
Genomic Imprinting and the
Evolutionary Psychology of Human
Kinship
![]()
Genomic imprinting is predicted to influence behaviors that affect individuals to whom an actor has different degrees of matrilineal and patrilineal kinship (asymmetric kin). Effects of imprinted genes are not predicted in interactions with nonrelatives or with individuals who are equally related to the actor’s maternally and paternally derived genes (unless a gene also has pleiotropic effects on fitness of asymmetric kin). Long-term mating bonds are common in most human populations, but dissolution of marriage has always affected a significant proportion of mated pairs. Children born in a new union are asymmetric kin of children born in a previous union. Therefore, the innate dispositions of children toward parents and sibs are expected to be sensitive to cues of marital stability, and these dispositions may be subject to effects of imprinted genes.
“The burden of making and the duty of exacting compensation ran on the mother’s side as well as the father’s. A father and son, or two half-brothers, would for the purposes of the blood-feud have some of their kindred in common, but by no means all.”
Pollock and Maitland (1895)
___________________
Department of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138. E-mail: dhaig@oeb.harvard.edu.
The opening quotation comes from a discussion of Anglo-Saxon law. An individual could be liable to pay wergeld for the slaying of his mother’s kinsman by his father’s kinsman and be entitled to receive wergeld for the same slaying, because each individual combined two lines of descent. The individual is divisible. Just as his loyalties can be divided by obligations to the two sides of his family, so too can his genome be divided between genes he shares with his mother and genes he shares with his father. Blood is thicker than water, and blood does not mix (in the sense that genes do not blend).
Genetically determined behaviors that benefit the father’s side of the family may be favored by natural selection when a gene has been transmitted by a sperm but not when the same gene has been transmitted by an egg. Conversely, a behavior that benefits the mother’s side of the family may be favored when a gene has been transmitted by an egg but not when the same gene has been transmitted by a sperm. In such circumstances, imprinted alleles, genes that are differently expressed when inherited via eggs and via sperm, can supplant unimprinted alleles that are expressed independent of parental origin (Haig, 1997, 2000b).
Imprinted genes have been considered prime candidates for involvement in disorders of human social interaction, such as autism and schizophrenia, because of their predicted role in interactions among kin (Badcock and Crespi, 2006; Isles et al., 2006; Crespi, 2008; Goos and Ragsdale, 2008; Úbeda and Gardner, 2010, 2011). Not all social interactions promote imprinted gene expression, however. The principal purpose of this paper is to clarify the rather specific conditions that favor stable maintenance of imprinted gene expression, but this task requires a broader understanding of how humans innately categorize kin. These questions will be addressed with a particular focus on effects of partner change and on internal genetic conflicts during sexual maturation and adolescence.
ASYMMETRIES OF RELATEDNESS
Consider an imprinted locus at which the established allele is silent when paternally derived but expressed at level x >0 when maternally derived. This pattern of expression is an evolutionarily stable strategy (ESS) when two conditions are met:
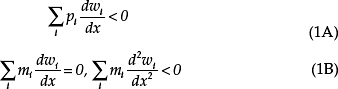
where dwi/dx is the effect of a change in x on the fitness of individual i, and pi and mi are coefficients of patrilineal and matrilineal relatedness of the category to which individual i belongs. Inequality (1A) specifies that extra x reduces patrilineal inclusive fitness. This condition maintains silence of paternally derived alleles. Eq. (1B) specifies that x is a local maximum of matrilineal inclusive fitness (Haig, 1997). These conditions are equivalent to
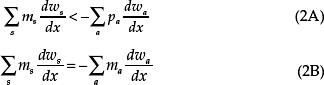
where s indexes symmetric kin (individuals for whom ms = ps) and a indexes asymmetric kin (individuals for whom ma ≠ pa). An individual’s symmetric kin include herself, her offspring, and her grandoffspring, but most other categories of kin are asymmetric, including “fullsibs,” because of uncertainty of paternity. Thus, the right-hand sides of 1A and Eq. (1B) can be considered to represent the marginal effect of x on the individual’s own survival and reproduction (individual fitness).
Eq. (2B) describes a tradeoff in the maximization of matrilineal inclusive fitness. At the ESS, the marginal effect of x on individual fitness is balanced by a marginal effect of opposite sign on indirect fitness obtained via asymmetric kin. If the value of Eq. (2B) is negative, then extra x increases individual fitness at a cost to matrilineal asymmetric kin. If the value of Eq. (2B) is zero, then x simultaneously maximizes both components of inclusive fitness (most plausible if x has no effects on matrilineal asymmetric kin). If the value of Eq. (2B) is positive, then extra x increases the fitness of matrilineal asymmetric kin at a cost to individual fitness.
Substitution of the right-hand side of Eq. (2B) for the left-hand side of 2A yields
![]()
which can be rearranged to give
![]()
where j indexes matrikin (individuals for whom mj > pj) and k indexes patrikin (individuals for whom mk < pk). This partition allows kin to be assigned to three mutually exclusive classes: symmetric kin (mi = pi),
matrikin (mi > pi), and patrikin (mi < pi). Inequality (3B) states that inactivation of the silent paternal allele is maintained when the summed effects of extra x on fitness are worse for patrikin than for matrikin, where fitness effects are weighted by the asymmetries of relatedness (terms in parentheses).
The ESS for a locus at which the established allele is silent when maternally derived but expressed at level z >0 when paternally derived is obtained by substitution of z for x and reciprocal substitution of m for p throughout the above analysis.
If maternal-specific expression of x has effects on two individuals only, (1A) and Eq. (1B) become

These conditions describe a tradeoff in which the two individuals’ fitnesses are differently weighted for genes of maternal and paternal origin. Condition (4A) can be expressed in a convenient form using a substitution from Eq. (4B):

Condition (4C) shows that the maintenance of paternal silence depends on a difference in the ratios of matrilineal and patrilineal relatedness for the two individuals affected.
If one of these individuals is the actor within whom x is expressed (p1= m1 = 1), then (4C) and Eq. (4B) become

Maternal-specific expression of x either benefits self at a cost to an individual who is patrikin (dw1/dx >0, dw2/dx < 0, p2> m2) or benefits an individual who is matrikin at a cost to self (dw1/dx < 0, dw2/dx >0, p2< m2). The ESS at a maternally silent locus, with paternal expression level z, is obtained by swapping m2 for p2 and z for x:
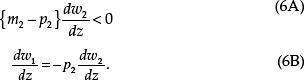
Paternal-specific expression of z either benefits self at a cost to an individual who is matrikin (dw1/dx >0, dw2/dx < 0, p2< m2) or benefits an individual who is patrikin at a cost to self (dw1/dx < 0, dw2/dx > 0, p2 > m2).
KINSHIP CATEGORIES
Other individuals evoke different innate dispositions in ego: some are sexual rivals, and others are potential mates; some are parents, and others are offspring; some are friends, and others are strangers. These dispositions constitute an implicit categorization of others that represents the way natural selection has parsed social interactions in a particular evolutionary lineage. The dispositions define the categories: all individuals who evoke a disposition belong to a category defined by the disposition. If the members of a category are, on average, related to ego, then the disposition will evolve, in part, shaped by its effects on the fitness of kin. An individual who evokes the disposition can be considered to be treated as a kinsman and the disposition can be considered a kin-directed behavior.
An actor can treat another individual as belonging to an innate category without recognizing that a category exists or recognizing particular individuals as members of the category. As a simple example, hormones secreted into the maternal circulation by a fetus affect another individual who necessarily carries copies of the maternally derived alleles of the fetus (Haig, 1996). However, if a behavior is to be preferentially directed toward a particular category of kin within a larger group of similar individuals, then the actor must discriminate among individuals and the individuals that belong to a category must be learned by social context. As a classic example, goslings have an innate disposition to follow “mother,” but the individual that is recognized as “mother” by a particular gosling is learnt through a process of imprinting (in an earlier sense of the word). Similarly, human children may possess innate dispositions in their interactions with “mother,” “father,” “brother,” or “sister,” but the particular individuals who evoke these dispositions must be learnt from social context.
We probably possess more-or-less discrete instinctive categories for primary kin, such as “self,” “mother,” “sister,” and “daughter,” and perhaps for some secondary kin, such as “sister’s daughter” or “daughter’s daughter.” However, for more distant kin, I suspect we possess a
vague sense of some individuals as closer kin than others, with behavioral dispositions that vary with degree of perceived kinship. Where we place a particular individual on this continuum will be determined by things we have been told, and how often, and in what contexts we have interacted with them, particularly during childhood.
A disposition evolves according to the average relatedness of the individuals who evoke the disposition, not according to the relatedness of any particular genealogical category. Thus, instinctive categories should be distinguished from the categories that would be determined by an omniscient geneticist. Suppose, for example, that a disposition is evoked by females, born within a few years of the actor, who live in close association with the actor’s “mother” during the actor’s own childhood. Such a disposition will often have been evoked by genealogical sisters, and the innate category can be labeled, for convenience, as “sister,” even though it may sometimes have been evoked by individuals who were not offspring of the actors’ mothers.
Hamilton’s second principle of the genetical evolution of social behavior was that “The situations in which a species discriminates in its social behaviour tend to evolve and multiply in such a way that the coefficients of relationship involved in each situation become more nearly determinate” (Hamilton, 1964b). In other words, natural selection will tend to favor actors who are able to subdivide beneficiaries into categories with a lower variance of genealogical relatedness. By this process, innate categories would more nearly come to approximate genealogical categories. However, this conclusion should be qualified by the observation that an individual may benefit from being classified by an altruistic actor as a member of a category with a coefficient of relatedness greater than the individual’s “true” relatedness. Thus, natural selection on actors to make ever finer discriminations of kinship may be opposed by natural selection on a subset of beneficiaries to confound such discrimination (Haig, 2000a).
Instinctive categories should also be distinguished from the cultural classification of kin (Feinberg and Ottenheimer, 2001). Cultural evolution exploits our innate dispositions for various cultural and rational ends. For example, by defining another individual as a brother, a cultural tradition or a political innovator attempts to evoke dispositions appropriate to innate brotherhood in interactions with that individual (Johnson, 1987; Salmon, 1998). To the extent that this evocation is successful, culture thereby changes the coefficients of relatedness associated with an innate category in ongoing natural selection. By this means, culture can shape the innate taxonomy of kin.
Symmetric Kin
“Self” and “offspring” are symmetric kin. These innate categories are evolutionarily ancient and undoubtedly have accrued a rich set of innate dispositions. “Grandoffspring” are also symmetric kin. Genes of maternal and paternal origin favor the same outcomes when fitness tradeoffs affect symmetric kin alone. Therefore, significant effects of imprinted genes on symmetric kin are predicted only if a gene’s expression also affects asymmetric kin. For example, gene expression might mediate a direct tradeoff between the fitness of symmetric kin (e.g., “self”) and asymmetric kin (e.g., “mother”).
Perfect symmetry of matrilineal and patrilineal relatedness is an ideal that is probably rarely realized, although selection favoring imprinted expression will be weak when asymmetries of relatedness are small. For example, fullsibs, considered as a genealogical category, are symmetrically related to ego. However, “fullsibs,” considered as an innate category, are ego’s matrikin because of the possibility of undetected cuckoldry. The asymmetry of relatedness associated with “fullsibs” will be small, however, whenever the probability of shared paternity is high. As another example, ego’s offspring will be asymmetrically related to ego when ego’s spouse is asymmetrically related to ego, as occurs under some forms of inbreeding (Haig, 1999; Wilkins and Haig, 2003a), but the asymmetries of relatedness will be small, except under close inbreeding.
Mother
“Mother” is the most important category of matrikin. Strong effects of imprinted genes are expected in an offspring’s relations with its mother, both prenatally and postnatally, because mothers have large effects on the fitness of offspring and are associated with an extreme asymmetry of relatedness from an outbred offspring’s genetic perspective (m - p = 1). Paternally expressed genes are predicted to increase the demands offspring impose on mothers, whereas maternally expressed genes are predicted to reduce these demands (Wilkins and Haig, 2003b; Haig, 2004). The strength of these effects will be attenuated when mothers establish stable breeding bonds with a particular male because paternally derived genes of an offspring then have an interest in a mother’s continued reproduction. Her future offspring are potentially also the offspring’s father’s future offspring. However, when a mother changes partner, her continued reproduction expends limited maternal investment on maternal halfsibs, who are unrelated rivals from the perspective of paternally derived genes of existing offspring.
Effects of imprinted genes during fetal development are broadly consistent with theoretical predictions that paternally expressed genes should enhance growth and maternally expressed genes should restrain overgrowth (Eggermann et al., 2008; Haig, 2010). Evolutionary speculation about postnatal effects of imprinted genes on a child’s relations with his or her mother has focused on the phenotypes of Prader-Willi syndrome (PWS) and Angelman syndrome (AS), with the former caused by loss of paternally expressed genes at 15q11–13 and the latter by loss of maternally expressed genes from the same region (Buiting, 2010). Therefore, PWS is predicted to exhibit absence, or weak development, of behaviors that elicit resources from mothers, whereas AS is predicted to exhibit an overdevelopment of such behaviors (Haig and Wharton, 2003; Úbeda, 2008).
Infants with PWS exhibit poor suck, weak cry, and excessive sleepiness, suggesting paternally expressed genes from 15q11–13 promote suckling, strength of cry, and wakefulness (all phenotypes that are expected to enhance maternal costs). From about the age of natural weaning, children with PWS develop an insatiable appetite associated with “foraging” behaviors. These phenotypes have been interpreted as a pathological expression of “weaning conflicts” that occurred when our ancestors were transitioning from predominant reliance on the breast to reliance on supplemental foods (Haig and Wharton, 2003; Úbeda, 2008).
The happy affect and smiling demeanor of children with AS contrast with the less effusive personality of children with PWS. Children with AS are proposed to express strongly behaviors that normally function to elicit maternal care, attention, and attachment (Isles et al., 2006; Brown and Consedine, 2004). However, the overtly social personality of children with AS is combined with a profound deficit in communication (Haig, 2008). Speech and gesture are largely absent. Therefore, maternally expressed genes at 15q11–13 appear necessary for the normal development of language. Badcock and Crespi (2006) have suggested that genes of maternal origin have been selected to act in the language centers of the child’s brain to promote attentiveness to maternal instruction and maternal example, coordinating maternal and child needs for the benefit of the matriline.
Father
Fathers are patrikin of their offspring. Therefore, maternally expressed genes in offspring are predicted to favor increased demands on fathers relative to the effects of paternally expressed genes. By
contrast, paternally expressed genes are predicted to show greater solicitude to the needs of fathers.
Most mammals probably lack an innate category of “father.” The evolution of more-or-less stable mating bonds between men and women has allowed fathers to recognize their offspring and offspring to recognize their fathers (often with a fair degree of confidence). Human fathers recognize offspring as babies born to women with whom they are involved in a more-or-less exclusive sexual relationship. Human offspring recognize fathers as adult males closely associated with their mother during infancy and early childhood (Chapais, 2008).
Recognition of fathers means fathers can be avoided as mates. From the genetic perspective of a daughter, mating with her father is associated with a direct cost of producing inbred, rather than outbred, offspring but an indirect benefit of an extra, albeit inbred, paternal half-sib (with the daughter herself as the mother). The direct cost is experienced equally by the daughter’s maternal and paternal alleles, but the indirect benefit is experienced solely by the daughter’s paternal alleles. Therefore, maternally expressed genes are predicted to promote strong aversions to sexual relations with fathers (Haig, 1999).
Sibs
Uterine sibs sometimes have different fathers and are thereby matrikin. Paternal-specific expression of imprinted genes is expected to benefit self at the expense of uterine sibs, whereas maternal-specific expression is expected to benefit uterine sibs at a cost to self. Two factors in human evolution have probably had opposing effects on the intensity of conflict between genes of maternal and paternal origin over relations with uterine sibs. On the one hand, the evolution of persistent pairbonds increased the proportion of uterine sibs that were fullsibs, thus reducing asymmetries of relatedness and mitigating potential conflicts. On the other hand, the evolution of shorter interbirth intervals and prolonged childhoods increased opportunities for competition among sibs (Kennedy, 2005; Sellen, 2007; Humphrey, 2010).
Sibs are both sharers of common genes and competitors for common resources; hence, the characteristic admixture of affection and aggression in many sibling relations. Innate dispositions toward sibs are expected to be sensitive to relative age, with rivalry more intense among sibs who are closer in age. An older sib usually has greater power to help or harm a younger sib than the younger has to help or harm the older (although younger sibs will often attempt to recruit the even greater power of a parent on their behalf). The age-related asymmetry
in power between a pair of sibs is expected to lessen as they grow older because they become closer together in relative age.
“Younger uterine sib” is likely to have been an evolutionarily salient category of matrikin because the arrival of a new baby will often have been accompanied by a reapportionment of maternal care away from older sibs. Consider two scenarios. In the first, a child grows up with his or her mother and a “father” who disappears and is replaced by an unfamiliar adult male, after which the mother has a new baby. From the perspective of the older child, the new baby is associated with a large asymmetry of relatedness (m - p = 0.5). In the second scenario, the “father” and mother remain together for the birth of a new baby. In this scenario, the new baby is associated with a much smaller asymmetry of relatedness because he or she is likely (although not certain) to be a fullsib of the older child.
New babies evoke a single instinctive category if the innate dispositions of older sibs are the same in the two scenarios. In this case, gene expression will have evolved according to a gene’s average relatedness to babies in the different scenarios, weighted by the long-term average frequencies of each scenario. If the two scenarios evoke different innate dispositions, then the babies belong to different instinctive categories and gene expression will have evolved according to scenario-specific coefficients of relatedness.
Innate dispositions of younger children toward “older uterine sibs” are probably less responsive to a mother’s change of partner than dispositions of older children to “younger uterine sibs” because a newborn child has not herself or himself experienced the change of partner and has few direct cues about the paternity of older sibs. The latter are more powerful and better informed than younger sibs, and thus may often set the tone of sibling relations. Infants and toddlers may, at first, express behaviors designed to ingratiate themselves to older sibs in an attempt to elicit help and avoid harm, with the overt expression of rivalry intensifying as disparities of power lessen with age.
Paternal halfsibs are patrikin, but it is unclear whether we have evolved innate dispositions that are specific for this category of kin. Relations with paternal halfsibs are usually less intimate than relations with maternal halfsibs because a father’s contact with his offspring becomes attenuated once his sexual relations with their mother ends, especially if he has offspring with another woman. Interactions with paternal halfsibs are more intense in polygynous households in which the offspring of two or more women compete for family resources (Jankowiak and Diderich, 2000), but this situation has probably been less frequent than living with maternal halfsibs.
Extended Kinship
Asymmetries of matrilineal and patrilineal relatedness are created whenever individuals of one sex disperse to reproduce, whereas individuals of the other sex remain in their natal group. If the variance of reproductive success is similar in the two sexes, then random pairs of individuals are more likely to share genes of maternal origin than genes of paternal origin in matrilocal groups with male-biased dispersal, but the reverse is true in patrilocal groups with female-biased dispersal (Haig, 2000a, 2010; Brandvain, 2010; Úbeda and Gardner, 2010, 2011; Van Cleve et al., 2010). Thus, differential dispersal of the sexes can result in genes having effects that discriminate between matrikin and patrikin without other individuals being explicitly recognized as belonging to the mother’s family or father’s family.
Whether human reproductive dispersal has been female-biased or male-biased, on average, is controversial (Ember, 1978; Alvarez, 2004). What is not controversial is that human groups exhibit a flexibility of social organization such that ties of matrilineal and patrilineal kinship predominate in different populations, with strong ties to both sides of the family maintained in many groups (Rodseth et al., 1991; Marlowe, 2004; Chapais, 2008). Most, if not all, cultures distinguish between matrilineal and patrilineal kin. A key unanswered question is whether this cultural distinction is reinforced by innate dispositions that distinguish “mother’s kin” from “father’s kin,” or whether the two kinds of kin are lumped together in a single instinctive category with asymmetries of relatedness determined by social context.
The recognition of particular individuals as belonging to particular categories of kin enables discrimination among members of social groups on the basis of degree of relatedness (nepotism). The evolutionarily oldest and strongest ties are between mothers and their offspring, and among uterine sibs. If adults maintain associations with their mothers and uterine sibs, then second-order ties are facilitated between children and their mother’s mother and mother’s sibs (Chapais, 2008). Strong ties of patrilineal kinship are more tenuous because recognition of fathers is less certain and (evolutionarily) more recent. Perhaps for these reasons, ties of patrilineal kinship are often reinforced by strong patriarchal ideologies.
KITH AND KIN
Queller (2011) distinguishes social effects mediated via kin from those mediated via kith or kind. Kin selection involves fitness effects on individuals who share genes via genealogical descent. Kind selection involves fitness effects mediated by identity by state rather than by
descent. Kith selection involves an actor’s effects on other individuals that feed back to the actor’s own individual or inclusive fitness. I will not discuss kind selection except to draw attention to Queller’s perceptive discussion of the relation between “phenotype matching” and green-beard effects (Queller, Chaper 1, this volume).
Each individual has two parents who may be genetically unrelated but have a common interest in the survival and reproduction of their mutual offspring. The parents are each other’s kith, and their relation engenders a complex intertwining of kith and kin effects because the affines of the father and mother are, respectively, matrikin and patrikin of the offspring. Put another way, an individual’s matrikin are kith from the perspective of paternally derived genes, whereas an individual’s patrikin are kith from the perspective of maternally derived genes. Further entanglement of kith and kin occurs when parents are themselves kin because of consanguineous matings.
A husband may benefit from investment in the health and well-being of his wife because this feeds back to increased fitness of his children. By extension, a husband is kith of his wife’s family, who are matrikin of his offspring. His investment in relations with his wife’s parents, and their investment in their son-in-law, may feed back to increased fitness of his children and their grandchildren. By further extension, a mother is kith of the paternally derived genes of her own offspring. These genes have an interest in her well-being to the extent that the offspring’s individual fitness depends on continued investment by a healthy mother. Moreover, the offspring’s patrilineal inclusive fitness may benefit from maternal investment in fullsibs.
Kith relations are contingent in ways that kin relations are not. The love of a child is more robust to bad behavior by the child than is love of a spouse to bad behavior by the spouse. The sharing of genes by descent is a brute fact that is unchanged by changes in the personal relations of kin, but spousal fitnesses are decoupled when either partner pursues other reproductive opportunities.
PARTNER CHANGE
In preindustrial societies, it was a lucky child who reached maturity living in a household with both biological parents because of high rates of parental death and divorce (Hewlett, 1991; Marlowe, 2005). Some of our ancestors undoubtedly grew up in families with both parents present, but others grew up in families in which one or both parents were absent. Behaviors that best promoted inclusive fitness are likely to have differed between intact and disrupted families because parents differed in their ability (or willingness) to invest in offspring and divorce
was associated with predictable changes in relatedness for the children of former marriages.
As long as a couple remains together, their mutual offspring are fullsibs and symmetric kin of existing offspring (ignoring, for the moment, children conceived by extrapair copulations). However, once parents change partners, subsequent offspring of the mother and father are, respectively, maternal halfsibs (matrikin) and paternal half-sibs (patrikin) of the parents’ mutual offspring. Therefore, a child’s innate dispositions toward parents and younger sibs should be sensitive to whether or not his or her parents remain together, and these dispositions may be particularly sensitive to influences of imprinted genes after divorce or parental death.
Conflict in a child’s relations with his or her parents is expected to intensify after parental separation, especially after parents acquire new partners (Emlen, 1995, 1997b), because genes of maternal origin in the child have no direct interest in the father’s continued reproduction, whereas genes of paternal origin have no direct interest in the mother’s continued reproduction. Therefore, genes of paternal origin are expected to promote reduced cooperation with mothers after divorce, either expressed as increased demands for maternal resources, increased competition with maternal halfsibs, or reduced expression of helpful behaviors. Genes of maternal origin are expected to have opposing effects. As a result, conflicts within the child’s genome are predicted to intensify after divorce.
When marriages dissolve, children usually remain with their mothers and contact with their fathers declines; social interactions with maternal halfsibs tend to be stronger than with paternal halfsibs; and ties to the mother’s extended family strengthen, whereas ties to the father’s family weaken (Furstenberg and Cherlin, 1991). If similar biases were present in our evolutionary past, then the dissolution of pairbonds would have been associated with a statistical shift toward greater interaction with matrikin and a concomitant shift in the selective forces acting on imprinted genes in children. Paternally derived genes of children would therefore favor a greater emphasis on self-beneficial behaviors and a reduced emphasis on kin-beneficial behaviors after parental divorce.
SEXUAL MATURATION
Age at sexual maturity is a pivotal life-history variable (Stearns, 1992). In standard life-history theory, risk of death is the primary factor favoring earlier reproduction because individuals who delay maturation may not survive to reproduce or may not remain alive long enough to raise their offspring. Other things being equal, higher mortality of
young adults favors earlier reproduction. Thus, high risks of subadult mortality have been proposed to explain early reproduction at small size in human pygmies (Walker et al., 2006; Migliano et al., 2007). Early maturation, in this case, is assumed to reflect a genetic change in the pygmy gene pool. Facultative responses are also possible. Thus, early reproduction by poor African-American women has been interpreted as a rational response to low life expectancy (Geronimus, 1997).
Theoretical discussions have focused on effects of pubertal timing on individual fitness with indirect effects on the fitness of relatives, for the most part, neglected. In this section, I will focus on indirect effects. My motivation is that a number of imprinted regions of the human genome influence pubertal progression and timing. This suggests that variation in age at maturity has affected the fitness of asymmetric kin, as well as individual fitness (Haig, 2010). I do not address the relative importance of direct and indirect effects. The selective forces acting on pubertal timing are undoubtedly complex, and a comprehensive review is beyond the scope of this paper.
The timing of ego’s transition to adulthood would have had varied consequences for the fitness of ego’s kin depending on ecological conditions: whether ego remained in his or her natal group or moved to another group, how much ego contributed to communal goods, and the extent to which ego’s offspring competed for limited resources with other group members (Haig, 2010). Rather than attempt a global analysis that sums fitness effects across all categories of kin, I will consider a simple model in which the level of x (expressed from one locus) accelerates ego’s pubertal development, whereas the level of z (expressed from another locus) decelerates pubertal development, and consider two ways in which ego’s age at maturity could affect the fitness of a younger uterine sib.
In the first scenario (another-mouth-to-feed), ego (m1= p1= 1) and a younger sib (m2= 0.5 > p2) compete for limited maternal investment until ego leaves the parental home. Earlier puberty reduces ego’s fitness (?w1/![]() x < 0,
x < 0, ![]() w1/
w1/![]() z > 0) at a benefit to the younger sib (
z > 0) at a benefit to the younger sib (![]() w2/
w2/![]() x >0,
x >0, ![]() w2/
w2/![]() z < 0), who acquires more resources because of reduced competition with ego. Ego’s genes of paternal origin have less of an interest in the younger sib’s welfare than ego’s genes of maternal origin. Therefore, this scenario predicts maternal-specific expression of accelerators of puberty and paternal-specific expression of decelerators of puberty. From Eq. (5B) and Eq. (6B), the joint ESS is characterized by
z < 0), who acquires more resources because of reduced competition with ego. Ego’s genes of paternal origin have less of an interest in the younger sib’s welfare than ego’s genes of maternal origin. Therefore, this scenario predicts maternal-specific expression of accelerators of puberty and paternal-specific expression of decelerators of puberty. From Eq. (5B) and Eq. (6B), the joint ESS is characterized by
![]()
Ego is predicted to undergo puberty at a younger age than is optimal for his or her individual fitness. Production of x is “altruistic” because it benefits the younger sib at a cost to self, whereas production of z is “selfish” because it benefits self at a cost to the younger sib.
In the second scenario (helper-at-the-nest), ego helps raise the younger sib by providing child care and contributing food to the household pot or otherwise reducing maternal workload (Turke, 1988; Bereczkei and Dunbar, 2002; Kramer, 2002; Hrdy, 2009), but this help is withdrawn when ego begins to reproduce on his or her own. Earlier puberty enhances ego’s individual fitness (![]() w1/
w1/![]() x >0,
x >0, ![]() w1/
w1/![]() z < 0) at a cost to the fitness of the younger sib (
z < 0) at a cost to the fitness of the younger sib (![]() w2/
w2/![]() x < 0,
x < 0, ![]() w2/
w2/![]() z >0). Ego’s genes of paternal origin have less interest in the fitness of the younger sib than ego’s genes of maternal origin. Therefore, this scenario predicts maternal-specific expression of decelerators of puberty and paternal-specific expression of accelerators of puberty. The joint ESS is characterized by
z >0). Ego’s genes of paternal origin have less interest in the fitness of the younger sib than ego’s genes of maternal origin. Therefore, this scenario predicts maternal-specific expression of decelerators of puberty and paternal-specific expression of accelerators of puberty. The joint ESS is characterized by
![]()
Ego is predicted to undergo puberty at an older age than is optimal for his or her individual fitness. Production of x is “selfish” because it benefits self at a cost to the younger sib, whereas production of z is “altruistic” because it benefits the younger sib at a cost to self.
Human sexual maturation is delayed relative to the other great apes. The two scenarios make different predictions about the reason for delayed maturation in the human lineage. In the another-mouth-to-feed scenario, delayed maturation is favored because it allows ego to accumulate more embodied capital and become a better parent (Gurven et al., 2006). In the helper-at-the-nest scenario, delayed maturation is favored because ego obtains greater returns from indirect investment in a younger sib than from direct investment in his or her own offspring (Reiches et al., 2009; Kramer and Ellison, 2010). The two scenarios are, of course, not mutually exclusive: A child can both compete with his or her sibs for limited resources and provide help to his or her parents. Moreover, experience gained in care of younger sibs is a form of embodied capital when a child has offspring of his or her own.
What would one expect if age at maturity were contingent on whether ego’s mother and father stayed together for the birth of the younger sib? Partner change causes a decrease in p2, thus discounting the cost of competition with the younger sib, or discounting the benefit of helping raise the younger sib, for genes of paternal origin. In the another-mouth-to-feed scenario, reduced relatedness to younger sibs
is predicted to favor increased production of z from alleles of paternal origin to slow the onset of puberty. Partner change, by itself, does not promote a change in x because m2 is unchanged. However, the acceleration of puberty due to increased z may favor enhanced production of x from alleles of maternal origin as a countermeasure. By contrast, in the helper-at-the-nest scenario, reduced relatedness to younger sibs directly favors increased production of x from alleles of paternal origin to hasten the onset of puberty.
Blended vs. Unblended Relatedness
What coefficient (or coefficients) of relatedness should be associated with an innate kinship category in models of inclusive fitness? Inclusive-fitness theory usually employs a coefficient that averages relatedness for alleles of maternal and paternal origin, as if maternal and paternal alleles were blended together in offspring rather than retaining their separate identities. By contrast, the present paper uses parent-specific coefficients. There has been surprisingly little discussion of if, and when, blending is appropriate given that the two approaches make different kinds of predictions about what should be observed in nature. I will use the helper-at-the-nest scenario to illustrate the difference of approach and predictions.
A “conventional” model of pubertal timing would use age at puberty, y, as the variable for direct optimization. Larger values of y would be associated with a benefit to the younger sib (dw2/dy) at a cost to self (-dw1/dy). At the optimal age of puberty,
![]()
where r2= (m2 + p2)/2 is a coefficient that blends matrilineal and patrilineal relatedness. By contrast, an “imprinting” model would treat y as a function of the level of expression of one or more genes [e.g., y = Φ(x, z), where x is a pubertal accelerator and z is a pubertal decelerator] [Eq. (8)]. The model then makes statements about levels of gene expression at evolutionary equilibrium.
This comparison immediately identifies the attraction of the conventional approach. The use of blended relatedness allows statements to be made about phenotype y, whereas the use of parent-specific relatedness views y as an issue in dispute between opposing parties. The latter models usually do not predict how the dispute will be resolved at the level of outward phenotype. The differences should not be overstated. When considering the effects of husband replacement on optimal age at puberty, r2 (the blended coefficient) changes in the same direction as p2 (the patrilineal coefficient), with m2 (the matrilineal coefficient) unchanged. Therefore, predictions about the direction of change
in phenotype will be similar for the two kinds of model, absent the possibility that genes of paternal origin have no influence. Nevertheless, I would argue that parent-specific coefficients are more appropriate than blended coefficients except in the special case where effects of imprinted alleles can be excluded. Even in the latter case, my preference is to use parent-specific coefficients and let the “blending” of relatedness emerge from the mathematics of the model.
Effects of Father Absence
Girls are reported to enter puberty and begin reproduction at younger ages when raised in households in which biological fathers are absent (Surbey, 1990; Maestripieri et al., 2004). Previous attempts to explain this association have focused on the role of father absence as a predictor of the daughter’s expected fitness when “choosing” among alternative reproductive strategies. Thus, absence of her father during a girl’s early childhood has been proposed to predict lower paternal investment by the potential fathers of her own offspring. Her poor prospects of finding a reliable spouse have been conjectured to favor an earlier onset of reproduction (Belsky et al., 1991). A somewhat simpler hypothesis is that daughters themselves expect less parental investment from delaying maturation in a family with a single parent than in a family in which both biological parents are present (Ellis, 2004). These hypotheses interpret the association between father absence and early maturity as the outcome of a conditional strategy of a common genotype. An alternative interpretation is that early maturation of daughters and absence of fathers are genetically correlated (Mendle et al., 2006, 2009).
The helper-at-the-nest scenario predicts earlier menarche in disrupted families, whereas the another-mouth-to-feed scenario predicts a delay in men-arche. Therefore, the association of early menarche with father absence is consistent with older daughters having been selected to delay reproduction to help mothers raise younger sibs when these are likely to be fullsibs but not when these are likely to be halfsibs. The assumptions of the model are simplistic, however. In particular, partner change is assumed to change patri-lineal relatedness to the younger sib but not to affect the form of the fitness functions. Thus, the model does not consider direct effects of father absence on the daughter’s expected fitness or the purported value of father absence as a cue to the quality of the mating market.
The helper-at-the-nest hypothesis is compatible with the effect of father absence being either the expression of a conditional strategy or the result of a genetic correlation. In the first instance, the presence of a girl’s father would be used as a cue to delay maturation. In the second instance, genes that predispose men to short-term relationships would become statistically associated with genes of paternal origin that predispose daughters to mature early, and thus avoid sacrificing personal reproduction for the benefit of maternal
halfsibs. Conversely, genes that predispose men to long-term relationships would become associated with genes that predispose daughters to help parents raise fullsibs.
Early reproduction by elder daughters maximizes the potential for reproductive overlap between mothers and daughters. Either could help the other raise offspring at the expense of personal reproduction. What determines who becomes the helper at whose nest? Cant and Johnstone (2008) have argued that when a young woman moves into the extended family of her husband, her mother-in-law is predisposed to become the helper at the younger woman’s nest because the older women is related to the younger’s offspring (and therefore has a genetic incentive to help), whereas the younger woman is unrelated to the older’s offspring (and therefore has no genetic incentive to help). Perhaps a similar dynamic can play out after divorce between a younger woman and her own mother. In this case, the older woman is symmetrically related to the younger woman’s offspring (she is their maternal grandmother), but genes of paternal origin in the younger woman are unrelated to potential offspring of the older woman. This would create a bias in favor of the older woman helping the younger.
Effects of Birth Order
Elder daughters probably provide most effective help for sibs several years younger than themselves. Therefore, the helper-at-the-nest hypothesis predicts earlier menarche for daughters with fewer younger sibs and later menarche for elder daughters in larger families. A British study found correlations broadly consistent with these predictions: Menarche was delayed in girls from larger families, but girls born later in a family of a given size had earlier menarche (Dann and Roberts, 1993). However, in a Spanish study, first-born daughters had earlier menarche than second-born and third-born daughters but later menarche than fourth-born or higher-born daughters (Apraiz, 1999). Effects of birth order on age at menarche may be complex and highly contingent. The greater competence of elder daughters to provide help is accompanied by a greater ability to compete for limited resources. Elder daughters have also spent early childhood in a smaller family than the family experienced by their younger sibs at the same age.
Expectations about the effects of family size are complicated because larger families contain more competitors for limited resources but also more opportunities for help. Analyses that consider effects of birth order commonly assign lowest birth order to oldest offspring, regardless of family size, and assign higher birth orders to younger sibs. In this formulation, birth order is linearly related to ego’s number of older sibs but provides no information about number of younger sibs (potential beneficiaries of help). From an evo-
lutionary perspective, it might be more informative to perform these analyses with numbers of older and younger sibs as independent predictors.
Effects of Imprinted Genes
Previous sections have explored the hypothesis that mothers (and maternally derived genes of daughters) benefit from delayed maturation because of help provided to mothers by older daughters. The effects of father absence on timing of menarche were construed as supportive of this hypothesis. However, evidence from the effects of imprinted genes on pubertal progression is not readily compatible with the helper-at-the-nest hypothesis.
PWS and Silver-Russell syndrome are caused by the absence of paternally expressed genes or increased dosage of maternally expressed genes. Both syndromes are associated with reduced linear growth in childhood and a weak (or absent) pubertal growth spurt (Davies et al., 1988; Wollmann et al., 1995; Hauffa et al., 2000). These phenotypes suggest matrikin benefited from slower childhood growth. Moreover, individuals who receive both copies of chromosome 14 from their mother experience precocious puberty (Kotzot, 2004). This phenotype suggests that earlier puberty benefited matrikin, perhaps via reduced competition for resources among uterine sibs. Thus, the effects of imprinted genes are more easily reconciled with the another-mouth-to-feed scenario than with the helper-at-the nest scenario.
Given the centrality of age of first reproduction to life-history theory, it is perhaps surprising how little we understand about the fitness tradeoffs that influence variation within and among human populations in age at puberty in either sex. A detailed study of the effects of imprinted genes on pubertal timing and progression promises to provide important clues about the evolution of the distinctive human life history.
Adolescence
Adolescence has been defined as the period from onset of puberty to independence from parents (Casey et al., 2010). The duration of adolescence, by this definition, is highly variable within and among human populations. Popular opinion views adolescence as a time of heightened conflict between parents and offspring and of internal turmoil within the adolescent psyche. Adolescence is both a period of reorganization of neural circuits within the brain (Casey et al., 2010) and a period in which decisions are made about where to live and whom to marry that may have divergent effects on matrilineal and patrilineal inclusive fitness of the child and of his or her parents.
Humans exhibit variation, even within sibships, in the degree to which adults maintain close ties with parents, siblings, and more distant kin. Relations with parents during adolescence are often perceived by young persons as a conflict between their desire for autonomy and parental attempts to control their choices (Surbey, 1998). Parents often perceive the adolescent as self-absorbed and as neglecting responsibilities to family. Adolescence is a life-history transition in which the expression of imprinted genes may have significant effects within the brain. The maternal and paternal genomes of the adolescent agree about individual fitness but may disagree over how much individual fitness should be killed for investment in indirect fitness via kin.
A long tradition in anthropology has emphasized the role of marriage as a form of exchange between patriarchal groups with young women as the commodity of exchange (Tylor, 1889; Durkheim, 1963; Lévi-Strauss, 1980). This was sometimes a direct exchange of daughters between groups, and it sometimes involved a transfer of family wealth, either a payment for a bride or a payment to place a daughter in a favorable situation. The freedom of young people to choose their own partners was curtailed. Conflicts between parents and offspring over the choice of marriage partners are the stuff of legend and literature. Parents usually believe they are acting in their child’s best interests (they believe they have more experience than their child in identifying a suitable spouse), but evolutionary theory recognizes that the genetic interests of parent and offspring may diverge.
Material benefits that a spouse brings to a marriage can be transmitted to affinal kin of the spouse, but genetic benefits are transmitted only to offspring of the marriage. Offspring are therefore expected to place a greater emphasis than parents on the genetic qualities, rather than material resources, provided by mates (Trivers, 1974; Apostolou, 2007a,b; Buunk et al., 2008). Mother and father may disagree over the relative value of material and genetic benefits provided by potential spouses of their child if material benefits flow unequally to matrikin and patrikin. For the same reason, maternal and paternal genomes of the child may disagree about the optimal attributes of a spouse.
The choice of where and with whom to reside may also have important fitness consequences for a young couple. Families can both provide support for personal reproduction and demand support for kin. The expression of imprinted genes within the brain raises the possibility that some of these conflicts, over where to live and who to marry, may be internalized within the adolescent psyche.
DISCUSSION
Life-history theory is concerned with tradeoffs in fitness: between the benefits of muscle and fat, between immune function and reproductive effort, between quantity and quality of offspring, and between reproduction now and reproduction later. Inclusive-fitness tradeoffs may involve the personal fitness of different individuals. Social tradeoffs exist between eating food now or bringing it back to the camp to be shared, between being a dad or a cad, between sponging on mother’s kin or father’s kin, and between helping one’s mother raise sibs or having a child of one’s own. Psychology is concerned with tradeoffs in mental function: between immediate and delayed gratification, between empathizing and systemizing, between focused and diffuse attention, and between impulsiveness and executive control. A key challenge for a synthesis of these fields will be to understand how psychological tradeoffs mediate life-history tradeoffs.
Our species’ innate taxonomy of kin is defined by evolved dispositions that are directed toward some individuals but not others based on environmental cues that are correlated with degree of relatedness. An innate disposition defines the membership of a category, and the membership defines the coefficient of relatedness associated with the category. All individuals who evoke a disposition belong to the category, and all members of the category determine the relatedness associated with fitness consequences of the disposition. Thus, innate kin categories need not correspond exactly to genealogical categories, and, given enough time and genetic variation, the cultural categorization of kin can shape our innate dispositions.
An unresolved issue is the richness of our innate categorization of kin both in terms of the number of different kinds of kin for whom we have distinct dispositions and in terms of the complexity of dispositions toward each particular category. No one would seriously argue that innate structure is absent in our interactions with mothers, but there is no similar consensus over whether we innately distinguish fullsibs from halfsibs, let alone mother’s brother’s daughters from father’s sister’s daughters.
When tradeoffs exist between the individual fitnesses of relatives, inclusive fitness assigns relative values to effects on different categories of kin based on each category’s degree of relatedness to an actor. For most categories of kin, relatedness differs for genes of maternal and paternal origin. The inclusive fitness of maternal and paternal alleles will be maximized by different allocations of fitness among kin, creating the potential for conflicting goals within individual organisms and a deep-seated biological ambivalence in relations among kin.
Maternal and paternal genes have a common interest in the effective functioning of the individual actor, but phenotypes that are determined by agents with different fitness functions are not expected to show the degree of integration and physiological efficiency one would expect of a phenotype determined by agents with identical interests. Perhaps such internal conflicts can partially account for inefficiencies of mental function and a high frequency of pathology in human social interactions.
ACKNOWLEDGMENTS
The paper has benefited from the comments of Bernard Crespi, Edgar Dueñez-Guzman, Sarah Hrdy, Karen Kramer, David Queller, Stephen Stearns, Robert Trivers, and two anonymous reviewers. The work was supported by a Collaborative Innovation Award from the Howard Hughes Medical Institute to Catherine Dulac.






















