
13
Ernst Mayr and the Modern Concept of Species
KEVIN DE QUEIROZ*
Ernst Mayr played a central role in the establishment of the general concept of species as metapopulation lineages, and he is the author of one of the most popular of the numerous alternative definitions of the species category. Reconciliation of incompatible species definitions and the development of a unified species concept require rejecting the interpretation of various contingent properties of metapopulation lineages, including intrinsic reproductive isolation in Mayr’s definition, as necessary properties of species. On the other hand, the general concept of species as metapopulation lineages advocated by Mayr forms the foundation of this reconciliation, which follows from a corollary of that concept also advocated by Mayr: the proposition that the species is a fundamental category of biological organization. Although the general metapopulation lineage species concept and Mayr’s popular species definition are commonly confused under the name “the biological species concept,” they are more or less clearly distinguished in Mayr’s early writings on the subject. Virtually all modern concepts and definitions of the species category, not only those that require intrinsic reproductive isolation, are to be considered biological according to the criterion proposed by Mayr. Definitions of the species category that identify a particular contingent property of metapopulation lineages (including intrin-
|
* |
Department of Vertebrate Zoology, National Museum of Natural History, Smithsonian Institution, Washington, DC 20560-0162. |
sic reproductive isolation) as a necessary property of species reduce the number of metapopulation lineages that are to be recognized taxonomically as species, but they cause conflicts among alternative species definitions and compromise the status of the species as a basic category of biological organization.
Species are one of the fundamental units of comparison in virtually all subfields of biology, from anatomy to behavior, development, ecology, evolution, genetics, molecular biology, paleontology, physiology, and systematics. In large part, the importance of species in biology derives from their importance in systematics, which is responsible for the taxonomic framework used in all branches of biology. Systematics is one of the oldest scientific disciplines and, from its beginning, one of its central concepts has been the concept of species. Systematics can be characterized generally as the branch of science devoted to the study of the different kinds of organisms (biological diversity, in contemporary terms), and the term “species” is Latin for “kind.” Moreover, systematics, for the last 250 years, has been strongly influenced by the familiar hierarchy of taxonomic categories originating from the work of Carolus Linnaeus (1753, 1758), of which the species is the lowest, and in some sense the most fundamental, of the principal categories (de Queiroz, 1997). According to one major dictionary, it is “the basic category of biological classification” (Flexner and Hauck, 1993).
The central role of species in systematics is reinforced by the relationship of systematics to evolutionary biology. Modern systematics continues to become thoroughly integrated with evolutionary biology, and evolutionary biology has, from its inception, granted a central role to species. This situation should be evident from the fact that the most important book in the history of this field, the one that more or less initiated the field itself, is titled On the Origin of Species (Darwin, 1859). The central role of species has continued into the more recent history of the discipline, including the period of the Modern Evolutionary Synthesis (Huxley, 1942; Mayr and Provine, 1980), which laid the foundation for much current research in systematics and evolutionary biology. Evidence for the central role of species is provided by the titles of two of the most important publications from this period, both of which highlight species through reference to Darwin’s title: Dobzhansky’s Genetics and the Origin of Species (1937) and Mayr’s Systematics and the Origin of Species (1942).
In the case of Mayr’s (1942) book, the importance of species is also attested to by the fact that one of the most important and enduring influences of this book, along with subsequent repetitions and elaborations (Mayr, 1963, 1970, 1982), concerns its discussion of species concepts, including a proposed definition of the species category that became a text-
book standard. Most contemporary biologists are familiar with the idea that “species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups” (Mayr, 1942), now commonly known as the “biological species definition” or the “biological species concept.” However, despite the important influence that Mayr’s writings concerning species had on systematics in particular, and on evolutionary biology in general, and despite the wide adoption of his proposed species definition in textbooks, these contributions did not solve the long-standing problem concerning the nature of species. In fact, Mayr’s proposed definition seems to have led to the emergence of new sources of disagreement.
In this paper, I will argue that the reconciliation of alternative and incompatible definitions of the species category is a natural outgrowth of the general concept of species for which Ernst Mayr was one of the primary developers and advocates. On the other hand, this proposed reconciliation is at odds with most contemporary species definitions, including the popular definition proposed by Mayr himself, at least as commonly interpreted. The incompatibility results from treating certain properties acquired by diverging population lineages as necessary properties of species, including potential interbreeding and its counterpart, intrinsic reproductive isolation, in the case of Mayr’s definition. Reinterpreting these properties as neither necessary nor sufficient for the definition of the species category eliminates the incompatibilities among alternative concepts of species, resulting in a unified species concept that brings taxonomic practice in line with common claims about the theoretical significance of species, and that is highly consistent with the general concept of species for which Ernst Mayr was arguably the most articulate and prolific advocate.
THE SPECIES PROBLEM
Despite the wide acceptance of Mayr’s proposed species definition (and perhaps partly because of it), this definition stimulated critiques as well as the proposal of alternatives. An early critique, including an alternative definition, was published by George Gaylord Simpson (1951, 1961), another leader of the Modern Synthesis (Fitch and Ayala, 1994). However, alternative species concepts did not really begin to proliferate until the 1970s, starting with a paper by Sokal and Crovello (1970), which proposed a phenetic species concept. By the late 1990s, literally dozens of alternatives had been proposed. Mayden (1997), for example, identified 24 named species concepts, including the now-familiar biological, phenetic, evolutionary, ecological, and phylogenetic (three versions) concepts and 16 others. The diversity of contemporary species concepts has been
reviewed in several recent publications (Coyne and Orr, 2004; de Queiroz, 1998; Harrison, 1998; Mayden, 1997) and will not be repeated here. For the present discussion, the important thing to recognize is that different contemporary species concepts are based, in part, on different biological properties. For example, the biological species concept emphasizes the property of reproductive isolation (Dobzhansky, 1970; Mayr, 1942), the ecological species concept emphasizes occupation of a distinct niche or adaptive zone (Andersson, 1990; Van Valen, 1976), one version of the phylogenetic species concept emphasizes diagnosability (Cracraft, 1983; Nixon and Wheeler, 1990) and another, monophyly (Donoghue, 1985; Rosen, 1979). For a more extensive list of properties that form the basis of alternative species concepts, see de Queiroz (1998).
As a consequence of these differences, many alternative contemporary species concepts are incompatible in that they lead to the recognition of different species taxa depending on which concept is adopted. In other words, they lead to different species boundaries and different numbers of recognized species. For example, adopting the diagnosable version of the phylogenetic species concept commonly leads to the recognition of many more species taxa than adopting the biological species concept (Cracraft, 1983; Martin, 1996; Zink, 1996). The existence of alternative. and at least partially incompatible, definitions of the species category, hereafter referred to as the “species problem,” creates difficulties given that species are used as basic units of comparison in diverse types of studies. On the one hand, species taxa recognized according to different species concepts often will not be comparable to one another with regard to the biological properties they possess. On the other hand, a study that uses species taxa based on a single species concept may yield very different results from one that uses species taxa based on a different species concept. This is not to deny that particular concepts are preferred by particular groups of biologists. Some such groups argue passionately about the superiority of their preferred concept over the alternatives. However, other groups argue just as passionately in favor of different species concepts. In addition, the species problem seems to be getting worse rather than better, which is to say the number of alternative species concepts has been growing rather than diminishing. Moreover, judging by the increasing numbers of critiques and proposed alternatives, Mayr’s species definition, although still perhaps the most widely adopted, seems to be less popular now than 20–30 years ago.
The existence of diverse species concepts is not altogether unexpected, because different concepts are based on properties that are of greatest interest to different subgroups of biologists (de Queiroz, 1998). For example, biologists who study hybrid zones tend to emphasize reproductive barriers, whereas systematists tend to emphasize diagnosability and monophyly, and ecologists tend to emphasize niche differences. Paleon-
tologists and museum taxonomists tend to emphasize morphological differences, and population geneticists and molecular systematists tend to emphasize genetic ones. Nevertheless, for those biologists who are able to set aside their own personal investments and research interests, all of the concepts seem to have some merit. It is certainly the case that all are based on important biological properties.
THE METAPOPULATION LINEAGE CONCEPT OF SPECIES
The reconciliation of alternative and incompatible species concepts derives from the recognition of a more general concept of species that is shared by all contemporary species concepts and definitions (de Queiroz, 1998, 1999, 2005). This general concept of species originated at least as early as the beginning of the 20th century [Mayr (1955, 1982) cited papers by Jordan (1896, 1905) and Poulton (1903) as early examples], but it became well established during the period of the Modern Evolutionary Synthesis (Huxley, 1942; Mayr and Provine, 1980), through the writings of the great leaders of that movement, including Sewall Wright, Theodosius Dobzhansky, George Gaylord Simpson, and particularly Ernst Mayr. All modern species concepts and definitions conform to this general species concept and can therefore be considered variants of it. This general species concept, not Mayr’s more restricted species definition, is the true biological species concept (see below).
Several influential discussions of the general species concept that became established during the Modern Synthesis emphasized the correspondence of species with metapopulations or gene pools. Species were equated with groups of interconnected populations that form an extended reproductive community and an unevenly distributed but unitary gene pool or field for gene recombination. The equation of species with metapopulations or gene pools is evident in a number of species definitions from the period of the Modern Synthesis, including those proposed by several of the most influential contributors to that movement. Thus, according to Sewall Wright (1940) “[species are] groups within which all subdivisions interbreed sufficiently freely to form intergrading populations wherever they come in contact, but between which there is so little interbreeding that such populations are not found.” Similarly, according to Theodosius Dobzhansky (1950), “The biological species is the largest and most inclusive Mendelian population” (a “Mendelian population is a reproductive community of sexual and crossfertilizing individuals which share in a common gene pool”). And finally, according to Ernst Mayr (1942), “Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups.”
All three of these definitions equate species with metapopulations (sets of connected subpopulations, maximally inclusive populations), and all of them suggest that the limits of species as metapopulations are set in one way or another by the limits of interbreeding (which requires not only mating but also the production of viable and fertile offspring), thus implying sexual reproduction. For cases involving purely asexual reproduction, there are two possibilities regarding species. One possibility is that purely asexual organisms do not form species (Dobzhansky, 1937; Ghiselin, 1997; Hull, 1980). The other possibility is there are processes other than the exchange of genetic material, such as natural selection, that determine the limits of species in purely asexual organisms (Meglitsch, 1954; Templeton, 1989; Van Valen, 1976). Both of these views are consistent with the equation of species with metapopulations. Either asexual organisms do not form metapopulations, and therefore they do not form species, or they do formmetapopulations (as the result of some process or processes other than interbreeding), and therefore they also form species.
The general metapopulation concept of species is also evident in species definitions that describe species as lineages rather than as populations. For example, according to George Gaylord Simpson (1961), “[a] species is a lineage (an ancestral-descendant sequence of populations) evolving separately from others and with its own unitary evolutionary role and tendencies.” Similarly, according to Leigh Van Valen (1976), “A species is a lineage (or a closely related set of lineages) which occupies an adaptive zone minimally different from that of any other lineage in its range and which evolves separately from all lineages outside its range.” The reason definitions that characterize species as lineages can be considered to represent the same general species concept as those that characterize species as (meta)populations is there is a close relationship between populations and lineages at the same level of biological organization (Fig. 13.1). On the one hand, populations can be considered to extend through time, in which case a population is equivalent to a lineage. Alternatively, populations can be considered to exist at an instant in time, in which case a population is equivalent to an instantaneous cross section of a lineage, and a lineage corresponds to a continuous series of populations (the ancestral-descendant sequence of populations in Simpson’s definition). In either case, this type of lineage (a single line of ancestry and descent) is not the same as a monophyletic group (a group of entities sharing an exclusive common ancestry), which is also commonly referred to as a lineage (although the term “clade” is more appropriate).
The important point is that virtually all contemporary definitions of the species category are based on a common general concept of species: the concept of species as (segments of) metapopulation lineages (de Queiroz, 1998, 1999, 2005). Definitions that describe species as popula-
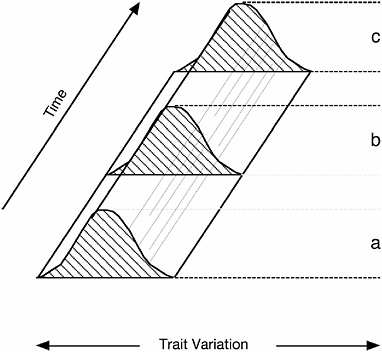
FIGURE 13.1 Populations as lineages (modified from Simpson, 1951). The population or population level lineage at a given instant in time is represented as a frequency distribution in two dimensions [x axis, trait variation; y axis, frequency (not shown)], whereas the time-extended population or population level lineage is represented by the 3D solid formed by extending the 2D frequency distribution through a third dimension (z axis, time). The three shaded distributions (a–c) represent cross sections of the time-extended population lineage at three different times. The population itself can be interpreted either as the 3D (time-extended) object, in which case it is equivalent to the lineage, or as one of the 2D (time-limited) objects, in which case it is equivalent to a cross section of the lineage.
tions simply view species over some relatively short interval of time, usually the present. In contrast, definitions that describe species as lineages tend to view species over longer time intervals. (In this context, it is not surprising that the first of the lineage definitions was proposed by Simpson, a paleontologist.) Because the population vs. lineage terminology simply reflects a different temporal perspective on entities of the same basic kind, I will hereafter use the term “metapopulation lineage” to encompass both views. Virtually all contemporary species definitions (by which I mean those advocated by some contemporary group of biologists) conform to this general metapopulation lineage concept of species (de Queiroz, 1998). They differ with regard to emphasizing (in addition to different temporal perspectives) the theoretical concept itself vs. the em-
pirical evidence and operational procedures that are used to apply it. They also differ with regard to the properties of metapopulation lineages that are considered necessary for those lineages to be regarded as species. Because the metapopulation lineage concept of species is general in the sense that it is shared by all modern concepts and definitions of species, it has previously been referred to as the “general lineage concept of species” (de Queiroz, 1998, 1999, 2005), although it might be more accurately termed the “general metapopulation lineage concept of species.”
THE SPECIES AS A FUNDAMENTAL CATEGORY OF BIOLOGICAL ORGANIZATION
An important corollary of the metapopulation lineage concept of the species is that the species is a fundamental category of biological organization. Although this corollary is now often taken for granted, it is important to recognize that it represents a significant departure from an older view of the species category. Under the older view, the species category was simply a rank in the hierarchy of taxonomic categories. More specifically, the taxa at all levels in the hierarchy were viewed as being of the same basic kind, namely, groups of organisms that shared particular traits (de Queiroz, 1997), but they were assigned to different ranks to indicate differences in relative inclusiveness. Species were included within genera, genera were included within families, and so forth. Thus, species were not viewed as constituting a fundamentally different kind of entity than genera or families; they were just smaller groups separated by smaller degrees of difference. This perspective was held by Darwin (1859), who stated that he viewed “… the term species, as one arbitrarily given for the sake of convenience to a set of individuals closely resembling each other, … it does not essentially differ from the termvariety, which is given to less distinct and more fluctuating forms” and that “the grades of acquired difference [between taxonomic groups are] marked by the terms varieties, species, genera, families, orders, and classes.” In short, Darwin viewed the species category as just another categorical rank, in particular, one applied to groups of organisms that differed more than varieties but less than genera.
In contrast, the view of species that emerged during the Modern Synthesis was that species are fundamentally different from the taxa above and below them in the taxonomic hierarchy. Species differ from genera (for example) not only in inclusiveness but also in kind. Species are metapopulation lineages, whereas genera are groups of species sharing a relatively recent common ancestry. Thus, according to Simpson (1961), “… there are units in nature that have a special evolutionary status not fully shared with taxa either above or below them in the hierar-
chy…. Many of them … recognized before Darwin had been called species, and it was inevitable that the term should be transferred to the evolutionary units.” Similarly, according to Mayr (1969a), “The unique position of species in the hierarchy of taxonomic categories has been pointed out by many authors…. It is the only taxonomic category for which the boundaries between taxa at that level are defined objectively.”
As a fundamental category of biological organization, the species category is roughly analogous to other such categories, including the cell and the organism. In Mayr’s (1982) words, “… the species is as important a unit of biology as is the cell at a lower level of integration.” Species may differ from cells and organisms in terms of the processes responsible for uniting their parts to form larger wholes, and they may exist at a higher level of organization (the population level), but all three categories are fundamentally similar in identifying particular kinds of biological entities that replicate or reproduce in the sense of generating other entities of the same kind. Cells divide to produce new cells, organisms reproduce to generate new organisms, and species speciate to produce new species.
Despite the relatively wide acceptance of the idea that the species represents a fundamental category of biological organization, some of the ways in which biologists continue to treat species taxonomically are inconsistent with that proposition. The practices in question appear to be holdovers from the earlier interpretation of the species category as a rank in the hierarchy of taxonomic categories (i.e., as opposed to its later interpretation as a fundamental category of biological organization). Moreover, those practices also appear to be responsible for the current species problem, that is, for the existence of alternative and at least partially incompatible definitions of the species category.
If the species category is to be truly analogous to the categories cell and organism and of similar general theoretical significance, then the species category must be the most general category at its particular level of biological organization. It cannot be a less general subset of this general category. Consider the analogous category organism. This category includes all living (and formerly living) beings, which are not required to possess any property beyond those used to define the general category, such as being born, or sexually mature, or fully grown, to be considered organisms. Embryos, juveniles, and adults are all considered organisms. Similarly, in the case of cells, once the entities are physically separated, they need not possess any additional property, such as having replicated their DNA or condensed their chromosomes, to be considered cells. Cells at various phases in the cell cycle are all considered cells. The general manner in which the categories cell and organism are conceptualized is what confers on these categories their general theoretical significance (which is greater than that of more restricted categories, such as prophase
TABLE 13.1 Properties, in Addition to Existence as a Separately Evolving Metapopulation Lineage, Commonly Treated as Necessary Properties of Species
|
Property |
Species Concept and/or Definitiona |
|
Potential interbreeding (intrinsic reproductive isolation) |
Biological species concept/definition (isolation species concept) (Mayr, 1942; Dobzhansky, 1970) |
|
Shared specific mate recognition or fertilization system |
Recognition species concept (Paterson, 1985) |
|
Same niche or adaptive zone |
Ecological species concept (Van Valen, 1976) |
|
Monophyly (as inferred from apomorphy or exclusive coalescence of gene trees) |
Monophyly version of the phylogenetic species concept (Rosen, 1979; Donoghue, 1985), genealogical species concept (Baum and Shaw, 1995) |
|
Form a phenetic cluster (quantitative difference) |
Phenetic species concept (Sokal and Crovello, 1970; Michener, 1970; Sneath and Sokal, 1973) |
|
Form a diagnosable group (fixed qualitative difference) |
Diagnosable version of the phylogenetic species concept (Cracraft, 1983; Nixon and Wheeler, 1990), some interpretations of the evolutionary species concept (Grismer, 1999, 2001) |
|
Form a genotypic cluster |
Genotypic cluster species definition (Mallet, 1995) |
|
aAccording to the classification of de Queiroz (1998). |
|
cell and adult organism). In the case of the species, however, most biologists continue to treat this category in a way that is at least partially inconsistent with the general theoretical significance commonly attributed to it. Under most current species definitions, the species is not the most general category at its particular level of organization. The reason is that those definitions commonly require separately evolving metapopulation lineages to possess some additional property before they are considered species.
Table 13.1 lists some of the additional properties that are commonly interpreted as necessary for a separately evolving metapopulation lineage to be considered a species (i.e., as defining properties of the species category). Those properties include intrinsic reproductive isolation, as in Mayr’s well known species definition (Mayr, 1942, 1963, 1970); the occupancy of a distinct niche or adaptive zone, as in the ecological species
definition proposed by Van Valen (1976); monophyly, as in definitions proposed by certain phylogenetic systematists (Donoghue, 1985; Rosen, 1979); and a number of others. Hereafter, I will refer to these properties as contingent properties, because they are properties that a metapopulation lineage may or may not acquire during the course of its existence (the longer it persists, the more likely it is to acquire them).
The problem with the interpretation of these contingent properties of metapopulation lineages as necessary properties of species is that it compromises the generality of the species concept. By requiring that a separately evolving metapopulation lineage be intrinsically reproductively isolated, or ecologically differentiated, or monophyletic, or anything else, before it is considered a species, the concept of species is restricted to only some members of the general category at the level of biological organization in question (i.e., to only some metapopulation lineages). As a consequence, the generality of the species concept is restricted. To use analogies from other levels in the hierarchy of biological organization, current definitions of the species category are analogous to considering only those membrane-bound parcels of cytoplasm that have reached the S phase of the cell cycle to be cells or only those living beings that have reached sexual maturity to be organisms.
The interpretation of properties such as intrinsic reproductive isolation, ecological distinctiveness, monophyly, and so forth, as necessary properties of species seems to represent a holdover from the earlier view of the species as a taxonomic rank used to distinguish among groups that differ only in inclusiveness (i.e., as opposed to representing a distinct category of biological organization). The reason is that, in effect, those properties are being used to decide which metapopulation lineages deserve to be ranked as species. Those metapopulation lineages that are separately evolving but have not yet acquired the stipulated property are not considered to merit the rank of species. Instead, they are commonly ranked as subspecies. Thus, even if the species category is being treated as fundamentally different from the genus, the family, and the other higher taxonomic categories, it is still effectively being treated as a different rank rather than a different kind relative to the lower ones, in particular, relative to the subspecies category. In other cases, metapopulation lineages that have not yet acquired the stipulated property are not granted any formal taxonomic recognition whatsoever (i.e., as opposed to being ranked as subspecies). In such cases, the subspecies rank has been eliminated, and the species has effectively become the only rank assigned to metapopulation lineages. But regardless of whether one considers this situation to represent a holdover from the interpretation of the species category as a rank, it still means the species category is not the most general category at its particular level of organization.
THE CAUSE OF THE SPECIES PROBLEM
In addition to restricting the theoretical significance of the species category, the interpretation of various contingent properties of metapopulation lineages as necessary properties of species is also the cause of the species problem. That is, it is the reason for the existence of incompatible alternative definitions of the species category. Because different authors adopt species definitions that treat different contingent properties of metapopulation lineages as necessary properties of species (e.g., intrinsic reproductive isolation, diagnosability, or exclusive coalescence of alleles), those authors commonly disagree about which metapopulation lineages deserve to be ranked as species (de Queiroz, 1998, 1999, 2005).
The reason emphasis on different contingent properties leads to incompatible species definitions is that those properties arise at different times during the process of separation and divergence among metapopulation lineages (i.e., speciation). Lineage separation and divergence can be conceptualized in terms of a few general evolutionary processes: mutation, natural selection, migration (or the lack thereof), and genetic drift. In contrast, the properties affected by those processes are highly diverse. They may be genetic or phenotypic, qualitative or quantitative, selectively advantageous, disadvantageous, or neutral, and they may involve many different aspects of biology, including genetics, development, morphology, physiology, and behavior.
With regard to the species problem, the important point is that the process of evolutionary divergence leads to the acquisition of a number of different properties by diverging lineages, including those that have been emphasized by different groups of biologists in their definitions of the species category. Thus, as lineages diverge, they (or their component organisms) become distinguishable in terms of quantitative traits. They become diagnosable in terms of fixed character states. Their genitalia, gametes, genomes, and developmental systems become incompatible. Their mate-recognition systems diverge to the point where they no longer recognize one another as potential mates. They evolve distinctive ecologies. And they pass through polyphyletic, paraphyletic, and monophyletic stages in terms of their component genes and organisms. These changes commonly do not occur at the same time, and they are not even necessarily expected to occur in a regular order. The problem is that each alternative species definition adopts a different one of these properties as a defining or necessary property of species. This is the reason that the alternative species definitions, despite their general agreement regarding the conceptualization of species as metapopulation lineages, imply different conclusions concerning which lineages deserve to be recognized as species.
The highly simplified diagram in Fig. 13.2 represents the process of metapopulation lineage divergence. The progressive darkening and light-
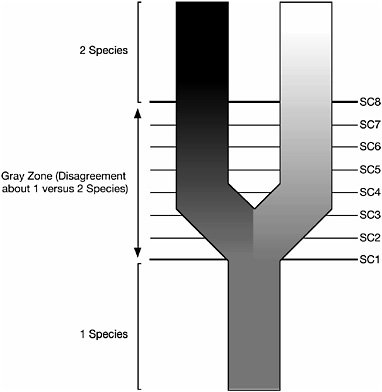
FIGURE 13.2 A highly simplified representation of the process of metapopulation lineage divergence (speciation) illustrating the conflicts caused by adopting different contingent properties of metapopulation lineages as necessary properties of species (modified from de Queiroz, 1998). Progressive darkening and lightening of the daughter lineages represent their progressive divergence through time (bottom to top), and the numbered lines labeled SC (species criterion) 1–8 represent the times at which the daughter lineages acquire different properties relative to one another (e.g., when they become phenetically distinguishable, diagnosable by a fixed character difference, reciprocally monophyletic, reproductively incompatible, ecologically distinct, etc.). Before evolution of the first property (SC1), authors will agree there is a single species, and after evolution of the last property (SC8), they will agree there are two. Between these events, however, there will be disagreement among authors about whether one vs. two species are involved. Those disagreements result from authors adopting different contingent properties (species criteria) as the basis for their species definitions.
ening of the daughter lineages represent their increasing divergence through time, and the numbered lines represent the times at which they acquire different properties relative to one another, for example, when they become phenetically distinguishable, diagnosable, reciprocally monophyletic, reproductively incompatible, ecologically distinct, and so
forth. The entire set of properties forms a broad gray zone where alternative species concepts come into conflict. On either side of the gray zone, there will be unanimous agreement about the number of species: before the acquisition of the first property, everyone will agree there is one species, and after the acquisition of the last property, everyone will agree there are two. In between, however, there will be disagreement. Some people will place the cutoff for treating the diverging lineages as separate species relatively early in the sequence, perhaps where loss or fixation of a character in one of the lineages makes them diagnosable. Others will place the cutoff later, perhaps where the lineages develop an intrinsic reproductive barrier. Still others will place the cutoff later still, perhaps where both lineages form exclusive groups in terms of multiple gene trees. This adoption of different thresholds as criteria for treating diverging lineages as different species is the cause of the species problem. It is the reason for the existence of multiple incompatible definitions of the species category despite widespread agreement about the general nature of species.
A SOLUTION TO THE SPECIES PROBLEM
Both the species problem itself and the discrepancy between the general theoretical significance commonly attributed to species and the treatment of species in taxonomic practice can be solved by making a simple yet fundamental shift in the way species are conceptualized (de Queiroz, 1998, 1999, 2005). This shift is highly consistent with the general concept of species that became established during the Modern Evolutionary Synthesis and for which Ernst Mayr was arguably the most important spokesman. It also represents the more complete acceptance of Mayr’s (1982) proposition that the species is one of the fundamental categories of biological organization. The proposed solution retains the element common to all contemporary concepts of species, and it eliminates the conflicts between those rival concepts without denying the importance of the properties that underlie their obvious differences.
The proposal has two components. First, it retains the element common to all contemporary concepts and definitions of species by adopting the general concept of species as separately evolving metapopulation lineages. Second, it eliminates the conflicts among rival concepts by treating this property, existence as a separately evolving metapopulation lineage, as the only necessary property of species. In other words, all of the other properties that have previously been treated as necessary properties of species, which created incompatibilities among alternative species concepts, are reinterpreted as no longer being defining properties of the species category. Instead, they are interpreted as contingent properties not
only of metapopulation lineages but also of species, properties that species as metapopulation lineages may or may not acquire during the course of their existence. In other words, metapopulation lineages do not have to be phenetically distinguishable, or diagnosable, or monophyletic, or reproductively isolated, or ecologically divergent, to be species. They only have to be evolving separately from other such lineages. Because the interpretation of various secondary properties of lineages as necessary properties of species is the cause of the incompatibilities among alternative species concepts, their reinterpretation as contingent rather than necessary properties also removes the incompatibilities. The result is a single, general, unified concept of species.
The reason the resulting species concept can be considered unified is that it does not deny the importance of any of the properties that have been emphasized in previous definitions of the species category. Under a general and unified concept of species, the various contingent properties, although no longer treated as necessary properties of species, remain important in two ways. First, they continue to serve as important lines of evidence relevant to assessing the separation of metapopulation lineages. Indeed, the properties in question (e.g., phenetic distinguishability, reciprocal monophyly, pre- and postzygotic reproductive isolation, fixed character state differences, etc.) are among the best lines of evidence regarding the separation of metapopulation lineages. Second, the various contingent properties can be used to define subcategories of the general species category, that is, to recognize different classes of species based on the properties possessed by those entities. Just as different subcategories of the general category organism are recognized based on properties possessed by organisms (e.g., sexually mature organisms, fully grown organisms, socially dominant organisms, etc.), similarly, different subcategories of the general category species can be recognized based on properties possessed by species (e.g., diagnosable species, reproductively isolated species, monophyletic species, etc.). Thus, under a general and unified species concept, all of the properties that have been considered important by previous authors remain important for determining the numbers and boundaries of species, and they take on new importance in identifying those species most relevant to addressing particular questions. The main difference is they are no longer treated as necessary properties of species.
Another beneficial consequence of this proposal is that it removes the inconsistency between the proposition that the species is a fundamental category of biological organization and the way in which species are treated taxonomically. Under the general and unified species concept described above, the species would be the most general category at its particular level of biological organization. Consequently, species would be more directly analogous to the members of other fundamental categories
of biological organization, such as cells and organisms. Just as living beings need not (for example) be born, or sexually mature, or fully grown to be considered organisms, metapopulation lineages would not (for example) have to be diagnosable by fixed character differences, or monophyletic, or intrinsically reproductively isolated to be considered species. In other words, all separately evolving metapopulation lineages would be species (de Queiroz, 2005). Reinterpreting the properties in question as contingent rather than necessary properties of species would thus increase consistency between taxonomic practice and common assertions about the general theoretical significance of species.
ERNST MAYR AND THE MODERN CONCEPT OF SPECIES
The proposed resolution of the conflicts among alternative definitions of the species category described above is at odds with the common interpretation of Ernst Mayr’s popular species definition, which treats intrinsic reproductive isolation as a necessary property of species. Nevertheless, the proposal is highly compatible with, and might even be considered the culmination of, the general metapopulation lineage concept of species for which Ernst Mayr was arguably the most important spokesman. Evidence for the greater theoretical significance of the general metapopulation lineage concept of species relative to Mayr’s concise species definition can be found in at least three components of Mayr’s own writings on species: First, the reason he used the adjective “biological” to describe his species concept and definition; second, the properties of species Mayr viewed as important for distinguishing the new biological concept he advocated from older species concepts; and third, the distinction Mayr made between species concepts and species definitions, particularly in his early writings, that is, before his views were challenged by a proliferation of alternatives.
Mayr’s choice of the adjective “biological” as in the terms “biological species definition” and “biological species concept” has sometimes been criticized for being overly general (Simpson, 1961; Van Valen, 1976), thus raising the suspicion that it was chosen more for its rhetorical value than for its descriptive accuracy. However, an examination of Mayr’s writings on species reveals he had good reason for selecting this adjective. According to Mayr (1969a,b), “This species concept is called biological not because it deals with biological taxa, but because the definition is biological. It utilizes criteria that are meaningless as far as the inanimate world is concerned.” The important idea for Mayr was that earlier concepts of species were based on properties, such as degree of difference, that could be applied just as easily to inanimate objects as to living things. Linnaeus (1766–1768), for example, recognized species not only of plants and ani-
mals but also of rocks and minerals. In contrast, a truly biological concept of species must be based on properties that are unique to biological systems, properties such as reproduction and interbreeding.
It should be noted, however, that all contemporary species definitions (i.e., all definitions based on the general conceptualization of species as metapopulation lineages) are biological in the sense just described. The reason is that inanimate objects such as rocks and minerals lack reproduction and thus do not form populations or lineages in the uniquely biological way that organisms do. Moreover, with regard to the contingent properties commonly adopted as necessary properties of species, potential interbreeding (and its counterpart, intrinsic reproductive isolation) are no more biological than are a number of alternative properties, such as mate recognition, monophyly, and heterozygote deficits, all of which are also unique to biological systems. Thus, although it is appropriate to use the term “biological species concept” for the general concept of species as metapopulation lineages adopted by Ernst Mayr and virtually all other contemporary biologists, it is misleading to use this term for Mayr’s proposed species definition, that is, for the idea that a metapopulation lineage is not a species until it has become intrinsically reproductively isolated from all other such lineages.
Equation of the term “biological species concept” with the general concept of species as metapopulation lineages rather than the specific criterion of intrinsic reproductive isolation is supported by Mayr’s own writings, in particular, by the properties he identified as being important for distinguishing the concept of species in its newer and uniquely biological sense from older conceptualizations of the species category. According to Mayr (1963, 1970), there are three properties that “raise the species above the typological interpretation of a ‘class of objects’” and represent “one of the earliest manifestations of the emancipation of biology from an inappropriate philosophy based on the phenomena of inanimate nature.” These properties are first, that the members of a species constitute a reproductive community; second, that a species is an ecological unit that interacts with other species in its environment; and third, that a species is a genetic unit consisting of a large intercommunicating gene pool. These three properties are very general ones that apply to species (at least those composed of sexually reproducing organisms) under all definitions that conform to the general metapopulation lineage concept, that is, not only to those species recognized on the basis of the additional criterion of intrinsic reproductive isolation. One might even question whether potentially (rather than actually) interbreeding organisms are part of the same reproductive community and intercommunicating gene pool (Hull, 1965).
Further support for the equation of the term “biological species con-
cept” with the general metapopulation lineage concept of species, rather than a definition based on the property of intrinsic reproductive isolation, can be found in the terminology Mayr used to distinguish among these distinct ideas, at least in his early writings on the species. In those writings, Mayr distinguished more or less clearly between the general biological or metapopulation lineage concept of species and his attempt to describe that concept with a concise definition. Thus, in Systematics and the Origin of Species (1942), Mayr referred to the general concept as “the new species concept,” and he referred to his proposed definition as “a biological species definition” (not “the biological species definition”) (1942, p. 120). He also called it “a practical species definition,” emphasizing its utility for the practicing taxonomist and implying it was a compromise between theoretical and practical considerations (1942, p. 120). Later, in Animal Species and Evolution (1963), Mayr stated, “A study of all of the species definitions published in recent years indicates that they are based on three theoretical concepts, neither more nor less” (Mayr, 1963, p. 16). He called these three concepts “the typological species concept’” (used to refer to older species concepts that could be applied to inanimate objects), “the nondimensional species concept” (for the concept adopted by naturalists working at a single time and place), and the “interbreeding-population concept” (for the concept he advocated). He referred to concise descriptions proposed by both Dobzhansky and himself as “biological species definitions” (Mayr, 1963, pp. 19–20), apparently viewing both as falling under the general interbreeding-population concept.
In Mayr’s later writings (Grismer, 2001; Mayr, 1969a; Simpson, 1951), the nondimensional species concept was replaced with the nominalistic species concept (used to refer to the view that species are mental constructs invented to permit reference to several individuals collectively), and the terminterbreeding-population concept was replaced with biological species concept, thus demonstrating the equivalence between these terms. However, in these later works, Mayr’s own concise definition was presented as if it followed more or less directly from the general concept (“The species definition which results from this theoretical species concept is: [Mayr’s definition]”), and Dobzhansky’s definition was no longer mentioned. Subsequently, particularly in the writings of other authors (both pro and con), the distinction between the general biological (interbreeding-population) concept of species and Mayr’s species definition became further obscured by common reference to both ideas as the biological species concept.
In sum, there is an important distinction between the general concept of species as metapopulations or metapopulation lineages (the true biological species concept) and Ernst Mayr’s concise species definition. The
former is a very general theoretical concept that underlies virtually all modern views on species, including all contemporary species definitions. The latter describes a more restricted concept that uses the property of intrinsic reproductive isolation to facilitate taxonomic decisions concerning which metapopulation lineages are to be recognized as species (a practice that retains elements of an older view of the species category as a taxonomic rank and prevents full acceptance of the proposition that the species is a fundamental category of biological organization). Although this distinction is fairly clear in Mayr’s early writings, it has become obscured in the recent literature on species concepts. In any case, the concise definition with its treatment of intrinsic reproductive isolation as a necessary property of species is an important part of the species problem (the existence of alternative and partially incompatible definitions of the species category), whereas the general theoretical concept of species as metapopulation lineages forms the basis of a solution to this problem that results in a unified concept of species.
CONCLUSION
Ernst Mayr is almost certainly the greatest of all biologists in terms of his contributions to the development and acceptance of modern views on species. However, with regard to theoretical advances and their practical consequences, his most important contribution in this area was not his widely adopted definition of species but rather the major role he played in the development and advocacy of the general metapopulation lineage concept of species. This contribution had tremendous significance both for systematics in particular and for biology in general. It represented a fundamental shift in the conceptualization of the species category that resulted in a uniquely biological concept of species and changed the species category from a more-or-less arbitrary rank in the hierarchy of taxonomic categories to a basic category of biological organization. Moreover, because this important change in the conceptualization of the species category still has not been fully accepted, it continues to have important consequences. For example, as discussed in this paper, its more complete acceptance provides a simple solution to the species problem, and this solution, in turn, brings the way in which species are treated in taxonomic practice into line with claims about the general theoretical significance of the species category. In sum, Ernst Mayr’s ideas had tremendous importance, among many other things, for the development and acceptance of the modern metapopulation lineage concept of species, and they continue to provide the foundation for advances regarding the theoretical concept of species and its practical application.
ACKNOWLEDGMENTS
I thank Jody Hey, Walter Fitch, and Francisco Ayala for the invitation to present this paper at the Colloquium “Systematics and the Origin of Species: On Ernst Mayr’s 100th Anniversary.” Jody Hey provided valuable comments on the original manuscript.
REFERENCES
Andersson, L. (1990) The driving force: Species concepts and ecology. Taxon 39, 375–382.
Baum, D. A. & Shaw, K. L. (1995) Genealogical perspectives on the species problem. In Experimental and Molecular Approaches to Plant Biosystematics, eds. Hoch, P. C. & Stephenson, A. G. (Mo. Bot. Gard., St. Louis, MO), pp. 289–303.
Coyne, J. A. & Orr, H. A. (2004) Speciation (Sinauer, Sunderland, MA).
Cracraft, J. (1983) Species concepts and speciation analysis. Curr. Ornithol. 1, 159–187.
Darwin, C. (1859) On the Origin of Species by Means of Natural Selection (John Murray, London).
de Queiroz, K. (1997) The Linnaean hierarchy and the evolutionization of taxonomy, with emphasis on the problem of nomenclature. Aliso 15, 125–144.
de Queiroz, K. (1998) The general lineage concept of species, species criteria, and the process of speciation: A conceptual unification and terminological recommendations. In Endless Forms: Species and Speciation, eds. Howard, D. J. & Berlocher, S. H. (Oxford Univ. Press, Oxford, U.K.), pp. 57–75.
de Queiroz, K. (1999) The general lineage concept of species and the defining properties of the species category. In Species: New Interdisciplinary Essays, ed. Wilson, R. A. (MIT Press, Cambridge, MA), pp. 49–89.
de Queiroz, K. (2005) A unified concept of species and its consequences for the future of taxonomy. Proc. Calif. Acad. Sci. 56, in press.
Dobzhansky, T. (1937) Genetics and the Origin of Species (Columbia Univ. Press, New York).
Dobzhansky, T. (1950) Mendelian populations and their evolution. Am. Nat. 84, 401–418.
Dobzhansky, T. (1970) Genetics of the Evolutionary Process (Columbia Univ. Press, New York).
Donoghue, M. J. (1985) A critique of the biological species concept and recommendations for a phylogenetic alternative. Bryologist 88, 172–181.
Fitch, W. M. & Ayala, F. J. (1994) Tempo and mode in evolution. Proc. Natl. Acad. Sci. USA 91, 6717–6720.
Flexner, S. B. & Hauck, L. C. (1993) Random House Unabridged Dictionary (Random House, New York).
Ghiselin, M. T. (1997) Metaphysics and the Origin of Species (State Univ. of New York Press, Albany, NY).
Grismer, L. L. (1999) An evolutionary classification of reptiles on islands in the Gulf of California, Mexico. Herpetologica 55, 446–469.
Grismer, L. L. (2001) An evolutionary classification and checklist of amphibians and reptiles on the Pacific islands of Baja California, Mexico. Bull. South. Calif. Acad. Sci. 100, 12–23.
Harrison, R. G. (1998) Linking evolutionary pattern and process. In Endless Forms: Species and Speciation, eds. Howard, D. J. & Berlocher, S. H. (Oxford Univ. Press, New York), pp. 19–31.
Hull, D. L. (1965) The effect of essentialism on taxonomy—Two thousand years of stasis (II). Brit. J. Philos. Sci. 16, 1–18.
Hull, D. L. (1980) Individuality and selection. Annu. Rev. Ecol. Syst. 11, 311–332.
Huxley, J. (1942) Evolution, the Modern Synthesis (Allen and Unwin, London).
Jordan, K. (1896) On mechanical selection and other problems. Novit. Zool. 3, 426–525.
Jordan, K. (1905) Der Gegensatz zwischen geographischer und nichtgeographischer Variation. Z. Wiss. Zool. 83, 151–210.
Linnaeus, C. (1753) Species Plantarum (Laurentii Salvii, Stockholm).
Linnaeus, C. (1758) Systema Naturae, 10th Ed. (Laurentii Salvii, Stockholm).
Linnaeus, C. (1766–1768) Systema Naturae, 12th Ed. (Laurentii Salvii, Stockholm).
Mallet, J. (1995) A species definition for the Modern Synthesis. Trends Ecol. Evol. 10, 294–299.
Martin, G. (1996) Birds in double trouble. Nature 380, 666–667.
Mayden, R. L. (1997) A hierarchy of species concepts: The denouement in the saga of the species problem. In Species: The Units of Biodiversity, eds. Claridge, M. F., Dawah, H. A. & Wilson, M. R. (Chapman & Hall, London), pp. 381–424.
Mayr, E. (1942) Systematics and the Origin of Species (Columbia Univ. Press, New York).
Mayr, E. (1955) Karl Jordan’s contribution to current concepts in systematics and evolution. Trans. R. Entomol. Soc. Lond. 107, 45–66.
Mayr, E. (1963) Animal Species and Evolution (Harvard Univ. Press, Cambridge, MA).
Mayr, E. (1969a) Principles of Systematic Zoology (McGraw–Hill, New York).
Mayr, E. (1969b) The biological meaning of species. Biol. J. Linn. Soc. 1, 311–320.
Mayr, E. (1970) Populations, Species, and Evolution (Harvard Univ. Press, Cambridge, MA).
Mayr, E. (1982) The Growth of Biological Thought: Diversity, Evolution, and Inheritance (Belknap Press of Harvard Univ. Press, Cambridge, MA).
Mayr, E. & Provine, W. B., eds. (1980) The Evolutionary Synthesis: Perspectives on the Unification of Biology (Harvard Univ. Press, Cambridge, MA).
Meglitsch, P. A. (1954) On the nature of species. Syst. Zool. 3, 49–68.
Michener, C. D. (1970) Diverse approaches to systematics. Evol. Biol. 4, 1–38.
Nixon, K. C. & Wheeler, Q. D. (1990) An amplification of the phylogenetic species concept. Cladistics 6, 211–223.
Paterson, H. E. H. (1985) The recognition concept of species. In Species and Speciation, ed. Vrba, E. S. (Transvaal Museum, Pretoria, South Africa), pp. 21–29.
Poulton, E. B. (1903) What is a species? Trans. Entomol. Soc. London 1903, lxxvii–cxvi.
Rosen, D. E. (1979) Fishes from the uplands and intermontane basins of Guatemala: Revisionary studies and comparative geography. Bull. Am. Mus. Nat. Hist. 162, 267–376.
Simpson, G. G. (1951) Evolution (Lawrence, Kans.) 5, 285–298.
Simpson, G. G. (1961) Principles of Animal Taxonomy (Columbia Univ. Press, New York).
Sneath, P. H. A. & Sokal, R. R. (1973) Numerical Taxonomy: The Principles and Practice of Numerical Classification (Freeman, San Francisco).
Sokal, R. R. & Crovello, T. J. (1970) The biological species concept: A critical evaluation. Am. Nat. 104, 127–153.
Templeton, A. R. (1989) The meaning of species and speciation: A genetic perspective. In Speciation and Its Consequences, eds. Otte, D. & Endler, J. A. (Sinauer, Sunderland, MA), pp. 3–27.
Van Valen, L. (1976) Ecological species, multispecies, and oaks. Taxon 25, 233–239.
Wright, S. (1940) The statistical consequences of Mendelian heredity in relation to speciation. In The New Systematics, ed. Huxley, J. (Oxford Univ. Press, London), pp. 161–183.
Zink, R. M. (1996) Bird species diversity. Nature 381, 566.






















