
4
Effects of Variations in Pollinator Populations on Pollination Services
Because pollinators provide essential ecosystem services in agricultural and natural areas, it is valuable to determine whether pollination services are threatened by their declining populations. Many plant species rely on pollinators for sexual reproduction, but whether a global “pollination crisis” is jeopardizing food supplies or wild biodiversity has been debated (Buchmann and Nabhan, 1996; Ghazoul, 2005a,b; Potts et al., 2005; Steffan-Dewenter et al., 2005). Chapter 2 noted that a pollinator shortage exists when the demand for pollination services exceeds available pollinator capacity. Shortages can be exacerbated by pollinator declines, but they can arise even when pollinator populations are stable or increasing over time. Some shortages, for example, those caused by the high seasonal demands of the California almond industry for honey bee colonies (for example, Norton, 2005), can be moderated by market forces, like the economics of supply and demand. In contrast, chronic and continuing pollinator declines could threaten the basic ecosystem services that many people take for granted. In the study of consequences for pollination and ecosystem services, it is useful to distinguish between transient pollinator shortages and declines.
POLLINATORS IN AGRICULTURE
Food Security and Nutrition
An evaluation of experimental evidence for pollination requirements of 107 globally traded fruit and vegetable crops (representing 40 percent of global plant-based food production) by Klein et al. (2007) shows that
animal pollination improves production in 75 percent of the crops studied. Most cultivars of another 10 percent of the crops require animal pollination. Another 8.5 percent of the crops do not benefit from animal pollination and its role in production of the remaining 6.5 percent crops is not known. Many crops, however—notably the staple grains that form the foundation of most human diets (rice, wheat, maize, sorghums, millets, rye, barley)—are self-pollinating or pollinated by the wind. Together, species that do not rely on pollinators account for most of the world’s food supply by weight (FAO, 2005).
Pollinator declines, therefore, do not fundamentally threaten the world’s caloric supplies. However, fruits and vegetables, which add diversity to the human diet and provide essential nutrients, tend to depend heavily on pollinators (Prescott-Allen and Prescott-Allen, 1990; Roubik, 1995). Seven of the nine crops that provide at least 50 percent of the vitamin C available to the human diet globally depend partially or entirely on animal pollination for the production of fruits or seeds (oranges, cabbages, green peppers, tomatoes, melons, tangerines, watermelon) (FAO, 2005; Free, 1993; McGregor, 1976; USDA-NASS, 2006b).
Animal-pollinated crops also tend to have greater economic value than do crops that are not animal-pollinated, and they provide relatively more income to farmers per unit of land (USDA-NASS, 2006b). Moreover, several oilseed crops depend on pollinators, and bee pollination is required to produce the seeds of major forage and hay crops, such as alfalfa and clover, that feed the animals that supply meat and dairy products. Thus, despite the fact that they do not provide the bulk of the human diet, animal-pollinated plants contribute importantly to the quality of the human diet. Although estimates of the proportion of the human diet that is attributable to animal pollination are occasionally attempted and frequently cited (for example, McGregor’s 1976 estimate that one-third of the human diet can be traced directly or indirectly to animal pollination), the proportion likely varies among countries and regions and depends on dietary preferences, seasonal availability, cultural practices, and economic status of consumers.
Major Crops
Plant species grown as agricultural commodities display a wide variety of breeding systems. Some readily self-pollinate; others are dioecious, monoecious, or self-incompatible and require animal pollination (Free, 1993; McGregor, 1976; Table 4-1). Dependence on pollinators also varies greatly among crops. In the United States, about 130 agricultural crops benefit from insect pollination (McGregor 1976; see examples in Table 4-1). Most North American crops can produce some fruits and seeds without animal pollination, but pollination often increases their value through higher yields or im-
TABLE 4-1 Value of Common Crops That Require or Benefit from Insect Pollination
|
Commodity |
Bearing Acres |
Production Unit |
Utilized Production (thousand units) |
Value of Utilized Production ($ thousand U.S.) |
|
Apple |
395,000 |
Ton |
4,261.6 |
1,783,952 |
|
Blueberry, cultivated |
41,720 |
Ton |
94.4 |
221,610 |
|
Blueberry, wild |
NA |
Ton |
40.2 |
28,540 |
|
Cranberry |
39,600 |
Ton |
309.2 |
208,025 |
|
Cherry, sweet |
73,940 |
Ton |
246.8 |
344,471 |
|
Cucumber, fresh |
58,600 |
Hundredweight |
55,000.0 |
187,391 |
|
Melon, cantaloupe |
86,000 |
Hundredweight |
22,107.0 |
371,721 |
|
Melon, watermelon |
149,600 |
Hundredweight |
38,208.0 |
342,918 |
|
Nut, almond |
550,000 |
Ton |
833.3 |
1,600,144 |
|
Peach |
145,530 |
Ton |
1,205.1 |
454,532 |
|
Pear |
64,150 |
Ton |
922.5 |
270,385 |
|
Squash, all |
50,700 |
Hundredweight |
7,685.0 |
197,020 |
|
SOURCE: Data from NASS Non-citrus Fruits and Nuts 2003 Summary. |
||||
proved quality (Klein et al., 2007). Coffee self-pollinates, but yields increase as a result of pollinator visits (Klein et al., 2003a,b,c; Ricketts et al., 2004; Roubik, 2002). A few U.S. crops—melons, cucurbit squashes, almond, and most pome fruits—are completely dependent on animal pollination to set fruit. Crops with many-seeded fruits, such as watermelon, pumpkin, and kiwi, have hundreds of ovules so they require many pollen grains per stigma. Thus, they are more susceptible to pollinator shortages than are those with few seeds per fruit (Free, 1993; Stanghellini et al., 1997).
In a detailed 1976 report, Insect Pollination of Cultivated Crop Plants, published by the U.S. Departmnent of Agriculture, McGregor (p. iv) noted that “the pollination of plants, essential in the perpetuation of most species, is so unobtrusively accomplished that it is often overlooked.” Although pollinators are critical to many agroecosystems, their importance varies not only among crop species but even among varieties of the same crop. In addition, wild-pollinator abundances can fluctuate dramatically among locations and over time (Kremen et al., 2002b; Price et al., 2005). Reliable data on the specific contributions of pollinators to crop yields are often lacking, and obtaining this information requires large-scale, long-term studies. Listed in Table 4-1 are the relative area and value of several major crops known to benefit from pollinators. Because managed honey bees are often used to pollinate them, the extent to which managed and unmanaged pollinators provide adequate pollination service for optimal yields remains for the most part undetermined.
Managed and Wild Crop Pollinators
Crop plants in North America are pollinated by many wild or managed native species of animals, in addition to managed colonies of introduced honey bees (Apis mellifera). The wild species include native bees, feral honey bees, other insects, birds, and bats (Free, 1993; Chapter 2). For some crops, one or more wild species could be more efficient than honey bees, although honey bees also might contribute to pollination (Free, 1993). Blueberry, cranberry, and kiwi are “buzz-pollinated”—a visiting bee vibrates its body to sonicate the pollen grains out of the flowers’ anthers. Bumble bees (Bombus spp.) and some other native bees are much more efficient blueberry pollinators than are honey bees because they buzz-pollinate (Buchman, 1983; Cane and Payne, 1990; Javorek et al., 2002; MacKenzie, 1997; MacKenzie et al., 1996), whereas honey bees cannot. However, honey bees are used to pollinate wild blueberries because apiculture is well developed. Even in crops that are routinely pollinated by managed honey bees, wild pollinators also can contribute pollination services. Watermelon growers in California use managed honey bees, but their crops also receive substantial pollination from a diverse community of native bees (Kremen et al., 2002a).
The adequacy of pollination services provided by managed and wild pollinators, including feral honey bees, can vary widely and is poorly documented for many crops (Free, 1993). Greenleaf and Kremen (2006b) studied field-grown tomatoes for which growers did not import honey bee colonies. They found that native bees (such as Anthophora spp. and Bombus spp.) were buzz-pollinating the crops. Although wild species are either known or likely to be important pollinators in many crop systems, the relative contributions of different groups of pollinators to crop yields are rarely defined. With such a paucity of relevant research, it is useful to review evidence from a sample of well-studied crops to evaluate the availability and importance of wild pollinators. For these exemplars, pollinator shortages often can be overcome by providing sufficient numbers of managed pollinators, especially honey bees, although in a few cases the crop production depends completely on specific pollinators (for example, midges or bats).
Watermelon
Watermelon (Citrullus lanatus) is grown in several parts of the United States and Mexico. U.S. production in 2005 was worth $410 million (USDA-NASS, 2006b). Because watermelon has separate male and female flowers, animal pollination is critical. Watermelon pollination requirements are high: 500–1,000 pollen grains must be deposited on stigmas to produce marketable fruits (Adlerz, 1966; McGregor, 1976), so the plant relies on multiple insect visits to each female flower for successful pollination (Stanghellini et al., 1997). In most areas of cultivation within North America, pollination
is provided by managed honey bees and a diverse set of native bee species (Kremen et al., 2002a).
In the central valley of California, Kremen and colleagues (2002a) measured pollination at 14 watermelon farms that differed in degree of isolation from natural habitat. At farms near natural habitat, the native bee community was sufficient to provide adequate pollination. At farms far from natural habitat, however, average pollen deposition by native bees alone was insufficient. The inadequacy was the result of a reduction in overall pollinator abundance and of local extinction of the most efficient pollinators (Kremen et al., 2004; Larsen et al., 2005).
Blueberry
Blueberries (Vaccinium spp.) grow in several Canadian provinces and in several U.S. states. High-bush blueberries are actively cultivated; low-bush blueberries are harvested from semicultivated wild plants. In the United States, about 44,000 acres (17,800 hectares) of high-bush blueberries are planted to produce a crop worth more than $275 million annually (USDA-NASS, 2006b). Like many other species in their family, blueberries are buzz-pollinated, primarily by bumble bees (Buchmann, 1983; Free, 1993). Introduced honey bees do not buzz-pollinate, so high-bush blueberries depend largely on native wild bees for pollination (Buchmann, 1983; Free, 1993; MacKenzie, 1997; MacKenzie et al., 1996).
Several research groups have investigated the dependence of blueberry production on wild bees. Cane and Payne (1988), for example, listed four species of wild bumble bee (Bombus spp.) and the anthophorine Habropoda laboriosa that buzz-pollinate high-bush blueberry fields in Alabama and Georgia. Honey bees from colonies at field borders were found to carry little or no blueberry pollen. In New Brunswick, Canada, pesticide applications in nearby forests were followed by devastating reductions in native populations of bumble bees, andrenid bees, and halictid bees, which might have contributed to a severe drop in low-bush blueberry production (Kevan, 1975b). When a less harmful insecticide was introduced, native bees appeared to recover (Kevan et al., 1997), and production returned to normal (Kevan and Plowright, 1995). These findings highlight both the dependence of the crop on wild pollinators and the interconnectedness of contiguous natural and cultivated habitats.
Sunflower
Sunflowers are grown throughout the world for oil and confection seed, and in 2005, 2.7 million acres were under production in North America, mostly in the United States (FAO, 2005). Sunflower, Helianthus annuus,
is a widely distributed native plant in North America, visited by many of its native pollinator species, including some bee species that specialize on sunflower pollen (for example, Svastra obliqua). Unlike the sunflower, many crops are grown outside their native ranges and therefore are visited by novel pollinators. The sunflower head consists of many individual florets, each of which passes first through a male stage and then a female stage (Free, 1993). Although the plant is self-compatible, bees often increase sunflower seed set by moving pollen from male-stage florets to female-stage florets within the same flower head and through cross-pollination between individual plants (Greenleaf, 2005; Greenleaf and Kremen, 2006b). Most sunflower varieties are sold as hybrid seeds, which require cross-pollination between pollen-producing lines and those that lack pollen. The hybrid sunflower seed production industry is worth $26 million annually in the United States (Greenleaf, 2005), and more than 90 percent of seed production occurs in northern California (Lilleboe, 2000).
Several studies have documented improvements in sunflower seed production as a result of insect pollination, although the degree of improvement varies among cultivars. For example, seed set on flower heads increased from 1 to 35 percent when bees provided self-pollination, transferring pollen among florets; when bees moved pollen between florets on different sunflower heads, seed set increased to 63 percent (Free, 1993). Although managed honey bee hives are stocked in hybrid sunflower fields in the United States, non-Apis bees also pollinate hybrid sunflowers and might do so far more efficiently than honey bees (DeGrandi-Hoffman and Watkins, 2000; Greenleaf, 2005; Parker, 1981). Non-Apis bee interactions with honey bees increase the pollination efficiency on hybrid sunflower by as much as five times, by disturbing them as they forage on florets and causing them to move among plants more often and cross-pollinate plants more effectively (Greenleaf, 2005). From those findings, Greenleaf (2005) calculated that the direct pollination provided by wild bees nationally to the hybrid sunflower seed industry was worth $1.9 million, the interspecific interaction between wild and honey bees was worth $10.4 million, and the direct contribution of honey bees was worth $13.8 million. Both proximity to natural habitat and crop rotation practices affect the amount of sunflower pollination provided by wild bees (Greenleaf, 2005; Greenleaf and Kremen, 2006b).
Canola
Canola is Canada’s fourth largest crop in terms of area under cultivation, and annual exports are currently worth more than $2 billion Canadian (Morandin and Winston, 2005). The two species in the mustard family grown to produce canola oil, Brassica napus and B. rapa, differ in dependence on pollinators. B. napus is self-compatible, but some cultivars
produce better yield with cross-pollination (Kevan and Eisikowitch, 1990). In contrast, B. rapa is self-incompatible and therefore requires pollinators for seed set. Although managed honey bees are used, many native species also pollinate canola crops (Morandin and Winston, 2005).
Morandin and Winston (2005) studied pollination of self-incompatible canola in Alberta, Canada, and compared results from fields with different management regimes. They reported that farms with more wild bees also showed smaller pollination deficits, which were defined as the extent to which yields were depressed because of insufficient pollination. The farms with the fewest bees were those planted in genetically modified, herbicideresistant plants. Because those fields were treated heavily with herbicides, they had a lower diversity and abundance of flowering weeds, which could account for their reduced capacity to support wild bees. Using those results, Morandin and Winston (2006) calculated that profits lost by retiring up to 30 percent of the field area from production would be more than offset by increased yield that would result from better pollination by the more diverse and abundant pollinators found in fields within 750 meters of uncultivated areas.
Coffee
Coffee (Coffea arabica and C. robusta) is grown extensively in Mexico and is one of the developing world’s most valuable export commodities. Worldwide, 11 million hectares are planted in coffee, and the industry employs 25 million people (O’Brien and Kinnaird, 2003). C. arabica, the species thought to produce the highest-quality coffee, self-pollinates, but yields increase 15 to 50 percent when flowers are visited by bees (for example, Free, 1993; Klein et al., 2003b; Roubik, 2002). Despite that, few coffee farmers recognize pollination as an issue for their crops (Ricketts et al., unpublished data).
Several recent studies have investigated the importance of wild bees to coffee production. Ricketts and colleagues (Ricketts, 2004; Ricketts et al., 2004) reported that in Costa Rica a diverse community of wild bees pollinates coffee. The pollinators were mainly wild Africanized honey bees and 10 species of social stingless bee (tribe Meliponini). The species depend on forest habitat for nest sites and food resources but flew into nearby coffee fields when they were in flower. Coffee fields near forest patches received more visits by a more diverse set of bees and had 20 percent higher yields than did fields more than 1 km away from a forest. Klein and colleagues (Klein et al., 2002, 2003c) also noted a diverse community of wild coffee pollinators in Indonesia. They reported that coffee yields increased with increased diversity of pollinators and that pollinator diversity increased near forest remnants and in less intensive farms managed with a more diverse
canopy of shade trees. Although these studies were performed outside North America, their findings are informative for coffee production in Mexico.
Cacao
Chocolate is made from the seeds of the cacao tree (Theobroma cacao), which is native to the western Amazon basin, but now grown in many humid tropical regions, including Mexico (Clay, 2004). Cacao trees can be either self-compatible or self-incompatible, but even self-compatible trees require insect visits to move pollen from anthers to stigmas and pollinate the flowers. Honey bees do not appear to be effective pollenators (Free, 1993). Although the identity of the principal pollinators of cacao eluded growers and scientists for many years, midges in the family Ceratopogonidae have been identified as the most important group (Free, 1993; Young, 1985). Many other wild insects also have been observed visiting cacao (bees in the subfamilies Halictinae, Meliponinae), but their pollinating efficiency is either low or undetermined (for example, Young, 1981, 1985).
There is substantial evidence that farm management practices influence the amount and quality of habitat for midges within and around cacao farms and therefore can affect yields (Free, 1993). Midges prefer shady, moist conditions; their eggs are laid and larvae develop in rotting wood and leaves on the ground (Free, 1993). Young (1982, 1986) showed that adding pieces of rotting wood to the leaf litter within cacao plantations increased the abundance of several midge species and that cacao plants near shade trees or rotting material often had more fruits than did trees that were farther away.
Agave
Tequila—one of several alcoholic beverages derived from agave—is produced exclusively from Agave tequilana var. azul in select regions of Mexico. Tequila revenues were close to $1 billion in exports and domestic sales in 2000 (Tequila Aficionado, 2001), and in 1998, Mexico had about 100,000 hectares (about 250,000 acres) containing roughly 200 million plants (Ramírez, 1998). After 1989, and at least until 2002, the tequila industry faced major agave shortages caused by several diseases that affected about 30 percent of the plants at different stages of development (Jiménez-Hidalgo et al., 2004; Larrea-Reynoso, 1998; Valenzuela-Zapata, 1994).
Although many species of agave, including A. tequilana, are naturally pollinated by bats and other animals (Arizaga et al., 2002; Rocha et al., 2005), the industry uses only asexually reproduced plants to replenish the producing population. This practice has prevented outcrossing of individuals and resulted in loss of genetic diversity compared to other agaves (Dalton,
2005). Recent studies have suggested links between the disease outbreaks to this reduction of genetic diversity (Valenzuela-Zapata and Nabhan, 2004), and producers are supporting research to find solutions (Dalton, 2005). The tequila industry very likely would benefit if genetic diversity were maintained and used in various combinations in the fields. Monocultures with little genetic variation are susceptible to pests and diseases.
Other Crops
Ecologists have investigated a variety of crops, both in North America and elsewhere, to determine the degree of pollination services provided by wild bees. Some studies demonstrate extensive pollination by wild pollinators (Roubik, 1993, 1995), whereas others find little support for this assertion (reviewed in Free, 1993; Klein et al., 2007). Additional studies on pollination of muskmelon (cantaloupe), almond, and tomato in California, New Jersey, and elsewhere are under way (Greenleaf and Kremen, 2006a; Kremen, unpublished data; Williams and Kremen, unpublished data; Winfree et al., 2006). The growing body of research will contribute to elucidating the degree to which wild pollinators benefit agricultural production and, more important, the circumstances and management regimes that affect pollination of crops by unmanaged wild pollinators.
Alleviating Pollinator Shortages
Traditional farmers and plant breeders have developed new cultivars that depend less on pollinators. Cultivated hybrid sunflowers (Helianthus annuus) self-pollinate even though their insect-pollinated wild ancestors are self-incompatible (Heiser, 1954). In the early 1900s, strawberries were developed with hermaphrodite flowers to improve fruit set (McGregor, 1976), although they still benefit from pollinators. Similarly, flowers of wild tomatoes and some heirloom tomato varieties must be visited by bumble bees to set fruit. Most cultivated tomatoes can self-pollinate if they are jostled mechanically (McGregor, 1976), but greenhouse tomatoes typically are buzz-pollinated by managed bumble bees (Box 3-1). In field production, some varieties of tomatoes also produce more and larger fruits if they are visited by bees (Greenleaf and Kremen, 2006a).
Bananas and some varieties of pineapple, grape, citrus, cucumber, and persimmon can produce fruit without sexual fertilization, a process known as parthenocarpy. Parthenocarpic fruit production solves problems associated with insufficient numbers of pollinators by bypassing them altogether. Parthenocarpy can be induced with the use of plant hormones and genetic modification (Gorguet et al., 2005; Pagnotta, 1999; Rotino et al., 1997). Mainland and Eck (1968) induced fruit production in blueberries with
applications of auxin, gibberellin, and kinin, and Hayata and colleagues (1995) and Maroto and colleagues (2005) used plant regulators to induce parthenocarpic fruit production in watermelons. In a report that detailed parthenocarpy induced by genetic modification, Acciarri and colleagues (2002) reported improved fruit production in eggplant using the DefH9iaaM auxin-synthesizing gene. Donzella and colleagues (2000) reported the same results. Mezzetti and colleagues (2002a,b, 2004) used the DefH9-iaaM gene to induce parthenocarpic fruit production in strawberry and raspberry cultivars. Barg and colleagues (2001) reported successful production of parthenocarpy in tomato cultivars by use of a chimeric transgene. Conventional and molecular techniques have been used to produce parthenocarpic cultivars of citrus (Koltunow et al., 2000). The development of additional parthenocarpic varieties through traditional breeding or recombinant DNA techniques could expand the number of crops that do not require pollination by animal vectors. Nonetheless, pollinators contribute to maintenance of plant genetic diversity (see Agave example above) and are important for breeding programs (such as hybrid or other seed production).
In some cases, the problem of insufficient cross-pollination can be addressed by better planting designs. For example, apples are self-incompatible and require cross-pollination not only between individuals but also between cultivars, and movements of honey bees from tree to tree are limited. Using genetic markers, Kron and colleagues (2001a,b) demonstrated that intercultivar pollen movement by pollinating honey bees in orchards of dwarf apple was limited to about four neighboring trees. Thus, plantings with too many rows of the same cultivar produced lower yields. In such a situation, the efforts of the pollinators (and the money spent by grower) are wasted, and in essence a pollinator shortage is created unnecessarily. Moreover, Kron and colleagues (2001a,b) noted that the pollen of some apple cultivars is more vigorous than is that of others. Planting arrangements can be designed to increase the efficiency of available pollinators. Another method of alleviating pollinator shortages is to protect and enhance the habitats of wild pollinators. Strategies for improving pollinator habitats in and around agricultural areas are discussed in Chapter 6.
Economic and Financial Consequences of Pollinator Shifts
Honey bees are responsible for the greatest percentage of agricultural pollinator activity in North America, and they constitute the greatest percentage of the commercial pollination market. Commercial beekeepers, who account for an estimated 99 percent of all reported pollination rentals in North America (Burgett, 2004), earn their income from pollination fees and honey sales. Honey revenues averaged $143 million annually from 1986 to 2005 (USDA-NASS, 1995, 1999, 2004b, 2005), and reached record highs
in 2002–2003 (USDA-NASS, 2005). Although pollination fee data are not reported nationally, the total has been estimated at $150 million: $75 per hive for each of the 2 million hives in commercial hands (Rucker et al., 2005).
Commercial beekeepers divide their honey bees’ time between crops that contribute to pollination income and crops that yield marketable honey. Honey production is high for alfalfa, clover, and vetch crops. Almonds, however, which create North America’s greatest demands for managed honey bees, do not provide commercially competitive honey. Pollination fees tend to be higher for crops that need pollination, but produce poor honey. Burgett and colleagues (2004) reported average pollination fees for honey crops (vetch seed, crimson clover seed, red clover seed, radish seed, blueberry) between 1987 and 2002 ranged from $2.67 to $23.29 per colony, in 2002 dollars. By contrast, fees for nonhoney crops (cucumber, pear, sweet cherry, squash, apple, cranberry, almond) were higher: $25.21 to $38.24. Rucker and colleagues (2005) reported that honey production and pollination services tend to compete for beekeepers’ services and higher honey prices are associated with higher fees for pollination.
The largest concentrations of honey bee colonies are maintained in four regions: the Dakotas and the northwestern tier states, where large areas of vetch and clover produce high-quality honey; California, with almond, fruit, and melon crops; Florida, with orange blossom as the major March and April crop and with winter vegetables and small fruits; and Texas, with winter vegetables (USDA-NASS, 2006). The mobility of commercial bee colonies on tractor trailers creates three major south-to-north pollination routes: West Coast, Midwest, and East Coast.
The economic consequences of pollinator shortages in agriculture hinge on six determining factors: a specific need for animal pollination, a crop yield gain contributed by the pollinator, the crop price, the cost of pollination services, the value of marketed byproducts (such as honey), and the availability of alternative means of pollination.
Predicting the direct, short-run economic consequences of population declines in honey bees—the principal managed pollinator species—is not straightforward. Microeconomic theory predicts two effects; both increase pollination fees. The first is a rise in cost of producing and maintaining commercial bees. This increase reduces the quantity of pollinators that can be supplied at any price (leftward shift in the supply curve, illustrated in Figure 4-1). The second is an increase in demand for commercial honey bee pollination caused by decreases in feral bee colonies and native pollinator numbers or by an increase in acreage or floral density of pollinator-dependent crops (rightward shift in the demand curve, Figure 4-1).
The economic effects of a decrease in abundance of insect pollinator are likely to be greatest for crops that rely heavily on insect pollination. Morse
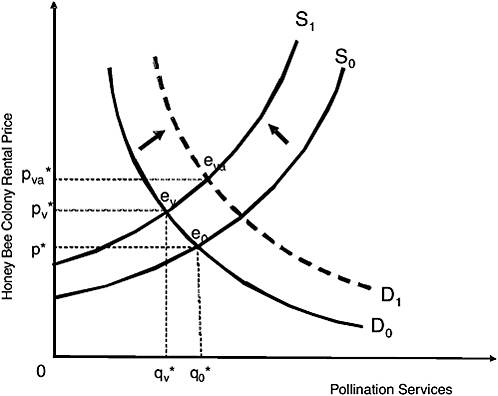
FIGURE 4-1 Honey bee population declines raise bee production and maintenance costs, reducing the commercial supply of pollination services offered at all price levels from S0 to S1, and raising the market price and reducing the marketed quantity of honey bee colony rentals from e0 to ev. Agricultural demand for pollination services could rise (from D0 to D1) as feral honey bees and native pollinators decrease in abundance or as crop acreage rises (as in almonds), increasing both the equilibrium price and the quantity to point eva.
and Calderone (2000) estimated that five major fruit crops are 100 percent reliant on insect pollination: almond, apple, avocado, blueberry, and cranberry (see Box 4-1). Most calculations of the value of pollination services use current market prices (for example, Morse and Calderone, 2000). However, if a major decrease in pollinator availability were to occur, the prices of crops that depend on pollination would rise, increasing the economic value of the remaining pollination services (Southwick and Southwick, 1992) and reducing consumer welfare (Kevan and Phillips, 2001). The combination of rising production costs and falling yields also could prompt financially marginal producers of pollination-dependent crops to exit the industry or shift to crops that do not depend on pollinators. Both increased food costs
|
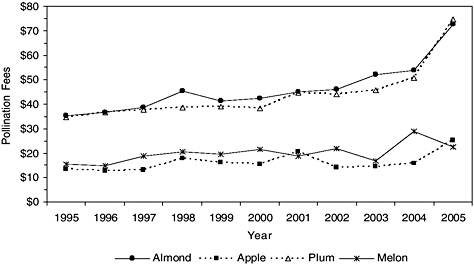
BOX 4-1 Pollination Fees for California Almonds The California almond industry is contending with record high prices for rentals of honey bee colonies. Average fees jumped from $35 per colony in the late 1990s to $75 in 2005 (Burgett, 2005; Burgett et al., 2004) (Figure 4-2), and fees can go as high as $150 per colony (http://beesource.com/pov/traynor/bcnov2005.htm). Given that the U.S. commercial honey bee population remained fairly stable (on a year-to-year basis) between 1996 and 2004 (USDA-NASS, 1999, 2004a, 2005), the increased cost cannot be attributed solely to honey bee decline. Three other factors are also responsible for the higher pollination fees. First, California almond acreage expanded more than 35 percent (from 405,000 to 550,000 acres) between 1991 and 2004, during which time varroa mites infestations were widely detected (Pollack and Perez, 2005). The current bearing acreage in almonds requires 1.4 million colonies for pollination during the February flowering period (Heinz, 2005), and that figure constitutes more than 60 percent of the U.S. supply of commercial honey bees. Second, honey bee colonies have experienced significant winter mortality that is attributable to varroa mite infestation that leads to seasonal colony shortages each February because beekeepers have not had time to rejuvenate colonies. Third, the price of honey has been relatively high since 2002. Almond nectar is bitter, so beekeepers who market honey avoid almonds as forage plants. Indeed, honey traditionally competes with pollination for the beekeeper’s business: when honey prices go up, pollination rental fees climb as well (Rucker et al., 2005). How high can almond pollination fees go? Recent almond plantings mean that bearing acreage will continue to rise. Given that almonds are almost |
and reduced agricultural employment would have negative multiplier effects in the reduction of spending on other goods and services.
The prices of pollination byproducts and the availability of alternative means of pollination also should be considered. A decline in managed honey bee populations should trigger a rise in the price of honey. And the rising cost of pollination and the income lost as a result of lower crop yields should trigger research and development of alternative means of pollination.
Evidence suggests that some of the hypothesized economic effects of pollinator decline have, in fact, occurred in the United States since the mid-1980s when tracheal and varroa mites began to spread. Statistical regression analysis of annual pollination fee data from Washington and Oregon shows that honey bee colony rental fees for pear, cherry, and apple rose by $4.40–$5.30 (in 2002 dollars) after 1991, when the varroa mite was widely
detected in the Pacific Northwest (Rucker et al., 2005). As noted by Burgett and colleagues (2004), if each honey bee colony is rented out two to three times per year, the increase in pollination fees roughly equals the estimated annual cost of varroa mite control at $10–$15 per colony. By extrapolation from their Pacific Northwest data to the 2 million commercial hives in the United States, varroa mite control has increased honey bee colony rental fees by nearly $30 million annually (ignoring any honey or crop yield losses that could result from honey bee shortages).
The rising cost of varroa mite control and the high prices of domestic honey during 2002–2003 (Figure 4-3) might also be responsible for the subsequent increase in honey bee pollination fees in 2003–2005. The rise in pollination fees was aggravated by the hard winter of 2004–2005, which caused a 5.7 percent decline in the reported U.S. honey bee population
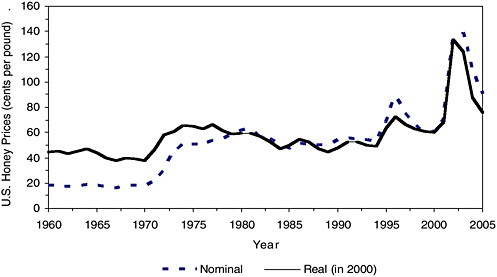
FIGURE 4-3 U.S. honey prices (cents per pound) in nominal and real (2000) terms.
SOURCES: Data compiled from prices listed by Hoff and Phillips (1989); USDA-NASS (1995, 1999, 2004b, 2005, 2006); producer price index data for farm products from the Council of Economic Advisors (2006).
(USDA-NASS, 2006). In response to the early season shortage of honey bee colonies, beekeepers imported “package” honey bees from Australia for the 2005 California almond pollination as substitutes for domestic honey bees (Sumner and Boriss, 2006; USDA-APHIS, 2004).
The U.S. commercial honey bee population was stable from 1996 to 2004, but if it were to continue to decline at the rates exhibited from 1947 to 1972 and from 1989 to 1996, it would vanish by 2035. Although that is conceivable, there are strong incentives to protect honey bees and, failing that, to develop alternative methods of pollination. The long-term economic consequences of resumed honey bee population decline will depend on how farmers, beekeepers, scientists, and others respond to smaller populations and higher fees. Writing about almond pollination, pomology consultant Wesley Asai predicted that “growers will have greater interest in new varieties that are self-fertile and possibly even self-pollenizing to reduce dependence on bees” (2005). Agricultural research has a long history of innovation (Hayami and Ruttan, 1985), and new work could focus on mite control, bee tolerance of mites, or crop pollination needs. Another potential response is the introduction of substitute pollinators—propagated bumble bees or the imported Australian packaged honey bees, like those used to pollinate California almonds in 2005 (Sumner and Boriss, 2006). Of course, a new long-term market equilibrium could be established with
higher pollination fees that eventually would be seen in higher prices at the supermarket.
Over the long term, the indirect effects of honey bee shortages will depend on how markets and technology respond. If the pollination markets adjust to a new, higher price equilibrium, then higher costs could undermine the profitability of crop production in marginal areas. Increased food prices also are possible, although whether they occur would depend on competition from fruit, nut, and vegetable producers outside North America.
POLLINATORS IN NATURAL AREAS
Pollinator Limitation of Seed Production
As with agricultural systems, pollinator limitation in natural plant populations occurs through a variety of mechanisms that decrease pollinator abundance below that required by plants for full reproduction, and pollinator shortages are expected to depress fruit and seed set in local plants. Low seed set can be caused by other factors as well, including severe weather, inadequate concentrations of soil nutrients, disease, damage caused by herbivores, partial sterility, or shortages of neighboring plants with compatible pollen. Thus, in plant populations where reproduction is low or appears to be declining, explanations other than inadequate pollination should first be ruled out (Figure 4-4). It is difficult to study the efficacy of pollination services because it is usually impossible to augment native pollinator populations artificially in controlled experiments (Thompson, 2001). Therefore, many studies rely on indirect evidence, such as correlations between pollinator abundance and seed set or the effects of hand-pollination treatments on seed set (Box 4-2).
In the most extreme examples of pollinator limitation, seed set fails because a plant’s primary pollinators are extinct or in precipitous decline. The best known cases in the United States involve endemic Hawaiian plant species that depend on pollinating birds or moths that are now extinct on one or more islands (Nabhan and Buchmann, 1996); some of them are maintained only by hand pollination. Two species of native lobeliads, Brighamia insignis and B. rockii (Shepherd et al., 2003), are examples whose principal sphingid moth pollinator is believed to be extinct (Kearns et al., 1998). In other cases, broadly applied insecticides have killed some native pollinators, resulting in low seed production in two species of wild plant (Thomson et al., 1985).
A more subtle form of pollinator limitation occurs when a plant species loses its pollinators to competition with other plants that have overlapping flowering seasons and share the same habitat (Kephart, 2005). Pollinators often prefer one plant species over another because of differences in floral
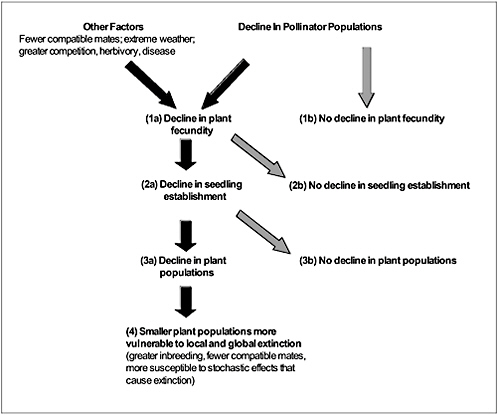
FIGURE 4-4 Possible effects of declining pollinator populations and other factors on the abundance and persistence of plant species. Effects represented by black arrows are cause for concern because they could lead to local or global extinctions.
rewards that can be collected in a given foraging period (Bernhardt, 1984). Plant fecundity is reduced when pollinators forage elsewhere and do not deliver enough conspecific compatible pollen to recipient plants (Campbell and Motten, 1985; Waser, 1978). Thus, competition for pollinators can depress plant fecundity—even when pollinators are locally common—because of the effects of plant community composition on pollinator behavior. In some plant species, no floral rewards are provided to pollinators and the flowers are thought to mimic rewarding flowers of other species or females of pollinating insects, whose males attempt copulation (Proctor et al., 1996) (Chapter 2). Such “deceptive” species could be especially prone to pollinator limitation because some pollinator species could learn to avoid them. For example, orchid species with flowers that lack food or chemical rewards for pollinators often exhibit very low fruit set, a condition commonly attributed to pollinator limitation (van der Cingel, 2001).
|
BOX 4-2 Pollen Supplementation Experiments Seed production is influenced by the supply of resources available to the plant for producing reproductive structures as well as by the availability of compatible pollen for siring seeds. A common way to test for pollen-limited fecundity (seed set) is to carry out pollen supplementation experiments (Ashman et al., 2004). The flowers of one set of plants are hand-pollinated with ample amounts of compatible pollen, and their seed set is compared with that of flowers on naturally pollinated control plants (Kearns and Inouye, 1993; Thompson, 2001). Thorough hand-pollination of all of a plant’s flowers throughout its lifetime can aid in differentiating between resource limitation and pollen limitation (Zimmerman and Pyke, 1988) in that resource limitation might prevent plants from producing fruits in successive years even when there is no shortage of pollination. Hand-pollination is impractical, however, in long-lived or many-flowered species like shrubs and trees. If only a fraction of the plant’s flowers are hand-pollinated, a separate set of control flowers should be used to determine whether artificially high seed set from hand-pollinated flowers causes artificially low seed set on the remaining, naturally pollinated flowers. Such an outcome could result because resources that are used early in the growing season are not available for later fruit set (Knight et al., 2005a; Stephenson, 1981). Similarly, to avoid overestimating the extent of pollen limitation, investigators should determine whether artificially high fecundity in one year is followed by lower fecundity or reduced vegetative growth in the next year, as occurred in moth-pollinated cranefly orchids (Tipularia discolor) in Maryland (Snow and Whigham, 1989). Pollen supplementation experiments should be carried out in several populations, including small, isolated populations, to account for temporal and spatial variation in pollinator service. Some studies probably over- or underestimate the extent of pollen limitation, but meta-analyses of pollen supplementation experiments can correct for many of these shortcomings (Knight et al., 2006). Careful experimentation can reveal the relative contributions of resource limitation and pollen limitation within a single species (Asikainen and Mutikainen, 2005; Campbell and Halama, 1993). |
Because several mechanisms can account for low fecundity (Figure 4-4), it is useful to distinguish between two types of pollen-related constraints on seed production: pollinator limitation (insufficient pollinator service) and pollen limitation (insufficient delivery of compatible pollen to receptive stigmas; see Thompson, 2001). Low numbers of pollinators often cause pollen
limitation of seed production, but not all examples of pollen limitation result from low numbers of pollinators. For example, pollen limitation can occur in self-incompatible species that are highly clonal (Duncan et al., 2004), in populations without compatible mating types (Demauro, 1993), or during mass flowering (Vance et al., 2004) even when pollinators are common because too little outcross pollen is delivered to receptive stigmas. Methods for quantifying pollen limitation of seed set are described in Box 4-2. Plant populations that experience chronic and increasingly severe symptoms of pollen limitation are prime candidates for further study to determine whether low fecundity can be attributed to declining pollinator populations.
Several recent reviews indicate that pollen-limited fecundity is widespread in natural populations and can be severe (Burd, 1994; Knight et al., 2006; Larson and Barrett, 2000). Burd (1994) reported that 62 percent of 258 species exhibited pollen limitation at some sites and times, and Ashman and colleagues (2004) found evidence for pollen limitation in 62 of 85 cases in which hand-pollination was carried out on whole plants. In agreement with previous surveys, Knight and colleagues (2005a) reported that pollen limitation was more common in self-incompatible species than in self-compatible ones, and plants with fewer pollinator species were more likely to be pollen-limited than were those with many. Vamosi and colleagues (2006) reported a strong positive correlation between plant species richness and pollen limitation on a global scale, but there was no clear explanation for this association. The investigators hypothesized that competition for pollinators is more prevalent in species-rich communities, but they were unable to determine whether competition for pollinators is a natural and long-standing phenomenon or whether lower fecundity is a consequence of recent declines in pollinator populations.
In general, little is known about whether pollen limitation of seed set in wild plants has become more chronic or more severe on a large scale in recent decades. In any given species, the ratio of flowering plants to foraging pollinators can vary greatly during the flowering season, among locations, and over time (Fenster and Dudash, 2001), so it is difficult to identify all but the most drastic pollinator shortages. Despite that, well-planned pollen supplementation experiments can provide useful tests for pollinator service. They also are more manageable than tracking pollinator population trends directly or attempting to correlate variations in pollinator abundance with plant fecundity. Pollen supplementation experiments provide information about the immediate consequences of pollinator shortages, especially when studies include flower-bagging treatments that completely exclude pollinators. Conservation biologists use hand-pollination to determine whether insufficient pollination might contribute to the decline of threatened and endangered species, although that often is not the case (Bigger, 1999; Johnson et al., 2004; Rasmussen and Kollmann, 2004). Long-term monitoring of
pollen-limited fecundity (Chapter 5) is a prerequisite for securing compelling evidence of the functional consequences of pollinator declines.
Direct and Indirect Effects of Pollinator Shifts
Specialization and Redundancy in Plant-Pollinator Interactions
Pollination is typically a mutualistic relationship, and disrupting it can have immediate and significant consequences—at least for the partners directly involved—although the consequences might not be easily or immediately discerned. Janzen (1974) and Kevan (1974) suggested that loss of mutualisms could be an insidious process that might go unnoticed but that eventually would have major consequences for the integrity of an ecosystem. The complex nature of many plant-pollinator relationships, which can involve many links among species, could mean that the loss of component species would eventually elicit a sequence of plant or pollinator extinctions (Memmot et al., 2004; Olesen and Jain, 1994; Rathcke and Jules, 1993). Few studies have investigated how the breakdown of mutualistic pollination relationships might contribute to the decline of endangered plants (Robertson et al., 1999), but there are examples of the extinction of the primary pollinators of some species (Cox and Elmqvist, 2000; Ladley and Kelly, 1995; Robertson et al., 1999; Weller, 1994). Memmot et al. (2004) used records of flowers and pollinators to simulate the consequences of progressive pollinator extinction by removing pollinator species and observing which plants were left without pollinators as a result. In the simulations, the proportion of plants visited by pollinators declined differentially depending on whether pollinators are removed at random, systematically from least specialized (generalist) to most specialized, or systematically from most specialized to least specialized. The percentage of plants visited decreases more or less linearly if generalist pollinators are removed first. If the pollinators are removed in random order or in the order from least to most specialized, the percentage of plants visited decreases in a steadily accelerating pattern. These results suggest that loss of generalist pollinators pose the biggest threat to pollination systems (Memmott et al., 2004).
Pollen limitation of fruit or seed set can serve as a useful indication of pollinator decline. However, although many researchers have demonstrated apparent pollen limitation, few have investigated its effects on the demographics of plant species (but see Johnson et al., 2004; Ward and Johnson, 2005). The decline or loss of specialist pollinators would appear to be an obvious case in which to find consequences for conservation. Steiner and Whitehead (1996) identified a case in which the floral form of a South African shrub (Ixianthes retzioides) suggested that a large oil-collecting bee was the coevolved pollinator, although no such bee could be found in the popula-
tions examined. The minimal seed set that occurs in the populations appears to be the result of visits by pollen-collecting bees. There are other cases in which the loss of a coevolved pollinator appears to have been ameliorated by the arrival of introduced species that visit and pollinate the flowers. For example, in Hawaii the Japanese white-eye Zosterops japonicus has taken over the role of extinct or rare native birds in pollinating at least two plant species (Cox, 1983; Lammers et al., 1987). Plants with long, curved corollas, which might have depended on pollinators with matching mouthparts, have had unusually high extinction rates on Pacific islands; these losses have been accompanied (or preceded) by extinctions of their pollinating birds (Pimm et al., 1994).
Although few North American plant species rely on a small number of pollinator species for seed production, many plants depend heavily on specific floral visitors, such as bumble bees, for effective pollination. Plants have evolved suites of floral traits that attract and reward pollinators and pollinators can be classified into functional groups based on the types of flowers they pollinate (reviewed by Fenster et al., 2004).
To explore the extent of specialization for pollinators, Fenster and colleagues (2004) examined visitation data for 278 animal-pollinated plant species in Illinois that had been studied almost 80 years earlier by Robertson (1929). Recognizing that not all floral visitors are effective as pollinators, Fenster’s group reported that 75 percent of the plant species were visited mainly by species in only one or two of nine functional groups (long-tongued bees, short-tongued bees, other Hymenoptera, Diptera, Coleoptera, Lepidoptera, Hemiptera, Neuroptera, and birds). In contrast, Waser and colleagues (1996) examined Robertson’s (1929) records and noted that 91 percent of 375 native plant taxa reported in that study had been visited by more than one pollinator species. These reviews suggest that flowers of many plant species are visited by generalist pollinators as well as by pollinators in particular functional groups (Tudor et al., 2004).
If pollinator declines occur differentially in different taxonomic groups, rather than across all categories of major pollinators equally and simultaneously, the consequences for plant reproduction depend on the degree of redundancy in pollinator communities and on changes in the composition of pollinator communities. Redundancy has been described for a variety of plant-pollinator relationships; angiosperms in general could thus be buffered against losses of individual pollinator species (Waser and Ollerton, 2006). Most pollinators visit a variety of plants, and many plants are attractive to a range of pollinators (Memmott, 1999; Memmott et al., 2004). Because most pollinators are not completely constant to particular flowers, plant-pollinator interactions represent a complex web. Redundancy is one characteristic of community-level analysis of plant-pollinator relationships in general (Memmott, 1999).
The importance of redundancy among co-occurring pollinators is apparent in the variation in floral availability among locations, over time (for example, Saavedra et al., 2003), and among pollinators. The redundancy of services is a safety net for the long-term survival of plants and pollinators alike. Sharing of pollinators also can facilitate interactions among plants (Moeller, 2004). An example of the dynamic nature of pollinator communities involves the creosote bush, Larrea tridentata, a plant visited and pollinated by many specialist bees in the southwestern United States and Mexico. There are few long-term studies of bee abundance and diversity, but one investigation in undisturbed sites revealed that only a minority of abundant bee species was typically persistent, even after 20 years (Cane et al., 2006). Most of the bee species in the Larrea pollinator guild were uncommon, geographically sporadic, and temporally unpredictable (Cane et al., 2006). Williams and colleagues (2001) surveyed published studies and reported that bee faunas are typically locally diverse, highly variable in space and time, and often rich in rare species. These attributes of pollinator guilds indicate that intense long-term sampling among sites is necessary to differentiate the changes that are attributable to specific impacts of pollinator declines from the natural dynamics of populations and communities. These attributes also suggest that a metapopulation model that accounts for local disappearance and reintroduction of insect populations may be typical of many pollinator species. Because plant-pollinator interactions are highly connected and plant and pollinator populations can shift in time and space, conserving mutualisms might require a better understanding of their population dynamics and interactions.
Pollinator Declines, Plant Demography, and Genetics
The degree to which declining pollinator abundance affects plant population dynamics depends on several factors (Harris and Johnson, 2004), as illustrated in Figure 4-4. First, a decline in pollinators will not necessarily result in lower seed production (step 1, Figure 4-4). The prevalence of pollen-limited fecundity (Knight et al., 2006) suggests that low seed set or fruit production could be exacerbated by declining pollinator populations, but that might not be the case if alternative pollinators are present or if the plant is autogamous (self-pollinating) when pollinators are absent. Even when seed set remains high, though, increased rates of selfing could lead to poor seed quality because of inbreeding depression (Charlesworth and Charlesworth, 1987). In species that typically outcross, the survival and reproduction of selfed progeny are often lower than they are for outcrossed progeny (Dudash, 1990). Plants with mixed mating systems could be forced to increase reliance on selfing instead of outcrossing for seed production (Goodwillie, 2000).
The second condition that must be met for population-level responses to pollinator declines is for lower seed production to translate into lower seedling establishment (step 2, Figure 4-4). If seedling establishment is limited by some factor other than seed production, such as dispersal, seed predation, germination microsites, or high seedling mortality, then reduced fecundity might not affect seedling recruitment. Few studies have demonstrated that plant population size is seed-limited (Brys et al., 2004; Juenger and Bergelson, 2000). Empirical study of seed limitation is challenging. Vast numbers of seeds never reach a suitable site for seedling establishment, which is often patchy and episodic, and the effects of small but chronic decreases in plant fecundity on seedling populations could be difficult to measure in the field. Therefore, plant population ecologists often rely on population modeling based on empirically derived transition probabilities to predict the effects of reduced or enhanced fecundity on seedling establishment (Miriti et al., 2001).
The third condition that must be met for population-level responses to pollinator declines is for decreased seedling establishment to cause decreases in the number of mature plants. As with seedlings, the survival of young plants to sexual maturity can be strongly limited by competition, herbivory, extreme weather, human activity, and numerous other factors. Population modeling is useful for determining how this stage of the plant’s life history affects the number of mature plants. Drastic reductions in recruitment could be detectable in natural populations, and small, undetectable effects of declining fecundity could be insignificant or important. If the plants are long-lived perennials, it might be many years for the loss of pollinator services to become apparent in the population structure. Many wild plant populations already are declining because of habitat loss, competition with invasive species, overgrazing by livestock or deer, and other factors (Ellstrand and Elam, 1993); more information is needed to ascertain whether declining pollinators could induce or exacerbate the loss of plant populations. Such a decline in wild plant populations because of pollination deficits might be in progress, but there is a dearth of published evidence for its occurrence in North America during recent decades.
If all the consequences of declining pollinator populations in fact occur (which seems unlikely), plant populations could become more vulnerable to an extinction vortex. The extinction vortex refers to the interacting factors that serve to reduce small populations progressively because of the increased risk of demographic and genetic consequences of small population size, including genetic erosion, inbreeding depression, decreased reproductive success, and greater susceptibility to catastrophes and random variations in environmental parameters (Kearns et al., 1998). One demographic risk is the Allee effect (inverse density dependence at low densities), which has
been demonstrated experimentally in plant populations (Groom, 1998; Hackney and McGraw, 2001) and observed in nature (Lamont et al., 1993). If there are no compensatory mechanisms for pollinator loss, and if reduced seed production from pollinator loss is directly reflected in smaller plant populations, these populations could enter an extinction vortex (Bond, 1994). Only careful research can disclose the necessary information about breeding systems, specificity and redundancy of pollinators, and plant life histories that will allow accurate prediction of the consequences of pollinator declines or losses.
At the other end of the spectrum, pollinator populations could increase and have a positive effect on plant abundance. One such case involves banyan trees, introduced ornamental fig species (especially Ficus benghalensis, F. altissima, and F. microcarpa) that became invasive pests in southern Florida subsequent to the inadvertent introduction about 25 years ago of the nonnative wasps that pollinate them (Nadel et al., 1992).
Indirect Effects of Pollinator Declines
There is a possibility that a cascade of ecological consequences could follow from the loss (or change in abundance) of roots, stems, leaves, flowers, fruits, and seeds—all of which can be resources for herbivores (including seed predators)—produced by plants. A broad range of herbivores and frugivores is supported by such resources, as are parasites and parasitoids of those species. Decreases in seeds, nuts, and fruits could be damaging to many species of insects, birds, and mammals, even if plant populations do not exhibit declines. More severe effects are expected if populations of mature plants become scarcer. For example, trophic cascades and the loss of shelter and nesting sites associated with plant species decline could cause reductions in bird populations. An example of how pollination itself can be part of a trophic cascade is provided by a study of the effects of fish as predators of dragonfly larvae in Florida (Knight et al., 2005b). Adult dragonflies are predators of some pollinators, so plants near ponds with dragonfly-eating fish receive more pollinator visits and are less pollen-limited than are those near fish-free ponds. This example suggests that pollinators are an important food resource for insectivorous predators.
Economic and Financial Consequences
Major shifts in natural pollinator populations could have economic repercussions either directly—in their appeal to nature lovers and collectors—or indirectly in their effects on the plants they pollinate. Some pollinators are appreciated aesthetically; butterflies and hummingbirds are
prime examples. That the honey bee is the state insect of 17 states and that various butterflies are state insects in 23 additional states (http://www.netstate.com/states/tables/state_insects.htm) is an indication of the visibility of pollinating species and the value placed on them. Apart from their intrinsic value, pollinators generate economic benefit through tourism. In 2000–2001, about 250,000 tourists visited the monarch butterfly reserve in west-central Mexico (Barkin, 2003). Colorado, Florida, Costa Rica, and Brazil host butterfly and bird tours (http://wingsbirds.com/birdsand.htm), although no estimates are readily available on total tourist expenditures at these sites.
Apart from ecotourism, at least one study shows evidence of park visitors’ expressed willingness to pay to protect endangered butterfly species (of Boloria aquilonaris in Germany; Degenhardt and Gronemann, 1998). The existence of various pollinator conservation groups also provides evidence of people’s willingness to pay to ensure the survival of endangered butterflies and birds. Examples include the British and Dutch Butterfly Conservation organizations (http://www.butterfly-conservation.org/ and http://www.vlinderstichting.nl/), the American Butterfly Conservation Initiative (http://www.butterflyrecovery.org/), the Xerces Society (www.xerces.org), and the Hummingbird Society (http://www.hummingbirdsociety.org/).
The loss of major groups of pollinators in natural areas could trigger changes in abundance or community composition, or even the loss of plant species that depend on pollinators for sexual propagation. The economic value of such losses stems from two sources. The first is the combination of existence, bequest, and cultural values that some people would be willing to pay to ensure species survival, enjoyment, or use by future generations. Also important are the aesthetic benefits or cultural satisfaction of watching pollinators at work. This broad category of value applies not only to the pollinator species, but also to plant species that depend on them for reproduction and to animal species that depend on associated fruits and seeds for food. No studies appear to have estimated willingness to pay to prevent such wild species losses that would attend pollinator decline.
The second source of economic value is the value of losing the potential opportunity to generate future income, for example by commercial pharmaceutical discoveries that could have come from a species that will be lost. The “bioprospecting” literature has attempted to estimate the value of conserving biodiversity hotspots. Published results ascribe low values to land conservation, except in areas with very high biodiversity, because the probability of finding commercially exploitable species is low (Rausser and Small, 2000; Simpson et al., 1996). Theoretical valuations from models of complex species interaction offer less definitive results (Simpson, 2002) but give no guidance for small-scale biodiversity loss attributable to pollinator shifts. Extrapolating from these findings, the likely value of the small
number of plant species that might be lost as a result of reproductive failure that is caused by a loss of pollinators is likely to be small. Finally, some economically important wild species (for example, ginseng) that suffer from pollinator limitation also are severely threatened by other stressors, such as overharvesting and deer grazing (McGraw and Furedi, 2005). In such cases, species demise is only partially attributable to pollinator limitation.
CONCLUSIONS
Severe shortages of pollinators could cause many common food crops to become more expensive and perhaps less available, but there is no strong evidence for a current pollination crisis in agricultural production in North America. Most animal-pollinated crops can be serviced by honey bees, and farmers are accustomed to paying more for these services when necessary. Chronic pollinator shortages should lead to market adjustments and other innovations, although the demand for supplemental pollination has been strong recently, especially among California’s almond growers. Importing managed pollinators from other countries or regions can lead to the introduction and spread of pathogens and unwanted bee species (Chapter 3). Therefore, it is prudent to sustain wild-pollinator populations as a means of buffering current and future shortages of managed pollinators of agricultural crops. In some cases, farmers could find that promoting wild pollinators that can supplement or substitute for managed pollinators could provide greater yields than will complete dependence on rented honey bee colonies (for example, Greenleaf, 2005). Intensive agricultural practices and pesticide use can reduce wild-pollinator diversity and abundance (for example, Kevan, 1975b; Kremen et al., 2002b; Larsen et al., 2005). Strategies for promoting abundance of wild pollinators are presented in Chapter 6.
In natural areas, the consequences of pollinator shifts on ecosystem services and possible solutions to these problems are varied and complex. Relatively few plant species rely on a single pollinator species or even on a single category of pollinators for reproduction. Because of the redundancy of most plant-pollinator communities, the ecological consequences of losing a small number of pollinator species could be limited. However, the loss of pollinator species reduces the redundancy of pollinator services in the ecosystem, and thus its resiliency, so that further losses of pollinator species would likely have more severe consequences for the ecosystem.
Plant communities in which massive pollinator declines are reported, for example because of persistent pesticide use or extensive habitat degradation, merit careful study of ramifying consequences for biodiversity. The ecological consequences of widespread pollinator declines could be substantial, but they could be difficult to detect because seed production varies dramatically in response to many biotic and abiotic factors (Figure 4-4). Moreover, the
effects of sustained declines in seed production on the size and abundance of plant populations could be subtle, slow processes that are real and overlooked by short-term studies. There is little available evidence to suggest that recent pollinator declines in North America have resulted in smaller populations of wild plants, possibly because the challenges of obtaining evidence are formidable. More research is needed to assess ecological consequences of current and future pollinator declines.



























