
Page 20
3
Linkages Between Climate, Ecosystems, and Infectious Disease
This chapter explores the basic processes underlying the linkages between climate, ecosystems, and infectious disease. In the first two sections, background concepts related to climate and to infectious disease dynamics are reviewed to provide a foundation for those who are not already familiar with these topics. The third section provides an overview of the different ways that climate can influence the emergence and transmission of infectious disease agents, and the fourth section reviews other factors that must be considered as part of a comprehensive understanding of disease dynamics.
WEATHER AND CLIMATE: BACKGROUND CONCEPTS
Climate Variability
The distinction between weather and climate is not always appreciated but is important to understand in the context of the topics discussed in this report. Weather refers to the day-to-day state of the atmosphere, characterized by meteorological factors such as temperature, humidity, precipitation, and winds. In contrast, climate refers to average meteorological conditions over a specified time period (usually at least a month), which may include information about the frequency and intensity of extreme events and other statistical characteristics of the weather. Climate varies naturally over a wide range of spatial and temporal scales.
Spatial climate variability includes well-known latitudinal and altitudinal temperature gradients. For example, under typical conditions in mountainous
Page 21
terrain, the average surface air or soil temperature decreases by about 6.5°C for every 1,000-meter increase in elevation, and along an equator-to-pole gradient a distance of 1,000 kilometers corresponds to an average surface temperature change of about 5°C. Superimposed on these large-scale gradients are more complex regional patterns of temperature, precipitation, storm frequency, and so forth, and on very fine geographic scales, microtopographic changes can generate large differences in surface temperature and soil moisture.
Temporal climate variations are most obviously recognized in “normal” diurnal and seasonal variations. The amplitude of the diurnal temperature cycle at most locations is typically in the range of 5 to 15°C. The amplitude of seasonal climate variability is generally larger than that of the diurnal cycle at high latitudes and smaller at low latitudes. Normal annual cycles are modulated by interannual variations in average seasonal conditions. Interannual variations in the mean annual surface air temperature at specific locations are typically on the order of a degree Celsius. Interannual variations in precipitation are more substantial, and can be comparable to the average annual value. Years of research on these types of seasonal-to-interannual variations have uncovered a set of commonly recurring pressure and wind patterns that are termed “modes” of climate variability. The best known of these modes is the El Niño/Southern Oscillation (ENSO) cycle. The irregular cycling back and forth between warm (El Niño) and cold (La Niña) phases of the ENSO cycle in the equatorial Pacific results from a complex interplay between the strength of surface winds that blow westward along the equator and subsurface currents and temperatures. El Niño events (like the very strong one that prevailed in 1997-1998) are marked by higher than average pressure over the western Pacific and lower than average pressure over the eastern Pacific.
ENSO, however, is not the only common mode of climate variability. The Pacific North American Oscillation and North Atlantic Oscillation (thought by some researchers to be part of a larger Arctic Oscillation) are also well researched and are routinely discussed in the climate research literature, although these modes are not yet as familiar to non-specialists. Much of current climate research has turned to the examination of climate variability modes with periods of 10 or more years. North America appears to have a particularly large decadal signal.
On shorter timescales the picture is less clear, but researchers have found “intraseasonal” modes of climate variability with periods of roughly one to two months. These intraseasonal modes are primarily identified by movements of large-scale rainfall patterns in the tropical Indian and Pacific oceans; they appear to interact with ENSO, but their relationships are not well understood. There are some studies suggesting that the tropical intraseasonal variability may have an influence on mid-latitude monthly and seasonal climate.
Finally, climate also varies on very long timescales, which cannot be observed directly but are inferred through a variety of “proxy” records in ice cores,
Page 22
Box 3-1Global Impacts of ENSOThe El Niño/Southern Oscillation (ENSO) cycle is one of the Earth's dominant modes of climate variability The ENSO cycle affects climate around the globe, but only a few regions have consistent documented impacts ( Figure 3-1) During warm episodes in the tropical Pacific Ocean (El Niño events), abnormally heavy rainfall over the warm Pacific waters extends into coastal Ecuador and northern Peru, while drought conditions often prevail in parts of Australia, Malaysia, Indonesia, Micronesia, Africa, northeast Brazil, Central America, and tropical Africa The impacts of El Niño over the United States vary from one event to the next, and they tend to be restricted to winter and early spring On average, Alaska, the Pacific Northwest, and the Northern Plains tend to be mild and dry, while the South tends to be wet Cold years in the tropical Pacific (La Niña events) tend to be marked by the opposite kinds of climate phenomena than in the above region Predictions of these ENSO-associated regional anomalies are generally given in probabilistic terms, because the likelihood of seeing any projected anomaly varies a great deal from region to region and also varies with the strength and specific configuration of the equatorial Pacific sea surface temperature anomalies during the season of interest The term “teleconnections” is often used to describe the statistical relationship between the ENSO cycle and rainfall or temperature anomalies observed in a particular geographical location |
tree rings, and numerous other means. On century-to-millennial scales, climate changes such as the “little ice age” occur; and over the past approximately million years the global climate record is characterized by larger glacial-interglacial transitions, with multiple periodicities of roughly 20,000, 40,000, and 100,000 years, associated statistically with the effects of Earth-Sun orbital variations. The amplitudes of these transitions are on the order of 5 to 10°C and are accompanied by large extensions and retreats of polar and glacial ice.
Climate Change
Since the beginning of the Industrial Revolution, emissions of greenhouse gases such as CO2, CH4, and N2O have been rising due to the growing global population, increasing per capita energy consumption (primarily through the burning of fossil fuels), and land uses such as deforestation and agriculture. These greenhouse gases increase the natural atmospheric trapping of infrared (heat) radiation, a process known as radiative forcing, which results in an enhanced greenhouse effect.
Observational evidence indicates that Earth's global mean temperature has been rising over the last century, particularly during the past 20 years (NRC, 2000). Most climate models project that global temperatures will continue to increase throughout the twenty-first century, with warming of several degrees
Page 23
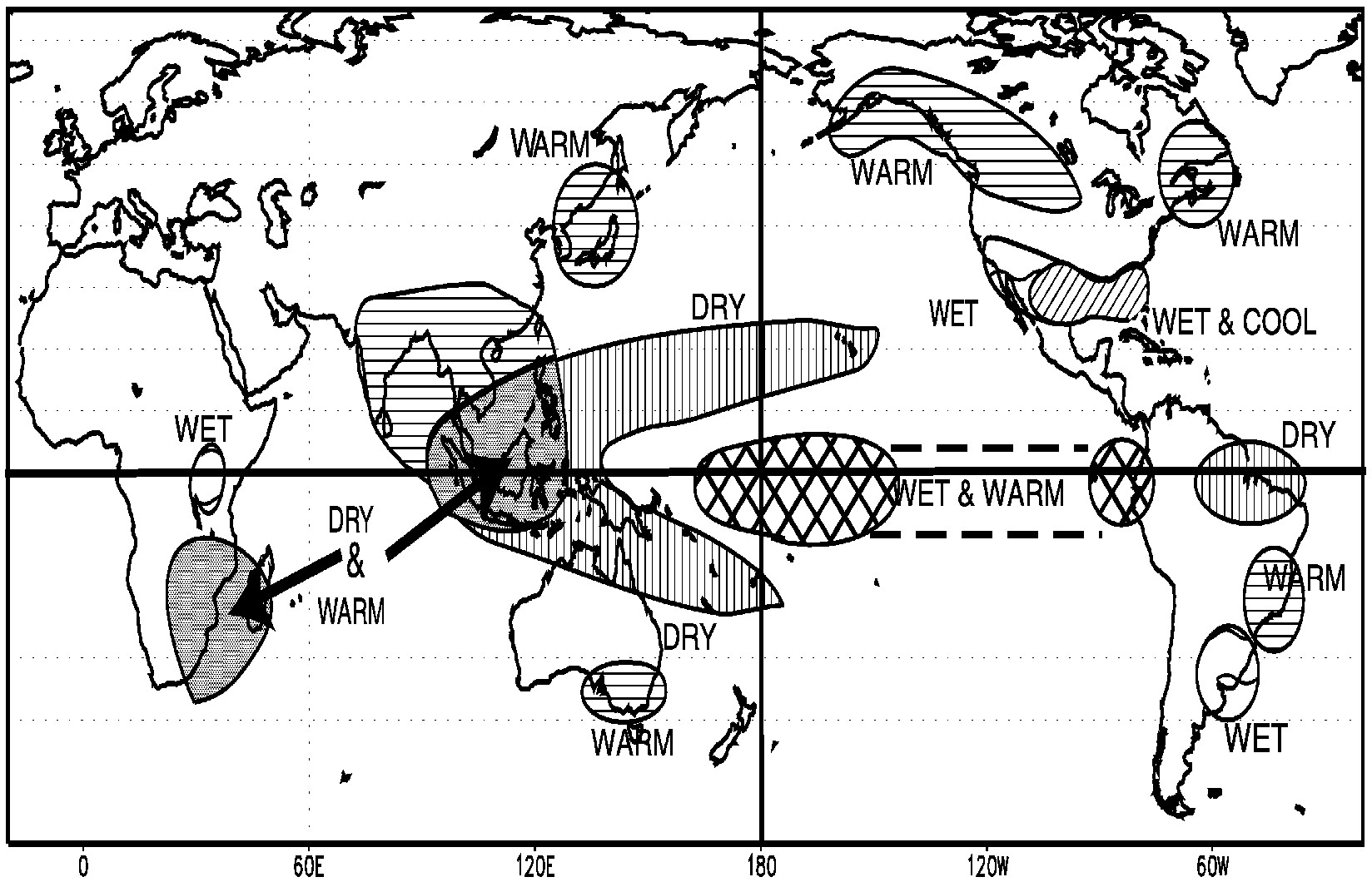
~ enlarge ~
FIGURE 3-1 Illustration of the global impacts of El Niño. Based on Ropelewski and Halpert (1987). Available online at http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/impacts/warm.gif .
Page 24
Celsius by 2100. Perhaps more important than mean global temperature changes, however, is understanding how climate change will affect the “spatial geography” of climate and expressions of other modes of climate variability. The actual effects of global warming on regional-scale temperature, precipitation, and humidity are expected to vary widely. There has been speculation that the long-term global warming trend could affect the occurrence or intensity of El Niño events, but the observational climate record is too short to determine if such changes are indeed occurring. The reader is referred to IPCC (2001) for a more extensive discussion of issues related to global climate change.
In addition to the impacts of greenhouse gases, there are other anthropogenically driven processes that can affect climate on local and regional scales. Industrial emissions can greatly enhance the natural background concentration of aerosols in the atmosphere, and these particles can exert either a net warming or cooling effect, depending on numerous factors such as the particles' chemical and physical properties, the height of the aerosol layer, and the albedo of the surface beneath. Aerosols can also augment the number and properties of droplets within clouds, which in turn affect temperature and precipitation. Recently, satellite data have revealed that plumes of aerosol pollution can have a wide-spread influence on continental precipitation by reducing cloud particle size (Toon, 2000; Rosenfeld 2000).
Changes in land use and vegetation cover can also affect climate over a wide range of spatial scales. Vegetation can, of course, provide shade for the ground underneath, but it can also affect regional precipitation patterns, since the water vapor “exhaled” by forests is a significant source of clouds and rainfall. A modeling study by Pielke et al. (1999) estimated that loss of the South Florida Everglades over the last century has decreased rainfall in the region by about 10 percent. Likewise, there is concern that deforestation-induced drought may be occurring in the Amazon and other parts of the tropics. Another land-use impact is the “urban heat island” effect, in which cities can be up to 12°C warmer than surrounding areas due to the extra heat absorbed by asphalt and concrete and the relative lack of vegetation.
Weather and Climate Forecasting
The skill of routine weather forecasting has increased dramatically since the middle of the twentieth century, commensurate with advances in computer systems and meteorological observations. Forecasts are based on the fundamental laws of physics, including Newton's laws for fluids combined with conservation of mass and momentum. Numerical weather models start with observations of the current atmosphere and integrate forward in time. These forecasts are often defined as solutions to “initial value” problems. Because the atmosphere is an inherently chaotic system, slightly different initial conditions can produce profoundly different forecasts. Theoretical studies indicate that because of the
Page 25
growth of errors in describing the initial state of the atmosphere, accurate weather forecasts are not possible beyond a period of about two weeks (Lorenz, 1982).
Entirely different modeling approaches are required for seasonal climate forecasting. Early attempts to predict climate variations on monthly to seasonal timescales relied almost exclusively on statistical analysis of the past record. These empirical/statistical climate prediction techniques produced marginally skillful seasonal forecasts, but until recently the source of the forecast skill was not well understood, and there was little basis to evaluate whether one seasonal forecast would be more or less accurate than another.
Not until the 1980s did climate scientists begin to capitalize on the premise that the physical basis for seasonal climate modeling does not rest solely in the atmosphere. Seasonal climate variability is now understood to be a manifestation of complex interactions between the atmosphere and underlying surface, primarily the world oceans. The coupling between the oceans and the atmosphere at seasonal timescales was first discovered through study of the El Niño Southern Oscillation.
The current approach to numerical computer modeling of seasonal climate (e.g., Mason et al., 1999) is through the solution of what are known as boundary value problems, where the mean state of the atmosphere is coupled to the mean state of the lower boundary, especially the ocean. Atmospheric models are run with a number of initial conditions for the same boundary conditions. The resulting set of forecasts is called an “ensemble,” and seasonal climate predictions are often given by the mean of ensemble forecasts. The confidence in these forecasts is sometimes gauged by the spread among the ensemble members. Given the nature of the seasonal prediction problem and the tools at hand, all such predictions are inherently probabilistic. Seasonal prediction also often incorporates empirical and statistical tools in addition to numerical model output (Barston et al., 1999; Goddard et al., in press). Typically predictions are expressed in terms of terciles (i.e., the probability that predicted seasonal rainfall or temperature will be in the upper, middle, or lower third compared to the historical record [ Figure 3-2]).
Most modeling and statistical efforts have focused on the equatorial Pacific Ocean basin, but there is good evidence that the Atlantic and Indian oceans also exert an influence on the atmosphere, albeit more locally constrained to the areas surrounding those basins. The ocean dynamics and coupling between the oceans and atmosphere in the Atlantic and Indian oceans are not as well understood, but there are research activities under way to improve this understanding. There is an expectation that improved seasonal forecasts will result once the Atlantic and Indian oceans' variability is better understood and modeled.
While ENSO research has shown the profound influence of the ocean surface on seasonal climate, all atmosphere/surface boundaries may be important, including those over land. Land/atmosphere interactions may be particularly significant in influencing the local climate. There have been major research
Page 26
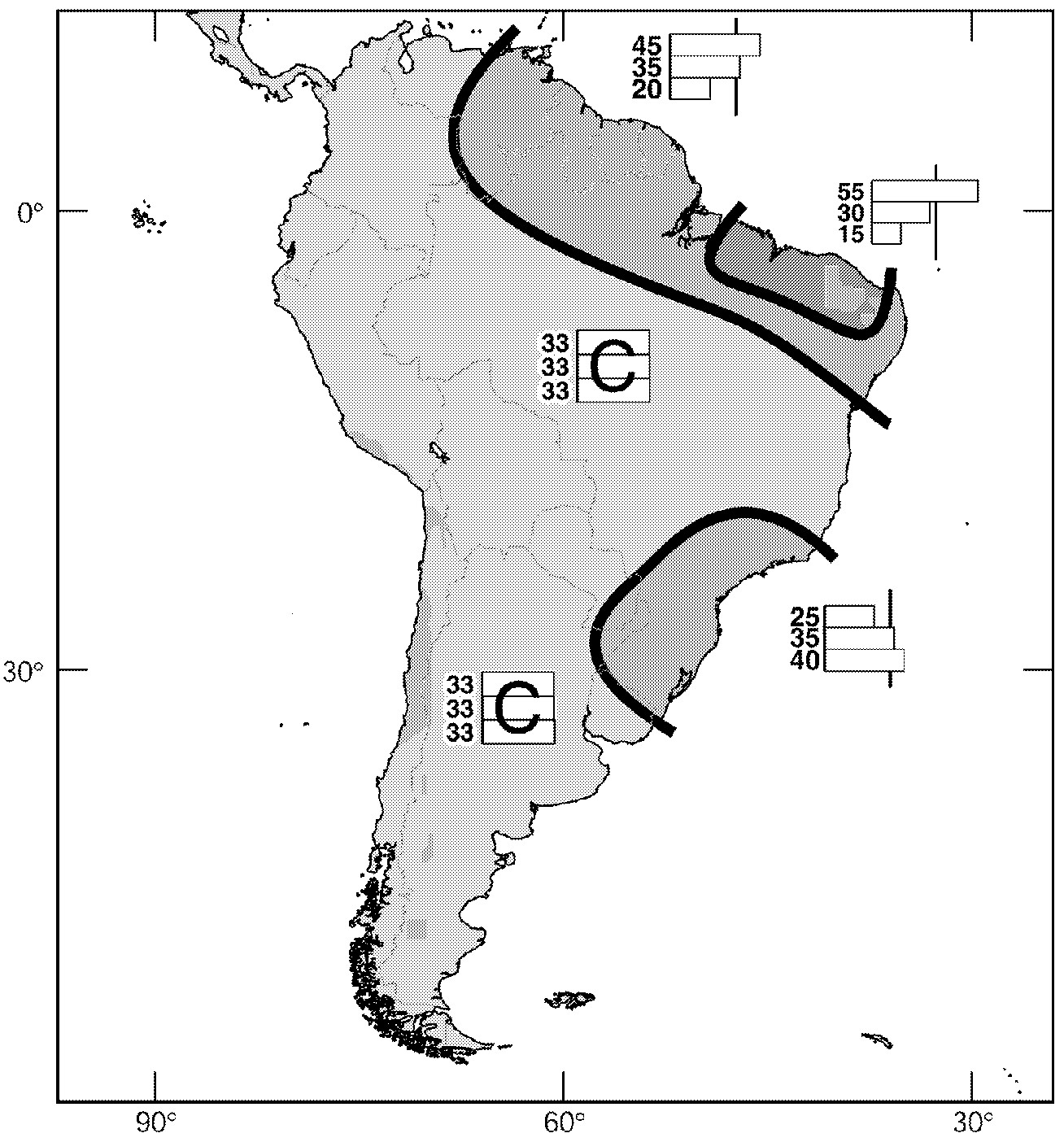
~ enlarge ~
FIGURE 3-2 Example of a seasonal forecast. A forecast for October-December 2000 (produced in September 2000) showing the percentage likelihood that the seasonal precipitation in particular regions will be above normal (top box), normal (middle box), or below normal (bottom box), compared to the mean for the period 1961 to 1990. “C” indicates that there is no basis for favoring any particular category. Source: International Research Institute for Climate Prediction, available online at http://iri.ldeo.columbia.edu/climate/forecast/net_asmt.
efforts in the past several years to monitor, model, and predict land/atmosphere interactions. The successful incorporation of coupled land surface models into seasonal climate forecast models is expected to enhance prediction skill.
Page 27
As described previously, coupling of the worlds' oceans with the global atmosphere provides a basis for climate prediction on seasonal timescales. For shorter time periods though, the influence of the ocean surface is overwhelmed by the “noise” introduced by weather. Thus, there is the conundrum that the tools for day-to-day weather prediction are constrained to less than two weeks, while the tools for climate prediction generally only work well on seasonal or longer timescales. Coupled ocean/atmosphere/land models may improve the ability of climate models to make predictions on shorter than seasonal timescales, and more sophisticated statistical techniques may improve the use of atmospheric weather models beyond 10 to 14 days. Decreasing this relative minimum in predictive capability will be one of the great scientific challenges for climate prediction research in the coming years.
Climate Change Projections
General circulation models (GCMs) represent the large-scale circulations and interactions of the atmosphere in three-dimensional form, and they are the primary tools used to make projections about how the Earth's climate will change in response to the anthropogenic addition of greenhouse gases and aerosols. Recently, scientists have begun to couple GCMs with similar models of the oceans and the biosphere in an effort to better understand the interactions within the Earth-atmosphere system and the subsequent impacts on climate.
Although GCMs are growing increasingly sophisticated, there are still several important sources of uncertainty to consider when using these models to assess possible future climatic changes. While all GCMs project increasing temperatures on a global scale, local- and regional-scale projections are often very different between models. Currently GCMs have a resolution of about 200 to 500 kilometers, yet many critical physical processes, particularly those related to clouds and precipitation, take place on much smaller scales and thus must be crudely parameterized in the models. Various techniques have been developed for “downscaling” climate model simulations, such as regional climate modeling and statistical downscaling (Giorgi and Mearns, 1991; Kattenberg et al., 1996). While these techniques have steadily improved on the simulation of regional climate in the past 10 years, regional-scale projections of climate remain highly uncertain.
The uncertainty in predicting future climate results equally from uncertainty in the modeling of climate itself and uncertainty in predicting emissions of greenhouse gases and aerosols. The latter source of uncertainty depends on projections of population growth, economic growth, and technological changes, among others. Since it is impossible to make quantitative predictions about all of these factors, a common approach is to develop a set of scenarios representing a plausible range of future developments.
Page 28
Box 3-2Terminology: Forecast/Projection/PredictionThere can be subtle but important distinctions in the standard use of common scientific terms among different disciplines, for example, “prediction,” “forecast,” and “projection” The terms forecast and prediction, which both refer to a statement about future events, are often considered to be synonymous Meteorologists, however, tend to make the distinction that a prediction is the result of a single numerical model, while a forecast comes from a synthesis of a number of predictions Future estimates of long-term climate change are usually discussed in terms of “projections,” which are generally considered to be less certain than predictions or forecasts Projections are based on scenarios of possible future changes (in population, economic growth, technological development, etc) with no specific probability associated with any of the scenarios |
INFECTIOUS DISEASE DYNAMICS: BACKGROUND CONCEPTS
Epidemiological Terminology
An infectious, or communicable, disease is an illness caused by a specific infectious agent that is transmitted from an infected person, animal, or reservoir to a susceptible host. Numerous terms are used to describe the occurrence of infectious diseases in a defined population at a particular point in time. For instance, disease “prevalence” refers to the proportion of people with disease in a particular population, while disease “incidence” describes the number of new cases appearing in a population during a particular interval of time.
A disease “epidemic,” often popularly described as an “outbreak,” refers to an excess of cases beyond that which normally occurs in a particular region and at a particular time of year. One or two locally acquired cases of a disease that had not recently been observed in a region may be considered an outbreak. In contrast, a large number of cases in a particular area may not be considered an epidemic unless there were more than had occurred historically during the same time of year. Thus, the number of cases that constitute an epidemic or outbreak will vary with each disease, location, and season. A further complication is the fact that numerous methods are used to estimate disease incidence, not all of which are highly accurate. For instance, relying on the number of hospital visits or clinically diagnosed cases could significantly undercount the incidence of diseases that are largely asymptomatic or that have symptoms shared with other diseases.
As distinct from epidemic, the term “endemic” refers to the constant presence of a disease in a region at a roughly steady incidence during a defined period. In some cases, a disease may be “hyperendemic,” that is, constantly
Page 29
present at a high level of incidence. Endemic diseases can also occur as periodic (often seasonal) outbreaks with varying numbers of cases depending on the intensity of transmission in a particular area, the geographic range of transmission, and the length of the transmission season.
“Emerging” diseases may be caused by a variety of pathogens that are considered to be undergoing change in particular ways. Emerging diseases can be characterized by at least one of the following criteria: (1) diseases caused by a new, previously unknown agent or syndrome; (2) disease symptoms that are more severe and/or more difficult to treat; (3) increased disease incidence in a region; and (4) widening global distribution.
Disease emergence can be due to a variety of causes, including increasing global traffic of goods and people, changes in human behavior and demographics, or a breakdown in public health measures. Other frequently identified factors in disease emergence (especially in outbreaks of previously unrecognized diseases) are climatic or ecological changes that place people in contact with a natural reservoir or host of an infection, by either increasing proximity or creating conditions that favor an increased population of the microbe or its natural host (NRC, 1992; Wilson et al., 1994; Morse, 1995).
Box 3-3Examples of Emerging Diseases(Note that the categories used are not mutually exclusive.) Newly recognized agent or syndrome
More pathogenic, more severe disease, decreased ability to treat
Increased local incidence
Greater distribution or range
|
Page 30
In some cases the emergence of new infectious disease organisms reflects the genetic adaptation of viruses and bacteria to changes in the external environment (e.g., ecological conditions, climate) or the internal environment of the host (e.g., immunity acquired in response to a previous exposure to the disease organism). The illness caused by these new disease strains may have increased severity, such as hemolytic uremic syndrome caused by new strains of toxigenic Escherichia coli, and toxic shock syndrome caused by new strains of group A streptococci. This enhanced virulence is probably the result of genetic events and evolutionary selection (Krause, 2001).
Modes of Disease Transmission
Classifying the different modes of transmission of infectious microbes permits examination of common attributes that may be used to evaluate how climate influences the distribution of disease incidence. First, most infectious diseases can be classified as either anthroponotic or zoonotic. Anthroponotic diseases are caused by microorganisms that normally exist in a transmission cycle involving only humans. Zoonotic diseases occur when microbes that are normally transmitted among nonhuman hosts are transferred to humans. A few microbes are regularly transmitted between and among humans and animals and may cause disease in both. Each of these categories can be further classified according to whether transmission occurs directly or indirectly.
Directly transmitted anthroponoses involve two components (microbial agent, human host) and are spread among people through contact or close association. For instance, the spread of microorganisms such as rhinovirus or Streptococci bacteria generally occurs via bodily fluids. Transmission of microbes such as tuberculosis or measles, for example, occurs via aerosolized droplets, fecal material, fomites, or other particulates. Indirectly transmitted anthroponoses involve three components (agent, vector, and human). Examples include many widespread diseases caused by microbes that are transmitted via arthropod vectors, such as dengue and malaria, which are transmitted by mosquitos.
Directly transmitted zoonoses involve three components (agent, reservoir, and human) and are spread by aerosolized particles or body fluids. People do not often experience this type of infectious contact with animals; rabies is an example of the relatively few diseases of this category. Indirectly transmitted zoonoses, which involve four components (agent, vector, reservoir, and human) infect humans via water, soil, feces, or arthropod vectors. There are numerous examples of these types of transmission cycles, including aerosol-borne hantaviruses, water-borne cryptosporidiosis, sandfly-transmitted leishmaniasis, flea-vectored plague, tick-associated Lyme disease, and mosquito-borne Rift Valley fever. Finally, there are several infectious diseases that are not communicable but that are acquired from the environment, such as Legionnaires, tetanus, and coccidioidomycosis.
Page 31
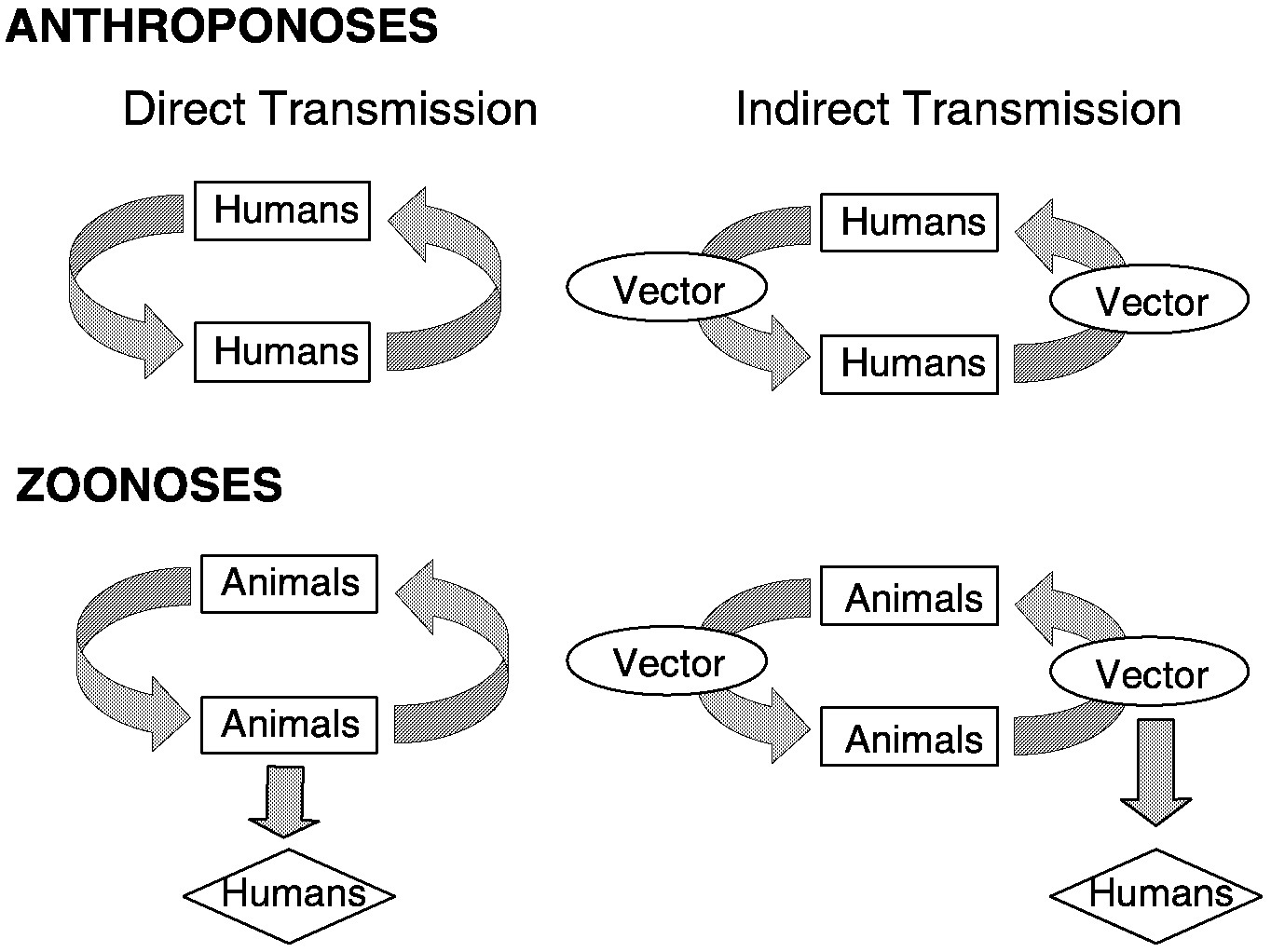
~ enlarge ~
FIGURE 3-3 Illustration of the different types of infectious disease transmission cycles.
These basic categories of transmission are illustrated in Figure 3-3. The impacts of various climate factors on transmission will largely depend on the number of variables that characterize a pathogen's life cycle and the climate sensitivity of each of these variables. Directly transmitted anthroponoses, with the fewest variables and simple links, generally appear to be the least sensitive to climate influences. On the other hand, vector-borne zoonoses, with many environmentally sensitive links, may be highly influenced by climate. At the same time, this larger number of climate-sensitive links makes it more difficult to forecast how climatic changes will alter risk.
The SEIR Framework
At the heart of many studies of infectious disease transmission dynamics is an SEIR modeling framework, which depicts the different states in the progression of a disease through a population: the proportions of individuals susceptible to infection (S); the proportion of people exposed to an infectious agent but not yet infectious (E); the proportion that are actually infectious (I); and those who are removed from the population of interest (R) either because they recover from
Page 32
the infection and are immune or because they die. A typical SEIR model is illustrated in Figure 3-4.
Usually this type of model consists of a series of differential equations that describe the rate of change from one state to another as a function of the relative proportions in each state and of numerous other disease- and population-specific parameters that are determined empirically from laboratory and field observations. For example, the change in the proportion of individuals in the susceptible state is determined by the initial number of susceptibles, by birth and death rates, and by the rate of contact between susceptible and infected individuals multiplied by a factor that reflects how likely this contact is to result in disease transmission.
The SEIR framework reflects the fact that disease dynamics are affected by many factors unique to a particular population, including the population size and density, demographics, connectivity patterns, and immunity levels. Infection of an individual may result in death, chronic infection, or recovery with immunity. At a population level, in some disease systems the gradual loss of widespread immunity due to the birth of susceptibles results in cyclical patterns of disease as
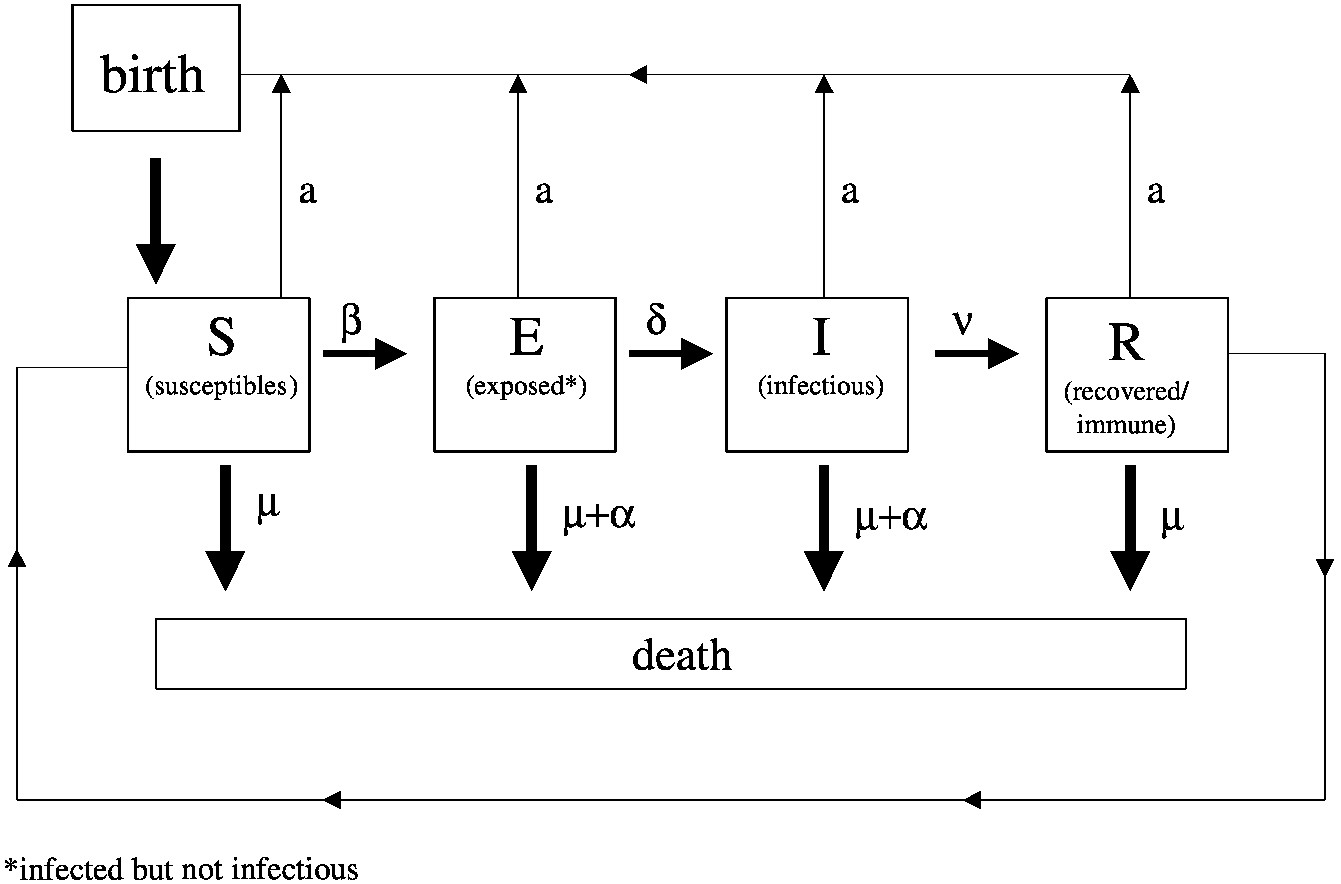
~ enlarge ~
FIGURE 3-4 Diagram of the SEIR framework used in modeling the transmission of disease agents. Definition of parameters: a = host per capital reproductive rate; µ = host per capita death rate; β = transmission coefficient; α = host per capita death rate from being infected; 1/γ = average duration of immunity; 1/δ = average duration of latent period; 1/v = average duration of infection.
Page 33
the population oscillates between resistance and receptiveness to new introductions of a pathogen. For a detailed discussion of these concepts and examples of how the SEIR framework is used in infectious disease modeling, see Mollison (1995) and Anderson and May (1992).
As discussed in Chapter 5, the SEIR modeling approach can lend itself to exploring the potential effects of climate and ecological changes on disease dynamics. In the simplest models of infectious diseases, the rates of change from one state to another in the SEIR model are assumed to be constant, but under changing environmental conditions, each rate is itself a variable. For example, the transmission rate from infected to uninfected hosts can be considered as a variable that is influenced by such factors as changing patterns of demographics and spatial aggregation in hosts and changing reproductive rates and genetics of pathogens. The effects of environmental change can be explicitly incorporated if the SEIR model is embedded in a larger system of dynamical equations that describe the relationships between the parameters of the SEIR model and climatic influences. These relationships can be derived either empirically or from first-principle mechanistic approaches.
WEATHER/CLIMATE INFLUENCES ON INFECTIOUS DISEASES: AN OVERVIEW
Disease agents and their vectors each have particular environments that are optimal for growth, survival, transport, and dissemination. Factors such as precipitation, temperature, humidity, and ultraviolet radiation intensity are part of that environment. Each of these climatic factors can have markedly different impacts on the epidemiology of various infectious diseases. Examples of typical environmental conditions and their effects on various infectious diseases are given in Table 3-1. Climate can directly impact disease transmission through its
|
Environmental Condition |
Disease Favored |
Evidence |
|
Warm |
Malaria, Dengue |
Primarily tropical distribution, seasonal transmission pattern |
|
Cold |
Influenza |
Seasonal transmission pattern |
|
Dry |
Meningococcal meningitis, Coccidioidomycosis |
Associated with arid conditions, dust storms |
|
Wet |
Cryptosporidiosis, Rift Valley fever |
Associated with flooding |
Page 34
effects on the replication and movement (and perhaps evolution) of disease microbes and vectors. Climate can also operate indirectly through its impacts on ecology or human behavior. These different mechanisms are discussed in further detail below.
Direct effects on microbial replication rate. Infectious microorganisms have a replication rate proportional to the ambient temperature. Human body temperature is sufficient for replication of microbes causing human diseases, but ambient temperatures may drop to well below the critical threshold for replication. Malaria, dengue, and other vector-borne diseases are all caused by temperature sensitive microorganisms that replicate in mosquitos, flies, ticks, or other cold-blooded arthropods. The period of time required for microbe replication in the vector species may vary dramatically with the ambient temperature, and below certain temperature thresholds, replication may cease altogether (Gillet, 1974; Molineaux, 1988). Although less well documented, it appears that some infections of the human mucosal surfaces and skin are also sensitive to regional body temperatures. For instance, some respiratory viruses grow preferentially in the upper airways where the cells are a bit cooler than core body temperatures, and bacteria such as leprosy grow preferentially in the cooler tissues of the extremities (Robard, 1981). Waterborne diseases such as cholera also are temperature-sensitive, with a minimum temperature required for replication in the environment.
Direct effects on microbial movement. In order to be transported over the relatively large distances from one host to another, many microbes must be passively borne through moving air or water. Some pathogenic microbes, such as that causing coccidioidomycosis, are picked up from the soil and carried in dry, dusty winds (Smith et al., 1946; Schneider, 1997). Others, such as cryptosporidiosis, may be washed by heavy rains into reservoirs of drinking water (Alterholt et al., 1998).
Direct effects on movement and replication of vectors and animal hosts. The geographic distribution of many arthropod vectors is limited by minimum and maximum temperatures, humidity, and the availability of breeding sites (which, for mosquitos, is often tied to precipitation levels). Meteorological variables may also affect the timing of the vector's life cycle and thus the rate at which it can transmit disease agents. For instance, as temperatures increase over a certain threshold, blood-feeding arthropods often increase their biting frequency and reproduction rates (Gillet, 1974; Shope, 1991; Bradley, 1993). Similarly, in the case of epizootic diseases, the abundance of the primary host (generally another vertebrate species) may depend on climate variables and affect the incidence of disease in human populations.
Page 35
Effects on evolutionary biology. The rapid growth rate and complex life cycles of many infectious diseases agents facilitate the evolution and emergence of new pathogens through horizontal gene transfers or cross-species transmission of microbes. For example, major epidemic influenza virus strains are known to arise by mixing and genetic reassortment of genes from influenza viruses of humans, pigs, and migratory waterfowl (Webster et al., 1992). In recent years there has been growing interest in understanding how disease agents evolve to enhance their own survival, in particular by developing resistance to antibiotics and other treatment drugs (e.g., Ewald, 1994). Climate and ecological changes could plausibly influence the evolution and adaptation of disease pathogens. Thus far, however, there has been very little research to explore such questions, and thus almost nothing is known about potential impacts.
Indirect effects operating through ecological changes. Weather and climate changes can lead to epidemic disease by altering local ecosystems. One example of this is Rift Valley fever, an epidemic viral illness in eastern Africa. Heavy rainfall with ground saturation and pooling of water in surface depressions (“dambos”) leads to increased hatching of the mosquito vectors of the virus and subsequently increases viral transmission (Linthicum et al., 1988). Another example is onchocerciasis (or river blindness) caused by a parasite common in tropical West Africa and South America, where the breeding of the disease vector, the Simulid fly, is governed by river water flow (WHO, 1985). The ecosystem instabilities brought about by climatic changes can give rise to new interactions among hosts and infectious disease agents, possibly accelerating the problem of emerging infectious diseases.
Indirect effects operating through changes in human activities. Changes in weather often bring about changes in human activities that can influence infectious disease transmission rates. For example, warm ambient temperatures often lead to greater use of central air-conditioning systems, which in turn may harbor and spread the bacillus responsible for legionnaires' disease (Garbe et al., 1985). Another example is the fact that school holidays often bring about a decline in influenza and other respiratory virus illnesses simply by reducing crowding and contacts. In the tropics, drought can lead to an increase in dengue because more people store water in open containers, thus increasing the number of breeding sites for mosquitos (Moore et al., 1978).
Ultimately, these climatic impacts must be placed in the context of all the other factors that influence infectious disease dynamics (see Figure 3-5), including land-use changes, transportation and migration patterns, urban crowding, widespread use of antibiotics, and changes in public health infrastructure. (These factors are discussed in depth in the following section.) The disease impacts of climatic changes can in some cases be dampened through natural immune re-
Page 36
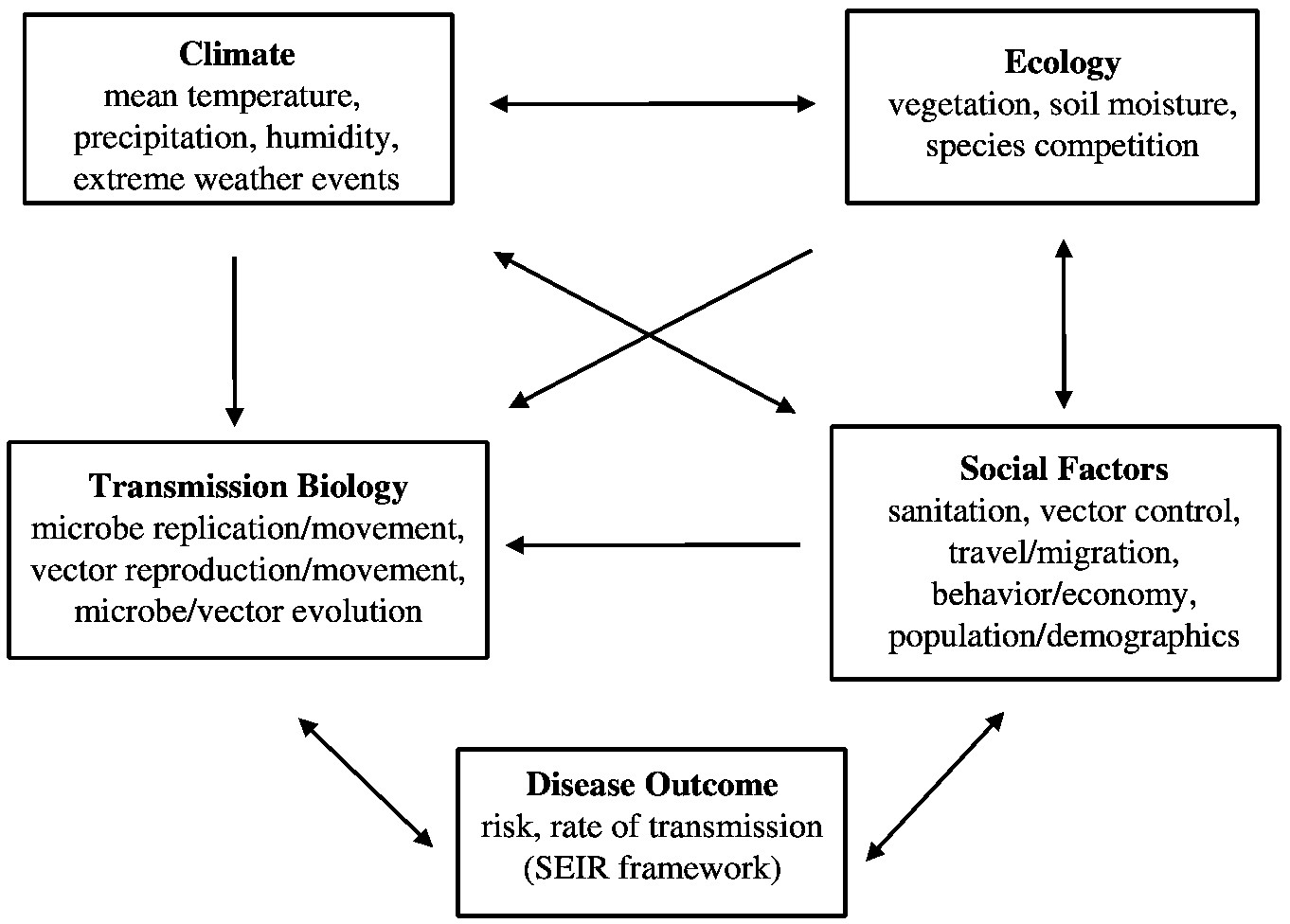
~ enlarge ~
FIGURE 3-5 The “web” of factors that influence transmission of infectious disease agents.
sponses at both the individual level and the population level (“herd immunity”). There are also numerous types of social responses that can modulate the impact of epidemics. Fleeing from urban epidemic epicenters and quarantine of travelers are centuries-old responses. Modern responses include the use of drugs and vaccines to interrupt transmission and/or provide immunity.
Over the long term, a wide array of other largely unforeseeable changes could greatly affect the type and global distribution of infectious diseases. For instance, it is possible that 50 years hence the potential to control diseases such as malaria and dengue could be greatly strengthened by vector eradication programs, improved sanitation/public health systems, or the development of drugs or vaccines. Likewise, it is possible that in the coming decades diseases that are now very rare will become more prevalent or that new strains of infectious agents will emerge. All of these uncertainties greatly complicate any attempts to assess future disease threats.
Extreme Weather Events and Infectious Disease Outbreaks
Extreme weather events such as floods, droughts, and hurricanes occur as isolated incidents but can also be manifestations of longer timescale climate
Page 37
Box 3-4Progression of Climate and Health AssessmentsOften in the study of a complex environmental issue, as scientific understanding of the issue progresses, investigators increasingly come to appreciate the broader context in which the issue is embedded The study of climate and disease linkages has followed this typical progression, and this is reflected in successive assessment reports from the Intergovernmental Panel on Climate Change (IPCC) The IPCC's first assessment report in 1990 contained only a couple of very general statements that “parasitic and viral diseases have the potential for increase and reintroduction in many countries” due to climate change Subsequent IPCC reports issued in 1995 and 1998 included more detailed discussions about climate influences on specific diseases, placed in different regional contexts These analyses considered the difference between direct and indirect effects and emphasized the role of socioeconomic factors in determining disease distribution The third IPCC assessment report (IPCC, 2001) elaborates further on some of these issues and explicitly addresses the methodological difficulties of attributing causation between climate and other coexistent factors This increasing number of caveats and uncertainties may at first appear to indicate a lack of progress in scientific understanding of climate/disease linkages In reality, though, the opposite is true, it reflects a growing understanding of the complexity of the issue and of the need for integrated research approaches. |
variations. For instance, El Niño events typically increase the frequency of tropical storms and hurricanes in the eastern Pacific, and La Niña events favor Atlantic hurricane formation (Saunders et al., 2000). Long-term climate change may increase the risks of extreme events such as floods and droughts occurring in particular regions and could possibly affect the frequency and intensity of mid-latitude storms and hurricanes; however, such projections are highly uncertain at this time.
While the primary risks associated with extreme weather events are injury and death that result directly from the event itself, there is also concern that weather-related disasters can lead to infectious disease outbreaks. The risk factors associated with disasters include (as discussed in Noji, 1997):
-
population displacement and crowding in emergency shelters, which increases the probability of disease communication, particularly when sanitary facilities are insufficient;
-
migration due to drought or other disasters, which exposes refugees to locally endemic disease against which they have no immunity;
-
disruption of healthcare services due to physical damage to roads, hospitals, etc.;
-
malnutrition, which makes people more susceptible to some diseases;
Page 38
-
unavailability of safe drinking water due to contamination of wells and reservoirs from overflow of sewage or failure of water treatment facilities;
-
increase in vectors due to a greater number of breeding sites or disruption of vector control programs;
-
greater exposure to vectors due to lack of shelter.
A common myth associated with natural disasters is that disease epidemics are inevitable. It must be recognized, however, that if disease-causing pathogens are not present in an affected area and are not introduced into the area after a disaster, then disease will not occur even if environmental conditions are ideal for transmission. This may explain why there have been almost no documented disease outbreaks associated with a natural disaster in the United States in over three decades. One possible exception is an outbreak of coccidioidomycosis in California in the wake of a major windstorm (Flynn et al., 1979). It is important to characterize accurately the disease risks associated with different types of extreme weather events, so that limited resources are not needlessly diverted from other, perhaps more critical, facets of a disaster relief effort.
The risks associated with waterborne disease are fairly well understood, and there are numerous examples of enteric disease outbreaks following floods, largely due to disruption in public health and sanitation services (i.e., water treatment). With vector-borne diseases, however, the picture is much less clear. Hurricanes and floods can affect vector populations in numerous and complex ways that can either increase or decrease disease transmission rates. For instance, mosquitos and other vectors can be physically washed away by strong winds and waterflows, which can temporarily suppress disease risk; but as the waters recede and leave behind pools of standing water, the vector populations may rebound dramatically with the greater availability of breeding sites. If the landscape is significantly altered by a disaster, this can have long-term effects on the vector ecology of a region.
There are few studies documenting infectious disease outbreaks associated with disasters, because collection of epidemiological data is not a top priority concern during a crisis situation. Bissell (1983) points out that many epidemiological studies of disasters cover only the first several weeks after an event and thus do not account for the significant lag time in the development of some epidemics (e.g., hepatitis A). Some diseases have a long incubation period and can take several months before reaching their maximum distribution.
While most studies focus on the effects of hurricanes and floods, droughts also have the potential to affect infectious disease risk. Droughts generally develop over a longer timescale than other natural disasters and thus offer more opportunity for interventions. Famine is the most direct risk associated with drought, but the resulting malnutrition can also increase risk of severe illness among infected individuals (Tompkins, 1986). There have been a few instances of cholera outbreaks associated with drought conditions, due to disruption of
Page 39
clean water supplies. Also, drought could increase the risk of meningoccocal meningitis in regions of Africa and Asia where the disease is associated with dry conditions and dust storms (Greenwood et al., 1984).
OTHER FACTORS THAT AFFECT INFECTIOUS DISEASE DYNAMICS
The primary focus of this report is on the relationships between climate and infectious diseases, but in order to understand this relationship it is also necessary to appreciate the many other factors that contribute to the distribution and dynamics of infectious diseases. These include factors such as land-use patterns; social, demographic, and geographical considerations; transportation and migration patterns; and public health interventions. Some of these factors are closely interrelated and are themselves influenced by climate, often in indirect and subtle ways. The complexity of these relationships gives credence to the need to appreciate the “web of causation” in public health.
As described by May (1958), for disease to occur, populations must behave in ways that allow the mutual contact of agents and hosts. Constraints on the geographical distribution of disease agents include landscape characteristics, proximity to breeding areas, and proximity to hosts themselves; while constraints on host behavior may include habitat, cultural influences, tradition, and transportation availability. These concepts, which are explored further in the following sections, are essential to understanding the ecology of disease.
Land Cover / Land Use
Land-use and land-cover changes can exert a profound influence on the distribution of infectious diseases. For instance, malaria vectors tend to be concentrated on land that is exposed to sunlight and that contains pools of water, thus land-cover changes that impact the amount of sunlight in vector-breeding sites can alter the dynamics of malaria transmission (Walsh et al., 1993). The snails that serve as intermediate hosts for schistosomiasis require immobile or slowly moving water. Rice paddies in China, slow-moving water such as the Nile upriver of the Aswan dam, and inland pools of water are all environments in which highly endemic schistosomiasis is frequently found, and modification of these environments has been important in schistosomiasis control efforts (Hairston, 1973).
The proliferation of Lyme disease in the United States has largely been the result of changing land-use patterns. Following European settlement of the northeastern United States, much of the forested land was cleared for agricultural use. As agricultural activity subsequently moved westward to allow for the growth of suburban and new semi-urban settlements, land on the periphery of towns was replaced by second-growth forest. This created an ideal habitat for the white-
Page 40
tailed deer, hosts of the ticks that transmit the disease. In recent years, population pressures and the demand for suburban housing located on the edge of forests have led to more human contact with deer habitat and thus with Lyme disease vectors (Mayer, 2000).
Transportation and Migration
The spread of infectious disease agents is greatly affected by human travel patterns. Migration is one of the means by which diseases spread, either because migrants bring new pathogens with them to their destinations or because the migrants themselves constitute susceptible populations and lack immunity to endemic diseases in their areas of settlement. This is true for both forced migration caused by political/religious oppression and natural disasters and for voluntary migration of people seeking new social or economic opportunities. For instance, migration has been linked to disease transmission patterns of malaria in Africa (Prothero, 1965), and there are examples of outbreaks of malaria among refugees with low levels of acquired malaria immunity in Thailand, Pakistan, Sudan, and Nepal.
Modern transportation has been implicated in the spread of disease pathogens and vectors. Jet transportation in particular makes it possible for an infectious disease to spread rapidly from area to area within a continent or from one continent to another. This has been the case with influenza, where it appears that new strains initially spread from Southeast Asia to other areas of the world. Infected people, who may be asymptomatic, can infect fellow passengers and susceptible people at their destinations. This is particularly the case for diseases with easy transmissibility such as tuberculosis (Kenyon et al., 1996) and other respiratory diseases.
Transportation has also been the putative means by which non-respiratory infectious diseases may be introduced into new areas. For example, antibiotic-resistant gonococci (which cause gonorrhea) initially were found in Asia and then spread to the United States (Gordon et al., 1996; Knapp et al., 1997), clearly following human sexual contact patterns. The movement of HIV/AIDS in Africa has been linked to the sex trade among those engaged in long-distance trucking in East Africa (Pickering et al., 1997).
The development of more highly interconnected areas as regions grow economically is a long-term factor in the transmission of infectious diseases. For instance, the spread of cholera through the United States in the nineteenth century directly reflected the changing connectivity of the urban system. In the earlier cholera epidemics of the century, the spread was based on distance from the portals of introduction of cholera into the country. As individual urban areas became more interconnected through the development of railroads and road systems, the disease spread rapidly from one major metropolitan area to another and to smaller towns within range of these cities (Pyle, 1969).
Page 41
Transportation vehicles themselves can contribute to the spread of vectors to new areas. The concept of “airport malaria” arose from numerous reports of limited malaria outbreaks among populations surrounding airports in temperate non-endemic areas such as the United States, England, and Northern Europe. The clustering of cases around international airports and the subsequent experimental confirmation that anopheline mosquitos could survive a long-distance flight in the wheel wells of jet aircraft demonstrates the potential for air transportation to facilitate the spread of disease vectors (e.g., Giacomini et al., 1997; Guillet et al., 1998). Similarly, one of the Asian vectors of dengue, the mosquito Aedes albopictus, is thought to have been transported to Houston in wet tires being carried aboard a container ship (Moore and Mitchell, 1997). This mosquito may be capable of transmitting dengue in the Americas as it spreads throughout the region.
Social and Demographic Patterns
Social and demographic patterns are major influences on the distribution and spread of infectious disease; and infectious diseases, in turn, exert a major influence on population structure. For example, the “epidemiological transition” is a commonly observed pattern in the development of most societies, wherein most mortality is initially due to infectious causes, but with improved sanitation and nutrition, mortality due to infectious diseases decreases while mortality due to non-infectious causes such as heart disease, stroke, cancers, and accidents increases. Along with this transition comes an increase in life expectancy, and the age structure of the population thus shifts to one where there are proportionately more middle-aged and older individuals.
Population density is another important factor to consider. Contagion is facilitated by population concentration because infected individuals have a higher probability of contact with susceptible members of the population. Population density has been linked with increasing ease of transmission of airborne infections, waterborne diseases, and sexually transmitted infections. Rapid urbanization and population growth in Southeast Asia, concurrent with spread of the urban mosquito vector, are considered to be primary reasons why dengue has become endemic in the region in recent years.
Poverty exerts profound effects on infectious disease patterns, although it is sometimes difficult to separate the effects of population density from other confounding effects of poverty. For instance, immunity can be compromised by malnutrition, HIV infection or other concurrent disease, or addiction to drugs or alcohol (Farmer, 1999). The typical pattern in the developed world is a disproportionate concentration of violence, poverty, and drug addiction in inner-city neighborhoods (Friedman et al., 1996). These neighborhoods have multiple characteristics that are conducive to higher prevalence and easier transmission of infectious diseases, including high rates of homelessness, co-infection and co-
Page 42
morbidity, and poor sanitation and nutrition. Sometimes hospitals and clinics can even provide a locus for spread of an infection to staff and other patients through inadequately sterilized equipment and inadequate isolation measures.
Human contact with previously uninhabited regions such as tropical rainforests has increased due to activities such as deforestation and land clearance schemes such as commercial rubber plantations in Malaysia. This enhances the possibility of human contact with infectious disease cycles in primates and other animals and therefore creates a greater risk of transmission of zoonotic agents to humans. Since the populations involved in these activities are usually very mobile, the potential for rapid transportation of pathogens out of their usual ecological niches is high. This is one reason why there is great concern over the transportation of virulent tropical hemorrhagic fevers such as Ebola, Marburg, and Lassa fevers from their endemic areas to more settled regions in both Africa and other continents. In tropical areas where previously endemic infectious diseases have remained highly localized, greater global interconnectivity is being reflected in the redistribution and spread of these diseases.
Household design and architecture can also influence patterns of vectored disease transmission. For example, the ecology of Chagas disease (American trypanosomiasis) has been transformed as the reduviid or “kissing” bugs that spread this highly prevalent disease have adapted to live in certain types of housing (Bastien, 1998). Second, perpetuation of the cycle of urban dengue fever depends on the proximity of pools of water in tires, cisterns, and on the roofs of houses. Third, some anophelines mosquitos find conducive habitats in urban and rural housing structures, which puts the residents of these houses at greater risk of contracting malaria.
Water development projects have been clearly linked with the intensification of certain disease cycles. Following completion of the Aswan Dam, upriver schistosomiasis increased in prevalence and shifted from a form that primarily affected young children to one that primarily affected young adults in the labor force (Mobarak, 1982). Rift Valley fever and malaria also were linked to the creation of lakes along the Nile and the increased length of the shoreline.
Health Services and Public Health Intervention
The availability of public health services can significantly affect the distribution of disease, since the very purpose of such services is to stem the spread of disease in populations. Vaccination is a specific intervention aimed at preventing the occurrence of disease in individuals and in so doing can reduce the incidence of an infectious disease in populations. Smallpox was the first infectious disease to be eliminated through a program of targeted intervention and vaccination. Polio is now relatively close to being eradicated through vaccination programs, and in many areas childhood diseases such as measles and chickenpox have been well controlled. This has all been due to public health pro-
Page 43
grams mandated by state or national laws and to the efforts of international public health agencies such as the World Health Organization and public service groups such as Rotary International.
The development of antimicrobial agents has also profoundly altered pattems of infectious disease. For instance, the widespread availability of penicillin made it possible to intervene in the progression of streptococcal pharyngitis to rheumatic fever; the development of anti-tubercular agents reduced the prevalence of tuberculosis; sexually transmitted infections such as syphilis, gonorrhea, and chlamydia are highly treatable with antimicrobials; and community-acquired bacterial pneumonias are similarly treatable. However, drug resistance and anti-microbial resistance are increasing problems with some of these diseases.
Public health measures such as improved sanitation, better nutrition, and environmental modification have been of great significance in altering patterns of disease transmission. Indeed, some diseases have been eliminated in specific regions because of these measures. For example, endemic malaria was eliminated in Trinidad through programs for pesticide application, draining swamps, and public education about reducing exposure to mosquitos. Also, the elimination of schistosomiasis in most Caribbean nations, Israel, and elsewhere has been a consequence of eliminating water pools, altering subsurface flora and water flow characteristics, and the application of molluscicides to eliminate the snails that are responsible for transmission of the disease.
Box 3-5Dengue on the U.S./Mexico BorderA striking example of the influence of non-climatic factors on disease distribution is the occurrence of dengue in regions that lie along the U S/Mexico border Between 1980 and 1996, there were 50,333 confirmed cases of dengue recorded in three Mexican states that border the Rio Grande, while there were less than 100 cases on the other side of the river in Texas, and most of these cases were among individuals who had recently visited Mexico (Texas Department of Health, personal communication) The climatic differences between these two regions are insignificant, and surveillance data indicate that mosquito density is actually higher in Brownsville, Texas, than in Reynosa, Mexico (3 8 vs 2 7 Aedes aegypti pupae per person, respectively) Thus, in this case the dramatic discrepancy in disease occurrence can be attributed to differences in living conditions The proportion of homes with air conditioning is higher in Brownsville than Reynosa (42 vs 2 percent) and the proportion of homes with adequate window screens is also higher in the U S community (57 vs 15 percent) Another important factor enhancing transmission of disease agents is the greater proportion of homes in Reynosa with adults and infants present during the day, the biting period of the vector These factors result in significant differences in human exposure to mosquito bites and transmission |
Page 44
Water treatment is a critical measure for protecting public health. Major advances in the control of waterborne diseases began about 1900, when diseases such as cholera and typhoid were dramatically reduced in the United States through the disinfection of drinking water with chlorine (Baker, 1948). By the 1940s and 1950s, sewers and primary treatment of wastes emerged, and secondary treatment came about in the 1970s along with disinfection of wastewater. Despite these advances, waterborne disease continues to occur in many places and affects food supplies through the contamination of irrigation waters and fishing and shellfish harvesting areas. Some of the reasons for this include aging infrastructure of sewage and water treatment facilities in the face of increasing water demands; inability to identify the microorganisms associated with many diseases; newly recognized microorganisms that are more resistant to disinfection (e.g., Legionella, Cryptosporidium, Giardia); contamination of water supplies with sewage overflow or concentrated fecal sources from animal farming operations; and increasing non-point sources of pollution, including septic tanks.
























