
20
Seasonality and the Structure of the Biosphere
JAMES W.VALENTINE
University of California, Santa Barbara
INTRODUCTION
This paper traces the major effects of seasonality on organisms, particularly in the sea, from both theoretical and observational data. The tilting of the Earth’s axis of rotation away from the axis of the ecliptic produces a regular annual pattern of variation in the solar radiation received on Earth, giving rise to seasons. The seasonality in solar radiation, greatest near the poles and least under the Sun’s track, entrains variation in a vast array of physical environmental parameters including temperature, rainfall, atmospheric and oceanic circulation, salinity regimes and nutrient upwelling in the sea, and humidity on land.
These parameters are highly important in the adaptation of organisms, and they affect in turn many other factors of great biological significance. The present pattern of seasonality is reflected in patterns in the biosphere. Patterns of seasonality must have been different in the past; the fossil record of patterns in ancient biospheres can be useful in understanding climatic change and its biological consequences.
POPULATION RESPONSES TO SEASONALITY
Intrinsic Population Features
The logistic equation of population growth is a convenient starting point from which to evaluate seasonal effects on populations. It is usually written
where N is population size, r is the intrinsic population growth rate, and K is the carrying capacity of the environment for that population. A population that is very small with respect to the carrying capacity will grow at a rate set by r until its density approaches K, when growth may be damped and the population size stabilized at an equilibrium value (Figure 20.1A). However, a wide range of other population behaviors is possible (May, 1975, 1979; May and Oster, 1976). If there is a time delay in response by a population to the operative factors in K (Hutchinson, 1948, 1965; May, 1973), the population becomes less likely to attain a stable equilibrium as the time lag in-
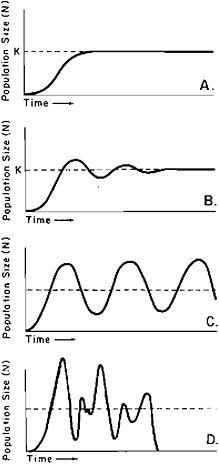
FIGURE 20.1 Diagrammatic representation of the dynamics displayed by populations of overlapping generations with a time delay in the regulatory mechanism. A, If time delay is sufficiently short a population may grow to a stable equilibrium level, K. B, With a larger time delay and sufficiently high r a population may at first penetrate the level of K, to which it may be returned via damped oscillations. C, With still larger time delay and sufficiently high r, a population may oscillate indefinitely in a limit cycle rather than seeking stability at an equilibrium level. D, As time delays increase, and if r is still sufficiently high, the limit cycle breaks down into random oscillations that can lead to extinction of the population. These concepts are reviewed by May (1979).
creases. Most time delays are on the order of a year and are related to seasonality. When r is sufficiently high and the factors in K are sufficiently slow to act, the population size can be driven above K temporarily. K may then be approached by damped oscillations that finally settle on the equilibrium value (Figure 20.1B). If the time lag is greater still, the population may penetrate K to such an extent that the subsequent crash carries it considerably below K; r may then return the population to its former peak level from which it will again crash, and the oscillations may continue indefinitely (rather than being damped) in a stable limit cycle (Figure 20.1C). Finally, if the time lag continues to increase, the stable cycle will break down into chaotic population density variations, the bounds of which increase so that at the lower bound the population size is below some critical low level from which it cannot recover; extinction ensues. Thus, intrinsic population properties even in a constant environmental regime can lead a population to range in behavior from stable to chaotic. Real populations must limit their downward fluctuations above some critical extinction point, and indeed to a point from which they may recover sufficiently to cope with subsequent inclement conditions.
Seasonal variation in environmental factors further complicates population dynamics. In general, the greater such intrinsic variation, the less the population, which can be driven through the same wide range of behavior as discussed above (see May, 1979). All real environments vary to some degree, evoking appropriate adaptive responses on the part of populations. Seasonality is the most regular pervasive source of environmental variation. It is logical to ask, what sorts of adaptations do populations evolve in order to cope with seasonality, and what are the consequences? Adaptations to seasonality are referred to here as seasonal strategies.
Extrinsic Density-Independent Factors
To endure in seasonal environments, populations must respond in ways that maintain their oscillations within some limited range. Two sorts of extrinsic factors are recognized (Smith, 1935). Density-independent factors affect organisms without regard to whether there are few or many individuals present. Most effects of physical climatic factors are of this sort. The lethality of a temperature change, for example, does not ordinarily depend on population size. Density-dependent factors, on the other hand, have increasing effects as population size increases. They form components of K in the logistic equation. Many biological and some physical factors are of this sort; commonly they involve a factor, such as food supply or habitat space, that can be used up. However, as emphasized by Andrewartha and Birch (1954), any factor that is density-independent can become density-dependent in some circumstances. It is better to speak of density-dependent and -independent processes or effects.
Density-independent processes play an important role in shaping the individual tolerances of organisms and therefore the modal niches of populations. This must be particularly true of seasonal factors because they impose annually recurrent effects, although with somewhat variable intensities. Selection arising from density-independent processes tends to act most severely near the margins of species’ distributions, where even small departures from normal conditions may exceed the tolerances of most individuals. It causes “stabilizing” or “centripetal” evolution. If the environmental regime changes, selection must adjust to tolerances and rates so as to be adaptive to the new conditions; this causes “directional” evolution. If the changes are too large or abrupt for evolution to track, extinction ensues.
Although they act independently of population size, it is possible for density-independent factors to control population densities at levels below those at which density-dependent effects occur (Andrewartha and Birch, 1954). For example, density-independent mortality may occur so frequently that populations never reach their carrying capacities. In this case it is postulated that selection favors a high-reproductive potential (r selection). When environments are relatively stable and populations at their carrying capacities, on the other hand, it is
postulated that selection favors adult efficiency in utilizing resources (K selection) (MacArthur and Wilson, 1967).
Extrinsic Density-Dependent Factors
The strategies that can be employed in response to seasonality have been investigated by Boyce (1979). He modeled the relative fitness of individuals with different combinations of three properties that seem particularly relevant to density-dependent selection: (1) intrinsic reproductive potential (r); (2) level of resource demand (D); and (3) population decay when resources are inadequate (z)—almost the opposite of (r). Boyce calculated the relative fitness of individuals with contrasts in these properties in different regimes of seasonality and at different population densities. The results are displayed in Figure 20.2.
In case A (Figure 20.2), individuals with higher r have the higher resource demand but resistance to inclement conditions is equal, compared with low-r individuals. The high-r individuals have the higher fitness over a range from low to high seasonality, except at very high population densities, when their high resource demand becomes a significant liability. Case D is similar in that the high-r individuals have the higher resource demand, but they have more resistance to inclement conditions than do low-r individuals. High-r individuals increase their margin of fitness in more highly seasonal conditions at low population densities. At very high densities, however, the high resource demand still poses a significant liability and individuals with the lower r become the more fit.
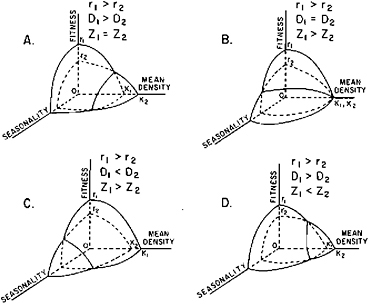
FIGURE 20.2 Models of the reproductive potential (r) and carrying capacity (K) of populations over a range of seasonalities and resource supplies (resource supplies are inversely proportional to mean densities). D is resource demand, Z is rate of population decrease. The population with the higher reproductive potential is always the fitter at low seasonalities, and it is also the fitter at high seasonalities if Z is equal to or lower than in the other population (cases A and D). However, the population with the greater resistance to decrease (smaller Z) is the fitter under highly seasonal conditions (after Boyce, 1979, copyright U. of Chicago Press).
Cases B and C have rather different outcomes. In both of these cases the high-r individuals have less resistance to inclement conditions than do low-r individuals. These high-r individuals remain most fit in the less seasonal environments but are least fit in highly seasonal conditions except at high densities. In case B both kinds of individuals have equal resource demands and equal fitness at the highest density. In case C the low-r individuals have the higher resource demands and therefore have less fitness at high densities.
It is evident from Figure 20.2 that adaptation is more difficult in environments with greater seasonality. Fitness always falls with increasing seasonality. Similarly, the carrying capacity is always lower in highly seasonal environments (as indicated by the curves of the fitness dimension of the seasonality-fitness faces in Figure 20.2). Similarly, the carrying capacity is always lower in highly seasonal environments (as indicated by the curves of the “mean density” dimension on the basal seasonality-mean density faces in Figure 20.2). The two main seasonal strategies are (1) to increase reproduction and (2) to fortify the populations against decrease during inclement conditions. The reproductive strategy has no special damping effect on population oscillations but prevents extinction by returning the population size rapidly toward K or beyond. It might result in rather large size fluctuations, but if successful, constrains them within limits (the lower significantly above zero) to give rise to a limit cycle. In the model these strategies (reproduction and fortification) seem about equally effective, although when greater resource demands are involved the fortification strategy is least affected.
Observation of species in highly seasonal environments reveals that these strategies are in operation in nature (Boyce, 1979). Mammals provide many terrestrial examples. Some have larger litter sizes in more seasonal (high-latitude) environments (Lord, 1960), following the strategies indicated by cases A or D. Other mammals in seasonal environments follow the fortification strategies. Some, such as hibernating forms, put energy into resource storage rather than into reproduction, following the strategy of case B; others grow to larger body sizes in seasonal regimes, buffering the effects of seasonality and thus following the strategy of case C.
Marine Populations and Seasonality
The fossil record of the marine shelf and shallow seas contains the most diverse fauna and embraces the greatest time span of all environmental realms. It is thus of interest to determine the seasonal strategies of shelf invertebrates. In the shelf realm there is a trend from high-reproductive potentials among species in regions with little seasonality to low-reproductive potentials in seasonal regions (Thorson, 1950; Mileikovsky, 1971)—just the opposite of what is expected of the reproductive strategies in cases A or D. Plausible explanations have been proposed for this trend in r. In less seasonal regions (such as the tropics) there is a relatively stable resource base in the water column—photosynthesis goes on at about the same intensity all year—and most shelf species have planktonic larvae that feed in the water column (planktotrophic larvae). Such species can produce many eggs, because nourishment need not be fur-
nished to the larvae. Planktotrophy is probably favored by selection because it aids dispersal and enhances recruitment in an environment densely populated and rich in rather specialized species and therefore patchy in opportunities. Invertebrates in stable shallow-water environments tend to be both r- and K-selected (Valentine and Ayala, 1978). In highly seasonal regimes (such as in high latitudes) most benthic invertebrates have a relatively small number of large yolky eggs; the young feed in the egg but hatch as nonfeeding larvae or as miniature adults. Each egg requires a considerable investment in energy and therefore fecundity is low, but clearly the chances of reproductive success per egg are higher than for planktotrophic forms, provided as they are with food and more or less protected during development, often by brooding. In terms of Boyce’s models, highly seasonal marine invertebrates follow case B or C strategies; the difference depends on whether their reproductive effort is equal (B) or greater (C) than invertebrates in less seasonal environments. It thus appears that marine invertebrates in seasonal environments are neither r- selected nor K-selected (Valentine and Ayala, 1978) but follow a strategy of developmental fortification.
Why this particular strategy has been selected, rather than an r strategy, is probably explained by the energetics of fluctuating populations. Imagine a population in a seasonal environment that is subject to such an inclement season as to suffer fairly heavy mortality—x percent. This must be made up via reproduction and (assuming deaths are random with respect to age) growth. In a more extreme (more highly seasonal) environment, mortality might be 2x percent. To make up for this heavier mortality, reproduction and growth must be correspondingly greater, and more energy must be consumed to support this greater effort. In each case the population has only until the next inclement season to return to a condition sufficient to cope with the accompanying wave of mortality. In general, then, under conditions of seasonally imposed mortality, the more a population fluctuates, the higher the cost in terms of energy use.
Because carrying capacities and fitnesses are low in highly seasonal environments (Figure 20.2), additional properties may be required of successful species. One strategy is to become ecologically generalized, occurring over a broad region in a wide variety of habitats and indulging in a catholic diet. In this way a species in a seasonal regime may maintain a large population but will cover the functional range of number of specialized species in stable environments. In a sense such adaptations represent a fortification strategy, since during inelement periods generalists are likely to be able to subsist on such food items as happen to be available and may occur in areas and in habitats that are least severly affected by the adverse conditions. It seems likely that a generalist strategy would maximize fitness under the circumstances.
DIVERSITY AND SEASONALITY
Diversity (species richness) patterns have attracted wide attention and controversy. They are often interpreted as climate related and if so would be useful in paleoclimatology for they represent one aspect of biotic structure that can be inferred from the fossil record. They are also of potential interest in predicting biotic response to future climatic change.
The strategies outlined above permit the survival of permanent communities in the “boom-and-bust” economies of highly seasonal environments, even with their low carrying capacities. At base, species persist by minimizing the effects of the seasonal fluctuations. The seasonal strategies, however, are resource-intensive. The available trophic resources must be apportioned in large shares. This places a limit on the number of species that can be accommodated in communities in seasonal regimes. In regions of low seasonality, resources are continuously available or nearly so, and carrying capacity is much higher per unit of resource. The resources may therefore be partitioned finely among numerous different species, which can have small populations and specialized habits since they can rely on a steady resource supply and need not regularly undergo energetically expensive fluctuations in population size.
Insofar as can be told from available data, the correlation of seasonality and diversity seems high in modern oceans (Valentine, 1971, 1973). The well-known latitudinal gradient in diversity parallels the latitudinal gradient in seasonality. Additionally, patterns of trophic resource seasonality that are created by hydrographic factors—alternations of nutrient-rich and nutrient-poor waters—are particularly important in ocean climates, because perturbations of the oceanic water column lead directly to nutrient supply (net upwelling) or removal (net downwelling). Horizontal currents also change nutrient-poor for nutrient-rich water on a seasonal basis in some regions (as along the northeastern American coast).
It would obviously be much to the point to compare the modern pattern of marine diversity, normalized to some standard measure, with the pattern of seasonality of productivity in the oceans today. Diversity data, though spotty, are improving and are probably sufficient to determine broad global patterns. The pattern of seasonality of productivity, however, is not known from observations, which are restricted to only a few scattered localities (there is a great lack of winter data). It must be inferred from general principles and from the hydrographic patterns. Not to be possessed of actual observations sufficient to produce a map of global marine seasonality of productivity is a serious gap in our knowledge of the oceans and of the global ecosystem. Generally, diversity displays an inverse relation to nutrient seasonality, just as it does to solar seasonality, whenever trends can be inferred (Valentine, 1973).
Factors other than seasonality affect local patterns of species richness. Some factors act to restrict the sizes of populations so as to release resources than can be utilized by additional populations. Predators may have this effect (Paine, 1966), and physical disturbances can also free resources, often by increasing spatial patchiness (Dayton, 1971). There is no evidence that global trends in predation or in disturbance regulate global diversities, however. Other possible diversity controls have been suggested, but when all are considered it seems reasonable to conclude that the effects of seasonality have the
largest claim to both theoretical and empirical bases (so far as the data are available) sufficient to explain global diversity patterns.
SEASONALITY PAST AND FUTURE
If seasonality is indeed the major determinant of global diversity patterns, then the major diversity trends as registered in the fossil record should indicate past patterns of seasonality. Diversity trends can be observed when the record is studied on a hemispheric or global scale (for example, Stehli, 1970). Furthermore, techniques are available to test hypotheses of seasonality based on diversity patterns. Seasonal growth rhythms are commonly recorded by tree rings and by shell-growth increments in shallow marine invertebrates. Indeed, rhythmic patterns in the growth of stromatolites (Panella, 1976) and in the deposition of iron formations (Trendall and Blockley, 1970) are interpreted as suggestive of seasonality in rocks over 2 billion years old.
Although seasonal patterns have not yet been systematically followed through time, it is clear from the data available that seasons have varied considerably. This has probably been in large part due to effects of plate tectonic processes, to the changing geographies of land and sea, to mountain ranges, to ocean currents, and to sea-level fluctuations. Biogeographic patterns are strongly affected by such changes, as indicated by other chapters in this volume. Seasonal patterns and their biotic reflections should also be affected.
In the geologic record there are indications that some periods were characterized by relatively broad, equable climates, when tropical (or at least low-latitude) faunas penetrated much further poleward than they do today, and latitudinal provinciality was relatively low. Indeed, during much if not all of the pre-Pleistocene, the record suggests a weaker latitudinal marine provinciality than today’s (Valentine et al., 1978). At some times, as in the Jurassic (see Chapter 17), there seem to have been only one or two marine provinces between equator and pole. Three or four may be inferred at other times (perhaps during the late Cretaceous). Today, by contrast, there are six or seven separate marine-shelf provinces along the northeastern Pacific shelf between the equator and pole (Valentine, 1966).
An important question is, how are patterns of marine seasonality affected when climates broaden? Because we can probably assume that the gradient in seasonality of solar radiation has been about the same during the Phanerozoic, the question becomes, how are patterns of seasonality of nutrient supplies affected when climates broaden? It is possible to develop hypotheses that suggest a general reduction of nutrient seasonality with warmer poles and broader climates, although the subject is so complicated (see Chapter 14) that the opposite notion, relating higher nutrient seasonality to a broader climate, cannot yet be ruled out. The question might be solved by contrasting diversity patterns of more broadly zoned with those of more narrowly zones periods. Additional important questions concern other factors that can significantly alter the nutrient regime and therefore the seasonal pattern of productivity in the world ocean. Such factors may include the pattern of continentality, of narrowness or breadth of oceans, of the width of continental shelves (the height of sea level), the prevalence of east-west versus north-south coastlines, and the geography of ocean gateways. Many such questions can probably be tested, with care, from fossil-diversity data.
In conclusion, seasonality appears to be a significant parameter in structuring the biosphere, and it would seem reasonable to encourage research efforts in two main areas, One is in recording and interpreting the pattern of seasonality of productivity in the present oceans; this is one of the more important and basic pieces of information about how the present world operates and its collection is quite technically feasible. The second area is in determining the global patterns of marine diversity of the past. When local effects are filtered out, these patterns should reflect the patterns of marine seasonality. Their relations can then be tested against the breadth of past climates and other relevant factors and employed in constructing models of pre-Pleistocene climates.
REFERENCES
Andrewartha, H.G., and L.C.Birch (1954). The Distribution and Abundance of Animals, U. of Chicago Press, Chicago, 789 pp.
Boyce, M.S. (1979). Seasonality and patterns of natural selection for life histories, Am. Nat. 114, 569–583.
Dayton, P.K. (1971). Competition, disturbance, and community organization: The provision and subsequent utilization of space in a rocky intertidal community, Ecol. Monogr. 41, 351–389.
Hutchinson, G.E. (1948). Circular causal systems in ecology, Ann. N.Y. Acad. Sci. 50, 221–246.
Hutchinson, G.E. (1965). The Ecological Theatre and the Evolutionary Play, Yale U. Press, New Haven, Conn., 139 pp.
Lord, R.D. (1960). Litter size and latitude in North American mammals, Am. Midl. Nat. 64, 488–499.
MacArthur, R.H., and E.O.Wilson (1967). The theory of island biogeography, Princeton Monogr. Pop. Biol. 1, 1–203.
May, R.M. (1973). Time-delay versus stability in population models with two and three trophic levels, Ecology 54, 315–325.
May, R.M. (1975). Stability and complexity in model ecosystems, Princeton Monogr. Pop. Biol. 6, 1–265,
May, R.M. (1979). The structure and dynamics of ecological communities, in Population Dynamics, R.M.Anderson, B.D. Turner, and L.R.Taylor, eds., Blackwell Science Publishers, Oxford, pp. 385–407.
May, R.M., and G.F.Oster (1976). Bifurcations and dynamic complexity in single ecological models, Am. Nat. 110, 573–599.
Mileikovsky, S.A. (1971). Types of larval development in marine bottom invertebrates, their distribution and ecological significance: A re-evaluation, Mar. Biol. 10, 193–213.
Paine, R.T. (1966). Food web complexity and species diversity, Am. Nat. 100, 65–75.
Panella, G. (1976). Geophysical inferences from stromatolite lamination, in Stromatolites, M.R.Waiter, ed., Elsevier, Amsterdam, pp. 673–685.
Smith, H.S. (1935). The role of biotic factors in the determination of population densities, J. Edon. Entomol. 28, 873–898.
Stehli, F.G. (1970). A test of the Earth’s magnetic field during Permian time, J. Geophys. Res. 75, 3325–3342.
Thorsen, C. (1950), Reproductive and larval ecology of marine bottom invertebrates, Biol. Rev. 25, 1–45.
Trendall, A.G., and J.G.Blockley (1970). The iron formations of the Precambrian Hamersley Group, Western Australia, Geol. Surv. Western Australia Bull. 199, 1–365.
Valentine, J.W. (1966). Numerical analysis of marine molluscan ranges on the extratropical northeast Pacific shelf, Limnol. Oceanogr. 11, 198–211.
Valentine, J.W. (1971). Plate tectonics and shallow marine diversity and endemism, an actualistic model, Syst. Zool. 20, 253–264.
Valentine, J.W. (1973). Evolutionary Paleoecology of the Marine Biosphere, Prentice-Hall, Englewood Cliffs, N.J., 511 pp.
Valentine, J.W., and F.J.Ayala (1978). Adaptive strategies in the sea, in Marine Organisms: Genetics, Ecology and Evolution, B. Battaglia and J.A. Beardmore, eds., Plenum, New York, pp. 323–346.
Valentine, J.W., T.C.Foin, and D.Peart (1978). A provincial model of Phanerozoic marine diversity, Paleobiology 4, 55–66.






