
CHAPTER NINE
Ecosystems, Ecosystem Services, and Biodiversity
Terrestrial and marine ecosystems supply the foundation for human well-being and livelihood through the food, water, timber, and other goods and services they provide. Advances over past decades have also revealed the importance of less visible but equally important services that ecosystems provide for society, such as water filtration, carbon storage, maintenance of biodiversity, protection from storm disturbance, and stabilization of local climates. Climate change has already led to a number of changes in both terrestrial and marine ecosystems, and future climate change will strongly influence biodiversity, ecosystem processes, and ecosystem services, adding to other stresses on ecosystems from human activities.
Some questions decision makers are asking, or will be asking, about ecosystems management in the context of climate change include the following:
-
How is climate change—including changes in temperature, precipitation, and the chemistry of the atmosphere and oceans—altering the distribution of species?
-
Will these changes have major economic and social consequences, such as the loss of pollination services or valuable fisheries?
-
How does climate change relate to other ecosystem stresses, such as pollution and habitat loss?
-
Can ecosystems be managed to improve their ability to adapt to anticipated changes?
-
Is it possible to manage forests and other ecosystems in ways that can help limit the magnitude of future climate change?
Decades of focused research on terrestrial and marine ecosystems and their biodiversity have improved our understanding of their importance for society and their interactions with other components of the Earth system. The findings have been the subject of many authoritative syntheses and assessments, including those by the Pew Oceans Commission (2003), the Pew Center on Global Climate Change (Parmesan and Galbraith, 2004), the U.S. Commission on Ocean Policy (2004), the Millennium Ecosystem Assessment (MEA, 2005), the Intergovernmental Panel on Climate Change (IPCC; Fischlin et al., 2007), the Heinz Center (2008a), the National Research Council (NRC,
|
BOX 9.1 Glacier National Park Glacier National Park is rapidly losing its namesake as summer temperatures rise and its glaciers disappear (see figure on facing page). The park, which straddles both the Continental Divide and the U.S.-Canada border in Montana, has lost about two-thirds of its glaciers since 1850 (Hall and Fagre, 2003). Plant and animal species are struggling to keep pace as suitable habitats retreat uphill as the climate warms. For example, pine trees are invading open grassland as the tree line migrates to higher elevation, in turn reducing fodder available for grazing mountain goats, bighorn sheep, and other ungulates. Glacier National Park exemplifies some of the key questions that land and natural resource managers face with climate change (Pederson et al., 2006):
All of these questions highlight the need for improved understanding of how plant and animal species will respond to climate change and other stresses. Projections of climate change on finer spatial scales would provide input for land managers to begin to assess the implications in their local context. Studies and models of the complex interactions among climate, biodiversity, ecosystem processes, and |
2008b), and the U.S. Global Change Research Program (CCSP, 2009b), among others. This chapter outlines some of the key impacts of climate change on terrestrial and marine ecosystems (see Box 9.1), including the effects of ocean acidification, and also briefly summarizes current scientific knowledge about the potential role of ecosystems in limiting the magnitude of climate change and possible strategies for helping ecosystems adapt to climate change and other environmental stresses. The last section of the chapter outlines key research needs in all of these areas.

TERRESTRIAL ECOSYSTEMS
Impacts of Climate Change on Land-Based Ecosystems and Biodiversity
A series of place-based observations, meta-analyses, and models indicate that climate shifts have already begun to change the geographical range of plants and animal species on land (IPRC, 2007c). In the extreme, some plants and animals have experienced
maximum range shifts over the past 30 years that approach the magnitude of those witnessed in the transition from last glacial maximum to the present (NRC, 2008b; Parmesan and Yohe, 2003). In the Northern Hemisphere, range shifts are almost wholly northward and up in elevation as species search for cooler temperatures (NRC, 2008b). Special stress is being placed on cold-adapted species located on mountain tops and at high latitudes where boreal forests are invading tundra lands and where Arctic and Antarctic sea ice is rapidly diminishing (e.g., polar bears and various species of seals and penguins [NRC, 2008b]). Warming of streams, rivers, and lakes also potentially affects cold-water fish, such as economically important salmon and trout, through impacts on reproduction, food resources, and disease. The IPCC estimates with medium confidence that approximately 20 to 30 percent of plant and animal species assessed so far are likely to be at increasingly high risk of extinction as global average temperatures exceed a warming of 3.6°F to 5.4°F (2°C to 3°C) above preindustrial levels (Fischlin et al., 2007).
The phenology of species (seasonal periodicity and timing of life-cycle events) is also changing with warming. Biological indicators of spring (e.g., timing of flowering, budding, and breeding) arrive in the Northern Hemisphere as much as 3 days earlier each decade, and the growing season is longer (Walther et al., 2002). Such changes can disrupt the synchronicity between species and their food and water sources, pollinators, and other vital interactions. It also affects the timing and severity of insect and disease outbreaks, wildfire, and other disturbances, challenging the capacity of ecosystems and those charged with managing them to deal with new disturbance patterns. For example, large and long-duration forest fires have increased fourfold over the past 30 years in the American West; the length of the fire season has expanded by 2.5 months; and the size of wildfires has increased several-fold (NIFC, 2008; Westerling and Bryant, 2008; Westerling et al., 2006). Recent research indicates that earlier snowmelt, temperature changes, and drought associated with climate change are important contributors to this increase in forest fire (Westerling et al., 2006). Climate change in the western United States is also increasing populations of forest pests such as the spruce beetle, pine beetle, spruce budworm, and wooly adelgid (Logan et al., 2003) and expanding their range into forested areas previously protected from insect attack. Climate change thus increases the complexity and costs of forest and fire management practices (Chapin et al., 2003; Spittlehouse and Stewart, 2003), which in turn are strongly affected by policy. These policies and practices can be better informed by linking downscaled climate models with hydrologic and fire-vegetation models to determine, under different projections of climate change, which ecosystems will be most vulnerable to wildfires (Westerling, 2009).
Climate change, including the higher levels of CO2 in the atmosphere that help to
drive it, also affects the functioning of terrestrial ecosystems and their living communities (Loreau et al., 2001; Tilman et al., 1997); this, in turn, changes how ecosystems influence the atmosphere and climate system (Steffen et al., 2004). Experimental and modeling studies (e.g., Field et al., 2007b; Reich et al., 2006) reveal that, in general, exposure to elevated CO2 and temperatures leads to increases in photosynthesis and growth rates in many plants, up to a point; thereafter, the trend may reverse owing to processes not yet fully understood (Woodward, 2002). Decomposition and associated release of CO2 back to the atmosphere also increase as temperatures warm. However, ecosystem processes such as plant growth and decomposition are also determined by interactions with other factors such as nitrogen and carbon supplies, soil moisture, length of growing season, land use, and disturbance (Eviner and Chapin, 2003). Despite this complexity, projections suggest that forest productivity, especially in young forests on fertile soils where water is adequate, will increase with elevated CO2 and climate warming. Where water is scarce and drought is expected to increase, however, forest productivity is projected to decrease (Janetos et al., 2008).
Climate warming alone is projected to drive significant changes in the range and species composition of forests and other ecosystems. Generally, tree species are expected to shift their ranges northward or upslope, with some current forest types such as oak-hickory expanding, others such as maple-beech contracting, and still others such as spruce-fir disappearing from the United States altogether (Figure 9.1). Importantly, however, whole forest communities or ecosystems will not shift their ranges intact. Plant and animal species will respond independently, according to their physiology and sensitivity to climate, resulting in the breakup of existing communities and ecosystem types and the emergence of new ones. The consequences of such reshuffling are not clear, either for the plants and animals that now exist together, or for the services those systems provide to humanity.
In addition to climate change, ecosystems and biodiversity are already being impacted by human activities. For example, human infrastructure such as farms, settlements, and road networks have directly or indirectly affected more than 50 percent of the ice-free, terrestrial surface of the Earth (Ellis and Ramankutty, 2008; Foley et al., 2005; Vitousek et al., 1997). As much as 41 percent of the vast expanse of the oceans has been affected by human activities, for example through eutrophication or fish stock depletion (Halpern et al., 2008). Considering indirect impacts, such as ocean acidification, ground-level air pollution, and climate change, virtually all ecosystems on Earth are being affected in some way by climate change, and other human pressures on ecosystems are also growing significantly (Auffhammer et al., 2006; Chameides et al., 1994; Orr et al., 2005).
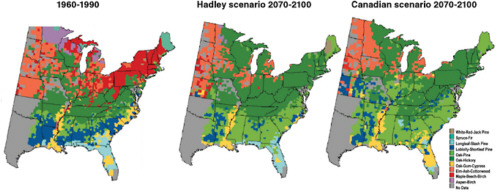
FIGURE 9.1 Potential changes in the geographic ranges of the dominant forest types in the eastern United States under projections of future climate change, based on the Hadley and Canadian climate models and a forest-type distribution model. Many forest types shift their ranges northward. Some types of forests, such as the loblolly-shortleaf pine in the Southeast (dark blue) or the maple-beech-birch forest type (red), shrink in area significantly or migrate to areas to the north and west. Oak-hickory (dark green) and oak-pine (light green) forest types expand their ranges. SOURCE: USGCRP (2001).
Managing the impacts of climate change on ecosystems and individual species already poses difficult challenges to land, resource, and conservation managers, and these challenges will undoubtedly increase. Past ecosystem conservation relied heavily on the assumption of a stable climate and focused on protecting individual species in place as well as preserving the habitat of entire species assemblages within protected areas. As climate change forces species to migrate to more suitable climates, ecosystems will be disassembled and reassembled in new locations, often outside the bounds of protection, and with new casts of characters. Some species will be lost, while other species will appear in new locations where they may become invasive and add to the pressures on existing species (NRC, 2008b).
Significant research is needed to better understand how climate change affects both individual species and entire ecosystems, and whether transitional or newly assembling ecosystems can continue to provide the ecosystem goods and services on which society depends (e.g., CCSP, 2008d; Fischlin et al., 2007). Moreover, social science research is needed to help land, resource, and conservation practitioners guide adaptive risk management in the face of altered species composition and a continually changing climatic and environmental baseline. In addition, very little is known yet about the social acceptability of new and evolving approaches to species conservation and land protection (including the Endangered Species Act under significant climate change, when many more species are at risk of extinction) or the social acceptability of a triage approach to species protection that may evolve as ecosystem functions are affected by climatic and species changes. Past experience with conservation management, however, indicates that societal values relative to species protection are significant to policy and practice. Integrated assessment and decision-support tools are also needed to help managers and the public understand and make wise judgments about the complex trade-offs that will be involved.
Role of Land-Based Ecosystems in Driving Climate Change
Modeling studies suggest that ecosystem responses to elevated CO2 result in a net carbon sink (that is, some of the elevated carbon generated by human activities is taken up and stored in plant tissues and soils, and the amount stored exceeds the carbon released through plant respiration and decomposition) and that this sink will persist through the twenty-first century (Schimel et al., 2000, 2001). When the models include temperature change as well as elevated CO2, however, they project that these carbon sinks could decrease, thereby increasing concentrations of CO2 in the atmosphere and reinforcing climate warming (e.g., Field et al., 2007a). Indeed, recent analyses suggest that the reduction in efficiency of land ecosystem sinks may already be
in decline (Canadell et al., 2007). Several major carbon sinks in terrestrial ecosystems face a high degree of risk from projected climate and land use changes (Fischlin et al., 2007). One of these is permafrost—frozen soil that covers vast areas of the northern latitudes and has locked away vast quantities of carbon. Permafrost temperatures are already rising due to high-latitude warming, creating a potential feedback that could drive further warming. Permafrost could also switch from a carbon sink to a source with thawing, releasing more carbon than in takes up (Dutta et al., 2006; Field et al., 2007a; McGuire et al., 2006; Norby et al., 2005; Zimov et al., 2006) and thus accelerating the pace of climate change. The potential for such a switch is one of several tipping points of concern in ecosystem-climate interactions (Barbier et al., 2008; Lenton et al., 2008; see also Chapter 6). Many other factors will ultimately determine whether terrestrial ecosystems provide a net feedback that enhances or slows the pace of climate change. Species redistributions, changes in major growth forms (e.g., from grass to woody plants, or from coniferous to deciduous trees), drought, length of growing seasons, air pollution, fire, insects and pathogens, deforestation and reforestation (Canadell and Raupach, 2008), and land use (Tilman et al., 2001) will influence uptake or release of CO2 and other greenhouse gases (GHGs) such as N2O and CH4 (Canadell and Raupach, 2008; Swann et al., 2010; Tilman et al., 2001).
Globally, as much as 35 percent of human-induced CO2 emitted over time to the atmosphere has had its origins in changes in land systems (both use and vegetative cover), principally deforestation (Foley et al., 2005). Biomass burning is also a major source of atmospheric aerosols (Andreae and Merlet, 2001). As discussed in Chapter 6, aerosols have direct effects on climate through scattering and absorbing solar radiation, and indirect changes in the properties and propensity for formation of clouds and hence precipitation, all of which can affect ecosystems (Lohmann and Feichter, 2005; Menon et al., 2002). Biomass burning is one of the largest sources of black carbon (soot) aerosols, a particularly potent warming agent that has been implicated in changing precipitation patterns and rapid ice melting in the Arctic (Flanner et al., 2007; McConnell et al., 2007; Wang, 2007). Finally, the emission of various trace gases by plants and from biomass burning leads to the formation of ground-level ozone, a gas that is both a climate-influencing agent and a pollutant that directly affects human and ecosystem health (Auffhammer et al., 2006; Chameides et al., 1994; Orr et al., 2005).
Land use change also influences climate by changing the reflective characteristics of the land surface and the exchange of water between the surface and the atmosphere. Deforestation, arid land degradation, and the transformation of ecosystems into built-up areas, for example, tend to increase reflectivity of the land surface and decrease evapotranspiration, leading to both local climate changes and, in combination with other land use changes, influencing large-scale climate forcing, feedbacks, and atmo-
spheric circulation patterns (Chapin et al., 2002; Pielke et al., 1998; Zhao et al., 2001). Deforestation tends to lead to warmer and drier climate conditions in the humid tropics, apparently due to reductions in evapotranspiration, (Bounoua et al., 2002; DeFries and Bounoua, 2004). Reductions in vegetation at high latitudes, on the other hand, tend to exert a cooling effect because more snow cover is exposed, increasing the reflection of solar radiation back to space (see Chapter 6 and Bonan, 1999). Afforestation (planting trees where they do not naturally occur), replanting forests in previously deforested areas, or shifts in evergreen species into previously shrub or forb areas could lead to increased absorption of solar radiation and thus increases in temperature (Bala et al., 2007). All these factors are important to the critical question of whether changes in terrestrial ecosystems accelerate or decelerate climate change, yet their combined role has not been evaluated. Importantly, these and other facets of ecosystem change not only influence the global climate system but also generate large local to regional climate implications as well (Cook et al., 2009; Durieux et al., 2003; Li et al., 2006; Malhi et al., 2008; Pielke et al., 1998).
Science to Support Managing Terrestrial Ecosystems to Limit the Magnitude of Climate Change
Managing land ecosystems provides opportunities to both limit the magnitude of climate change and ameliorate its negative consequences for society. Tropical deforestation and degradation, for example, contributed approximately 17 percent of anthropogenic carbon emissions in 2004 (Barker et al., 2007a). The opportunity to reduce emissions from deforestation and degradation (REDD) has been recognized within the United Nations Framework Convention on Climate Change as a relatively low-cost option to limit climate change (Gullison et al., 2007; Stern, 2007). Research is needed to support and improve such policies. While it is now a feasible goal to monitor changes in forest area by satellite throughout the tropics (DeFries et al., 2007; GOFC-GOLD, 2009), substantial uncertainties remain about the amount and distribution of biomass (carbon contained organic plant material such as leaves, branches, and roots). Accurate biomass estimates are critical for improving estimates of GHG emissions generated by deforestation (Houghton, 2005). Both ground-based measurements and new satellite technologies (e.g., Asner, 2009) for estimating above- and below-ground carbon are needed to improve these estimates.
Understanding the socioeconomic and ecological drivers of deforestation and degradation is also critically important for developing effective policies to reduce deforestation. Global-scale drivers, from international trade in agricultural products to subsistence needs by small-scale farmers, are complex and vary in different locations
(Nepstad et al., 2006; Rudel, 2005). Research focused on ecosystems needs to include intertwined climatic, ecological, and socioeconomic factors. For example, more clearing and more fires occur during relatively dry years in the tropical forests of southeast Asia, creating a positive feedback between emissions and climate change (van der Werf et al., 2008). The synergies and trade-offs between REDD and biodiversity conservation, watershed protection, and livelihood needs for local people require more rigorous analysis. (Research needs are discussed at the end of this chapter.)
Ecosystems also provide the opportunity to limit climate change through the enhancement of carbon storage or surface reflectivity. In forest ecosystems, protection from fire, insect damage, and forest thinning through logging and other human use can enhance carbon storage as can secondary regrowth of forests in abandoned croplands, tree plantations, and agroforestry (Rhemtulla et al., 2009; Gough et al., 2008). The extent to which these strategies might be able to offset GHG emissions on a global scale is poorly known. As noted above, land use and land cover changes also alter the reflectivity of the land surface, and this fact could potentially be exploited to limit the magnitude of climate change. Research is needed to evaluate these many interacting factors and quantify the potential and of these strategies relative to costs of adapting to climate change (Bala et al., 2007; Bonan, 2008; Jackson et al., 2008; Ollinger et al., 2008).
Ecosystems management is also a potential strategy to ameliorate some of the societal impacts of climate change. Restoration of wetlands in the Gulf of Mexico, for example, can reduce damage from hurricanes by damping wave action and diminishing wind penetration (Day et al., 2007). Mangroves protected people from a 1999 Asian cyclone (Das and Vincent, 2009) and will potentially provide some protection against storm surges that will move further inland with sea level rise. Quantitative and rigorous analysis of these ecosystem management opportunities, including their effectiveness and costs, is needed to assess their potential in different locations.
MARINE ECOSYSTEMS
Marine ecosystems are fundamental to the large role the oceans play in regulating the climate system. For example, the oceans contain many times more carbon than the atmosphere and terrestrial ecosystems combined, and are thus critical in regulating the amount of CO2 in the atmosphere. Climate change will have broad effects on marine ecosystems, their capacity to take up CO2 from the atmosphere, and the diverse ecosystem services they provide to society. These ecosystem effects will be driven by projected changes in ocean temperature, circulation (Bryden et al., 2005), storms, and
chemistry (Doney et al., 2009). Unlike on land, the majority of food that humans derive from the sea is still harvested from wild populations (FAO, 2008). Therefore, the oceans’ capacity to provide seafood, a major protein source for more than a billion people, will be directly affected by climate impacts on marine ecosystems (see Chapter 10).
Climate Change Impacts on Ocean Ecosystems
Over recent decades, marine scientists have detected widespread poleward shifts in species distributions that are consistent with patterns of a warming ocean (Alheit and Hagen, 1997; Holbrook et al., 1997; Mueter and Litzow, 2008; Sagarin et al., 1999; Southward et al., 1995). Marine species can be highly mobile, both as adults and as microscopic young drifting in the plankton (Kinlan and Gaines, 2003). This mobility can lead to larger and faster geographic shifts than in terrestrial ecosystems. For example, two-thirds of the 36 most common bottom-dwelling fish in the North Sea have shifted the geographical center of their range north toward the pole over just 25 years (Perry et al., 2005) (Figure 9.2). Such shifts, if they continue, could move the fish beyond the range of national fisheries. More broadly, because species move at different rates depending on their unique life histories, such shifts could lead to rapid rearrangements of the species composition of some ocean ecosystems (Cheung et al., 2009). The unpredictability of responses by different species is a key barrier to anticipating and adapting to the resulting ecosystem rearrangements.
Given the prominent role of oceans in storing carbon, climate impacts on ocean productivity could also alter their role in the carbon cycle. Overall, oceans contribute roughly half of the globe’s net primary productivity (NPP; Field et al., 1998), defined as the net carbon gain by ecosystems over a specific time period, typically annually. Some ocean habitats (polar seas, coastal upwelling systems) may see increased productivity under projected climate change (Arrigo et al., 2008; Bakun, 1990; Behrenfeld et al., 2006; Pabi et al., 2008; Polovina et al., 1995; Snyder et al., 2003). Most of the ocean, however, is permanently stratified with shallow, warm, nutrient-depleted water isolated from cold, nutrient-rich water below. In these seas, warmer surface temperatures generally decrease phytoplankton productivity (Figure 9.3). Given the prominence of these stratified seas, a substantially warmer ocean would “inevitably alter the magnitude and distribution of global ocean net air-sea carbon exchange, fishery yields, and dominant … biological regimes” (Behrenfeld et al., 2006).
Just as on land, high-latitude marine ecosystems may experience more stress than lower-latitude marine ecosystems, since rates of warming are higher (Gille, 2002; Hansen et al., 2006) and the opportunity for poleward range shifts is limited. Sea ice
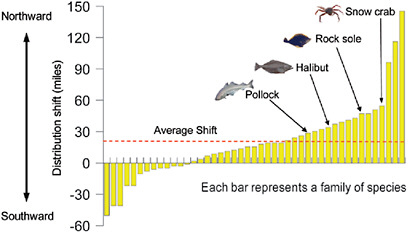
FIGURE 9.2 Observed northward shift of marine species in the Bering Sea between the years 1982 and 2006. Length of the yellow bars indicates the distance that the center of a species range has shifted. The average shift among the species examined was approximately 19 miles north of its 1982 location (red line). The northward shift is primarily linked to warming of the Bering Sea during this period. SOURCES: Mueter and Litzow (2008) and USGCRP (2009a).
creates critical habitat for a diverse array of marine species, including many mammals and birds (Hunt and Stabeno, 2002). Major declines in sea ice thickness and extent have been observed in the Arctic (see Chapter 6) and are projected for the next few decades (Overland and Stabeno, 2004; USGCRP, 2009a). Ice dynamics, which are highly sensitive to climate, drive dynamics of ocean primary productivity, which in turn has impacts throughout the marine food web in ways that are not clearly understood (Moore and Huntington, 2008; Smetacek and Nicol, 2005). Declines in sea ice can lead to large blooms in phytoplankton (e.g., Arrigo et al., 2008; Pabi et al., 2008) and declines in production from benthic (seafloor) habitats. These changes alter both the food webs of animals that ultimately depend on these different sources of productivity, including humans (Grebmeier et al., 2006; Mueter and Litzow, 2008; USGCRP, 2009a), and the role of high-latitude ocean ecosystems in the carbon cycle. Although the details are highly uncertain, many high-latitude ocean ecosystems appear to be at the threshold of major ecosystem changes (USGCRP, 2009a), especially since climate-induced changes may soon be joined by new human uses and stresses (e.g., oil and mineral exploration, expanded maritime use, and new fisheries in the Arctic) made possible by reductions in sea ice.
Some of the most productive ocean ecosystems are coastal regions where winds push
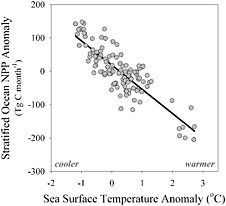
FIGURE 9.3 Relationship between changes in sea surface temperature and net primary productivity (NPP) from 1999 to 2004 based on satellite observations. Warmer ocean temperatures typically lead to reduction in the productivity of phytoplankton, which means that they remove less carbon from the atmosphere. SOURCE: Updated from Behrenfeld et al. (2006).
surface waters offshore and draw deep, cold, nutrient-rich waters to the surface (e.g., the west coast of North America). The nutrients fuel plankton blooms that support diverse and abundant food webs and fisheries. These upwelling regions may become even more productive under climate change if forecasts of increasing upwelling and favorable winds hold true (Bakun, 1990). Substantial increases in upwelling, however, can also have catastrophic consequences if the system crosses key thresholds (Chan et al., 2008; Helly and Levin, 2004). Deep ocean waters are typically extremely low in oxygen (hypoxic). Strong upwelling of deep cold waters can pull such hypoxic water onto shallow ocean shelves with devastating impacts on many marine species (Grantham et al., 2004). Hypoxia of coastal waters is more commonly associated with nutrient-laden runoff from land (NRC, 2000; Rabalais and Turner, 2001), but climate-driven changes in winds, ocean temperature, and circulation can cause similar devastation even in areas without runoff from land (Bakun and Weeks, 2004; Chan et al., 2008). The system can rapidly switch from high productivity to “dead zones,” where most species cannot live. For example, this transition has recently occurred in summers off the coasts of Oregon and Washington (Chan et al., 2008). Over more than 50 years of observations in the 20th century, hypoxia was rare or absent from these near-shore waters. In the past decade, however, hypoxia has become common and caused major die-offs of coastal species. By 2006, these once highly productive waters were oxygen-depleted along much of the coastline as upwelling winds increased (see Figure 9.4).
In the tropics, warm temperatures pose a “bleaching” threat to corals. Coral reef ecosystems have been compromised by a diverse set of activities including overfishing, damaging fishing practices, eutrophication, and sedimentation, among others (USGCRP, 2009a). On top of these human-caused stresses, recent decades have brought an
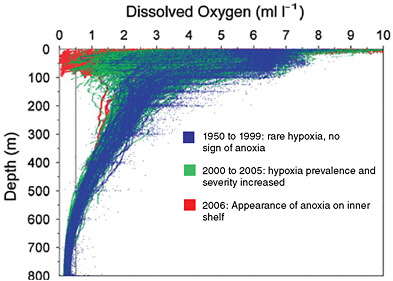
FIGURE 9.4 Hypoxia and anoxia in shallow waters. Values below 0.5 ml/l (left of black vertical line) represent severe hypoxia. Over the latter of half of the 20th century, hypoxia was only found in deep waters. In recent years (red and green), hypoxia has extended into waters close to the surface. SOURCE: Modified from Chan et al. (2008).
increase in widespread bleaching events, where corals eject their symbiotic algae in the face of extreme temperatures (Figure 9.5). In some cases, the bleached corals recover with new symbionts (Lewis and Coffroth, 2004). In other cases, the coral is killed. Periods of mass bleaching have occurred globally since the late 1970s (Glynn, 1991; Hoegh-Guldberg, 1999), with the most severe event in 1998, an El Niño year in which an estimated 16 percent of the world’s reef corals died (Wilkinson, 2000). The extent of bleaching varies greatly among species and locations. Some of the variability is tied to the level of other human stresses, which argues for managing reefs for greater resilience to climate change by reducing other stressors (Hughes et al., 2003). The next subsection discusses ocean acidification, which serves as an additional and potentially devastating stressor to corals. Recent models of coral-symbiont dynamics suggest that adaptation could greatly reduce coral bleaching catastrophes if the pace of climate warming is not too rapid (Baskett et al., 2005).
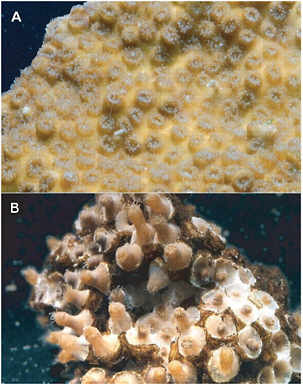
FIGURE 9.5 Photos of corals under normal (top) and acidified (bottom) conditions. The bottom coral lack a protective skeleton (appearing as light yellow in the top panel) and are sometimes called “naked coral.” SOURCE: Doney et al. (2009).
Ocean Acidification
In addition to its climate impacts, CO2 released by human activities can influence ecosystem dynamics in aquatic systems by altering water chemistry—in particular, the reaction of CO2 with water to form carbonic acid (H2CO3), which lowers (acidifies) ocean pH. Roughly one-third of all CO2 released by human activities since preindustrial times has been absorbed by the sea (Doney et al., 2009; Sabine and Feely, 2005; Sabine et al., 2004; Takahashi et al., 2006); consequently, ocean pH has decreased by approximately 0.1 units since preindustrial times. While this might not seem like a large change, it actually represents a 25 percent increase in acidity, because pH is measured on a logarithmic scale. By the end of this century, the oceans are projected to acidify by an
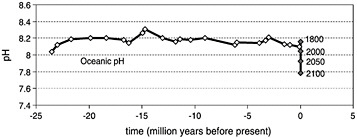
FIGURE 9.6 Estimates of ocean pH over the past 23 million years (white diamonds) and for contemporary times (gray diamonds). Projections are made for the future using IPCC projections of atmospheric concentrations of CO2. The projected changes in pH are extremely large and rapid, considering the relative stability of oceanic pH in the past. SOURCE: Blackford and Gilbert (2007).
additional 0.3 to 0.4 units (Orr et al., 2005) under the highest IPCC emissions scenario (Figure 9.6).
Because pH interacts with temperature to determine saturation levels for various related chemical species, cold-water ocean areas are projected to become undersaturated with calcium carbonate (CaCO3)—a key building block for the shells of many marine organisms—as early as 2050 (Orr et al., 2005). A broad array of marine species produce CaCO3 skeletons during at least part of their life cycle, so ocean acidification threatens nearly all ocean ecosystems by altering calcification rates while simultaneously increasing the rate of CaCO3 dissolution (Yates and Halley, 2006). Physiological studies suggest wide variations in the ability of organisms to cope with such changes (Doney et al., 2009). Acidification is especially challenging for coral reefs, which are defined by the CaCO3 skeletons of corals. Acidification, in tandem with elevated temperatures and other human stresses, decreased calcification rates on the Great Barrier Reef by 21 percent between 1988 and 2003 (Cooper et al., 2008). Numerous controlled experiments under elevated pH now complement these field observations (e.g., Doney et al., 2009). Projections of future ocean chemistry and climate change indicate that, by the time atmospheric CO2 content doubles over its preindustrial value, there will be virtually no place left in the ocean that can sustain coral reef growth (Cao and Caldeira, 2008; Silverman et al., 2009). Ocean acidification could also have dramatic consequences for polar food webs since several prominent species at the base of the food web may be unable to form shells—including species that salmon and other iconic species depend on for survival. Overall, ocean acidification has the potential to alter marine ecosystems catastrophically, but the details and consequences of these impacts are only beginning to be understood (see also NRC, 2010f).
The Role of Ocean Ecosystems in Managing Carbon and Climate Change
The ocean contains far more carbon than the atmosphere or land ecosystems. Storage of carbon in the ocean occurs by several mechanisms whose rates can be altered by human activities. In the ocean, CO2 dissolves directly in sea water; CO2 is sequestered when marine plants photosynthesize, and organic carbon ultimately sinks to great depths; and CO2 is also sequestered by conversion to CaCO3 by plankton, invertebrates, and fish (Wilson et al., 2009), CaCO3 that either forms sediments or sinks to deep water after the organism dies. None of these forms of storage is permanent, but sequestration rates can be modified greatly by a variety of factors (e.g., water temperature, pH, and the abundance of fish and plankton), ultimately affecting how much CO2 remains locked away or returns to the sea.
Because the oceans provide such an enormous reservoir for carbon storage, it may be possible to manipulate (i.e., geoengineer—see Chapter 15) ocean ecosystems to cause a transfer of CO2 from the atmosphere to the oceans. Several different approaches have been proposed to achieve this end, most of them involving the introduction of some kind of fertilizer to the upper ocean. The basic hypothesis is that fertilization may stimulate the incorporation of dissolved CO2 into organic matter through phytoplankton blooms, which could then sink to the deeper ocean. Some of the carbon that sinks out of the upper ocean should be replaced by CO2 from the atmosphere, thus reducing atmospheric CO2 concentrations.
Most of the attention given to the ocean fertilization hypothesis has focused on iron (Martin and Fitzwater, 1988; see also Limiting the Magnitude of Future Climate Change [NRC, 2010c]). In some parts of the ocean, especially the Southern Ocean and parts of the equatorial Pacific Ocean, marine biological productivity is limited by the availability of iron. The ratios of carbon to iron in marine phytoplankton typically exceed 10,000 to 1, so there is the potential that small amounts of iron could lead to substantial carbon uptake in the form of phytoplankton blooms. While there is still considerable uncertainty, the prevailing view is that this approach could store some carbon, but maximum achievable sustainable rates might be only a small fraction of the total carbon emitted due to fossil fuel emissions (Buesseler et al., 2008). There have been various proposals to fertilize the ocean with other nutrients, such as phosphate or nitrogen, or to fertilize the oceans by bringing up nutrients from the deep ocean, but these approaches have received even less study and attention on either their potential efficacy in reducing atmospheric CO2 or their broader environmental impacts.
In general, significant uncertainties remain about the effectiveness of ocean fertilization at removing CO2 from the atmosphere, as well as the length of time this CO2
would stay isolated from the atmosphere. Furthermore, there is considerable uncertainty about the impact of these manipulations on marine ecosystems and the services they provide to society, particularly since CO2 causes ocean acidification, which is expected to harm marine ecosystems. Much effort has been focused on trying to protect marine ecosystems by keeping CO2 out of the ocean, whereas ocean fertilization proposals seek to do the opposite. Because large parts of the oceans are a global commons, regulation of such activities represents a significant issue that has yet to be addressed. Furthermore, verification of amounts of carbon stored by ocean fertilization activities would be challenging, at best.
In summary, it is feasible that human manipulation of marine ecosystems could store at least some extra CO2 in the oceans. While maximum storage rates are projected to be at most a few percent of total human-generated GHG emissions, significant questions remain regarding exactly how much carbon could be stored, and for how long, using these approaches. Furthermore, considerations such as ocean acidification and the difficulty of predicting responses of marine ecosystems make it doubtful whether such manipulations could contribute to overall environmental risk reduction.
RESEARCH NEEDS
Improve understanding of the effects of climate change and impacts of enhanced CO2on ecosystems, ecosystem services, and biodiversity. Given the complexity of the impacts of different scenarios of climate change and elevated CO2 levels on ecosystem function, services, and biodiversity, further research is needed to evaluate the consequences of multiple interacting changes. For example, movement of species, changes in phenology and synchronicity, changes in productivity and carbon cycling processes, and changes in disturbance regimes in response to temperature, moisture, and CO2 have not been well assessed, especially at regional scales. Enhanced capacity for linking models of physical change in the climate system to species response models would help meet these challenges. Research is also needed to identify those ecosystems, ecosystem services, species complexes, and people reliant on them that are most resilient or most vulnerable (see Box 9.2).
Evaluate the climate feedbacks from changes in ecosystems and biodiversity. Changes in ecosystem biogeochemical processes (including GHG emissions) and biodiversity (including changes in reflectance characteristics) have the potential to exacerbate or offset certain aspects of climate change (i.e., act as feedbacks). Models and experiments that integrate knowledge about ecosystem processes, plant physiol-
|
BOX 9.2 National Marine Sanctuary Ocean ecosystems face growing threats globally from overfishing, habitat damage, pollution, and especially acidification (Halpern et al., 2008). As a result, the persistence of several marine species is at risk, and ecosystem services provided by intact coastal ecosystems could be compromised. Compared to the land, a minute fraction of the sea is set aside for protection. In response to growing threats, a number of nations, including the United States, are establishing networks of new marine protected areas (MPAs) with special protections (Airame et al., 2003; Fernandes et al., 2005). In the United States, the largest network of MPAs is being established along the coast of California, where dozens of new protected areas are currently being designed and implemented. Although MPAs can be dramatically successful at restoring depleted ocean ecosystems (Lester et al., 2009), many questions remain:
|
ogy, vegetation dynamics, and disturbances such as fire need to be further developed and included in advanced Earth systems models.
Assess the potential of land and ocean ecosystems to limit or buffer impacts of climate change. How can specific land uses (including managed and unmanaged forests and grasslands, agricultural systems, fisheries, urban systems, and aquatic systems) be managed for provisioning services as well as for their effects on GHG emissions, carbon storage, reflectivity, and evapotranspiration? What ecosystem management strategies can provide co-benefits that meet multiple goals, including carbon storage, biodiversity conservation, and watershed protection? To address these questions, new tools and approaches need to be developed for evaluating different land and ocean uses for their potential in helping to limit the magnitude of climate change. Such research needs to address the trade-offs between alternative land management options, including economic costs and impacts on ecosystem services that are difficult
to quantify in economic terms. The efficiency and efficacy of overlapping systems of governance and management structures to address trade-offs and determine management strategies is also a critical area of research.
Assess vulnerabilities of ecosystems and the benefits society derives from them to climate change. Ecosystems on land and in the ocean, and the services they provide, are key components of the maintenance of environmental functions and human well-being. Climate change affects this maintenance, with potentially significant societal consequences. Identifying critical linkages and feedbacks among changing ecosystems, their services, and human outcomes (e.g., crop yields, water supply) is essential. To do this requires analytical frameworks and methods for assessing vulnerability of coupled human-environment systems, and the ability of the social and environmental components of such systems to adapt to change. Complicating these assessments is the need to address climate change in the context of other changes, such as land use, acid rain, and nitrogen deposition.
Improve observations and modeling. There is a great need for global-scale, long-term, and continuous observations of land and ocean ecosystems and ongoing changes within them. Such observations will enable measures of ecological processes at relatively fine spatial and temporal scales, which are needed both to provide critical inputs to Earth system models and to track gradual and abrupt change in Earth system processes. The development of indicators of ecosystem health and ecosystem vulnerability is also needed as part of an early warning system (see NRC, 2009i). As mentioned earlier, new Earth system models that address multiple drivers and feedbacks from climate-ecosystem interactions are needed, and they will be most effective if linked to climate models that function at regional scales.




















