rule for larval dispersal or retention (Jones et al., 1999; Swearer et al., 1999; Cowen et al., 2000).
Many larvae, especially invertebrate larvae, require chemical cues to initiate settlement and metamorphosis. Suitable settlement sites that provide these cues are typically biogenic habitats—corals, algae, eelgrass, bryozoans—that also provide refugia and food for juveniles (Dayton et al., 2000). When these kinds of habitat are limiting, recruitment bottlenecks occur (Dayton et al., 1995; Lipcius et al., 1997; Bohnsack, 1998). Recruitment success in areas within and outside reserves will depend on the availability of suitable settlement sites.
Closed areas in the New England fishing grounds provide evidence of improved recruitment for some species. On Georges Bank and in the Gulf of Maine, more than 5000 nmi2 were closed to bottom trawling and dredging in December 1994 in response to the critical decline of groundfish stocks. The intent was to improve recruitment by reducing bycatch of juveniles and preventing the disturbance of juvenile habitat in the closed areas. Collie et al. (1997) and Fogarty and Murawski (1998) offer excellent reviews of the history of direct and indirect impacts of fishing on Georges Bank where the cumulative impacts
Page 90
|
Genus and Species |
Common Name |
Average Dispersal (km) |
Evidence |
Reference |
|
Echinometra meethaii |
Pacific urchins |
20 |
Genetics |
Palumbi, in review |
|
Solea vulgaris |
Common sole |
30 |
Genetics |
Palumbi, in review |
|
Riftia pachyptila |
Tubeworm |
20 |
Genetics |
Palumbi, in review |
|
Littorina cingulata |
Gastropod |
4 |
Genetics |
Johnson and Black, 1998 |
|
Littorina littorea |
Gastropod |
20 |
Invasion rate |
Carlton, 1982 |
|
Elminius |
Barnacle |
40 |
Invasion rate |
Crisp et al., 1958 |
|
Carcinus mineas |
Green crab |
20 |
Invasion rate |
Crisp et al., 1958 |
|
Cancer magister |
Dungeness crab |
50 |
Settlement pattern |
Botsford et al., 1997 |
had been extremely damaging to demersal fish species and sea scallops (Placopecten magellanicus). They concluded that the three large closed areas aided recovery of yellowtail flounder (Limanda ferrugineus) and dramatically improved the abundance of large scallops (Collie et al., 1997; Fogarty and Murawski, 1998; Murawski et al., 2000). Scallop biomass tripled in the first 20 months after closure of the areas on Georges Bank (Anderson et al., 1999). Importantly, the current patterns and clustering of scallop fishing vessels at the downstream margins of the closed areas suggest that scallop larvae are exported and recruit to downstream areas (Collie et al., 1997; S.A. Murawski, National Marine Fisheries Service, personal communication, 1999).
Larval Dispersal Distances
Dispersal is facilitated for marine species having a planktonic larval stage. In fact, larvae of coastal species frequently appear in mid-ocean plankton samples (Scheltema, 1986). Most estimates of larval dispersal are obtained indirectly (Levin et al., 1993), derived from inferences about oceanography (Lee et al., 1994), larval biology (Emlet et al., 1987) or the genetics of adult populations (Palumbi, 2000; see Table 5-4). There are few direct observations of larval dispersal distances, except for species with low rates of dispersal (Olson, 1985; Stoner, 1992). Tegner (1992, 1993), after transplanting green abalone to a protected area depleted of adult stock, found larval recruitment significantly enhanced, with local recruitment observed from the transplants. This experiment terminated prematurely when the transplanted abalone were collected illegally.
Page 91
Theoretically, species with relatively long larval periods (extending from a week to several months), experiencing current speeds of 0.1 m/s, could disperse more than 100 km downstream (Boehlert, 1996; Roberts, 1997b; Grantham et al., in review). However, recent reports suggest that average dispersal in some marine species is low and, hence, that long-distance dispersal may be less common at ecological time scales (Palumbi, 2000). For instance, species in many benthic habitats, such as sponges, several seaweeds, ascidians, and corals, have non-planktonic or demersal larvae and thus limited dispersal abilities. Also, long-term persistence of populations of marine species endemic to extremely small and isolated islands indicates that local retention must be highly significant (Hawkins et al., 2000). In simulation studies where larval movement is assumed to be passive, the results indicate that transport may be affected strongly by local eddies and current reversals (Lee et al., 1994; Limouzy-Paris et al., 1997), with limited dispersal from release points (McShane et al., 1988; Sammarco and Andrews, 1988). In his Caribbean study, Roberts (1996) concluded that surface currents might define dispersal patterns, with very large, often order-of-magnitude differences possible between the upstream and downstream settlement densities. Models of larval dispersal in reef habitats suggest that local retention on a natal reef (i.e., spawning site) is 10 times more likely than transport to downstream reef sites unless the spacing between reefs is about the same as reef diameter (Black, 1993). These studies indicate that knowledge of the dispersal patterns for larval replenishment will be important for effective reserve design and placement (see Chapter 6).
The issue of long-distance dispersal or local retention plays an important role in guiding decisions about the design of MPAs and reserves. Low transport rates for larvae of coral reef fish were determined using two methods: (1) micro-constituent analysis of otoliths in the U.S. Virgin Islands (Swearer et al., 1999), and (2) a mark-and-recapture tagging study involving millions of fish larvae around Lizard Island on the Great Barrier Reef (Jones et al., 1999). In both cases, a large proportion (roughly 50%) of larvae appeared to stay in local waters and recruit back to the home ranges of their parents. These studies are described in more detail in Chapter 7.
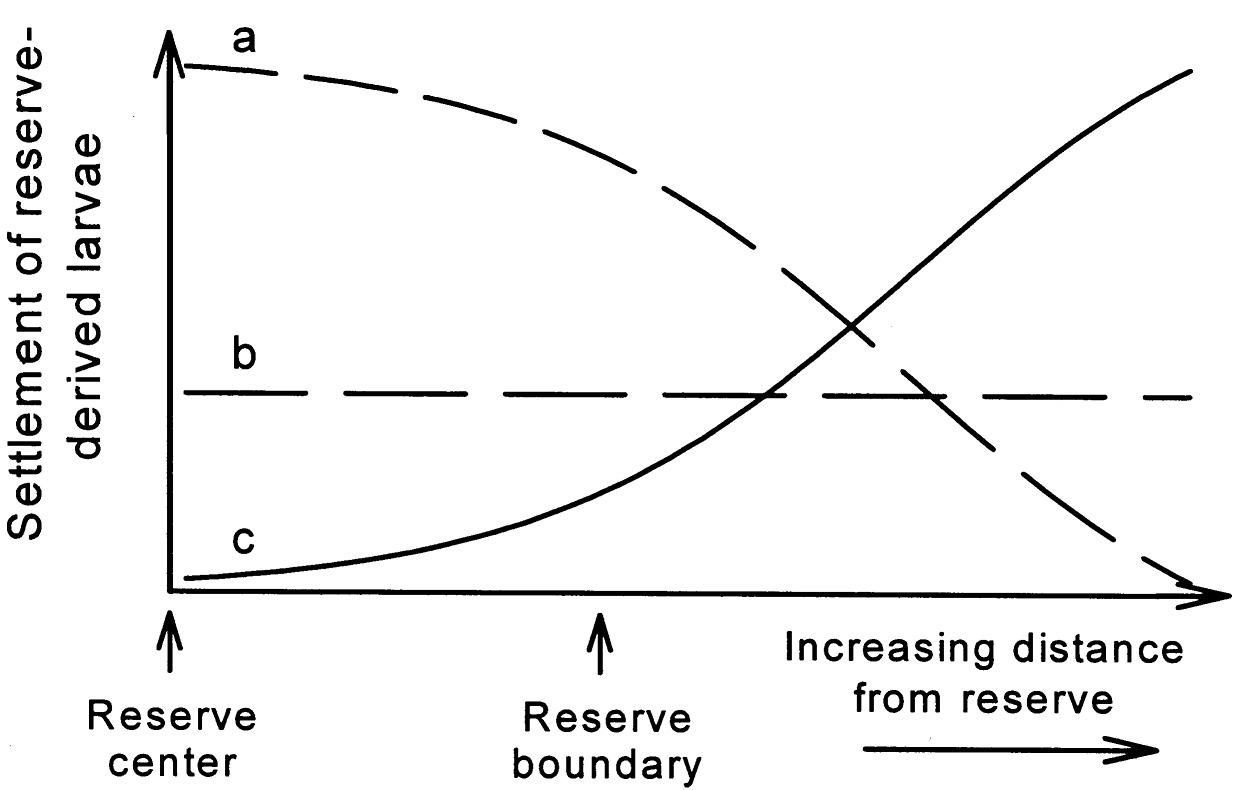
In other studies, local retention was rarer (e.g., Bertness and Gaines, 1993), but the fraction of larvae dispersing long distances was not determined. Larvae of species in protected areas appear to be more abundant just outside the protected area boundaries than at more remote sites, based on empirical and modeling results (Black, 1993; Palumbi, in review, 2000). If larval dispersal is low, even for species with planktonic larvae, then marine species, including some fishes, may require management on much finer spatial scales than previously assumed. Dispersal distances will affect the larval spillover distances to surrounding areas with implications for designing reserve networks that are self-sustaining (Figure 5-2).
Page 92
~ enlarge ~
FIGURE 5-2 Three different dispersal scenarios and their implications for export of eggs and larvae from reserves: (a) short-distance dispersal leading to significant local retention and low dispersal away from reserve; (b) mixed strategy with both long- and short-distance dispersal leading to significant retention and export; (c) long-distance dispersal with low local retention and significant export. Reproduced from Roberts, 2000.
Genetic Analysis of Dispersal
In general, high dispersal potential is associated with high gene flow and hence low levels of genetic differentiation between populations over scales of hundreds to thousands of kilometers (Palumbi, 1992, 1994). Therefore, the level of genetic differentiation in adult populations can be used as an indication of the long-term dispersal history of a species. When a population is genetically distinct, this is an unambiguous indication of isolation in terms of recruitment from other populations. However, because a low level of gene flow is sufficient to maintain genetic homogeneity, the lack of genetically distinct populations does not necessarily correlate with high levels of interannual dispersal.
Johnson and Black (1998) documented an increase in genetic differentiation with geographic distance over a few hundred kilometers by measuring genetic relatedness in marine species over fine spatial scales. Palumbi (in review) used a similar approach to estimate average larval dispersal for a variety of marine species. These values ranged from 10 to 50 km and are similar to values estimated from the spread of invasive marine species (Crisp, 1958; Shanks and Grantham, in review) and from settlement patterns along coastlines (Botsford et al., 1997; see also Table 5-4).
Page 93
Recruitment Sources and Sinks
The capacity of a protected area to replenish populations both inside and outside its boundaries may depend on whether the area will serve as a source or sink for new recruits (Allison et al., 1998). Sources produce “excess” recruits that spill over into surrounding areas, while sinks receive recruits but produce few of their own. In essence, a source is a productive habitat. Lipcius et al. (1997) have successfully measured sources and sinks for the spiny lobster (Panulirus argus) by measuring adult abundance, post-larval supply, juvenile density and nursery habitat. Evaluating these factors is critical for choosing a marine reserve site that will function as a source for the surroundings. Poor habitats are likely to be sinks, but often are suggested as potential sites for reserves because, typically, there will be fewer conflicts with users, especially fishers (Crowder et al., 2000; Dayton et al., 2000). However, locating a reserve in a sink area may increase fishing pressure on source populations and thus exacerbate population decline (Crowder et al., 2000).
The contribution of a reserve (located in a source area) to a fishery can be estimated conservatively as the fraction of the total biomass of the stock that resides in the reserve. There are several circumstances under which the reserve's contribution to recruitment is likely to be greater than this fraction:
1. Where a fishery removes most fish before they have attained sexual maturity: For example, in the fishery for red hind (Epinephelus guttatus) in Puerto Rico, 95% of the catch consists of immature fish (Sadovy, 1994). In this case, the reproductive potential within a reserve would exceed the potential in the surrounding fished areas. One way to estimate the reserve contribution to recruitment would be to calculate the reproductive biomass in reserves as a fraction of the total reproductive biomass in reserves plus fishing grounds.
2. Where a species experiences strong Allee effects at spawning: In this case, high population densities are necessary to trigger spawning and achieve fertilization. Allee effects are especially prevalent in sessile or sedentary invertebrates (Lillie, 1915; Levitan et al., 1992; Levitan, 1998; Dayton et al., 2000), such as scallops, clams, sea urchins and abalone. For such species, reserves may represent the only places where sufficiently high densities exist for successful spawning. In species subject to Allee effects, it is conceivable that a relatively small fraction of the biomass protected by the reserve could be responsible for the majority of reproduction (see Chapter 6, Figure 6-2).
3. Where fishing reduces the abundance of one sex: This is a particular concern for sequential hermaphrodites, like many grouper species, that change sex from female to male during their lifetime (see Box 2-1). Fishing skews sex ratios when it selectively removes larger fish that are predominantly male and thereby jeopardizes reproduction (Coleman et al., 1996, 1999), although there may be some compensation if sex change occurs at smaller sizes when males are
Page 94
scarce (Roberts and Hawkins, unpublished data). The same pattern is found in many grouper species throughout the world (Shapiro, 1978; Nagelkerken, 1981; Bannerot, 1984; Beets and Friedlander, 1992, 1999; Buxton, 1993; McGovern et al., 1998). Potentially, fishery reserves could enhance recruitment by increasing male survival and sustaining a higher fraction of mature fish to breed.
Throughout the warm-temperate and tropical regions, many groupers, representing some of the more valuable food fish of these regions, undergo seasonal migrations to aggregate on traditional spawning sites. Fishers rely on this aggregation behavior to increase their catch rates by targeting fish during these vulnerable times (Johannes, 1978; Poizat and Baran, 1997). For example, the Nassau grouper (Epinephelus striatus) once had spawning aggregations that numbered in the tens of thousands of individuals but now have been drastically reduced (Colin, 1992; Sadovy, 1993; Sadovy and Eklund, 2000). The same problem afflicts dozens of other species, and one proposed solution is to place spawning aggregation sites in fully-protected reserves (Domeier and Colin, 1997; Johannes, 1998; Roberts, 1998b; Coleman et al., 1999).
Although protecting spawning sites would seem to be a sensible solution, Horwood et al. (1998) suggest that reserves in spawning areas could redirect fishing effort from spawning to nursery habitats and thus from mature to immature fish. Redirection of effort could then lead to more intensive fishing elsewhere, with possible habitat damage and increased bycatch of non-target species. Hence, redistribution of fishing effort is an important consideration in designing MPA networks. A combination of conventional management tools and reserves in spawning and nursery areas may be needed for some species.
Reducing Risk of Population Collapse
The concept of using reserves as buffers against overfishing has been explored to an extent using simulation models of fluctuating populations when catch rates are imperfectly controlled (e.g., Guénette and Pitcher, 1999; Sladek-Nowlis and Roberts, 1999; Crowder et al., 2000). Lauck et al. (1998), for instance, demonstrated that reserves could minimize risk by adjusting the percentage of closed fishing area relative to the catch rate in open areas. When the target catch rate in the open area greatly exceeded the level that maximized sustainable yields, a very large fraction of the area (greater than 60% in this example) had to be closed to maintain the stock at or above the level of maximum productivity. At lower catch rates, smaller closures achieved similar yields. Under the simplifying assumptions used in this model, the fishery reserve effectively protected a fraction of stock equal to the fraction of area closed.
In actuality, the level of protection conveyed by a reserve will depend critically on movement rates across the boundary (Guénette et al., 1998; Fogarty, 1999; Kramer and Chapman, 1999). When spillover of adult fish is high, more
Page 95
of the stock is exposed to fishing, and protection is lower. This “dispersal imbalance” (i.e., more fish leave the reserve than enter) will intensify if fishing effort concentrates near the reserve boundary (Walters, 2000). Simulation models often indicate that a large fraction (20–50%) of productive fishing areas may have to be designated as reserves to provide the desired level of insurance (Roberts, 2000).
Reserves for Migratory Species
When the mobility of adults is high, as in many pelagic and migratory fish species, reserves have often been discounted as an effective management tool. Whereas many coral reef fish have small territories as adults and may disperse during their planktonic larval stage, numerous fish species migrate hundreds or even thousands of kilometers annually (Harden Jones, 1968). Many high-value fish species, including cod (Gadus morhua) and herring (Clupea harengus) migrate long distances and so would obtain only intermittent protection from reserves as they pass through them. However, reserves on spawning grounds or in nursery areas for such species can offer protection and may be a management option. Even for highly migratory species such as swordfish (Xiphias gladius) or tunas, MPAs that protect nursery areas or vulnerable population bottlenecks may be effective as management tools.
Although it is generally assumed that all individuals of a migratory species migrate, recent evidence suggests that many individual fish cover relatively short distances. For example, most of 11,000 galjoen (Dichitius capensis) tagged in a marine reserve in South Africa remained within a few kilometers of their tagging sites, while about 18% dispersed tens to hundreds of kilometers (Attwood and Bennett, 1994). Hence, reserves may protect the less mobile individuals of these migratory species.
Effects on Habitat
Fishing also affects habitat and non-commercial species. However, most studies of reserve performance evaluate only a narrow range of taxa, focusing on fish assemblages. Studies of East African coral reefs demonstrate that fishing has broader impacts on the ecosystem, including changes in biogenic habitat. In this case, the fishery removed keystone predators of sea urchins, leading to urchin population explosions. As urchin grazing intensified, there was increased bioerosion of coral with reductions in coral cover (McClanahan and Shafir, 1990). In addition, sea urchins out-competed herbivorous fish for algae, leading to declines in the fishery for these herbivorous species (McClanahan et al., 1994).
In Jamaica, the ultimate cause of reef degradation following a mass mortality of sea urchins and two hurricanes was attributed to overfishing of herbivorous fish. After the removal of herbivorous fish, sea urchins were the primary
Page 96
players in controlling algal growth on the reefs. Following the die-off of the urchin population and damage to the reefs from two hurricanes, algal growth smothered the remaining coral (Hughes, 1994). A similar occurrence in St. Lucia confirmed that overfishing led to algal overgrowth on the reefs. In St. Lucia, shifts from coral to algal domination occurred only in fished areas, while in reserves, higher densities of grazing parrotfish controlled algal growth and prevented coral losses (Hughes, 1994). At a broader scale, coral cover in the Caribbean appears to be increasing only where there are well-managed marine parks and reserves (Ogden, 1997).
The above examples illustrate how reserves can prevent or reverse indirect fishing effects on habitat. Of greater concern in some regions are the direct and indirect effects of mobile fishing gears such as trawls and dredges (Safina, 1998a; Watling and Norse, 1998). This gear can destroy delicate biogenic habitats that may have taken centuries to develop (Dayton et al., 1995; Koslow, 1997). Most of the Oculina coral reefs of southeastern Florida have been reduced to rubble by trawling (Scanlon, 1998; Koenig et al., 2000). To avoid a similar fate, Norway established two reserves to protect deep-water Lophelia coral beds that recently became vulnerable after the introduction of “rock-hopper” trawls for fishing on rough seabeds. Also, there have been proposals to establish reserves on the Scotian Shelf off Nova Scotia to protect stands of deep-ocean soft corals that are vulnerable to damage from trawling gear (Kenchington et al., in press; Willison et al., in press).
Marine reserves clearly offer a reliable means to protect habitat, especially where fishing gear has been shown to destroy fragile, slow-growing, biogenic habitat such as corals. Often, there are no clear alternatives for protecting spawning sites and nursery grounds (Minns et al., 1996), although gear restrictions might be effective in some situations.
Research in marine reserves is now a fast-growing field of endeavor. Although they rarely appeared in the literature previously, the terms “marine reserves” and “marine protected areas” have increased dramatically in frequency in journal articles since 1993 (Conover et al., 2000). Studies evaluating reserve effects can be found in Roberts and Polunin (1991, 1993a); Dugan and Davis (1993); Rowley (1994); Bohnsack (1996); Allison et al. (1998); and Guénette et al. (1998). However, in the United States, there are very few closed areas that can be studied. The recent executive order from President Clinton ( Appendix E), directing the Departments of Commerce and Interior to establish a national system of marine protected areas, could change this significantly.


























