
8
The Terminal Cretaceous Extinction Event and Climatic Stability
HANS R.THIERSTEIN
Scripps Institution of Oceanography
WHY THE LATEST CRETACEOUS?
The most outstanding feature of late Phanerozoic climatic evolution is the gradual cooling of the world’s high-latitude areas. This cooling trend started about 60 million years ago (Ma) and has been particularly well documented in changes of the oxygen isotopic composition of fossil carbonate shells (Savin, 1977). The time just prior to the onset of Cenozoic cooling may therefore be considered as the climatic state farthest removed from the present or Pleistocene one and thus may represent the youngest non-Pleistocene climatic end-member. A second major source of interest in the late Cretaceous derives from the fact that it preceded one of the most severe evolutionary breaks recorded in the Phanerozoic history of life (Russell, 1979). Because climate today is the most important single factor determining the abundance and distribution of living organisms, paleontologists have long been including climatic feedback mechanisms in their proposals on possible causes for the terminal Cretaceous mass-extinctions.
THE CRETACEOUS-TERTIARY BOUNDARY EVENT
Until quite recently, the terminal Cretaceous extinctions were a subject that was discussed almost exclusively by paleontologists. The main topic of interest was the disappearance of several major groups of organisms that had dominated Mesozoic fossil assemblages, such as the dinosaurs, most of the cephalopods, the rudists, and the inoceramids (e.g., Schindewolf, 1954), as well as the marked reduction in the diversity observed in virtually all fossil communities between the Cretaceous and the Paleocene (e.g., Newell, 1962). The discovery that a rapid evolutionary turnover was also recorded in oceanic plankton communities (Luterbacher and Premoli Silva, 1964; Bramlette, 1965) appeared to confirm the postulated global extent of the extinction event and led to a rapid diversification of ideas on possible causes, which included geochemical as well as climatic feedback mechanisms (Bramlette, 1965; Tappan, 1968; Worsley, 1971; Gartner and Keany,
1978; Thierstein and Berger, 1978; Gartner and McGuirk, 1979; Alvarez et al., 1980).
Which processes are involved in these models? How compatible are the proposals with the available evidence? What are some of the crucial gaps in our knowledge?
EVOLUTION AND CHRONOLOGY
There are several questions of major concern: Are the massextinctions real? Are they synchronous? How rapidly did they occur? Were they selective?
The reality and suddenness of the mass-extinction has been established beyond doubt for the planktonic foraminifera and for the calcareous phytoplankton by high-resolution stratigraphy (Figure 8.1) and by the verification of continuous sedimentation provided by paleomagnetic dating (e.g., Kent, 1977). A number of apparently complete sedimentary sections have been identified by these methods and have led to an estimation of the time of evolutionary turnover of less than a few thousand years (Thierstein, in press).
Deep-water benthic foraminifera show no evolutionary change across the Cretaceous-Tertiary boundary (Beckmann, 1960). However, it appears to be real wherever it could be well documented (e.g., Birkelund, 1979; Hakansson and Thomsen, 1979). Gradual evolutionary changes, mainly expressed as declining diversities through the latest Cretaceous, have been related to a gradual deterioration of unspecified nature in neritic environments (Kauffman, 1979), which could be related to local or global sea-level changes. Such changes may be real, but their relationship to the extinction event, if any, is obscure.
Claims of a gradual late Cretaceous evolutionary decline of the terrestrial large vertebrate communities in North America have been shown to be unsubstantiated when sample sizes are taken into account (Russell, 1979). Recent attempts to correlate the youngest fossilized dinosaur remains with the paleomagnetic stratigraphy have either been marred with controversies over local correlation (Butler et al., 1977; Alvarez and Vann, 1979) or were based on rocks with only marginally preserved remanent magnetization (Lerbekmo et al., 1979; Danis et al., in press). There is evidence for time transgressiveness of the dinosaur extinctions relative to the rapidly diversifying mammalian faunas, with some late reptilian survivors reported from the Paleocene of southern China (R.A.Sloan, U. of Miami, personal communication, 1979). Whether the evolution of mammals or the extinction of the dinosaurs are time transgressive and predate or postdate the plankton extinctions is obviously one of the more important issues, which will eventually have to be resolved by magnetic stratigraphy.
The severity of the change in the terrestrial vegetation is also controversial (Krassilov, 1978). Some of the best-documented data, however, appear to indicate at least a moderate change, representing climatic cooling and an increase in seasonality (Russell, 1979).
There is growing evidence that the extinctions were selective. A geologically instantaneous extinction of virtually the entire late Cretaceous oceanic calcareous phytoplankton has been well documented (Figure 8.1); this was followed by blooms of neritic taxa such as Braarudosphaera and the dino-
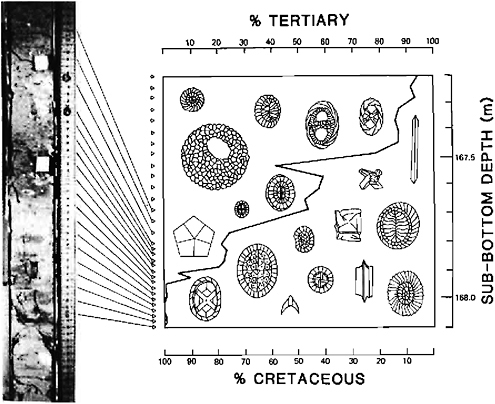
FIGURE 8.1 Replacement of the late Cretaceous by early Tertiary calcareous nannofossils within 70 cm of sediment thickness at DSDP Site 384, western North Atlantic (after Thierstein and Okada, 1979).
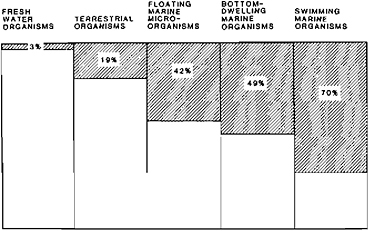
FIGURE 8.2 Terminal Cretaceous reduction in generic diversity of major fossil groups in different environments (after Russell, 1979).
flagellate cyst Thoracosphaera. The Maastrichtian-Paleocene reduction in the number of genera in different environments was clearly most severe in the oceanic realm and was less significant in terrestrial and particularly lacustrine environments (Figure 8.2). A reduction in generic diversity, however, must be considered a rather poor indicator of evolutionary change, as it may include taxonomic and sampling bias and possibly conceal a much larger evolutionary change at the specific level. For example, the ratio of Maastrichtian to Paleocene calcareous nannoplankton genera in the well-preserved sequence at Deep Sea Drilling Project (DSDP) Site 384 (Okada and Thierstein, 1979) is 30:24, a reduction of only 20 percent, although the assemblages are composed of entirely different species. More detailed data on the changes at the specific level for most groups of organisms are clearly required.
TECTONICS AND SEA LEVEL
There is widespread evidence for sea-level changes close to the Cretaceous-Tertiary boundary. However, little agreement appears to exist on their timing and direction in various parts of the world (Russell, 1979). Changing water depths certainly would exert a strong control on the local stratigraphic succes.sion of fossil assemblages in shallow-water environments (Kauffman, 1979). The rates of pre-Pleistocene eustatic sealevel changes, estimated by Pitman (1978) to be less than 1 cm per 1000 yr (excluding buildup and melting of continental ice) are considered too slow to be solely responsible for the quite sudden extinction events; such sea-level changes may have been instrumental, however, in triggering positive climatic feedback mechanisms or the isolation of ocean basins and their reconnection with the world oceans.
HISTORY OF CARBONATE DEPOSITION
Bramlette’s (1965) proposal of nutrient starvation in oceanic surface waters due to low erosion rates caused by tectonic quiescence was the first mechanism proposed that involved changes in the geochemical cycling of elements. Subsequently, Tappan (1968) pointed out that such a lowering of oceanic productivity may have resulted in additional environmental stress by significantly altering the oxygen-carbon dioxide balance in oceans and atmosphere. After the first four legs of the DSDP had failed to recover core representing a continuous Cretaceous-Tertiary transition, Worsley (1971) elaborated on these ideas and proposed that a late Cretaceous progressive shallowing of the carbonate compensation depth (CCD) into the photic zone might have affected the calcareous plankton directly. It has since been shown that carbonate deposition in at least some deep ocean basins was continuous across the Cretaceous-Tertiary boundary and that preservation of carbonate microfossils even improves between late Maastrichtian and earliest Danian (Thierstein, in press).
CLAY MINERALOGY
There are few data available on mineralogical changes in the carbonates and deep-sea clays across the Cretaceous-Tertiary boundary. Christensen et al. (1973) reported that increases in detrital quartz, silt, and trace-element concentrations in the boundary clay (Fiskeler) in Denmark were correlated with a decrease in carbonate contents, suggesting deposition of locally derived detritus in a starved, anoxic basin. Chamley and Robert (1979) in a study of DSDP sites in the Atlantic Ocean found that the stratigraphically and geographically variable supply of dominantly primary detrital clays through the late Cretaceous was suddenly terminated in the Paleocene. The trend, they concluded, was inconsistent with a latest Cretaceous cooling, which had been proposed based on oxygen isotopic data.
TRACE-ELEMENT GEOCHEMISTRY
The recent discovery of a strong increase in the concentration of iridium in the Cretaceous-Tertiary boundary clays (Alvarez et al., 1980; Ganapathy, 1980; Smit and Hertogen, 1980) lends strong support to Urey’s (1973) proposal of a meteorite impact as cause for the observed mass-extinctions. Noble element concentrations in differentiated crustal material are much lower than in undifferentiated extraterrestrial materials: measured iridium concentrations in Holocene deep-sea sediments average 0.3 part per billion (ppb); in manganese nodules, owing to low bulk accumulation rates, they average 8.5 ppb; and in iron meteorites they go up to 500 ppb (Crocket et al., 1973).
Alvarez et al. (1980) explained the observed increase in iridium in the noncarbonate fraction from an average of 0.3 ppb in the uppermost Cretaceous limestones at Gubbio (Italy) to a maximum of 8.4 ppb in the boundary clay as a consequence of an asteroid impact. With isotopic data they ruled out the possibility of a supernova or other galactic cause for the extinction event (Russell, 1979; Napier and Clube, 1979) and argued convincingly for a solar-system source of their impacting object. The mass-extinction in their scenario would be
caused by a thick veil of dust thrown up into the stratosphere by the impact, which then, through shielding of sunlight for a few years, would inhibit photosynthesis. There are several aspects of this proposal, however, that appear to require further analysis and scrutiny (see also Chapter 5). Light attenuation, for instance, would have to exceed 99 percent for months to prohibit survival of calcareous phytoplankton (Blankley, 1971). To achieve this attenuation, mass concentrations of about 500 µg m−3 over a stratospheric dust layer of 10-km thickness would have to be maintained (Cadle and Grams, 1975), assuming average grain-size distributions similar to those observed in the stratosphere during recent large volcanic explosions (e.g., Mossop, 1964). Additional uncertainties revolve around the possibility of noble-element enrichment processes within the global geochemical cycle, such as in the biosphere, in soils, or in hard grounds.
Excessive heat production during a meteorite impact and cyanide poisoning from a cometary collision have also been proposed recently (Emiliani, 1980; Hsü, 1980).
Future studies of other boundary sections, of the geochemical cycle of noble elements, and of rocks associated with known meteorite impacts may well yield conclusive evidence for the proposed model.
STABLE ISOTOPES
Some of the most important information known about climates in the late Mesozoic is derived from comparatively few studies on the oxygen isotopic composition of fossil carbonate shells. These studies were concentrated on an evaluation of major stratigraphic trends and paleolatitudinal gradients (Frakes, 1979). Recent results show that oxygen isotopic gradients preserved in late Cretaceous microfossils were very weak, both vertically and latitudinally, when compared with those of the Holocene (Table 8.1). The deepest benthic forami
TABLE 8.1 Oxygen Isotopic Temperature Gradients Preserved in Holocene and Latest Cretaceous Carbonate Microfossils
|
|
Holocene |
Latest Cretaceous |
|
Planktonic forams (vertical, 0–200 m) |
9°Ca |
5°Cb |
|
Planktonic/benthic forams (vertical, 0–2500 m) |
21°Cc |
5°Cd |
|
Nannofossils (0–55° paleolatitude) |
25°Ce |
13°Cb |
|
aFrom Berger et al. (1978). bFrom unpublished data by author. cFrom Vincent et al. (in press). dFrom Savin (1977). eFrom Goodney et al. (1980). |
||
nifera measurements available for the Cretaceous, however, are from a paleodepth of only 2.5 km. Truly abyssal waters, representative of polar winter temperatures, may well have been considerably colder than the 7 to 10°C measured at intermediate depths. The possibility of lowered temperature gradients suggests a less-vigorous deep-water circulation than observed today, which may well have led to an amplification of salinity gradients in surface waters. The present-day range of average annual salinity anomalies from the expected latitudinal values is 11 ‰ between the eastern Mediterranean, a significant deep-water contributor, and the Panama Basin, which is unaffected by river runoff (Dietrich et al., 1975). This difference is equivalent to a range in the oxygen isotopic composition of about 6.5 ‰ (Craig and Gordon, 1965). Because oceanic circulation in the Cretaceous was probably less intense than today’s, regional variation can be expected in the isotopic composition of oceanic surface waters, as evaporation and precipitation were at least as large as at present. In particular, the latitudinal surface salinity gradients may have been considerably larger than today’s. These gradients could have been maintained by increased moisture transport from low to high latitudes, which at the same time would have provided an efficient heat-transport mechanism. Recent sensitivity studies of global climate models suggest an increase in latitudinal moisture transport with increasing atmospheric CO2 content (Manabe and Stouffer, 1980). Higher atmospheric CO2 contents may have been caused by the lowered solubility of gases in warmer ocean waters (Plass, 1972).
The possibility of considerable latitudinal salinity fractionation of oceanic surface waters in the late Cretaceous is a basic premise of the brackish-water injection model (Gartner and Keany, 1978). Preliminary oxygen isotope data from the coccolith fraction at DSDP Site 356 (South Atlantic) appear to support a sudden freshening of oceanic surface waters at the Cretaceous-Tertiary boundary (Thierstein and Berger, 1978). The oxygen isotopic shift, however, could not be confirmed at other sites or in other fossil groups. In addition, the original biostratigraphic evidence used by Gartner and Keany (1978) has been reinterpreted as a submarine slump, further weakening the brackish-water injection hypothesis (Thierstein, in press).
Recent studies on the carbon isotopic variability of the dissolved carbon dioxide in ocean water as well as in Recent and Neogene carbonate shells indicate that carbon isotopes may be used to trace fertility patterns (Goodney et al., 1980) as well as major changes in the partitioning and cycling of carbon between the main carbon reservoirs. Changing flux rates between the reservoirs leave their traces in the carbonate skeletons of the transport vehicles. Such major changes have been documented in the middle Cretaceous (anoxic events), the Cretaceous-Tertiary boundary (collapse of the oceanic plankton community), and the late Miocene (beginning of Messinian salinity crisis) (Thierstein and Berger, 1978; Scholle and Arthur, 1980; Vincent et al., 1980).
A virtually unexplored field of great paleoclimatic potential is the study of seasonal variability preserved in the stable isotopic composition of carbonate shells of macroinvertebrates.
THE ENIGMA OF THE WARM AND STABLE CLIMATES IN THE LATE CRETACEOUS
The Cenozoic cooling trend in high latitudes was not gradual but proceeded in spurts and relapses as documented in the paleobotanical and oxygen isotopic evidence (Figure 8.3). The high-latitude cooling trend is generally thought to be caused by tectonic processes, such as the movements of continents into polar areas, global regression, and mountain building, interrupted by more local events, such as closure or opening of oceanic gateways, the isolation and reconnection of ocean basins, volcanism and the like, with amplification or attenuation by climatic feedback mechanisms, such as changes in albedo, in oceanic circulation, or in the carbon cycle. High-resolution stratigraphy may provide clues to these climatic feedback mechanisms and to the stability of the various climatic states that the Earth has experienced in glacials and interglacials, in the warm and stable late Cretaceous, and during the mid-Cretaceous period of widespread deep-water anoxia.
What were the conditions that made the late Cretaceous climates so stable? Was it climatic stability in itself, which, when disrupted, helped to produce the unprecedented biotic extinctions? The enigma of the properties of the late Cretaceous climatic system may be just as important as that of the disturbance that led to the extinctions. How different were late Cretaceous climates from Pleistocene ones?
From a late Phanerozoic viewpoint, today’s oceans are characterized by their small and highly dispersed areal coverage, by their pronounced temperature stratification into a thin, warm surface layer and a large cold deep layer, and by vigorous meridional surface circulation, leading to a strong geographical separation of fertility. The deep-water reservoir
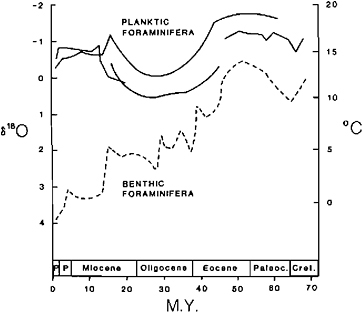
FIGURE 8.3 Isotopic paleotemperature data for latest Cretaceous and Cenozoic planktonic and benthic foraminifera, illustrating the post-Cretaceous irregular cooling trend in high latitudes (after Savin, 1977).
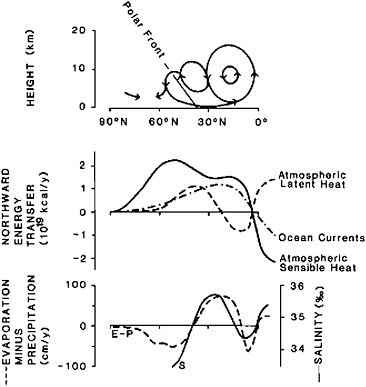
FIGURE 8.4 Schematic representation of vertical, meridional atmospheric circulation, annual latitudinal heat flux, evaporation/precipitation pattern, and surface salinities for the northern hemisphere (after Frakes, 1979; Sellers, 1965; Dietrich et al., 1975).
is mainly replenished in the cold polar areas, which are characterized by high surface albedo. Net oceanic sensible heat transport is limited to low latitude surface waters (Figure 8.4). A major portion of the poleward heat transport occurs as sensible heat in the atmosphere through the upper part of the subtropical Hadley cells and across the polar front. The latent heat transport (heat stored during evaporation and released during precipitation) is closely tied to the atmospheric circulation patterns and is strongly mirrored in the average latitudinal surface salinities in tropical and subtropical areas (Figure 8.4).
In the late Cretaceous, on the other hand, the overall climatic circulation patterns must have been considerably different for the following reasons. Lower surface albedo, on a global scale, because of higher sea level and the absence of ice in high latitudes, must have resulted in a more efficient global heat absorption. If there was indeed no significant temperature decrease below 2500-m water depth or in polar latitudes, the density of oceanic waters would have been determined to a much larger degree by salinity, rather than temperature, suggesting the possibility of a deep-water circulation reversal and thus an increase of the oceanic sensible heat transport into high latitudes. High-latitude upwelling of saline, but relatively warm water, should have been countered efficiently, however, by negative feedback through latent and sensible heat loss and increase in salinity. Could the resulting density increase have been balanced by dilution with precipitation and runoff surplus? A significantly longer residence time in the deep-water reservoir compared to today appears unlikely, as
there is no evidence of latest Cretaceous deep-water anoxia or greatly lowered fertility, as judged from calcium carbonate sedimentation rates and sedimentology. Did high-latitude upwelling occur locally or seasonally? Increased seasonality in high latitudes should have derived from higher heat retention due to decreased albedo (no ice), particularly during summers, when the total daily radiation is higher on the poles than anywhere else. Could evidence for seasonality be derived from the annual variability in the stable isotopic composition of highlatitude macroinvertebrate shells and be used to test Wolfe’s (1978) claim for a significantly lowered inclination of the Earth’s rotational axis?
A number of open questions concerning late Cretaceous climates can be answered by careful study of available sediments and fossils and by using available techniques, whereas others will have to be addressed by way of analogy and inference from the longer-range Phanerozoic climatic history of the Earth.
REFERENCES
Alvarez, W., and D.W.Vann (1979). Comments and replies on “Biostratigraphy and magnetostratigraphy of Paleocene terrestrial deposits, San Juan Basin, New Mexico,” Geology 7, 66–71.
Alvarez, L.W., W.Alvarez, F.Asaro, and H.V.Michel (1980). Extraterrestrial cause for the Cretaceous-Tertiary extinctions: Experiment and theory, Science 208, 1095–1108.
Beckmann, J.P. (1960). Distribution of benthonic foraminifera at the Cretaceous-Tertiary boundary of Trinidad (West Indies), Report 21st Geol. Congr., Norden, part V, 57–69.
Berger, W.H., J.S.Killingley, and E.Vincent (1978). Stable isotopes in deep-sea carbonates: Box core ERDC-92, west equatorial Pacific, Oceanol. Acta 1, 203–216.
Birkelund, T. (1979). The last Maastrichtian ammonites, in Cretaceous/Tertiary Boundary Events Symposium. I. The Maastrichtian and Danian of Denmark, T.Birkelund and R.G.Bromley, eds., U. of Copenhagen, Copenhagen, Denmark, pp. 51–57.
Blankley, W.F. (1971). Auxotrophic and heterotrophic growth and calcification in coccolithophorids, Ph.D. Thesis, U. of California, San Diego, 186 pp.
Bramlette, M.N. (1965). Massive extinctions in biota at the end of Mesozoic time, Science 148, 1696–1699.
Butler, R.F., E.J.Lindsay, L.L.Jacobs, and N.M.Johnson (1977). Magnetostratigraphy of the Cretaceous-Tertiary boundary in the San Juan Basin, New Mexico, Nature 267, 318–323.
Cadle, R.D., and G.W.Grams (1975). Stratospheric aerosol particles and their optical properties, Rev. Geophys. Space Phys. 13, 475–501.
Chamely, H., and C.Robert (1979). Late Cretaceous to early Paleogene environmental evolution expressed by the Atlantic clay sedimentation, in Cretaceous/Tertiary Boundary Events Symposium. II. Proceedings, W.K.Christensen and T.Birkelund, eds., U. of Copenhagen, Copenhagen, Denmark, pp. 71–77.
Christensen, L., S.Fregerslev, A.Simonsen, and J.Thiede (1973). Sedimentology and depositional environment of lower Danian fish clay from Stevns Klint, Denmark, Bull. Geol. Soc. Denmark 22, 193–212.
Craig, H., and L.I.Gordon (1965). Deuterium and oxygen-18 variations in the ocean and the marine atmosphere, in Stable Isotopes in Oceanographic Studies and Paleotemperatures, Spoleto, July 26–30, 1965, Cons. Naz. Richerche, Lab. Geol. Nucleare, Pisa, pp. 9–130.
Crocket, J.H., J.D.MacDougall, and R.C.Harriss (1973). Gold, palladium and iridium in marine sediments, Geochim. Cosmochim. Acta 37, 2547–2556.
Danis, G.P.L, J.A.Foster, and D.A.Russell (in press). The paleomagnetic record and the Cretaceous-Tertiary boundary in North America, Syllogeus.
Dietrich, G., K.Kalle, W.Krans, and G.Siedler (1975). Allgemeine Meereskunde, Gebr. Borntraeger, Berlin, 593 pp.
Emiliani, C. (1980). Death and renovation at the end of the Mesozoic, EOS: Trans. Am. Geophys. Union 61, 505–506.
Frakes, L.A. (1979). Climates Throughout Geologic Time, Elsevier, Amsterdam, 310 pp.
Ganapathy, R. (1980). A major meteorite impact on the earth 65 million years ago: Evidence from the Cretaceous-Tertiary boundary clay, Science 209, 921–923.
Gartner, S., and J.Keany (1978). The terminal Cretaceous event: A geologic problem with an oceanographic solution, Geology 6, 708–712.
Gartner, S., and J.P.McGuirk (1979). Terminal Cretaceous extinction: Scenario for a catastrophe, Science 206, 1272–1276.
Goodney, D.E., S.V.Margolis, W.C.Dudley, P.Kroopnick, and D.F.Williams (1980). Oxygen and carbon isotopes of Recent calcareous nannofossils as paleooceanographic indicators, Mar. Micropaleontol. 5, 31–42.
Hakansson, E., and E.Thomsen (1979). Distribution and types of bryozoan communities at the boundary in Denmark, in Cretaceous/ Tertiary Boundary Events Symposium. I. The Maastrichtian and Danian of Denmark, T.Birkelund and R.G.Bromley, eds., U. of Copenhagen, Copenhagen, Denmark, pp. 78–91.
Hsü, K.J. (1980). Terrestrial catastrophe caused by cometary impact at the end of the Cretaceous, Nature 285, 201–203.
Kauffman, E.G. (1979). The ecology and biogeography of the Cretaceous-Tertiary extinction event, in Cretaceous/Tertiary Boundary Events Symposium. II. Proceedings, W.K.Christensen and T.Birkelund, eds., U. of Copenhagen, Copenhagen, Denmark, pp. 29–37.
Kent, D.V. (1977). An estimate of the duration of the faunal change at the Cretaceous-Tertiary boundary, Geology 5, 769–771.
Krassilov, V.A. (1978). Late Cretaceous gymnosperms from Sakhalin and the terminal Cretaceous event, Paleontology 21, 893–905.
Lerbekmo, J.F., M.E.Evans, and H.Baadsgaard (1979). Magnetostratigraphy, biostratigraphy and geochronology of Cretaceous-Tertiary boundary sediments, Red Deer Valley, Nature 279, 26–30.
Luterbacher, H.P., and I.Premoli Silva (1964). Biostratigrafia del limite Cretaceo-Terziario nell’ Appennino Centrale, Riv. Ital. Paleontol. LXX, 67–128.
Manabe, S., and R.J.Stouffer (1980). Sensitivity of a global climate model an increase of CO2 concentration in the atmosphere, J. Geophys. Res. 85, 5529–5554.
Mossop, S.C. (1964). Volcanic dust collected at an altitude of 20 km, Nature 203, 824–827.
Napier, W.M., and S.V.M.Clube (1979). A theory of terrestrial catastrophism, Nature 282, 455–459.
Newell, N. D. (1962). Paleontological gaps and geochronology, J. Paleontol. 36, 592–610.
Okada, H., and H.R.Thierstein (1979). Calcareous nannoplankton—Leg 43, Deep Sea Drilling Project, in Initial Reports Deep Sea Drilling Project 43, U.S. Government Printing Office, Washington, D.C., pp. 507–573.
Pitman, W.C., III (1978). Relationship between eustacy and stratigraphic sequences of passive margins, Geol. Soc. Am. Bull. 89, 1389–1403.
Plass, G.N. (1972). Relationship between atmospheric carbon dioxide amount and properties of the sea, Environ. Sci. Technol. 6, 736–740.
Russell, D.A. (1979). The enigma of the extinction of the dinosaurs, Ann. Rev. Earth Planet. Sci. 7, 163–182.
Savin, S.M. (1977). The history of the Earth’s surface temperature during the past 100 million years, Ann. Rev. Earth Planet. Sci. 5, 318–355.
Schindewolf, O.H. (1954). Über die möglichen Ursachen der grossen erdgeschichtlichen Faunenschnitte, Neues Jahrb. Geol. Palaeontol. Monatsh. 10, 457–565.
Scholle, P.A., and M.A.Arthur (1980). Carbon isotope fluctuations in Cretaceous pelagic limestones: Potential stratigraphic and petroleum exploration tool, Am. Assoc. Petrol. Geol. Bull. 64, 67–87.
Sellers, W.D. (1965). Physical Climatology. U. of Chicago Press, Chicago, 272 pp.
Smit, J., and J.Hertogen (1980). An extraterrestrial event at the Cretaceous-Tertiary boundary, Nature 285, 198–200.
Tappan, H. (1968). Primary production, isotope extinctions and the atmosphere, Palaeageogr. Palaeoclimatol. Palaeoecol. 4, 187–210.
Thierstein, H.R. (in press). Late Cretaceous calcereous nannoplank ton and the change at the Cretaceous-Tertiary boundary, in The Deep Sea Drilling Project: A Decade of Progress, R.G.Douglas and E.L.Winterer, eds., Soc. Econ. Paleontol. Mineral. Spec. Publ. 30.
Thierstein, H.R., and W.H.Berger (1978). Injection events in ocean history, Nature 276, 461–466.
Thierstein, H.R., and H.Okada (1979). The Cretaceous/Tertiary boundary event in the North Atlantic, in Initial Reports Deep Sea Drilling Project 43, U.S. Government Printing Office, Washington, D.C., pp. 601–616.
Urey, H.C. (1973). Cometary collisions and geological periods, Nature 242, 32–33,
Vincent, E., J.S.Killingley, and W.H.Berger (1980). The magnetic Epoch-6 carbon shift: a change in the oceans’ 13C/12C ratio 6.2 million years ago, Mar. Micropaleontol. 5, 185–203.
Vincent, E., J.S.Killingley, and W.H.Berger (in press). Stable isotope composition of benthic foraminifera from the equatorial Pacific, Nature.
Wolfe, J. (1978). A paleobotanical interpretation of Tertiary climates in the northern hemisphere, Am. Sci. 66, 694–703.
Worsley, T.R. (1971). Terminal Cretaceous events, Nature 230, 318–320.







