
![]()
In cooperatively breeding systems in which some individuals delay reproduction to help raise others’ offspring, environmental variation in space and time influences individual reproductive strategies as well as interspecific patterns of sociality. Although most environmental explanations for cooperative breeding emphasize the mean fitness gains of living socially, the fittest individuals are not always those that produce on average the highest number of offspring. At times, variance in fecundity can influence fitness as much as mean fecundity, particularly in small populations like those of cooperative breeders. Cooperative breeding behavior could therefore be a risk-averse strategy to maximize fitness by reducing environmentally induced fecundity variance. Such a within-generation bet-hedging hypothesis for social evolution predicts that (i) variance in reproductive success should be related to environmental variation, (ii) variance in reproductive success should be related to the potential for cooperation in a group, and (iii) the potential for cooperation should be related to environmental variation. Using data from a 10-year study of cooperatively breeding superb starlings (Lamprotornis superbus) living in a temporally and spatially variable savanna ecosystem, I found that variance in reproductive success declined with increasing environmental quality (temporal variation), increasing territory quality (spatial variation), and increasing group size (potential for cooperation), which is itself
_______________
Department of Ecology, Evolution and Environmental Biology, Columbia University, New York, NY 10027. E-mail: dr2497@columbia.edu.
related to environmental variation. To understand the adaptive value of cooperative breeding behavior in variable environments, researchers must consider both mean and environmentally induced variance in fecundity. Determining how spatiotemporal environmental variation drives risk-averse strategies may provide insights into the evolution of complex social behavior.
Kin selection, or reproductive strategies that favor an organisms’ relatives, is often invoked to explain the evolution of cooperation and the formation of complex animal societies (Hamilton, 1964a; West-Eberhard, 1975). In cooperatively breeding systems in which some individuals delay independent breeding to help raise the offspring of others, the inclusive fitness benefits of helping genetic relatives may outweigh the potential costs of trying to breed independently (Brown, 1987). Recent theoretical (Boomsma, 2007, 2009) and comparative work in both invertebrates (Hughes et al., 2008) and vertebrates (Cornwallis et al., 2010) suggests that high relatedness among group members may be critical to the evolution of complex animal societies. However, despite renewed interest in determining how genetic relatedness among group members can influence social interactions and the evolution of family groups (Boomsma, 2007, 2009; Hughes et al., 2008; Hatchwell, 2009; Nam et al., 2010; Sharp et al., 2011), relatedness alone cannot explain why some individuals in a group breed whereas others do not, or why some species breed cooperatively whereas other closely related ones do not. In other words, relatedness may set the stage for cooperation in animal societies, but it is not sufficient to explain many individual differences in reproductive strategies or interspecific patterns of social diversity (Rubenstein and Lovette, 2007; Jetz and Rubenstein, 2011).
Environmental factors are known to influence complex vertebrate social behavior (Alexander, 1974; Jarman, 1974), as well as explain many of the individual differences in reproductive strategies (Emlen, 1982a; Komdeur, 1992; Covas et al., 2004; Rubenstein, 2007a) and interspecific patterns of sociality (Rubenstein and Lovette, 2007; Jetz and Rubenstein, 2011). The role of environmental factors in shaping animal societies is central to the ecological constraints hypothesis (Emlen, 1982a), which argues that when barriers to dispersal are high, offspring will be selected to delay dispersal and remain at home as part of a group because the probability of reproducing successfully outside the group is low. The ecological constraints hypothesis (Emlen, 1982a) and its other derivations (Koenig and Pitelka, 1981; Koenig et al., 1992) predict the environmental conditions under which delayed dispersal is likely to occur (Hatchwell and Komdeur, 2000). These conditions include a shortage of vacant breeding territories (i.e., habitat saturation), the costs
of dispersal, difficulties in finding a mate, and a low chance of successful reproduction once a territory is established [reviewed in Hatchwell and Komdeur (2000)]. Although each of these conditions represents an external constraint or cost associated with dispersal and independent breeding, such conditions represent only one side of the cost–benefit equation of social living. An alternative theory, the benefits of philopatry hypothesis, instead argues that delayed dispersal is the result of intrinsic benefits gained by remaining on the natal territory (Stacey and Ligon, 1987, 1991). These benefits include enhanced survival, indirect fitness gains from helping relatives, opportunities for obtaining a nearby breeding vacancy in the future, and the chance to inherit the natal breeding territory itself [reviewed in Cockburn (1998)]. Despite much initial debate over the relative importance of these two hypotheses in shaping cooperative groups (Emlen, 1994), it is now widely accepted that they are more similar than they are dissimilar, because they place different emphasis on the costs of dispersing vs. the benefits of not dispersing as a result of environmental constraints (Emlen, 1994, 1997a; Hatchwell, 2009).
Nearly all of the early environmental hypotheses for cooperative breeding behavior have focused primarily on the costs and benefits associated with breeding on territories of varying quality, or the fitness consequences of living in a spatially heterogeneous landscape where suitable territories are limiting (Emlen, 1982a; Stacey and Ligon, 1991; Koenig et al., 1992). However, spatial constraints on dispersal (i.e., habitat heterogeneity) are not the only form of environmental variation that can influence cooperative breeding behavior. Environmental variation in time can also influence social behavior, including dispersal decisions and the adoption of different breeding roles. Although the ecological constraints hypothesis is generally used to emphasize the role of habitat heterogeneity in influencing dispersal decisions, its original description also recognized the importance of environmental unpredictability in driving cooperative breeding (Emlen, 1982a), although this idea went largely untested for decades [but see Curry (1989) and Curry and Grant (1990)]. Recent work in cooperatively breeding birds living in unpredictable environments suggests that erratic and variable climatic patterns can also influence social complexity. Temporal environmental variation resulting from unpredictable patterns of rainfall (i.e., climatic uncertainty) has been shown to influence not only individual behavioral decisions and reproductive roles (MacColl and Hatchwell, 2002; Canario et al., 2004; Rubenstein, 2007a; Covas et al., 2008) but also interspecific patterns of sociality on continental and global scales (Rubenstein and Lovette, 2007; Jetz and Rubenstein, 2011). Together, spatial and temporal patterns of environmental variation explain many of the individual-level coopera-
tively breeding behaviors, as well as the broadscale interspecific patterns of social diversity.
Although the ecological constraints and benefits of philopatry hypotheses are said to be qualitatively similar (Emlen, 1994), one key difference seems to have been largely overlooked. Ecological constraint hypotheses primarily focus on mean reproductive success (Koenig and Pitelka, 1981; Emlen, 1982b; Stacey and Ligon, 1987; Koenig et al., 1992), emphasizing the average number of young produced when breeding alone or as part of a group. In contrast, the benefits of philopatry hypothesis were formulated on the idea of variance in reproductive success and emphasized variation in young produced on occupied territories through time in cooperative and noncooperative species (Stacey and Ligon, 1991). This key difference in fitness measures (i.e., mean vs. variance in fecundity) has important consequences for understanding how natural selection acts to promote cooperative behavior. Gillespie (1974, 1975, 1977) demonstrated that the fittest individuals are not always those that produce on average the highest number of offspring. Instead, he showed that in small populations, variance in fecundity can determine fitness as much as mean fecundity because the intensity of selection on reducing fecundity variance is inversely proportional to population size (Gillespie, 1974). Integrating these bet-hedging ideas into an inclusive fitness game theoretic framework, Lehmann and Balloux (2007) showed that helping behavior is selected for when fecundity variance is high. Thus, the simultaneous examination of mean offspring production and variance in offspring production [i.e., considering helping behavior and cooperative breeding as a bet-hedging strategy (Cockburn and Russell, 2011)] may shed light on the evolution of cooperative breeding behavior, particularly as it relates to spatiotemporal environmental variation.
To understand the role of fitness optimization in the evolution of cooperatively breeding behavioral phenotypes (i.e., breeding roles) in variable environments, we must consider the concept of bet-hedging, or risk aversion. Population geneticists have long understood that fluctuating selection resulting from environmental variability can favor the evolution of risk-averse strategies (Gillespie, 1974, 1975, 1977; Frank and Slatkin, 1990). Bet-hedging itself can be traced back more than 250 years to Bernoulli (1954; Stearns, 2000). In an evolutionary sense, bet-hedging strategies generally spread risk over multiple generations (i.e., years) by reducing variance in offspring production, which ultimately leads to an increase in the geometric mean lifetime reproductive success, but often a reduction in the arithmetic mean (Philippi and Seger, 1989). Although much rarer than these among-generation bet-hedging strategies, risk aversion can also operate within generations (Hopper et al., 2003). Within-generation bet-hedging spreads risk within
a single generation and involves variability in the selection pressures to which a phenotype is exposed (Hopper et al., 2003). Importantly, within-generation bet-hedging encompasses any behavioral strategy that avoids having no or few offspring in any given generation, rather than maximizing the expected number of offspring (Sarhan and Kokko, 2007). Examples of within-generation bet-hedging are rarer than those of among-generation bet-hedging because within-generation bet-hedging only evolves under a much narrower set of demographic conditions (Hopper et al., 2003). Specifically, within-generation bet-hedging is only likely to evolve in small populations because the intensity of selection on reducing fecundity variance is inversely proportional to population size (Gillespie, 1974). For cooperatively breeding species in which populations are subdivided into kin-based social groups that are connected via dispersal, the conditions for within-generation bet-hedging to evolve are likely to exist (Lehmann and Balloux, 2007; Shpak, 2005). Such within-generation bet-hedging strategies could apply in any cooperatively breeding species in which, in addition to opportunities for helping, subordinates have options for direct reproduction, either by dispersing to breed independently outside of the group, becoming a breeder in the natal group, or gaining reproduction through extrapair paternity. Within-generation bet-hedging strategies to avoid having no or few offspring in any given generation may be most evident in cases of redirected helping in species like the long-tailed tit (Aegithalos caudatus), in which temporally variable ecological constraints drive individuals to switch from independent breeding to helping others (often relatives) later in the breeding season (MacColl and Hatchwell, 2002; Hatchwell and Sharp, 2006). Thus, when individuals have simultaneous opportunity to accrue fitness directly and indirectly within a given breeding season, environmentally induced selection to reduce fecundity variance can operate on risk-averse breeding strategies.
Although the conditions under which variance in fecundity can influence kin structure and cooperative breeding behavior have been modeled (Lehmann and Balloux, 2007), to my knowledge they have not been studied empirically. Here, I will study the effects of spatial and temporal environmental variation on fecundity variance in an avian cooperative breeder. Specifically, I will examine how mean and variance in group reproductive success change with increasing potential for cooperation, and how the potential for cooperation relates to environmental variation. I will test the hypothesis that cooperative breeding behavior is a risk-averse strategy to maximize fitness by reducing environmentally induced variance in fecundity. Such a within-generation bet-hedging hypothesis for social evolution predicts that (i) variance in reproductive success should be related to environmental variation in space and/
or time, (ii) variance in reproductive success should be related to the number of helpers in a group, and (iii) the numbers of helpers in the group should be related to environmental variation. Having helpers at the nest has been proposed to reduce the risk of complete clutch failure within a breeding season, either by preventing nestling starvation or depredation during harsh conditions, and/or by allowing for more clutches to be laid during benign conditions, resulting in an extended breeding season (Rubenstein and Lovette, 2007). Moreover, parental care that positively influences offspring survival may be favored in variable environments (Bonsall and Klug, 2011), and delayed reproduction strategies in general may be favored in unpredictable environments (Koons et al., 2008). Therefore, cooperative breeding itself may be a risk-averse strategy to maximize fitness by reducing variance in the number of offspring produced in a social group. Using data from a 10-year study of cooperatively breeding superb starlings (Lamprotornis superbus) living in a temporally and spatially variable savanna ecosystem (Rubenstein, 2009), I will explore how environmental variability in space (habitat heterogeneity) and time (climatic uncertainty) can directly influence fecundity variance. Moreover, I will examine whether fecundity variance relates to the potential for cooperation (number of helpers in the group), which may itself be related to environmental variability. Thus, this study will examine how spatiotemporal environmental variation influences fitness in cooperatively breeding birds living in unpredictable and heterogeneous environments, thereby providing insights into risk-averse social behavior and the evolution of complex animal societies.
RESULTS
To integrate spatial and temporal environmental variation into a framework for understanding the evolution of complex animal societies, I examined the environmental correlates of reproductive success in the cooperatively breeding superb starling using data from a 10-year field study representing 20 breeding seasons; birds typically breed twice a year during both the long and short rains. Superb starlings are endemic to the savanna of East Africa, which like most semiarid ecosystems is a spatially and temporally variable environment (Rubenstein, 2009). Superb starlings are obligate plural cooperative breeders, meaning that all groups have helpers and multiple breeding pairs that nest separately. They live in spatially subdivided populations (hereafter social groups) with high kin structure (Rubenstein, 2007c), thereby meeting the primary criteria to empirically examine within-generation bet-hedging strategies (Lehmann and Balloux, 2007). Because superb starlings live in such complex social groups with multiple breeding pairs, I quantified
mean and variance in reproductive success at the level of the group as the proportion of eggs laid that fledged young in each nest for each group in each breeding season. This assumes that selection can operate on both the direct and indirect components of fitness, as has been shown theoretically in these types of kin-structured groups (Shpak, 2005; Lehmann and Balloux, 2007).
Rainfall in this region of Kenya is extremely variable from month to month and year to year. From 1998 through 2010, rainfall generally peaked three times per year; there were large peaks during both the short (November) and long (April–May) rainy seasons, as well as a small peak during July and August (Fig. 4.1A). The mean ± SD in annual rainfall was 529 ± 138 mm, which is characteristic of semiarid ecosystems worldwide (Austin et al., 2004). Annual rainfall ranged from 280 mm in 2000 to 822 mm in 2010. There was a negative relationship between mean monthly rainfall and the coefficient of variation in mean monthly rainfall (correlation: F1,10 = 19.51, P = 0.0013, r = 0.81; Fig. 4.1B), showing that the drier months during the prebreeding season were more unpredictable than the wetter months during the breeding season. Thus, there was both high within-year (i.e., seasonality) and among-year variation (i.e., temporal variability) in rainfall in this unpredictable environment.
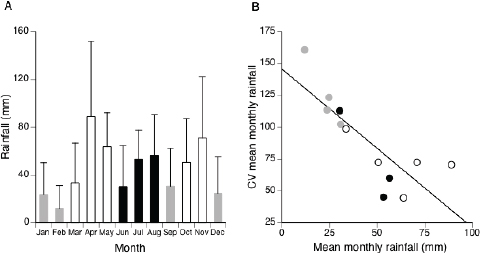
FIGURE 4.1 Rainfall patterns at the Mpala Research Centre, Laikipia, Kenya, from 1998 to 2010. (A) Mean ± SD in monthly rainfall is plotted. Peaks in rainfall were trimodal, highlighting the long and short rainy seasons, as well as a third peak in July and August. White bars indicate the primary breeding months, whereas graybars indicate the primary dry season months. (B) Mean monthly rainfall was negatively correlated with the coefficient of variation (CV) in mean monthly rainfall, showing that drier months were more variable than wetter months.
The high among- and within-year variation in rainfall influenced territory quality: There were significant differences among territories and among months in vegetation cover (generalized linear mixed model: territory, F6,1622 = 22.85, P < 0.0001; month, F11,1563 = 4.24; P < 0.0001). Vegetation cover is highly correlated with insect abundance and is thus a strong indicator of territory quality and a correlate of reproductive behavior (Rubenstein, 2007c). However, there was no effect of the interaction between territory and month on vegetation cover, suggesting that relative territory quality does not change much in this ecosystem (generalized linear mixed model: territory month, F66,1623 = 0.86; P = 0.78). In other words, although habitat quality differs among territories, high-quality territories remain better relative to low-quality territories in all months, seasons, and years.
Although the overall mean annual reproductive success was low in this population, as only 13% of all eggs laid fledged, there was significant variation in reproductive success among years (Wilcoxon test: X2 = 21.26, df = 9, P = 0.012) and among territories (Wilcoxon test: X2 = 17.31, df = 8, P = 0.027). However, mean fecundity was not related to variation in breeding conditions through time (i.e., climatic uncertainty or habitat heterogeneity); there was no relationship between mean reproductive success and breeding season rainfall (regression: F1,7 = 0.073, P = 0.79, R2= 0.004; Fig. 4.2A) or vegetation cover (regression: F1,5 = 0.012,
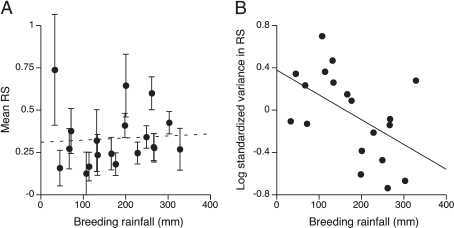
FIGURE 4.2 Reproductive success and climatic uncertainty. Reproductive success (RS) was estimated as the proportion of eggs fledged in each nest averaged for each group. Standardized variance in reproductive success was calculated as (variance in reproductive success)/(mean reproductive success)2. Each point represents a breeding season (n = 19). (A) Mean ± SE reproductive success did not vary with breeding season rainfall, but (B) standardized variance in reproductive success was negatively related to breeding rainfall. Thus, fecundity variance decreased with increasing environmental quality or conditions.
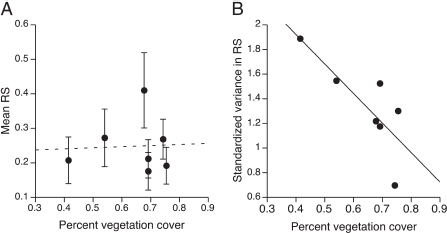
FIGURE 4.3 Reproductive success and habitat heterogeneity. Reproductive success (RS) was estimated as the proportion of eggs fledged in each nest averaged for each group. Standardized variance in reproductive success was calculated as (variance in reproductive success)/(mean reproductive success)2. Each point represents a territory or group (n = 7). (A) Mean ± SE reproductive success did not vary with percentage vegetation cover, but (B) standardized variance in reproductive success was negatively related to percentage vegetation cover. Thus, fecundity variance decreased with increasing territory quality.
P = 0.92, R2 = 0.002; Fig. 4.3A). In contrast, the variance in fecundity in time and space was related to environmental variation among territories and across years. There was a significant negative relationship between standardized variance in reproductive success and breeding season rainfall (regression: F1,17 = 6.17, P = 0.024, R2= 0.27; Fig. 4.2B), suggesting that fecundity variance among territories declines with increasing environmental quality or conditions. There was also a negative relationship between standardized variance in reproductive success and vegetation cover (regression: F1,5= 8.42, P = 0.034, R2 = 0.63; Fig. 4.3B), suggesting that fecundity variance among years declines with increasing territory quality. Together these results show that environmental variation in space and time had significant effects on variance in fecundity but not on mean fecundity.
Previous work in this system demonstrated that having helpers is critical, as nests with more helpers fledged more young (Rubenstein, 2007b). However, given the relationship between variance in reproductive success and environmental variability, does having more helpers actually increase mean reproductive success and reduce variance in reproductive success, and does helper number vary as a function of environmental variation? Using group size as an estimate of the potential
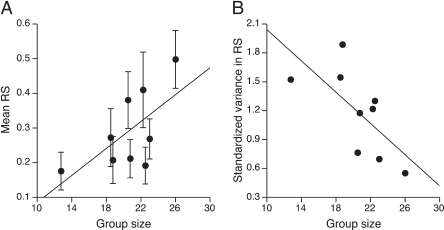
FIGURE 4.4 Reproductive success and helper number. Reproductive success (RS) was estimated as the proportion of eggs fledged in each nest averaged for each group. Standardized variance in reproductive success was calculated as (variance in reproductive success)/(mean reproductive success)2. Because all superb starling groups have helpers, group size is a good estimate of the number of helpers in a group (Rubenstein, 2007b; Rubenstein and Shen, 2009). Each point represents a territory or group (n = 9) (A) Mean ± SE reproductive success showed a nonsignificant trend to increase with group size, whereas (B) standardized variance in reproductive success was negatively related to group size. Thus, mean fecundity tended to increase with increasing numbers of helpers, whereas fecundity variance decreased with increasing numbers of helpers.
number of helpers available in a group (Rubenstein, 2007b; Rubenstein and Shen, 2009), I found that mean reproductive success showed a trend to increase with increasing group size (regression: F1,7= 4.82, P = 0.064, R2 = 0.41; Fig. 4.4A), whereas standardized variance in reproductive success declined with increasing group size (regression: F1,7= 6.09, P = 0.043, R2= 0.47; Fig. 4.4B). Additionally, group size was related to environmental variation in time but not in space. Group size, which was estimated during the long rains breeding season, was not influenced by vegetation cover (regression: F1,5= 0.48, P = 0.52, R2= 0.087) or breeding rainfall (regression: F1,2= 1.15, P = 0.40, R2= 0.36). However, just as many reproductive behaviors and components of superb starling physiology are influenced by rainfall in the prebreeding period leading up to the long rains breeding season (Rubenstein, 2007a,b; Rubenstein et al., 2008), so too is group size related to prebreeding rainfall (regression: F1,2 = 27.32, P = 0.035, R2= 0.93); groups were larger after relatively wetter dry seasons, suggesting that more helpers were available after favorable dry season conditions. Thus, fecundity (mean and variance) is related to
the potential for cooperation, which is itself related to a different seasonal component of temporal environmental variation.
DISCUSSION
Having helpers at the nest is beneficial for superb starlings, as nests with more helpers fledge more young (Rubenstein, 2007b). Here, I further demonstrate that mean reproductive success increased with increasing group size [i.e., the number of available helpers (Rubenstein, 2007b; Rubenstein and Shen, 2009)] and that variance in reproductive success decreased with increasing group size. Group size was also directly related to prebreeding rainfall or temporal environmental variation in the dry season immediately before the primary breeding season. Rainfall during this period not only influences the potential for cooperation but also breeding behavior directly (Rubenstein, 2007a,b), as well as stress physiology (Rubenstein, 2007a) and immune function (Rubenstein et al., 2008). These results suggest that not only is having helpers beneficial but also that living in larger groups has added reproductive benefits. Thus, cooperative breeding and the formation of large, complex family groups in superb starlings may be related directly to environmental variation.
If cooperative breeding behavior within these large family groups is indeed a within-generation bet-hedging strategy to maximize fitness by reducing fecundity variance in spatially heterogeneous or temporally unpredictable environments, then variance in reproductive success should also be related to environmental variation in space and/ or time. In support of this prediction, I found that reproductive success varied greatly among years and among territories, and that the variance in reproductive success was related to both climatic uncertainty and habitat heterogeneity. Variance in reproductive success among territories decreased with increasing environmental conditions across years (breeding season rainfall), whereas variance in reproductive success among years decreased with increasing territory quality (vegetation cover). Thus, mean reproductive success is similar in both good and bad times, as well as on high- and low-quality territories. As would be predicted from a bet-hedging hypothesis, constant levels of mean reproductive success are maintained across all environmental conditions, and importantly, reproductive success does not decrease as conditions deteriorate. Poor rainfall years, however, seem to exacerbate the differences between high- and low-quality territories, leading to greater variance in reproductive success among territories. Conversely, high rainfall can apparently mask the inherent differences in territory quality that drive patterns in reproductive success. Similarly, low-quality territories amplify the consequences of annual differences in breeding rainfall more than high-quality territories.
Together, these results are consistent with the hypothesis that cooperative breeding in starlings may be a risk-averse or within-generation bet-hedging strategy to maximize fitness by minimizing variance in fecundity in temporally and spatially variable environments. Additionally, these results are also consistent with both the ecological constraints and benefits of philopatry hypotheses. Ecological constraints clearly limit breeding opportunities in this species (Rubenstein, 2007a,c), but fecundity variance also seems to play an important role in shaping individual reproductive decisions. Additional studies in cooperatively breeding species in which subordinates have greater opportunities for independent breeding outside of the group will be needed to further disentangle these hypotheses. If cooperative breeding is a strategy to reduce risk in variable environments, do spatially and temporally variable environments influence social behavior in similar ways, as has been proposed previously (Emlen, 1982a,b), or do the mechanisms underlying group formation differ in the different types of environments? Insider-outsider conflict theory (Giraldeau and Caraco, 1993; Higashi and Yamamura, 1993) provides a possible framework to explore how the tension over group membership between current group members (i.e., insiders) and potential joiners (i.e., outsiders) could differ in temporally and spatially variable environments. Insider-outsider conflict theory has recently been expanded to consider conflict resolution during group formation in cooperatively breeding species in which relatedness among group members can be high (Shen and Emlen, 2010). Instead of focusing on just offspring delayed dispersal, the theory emphasizes the importance of simultaneously considering both insider (parents) and outsider (joining mature offspring) interests to fully understand the evolution of cooperative breeding. In temporally variable environments where breeding conditions are unpredictable from year to year, insiders may be more likely to allow outsiders into the group to maintain a pool of available helpers as a form of insurance (Emlen, 1982a; Covas et al., 2004, 2008; Rubenstein, 2007a). However, outsiders may be less inclined to join groups in all but the poorest years without insider concessions (i.e., a share of reproduction) because outside breeding opportunities are likely to be available (Shen and Emlen, 2010). This environmentally induced conflict would not only lead to the formation of larger groups (i.e., a larger insurance pool) but also potentially to greater reproductive sharing within groups or lower reproductive skew. Once groups have formed, year-to-year environmental differences could still influence reproductive conflict and the degree of reproductive skew (Emlen, 1982a; Covas et al., 2004). Thus, temporally variable environments might favor large groups with multiple breeders and low reproductive skew (i.e., plural breeding), in which outsiders exert relatively greater control over group membership and reproductive
conflict is high. In spatially variable, heterogeneous environments where territory quality varies across the landscape, the conditions on a given territory are more predictable from year to year than in temporally variable environments. Therefore, insiders might not only be less willing to accept outsiders into the group but also less inclined to share any reproduction with them. In contrast, outsiders may be more willing to join groups without concessions because outside breeding opportunities are likely to be limiting in all years. Thus, spatially variable environments might favor smaller groups with one breeding pair and high reproductive skew (i.e., singular breeding), in which insiders might exert relatively greater control over group membership and reproductive conflict is low. Overall, the mechanisms underlying group formation, the individuals that control group membership, and the types of social groups themselves (i.e., singular vs. plural) may differ in temporally and spatially variable environments.
Although the data presented here are consistent with the hypothesis that spatiotemporal environmental variation promotes cooperative breeding as a risk-averse behavioral coping strategy, it is not the only possible explanation. Kin-structured populations, or kin neighborhoods, resulting from environmental constraints could also influence avian cooperative breeding behavior (Hatchwell, 2009). Hamilton (1964b) was the first to realize that populations with limited dispersal, or population viscosity, will lead to greater opportunities for kin to interact. This idea, namely that reduced dispersal can lead to cooperation among relatives, forms the basis of most ecological constraints models of cooperative breeding (Emlen, 1982a; Koenig et al., 1992; Covas and Griesser, 2007), which are based largely on ideas of spatial variation in the environment. In general, habitat heterogeneity leads to reduced dispersal opportunities and therefore greater natal philopatry and the formation of kin neighborhoods that ultimately may give rise to kin groups. However, kin neighborhoods could also result from high temporal environmental variation, independent of processes like population viscosity. Variation in reproductive success, which could be driven largely by climatic uncertainty, is predicted to lead to an increase in the relatedness between group members because it decreases the number of effective relatives within a group (Lehmann and Balloux, 2007). Climatic uncertainty could therefore influence the formation of kin neighborhoods and ultimately kin groups, which is consistent with comparative results showing that cooperatively breeding species tend to live in temporally variable environments (Rubenstein and Lovette, 2007; Jetz and Rubenstein, 2011). Thus, spatial and temporal environmental variation can both influence demographic structure and the formation of kin neighborhoods, but for different reasons. Habitat heterogeneity could lead to kin-structured
populations via spatial constraints on dispersal, whereas climatic uncertainty could lead to higher relatedness within groups because of decreased offspring production.
Whereas most theoretical and empirical studies examining the role of environmental constraints in the evolution of cooperative breeding have focused on mean reproductive success (Emlen, 1982b; Koenig et al., 1992), the results presented here suggest that we should also consider variance in reproductive success when studying social evolution. In superb starlings and other birds, cooperative breeding may be a risk-averse strategy to maximize fitness in a range of environmental conditions by reducing fecundity variance. In particular, when mean reproductive success does not differ between high- and low-quality territories, or between good and bad years, selection on variance may be important. Selection on variance is maintained when group sizes remain small and/ or when dispersal rates are low (Lehmann and Balloux, 2007), both of which are hallmarks of cooperative breeders and especially likely to occur in temporally or spatially variable environments. However, spatial and temporal environment variation may influence cooperative breeding behavior in different ways. The mechanisms underlying group formation, the individuals that control group membership, and the types of social groups themselves may differ in temporally and spatially variable environments. Thus, to understand the adaptive value of cooperative breeding behavior in the heterogeneous and unpredictable environments where social species disproportionately occur (Jetz and Rubenstein, 2011), researchers must consider both the mean and environmentally induced variance in reproductive success. In addition to the within-generation bet-hedging hypothesis tested here, researchers should also consider social evolution in the context of among-generation bet-hedging, or risk spreading over multiple generations to maximize geometric mean lifetime reproductive success. Finally, the ideas presented here are not limited to birds: many species of social mammals (Solomon and French, 1997) and insects (Wilson, 1971; Costa, 2006) live in temporally variable environments or habitats where resources are distributed heteroge-neously on the landscape. Ultimately, determining how spatiotemporal environmental variation drives patterns of and variation in fitness will provide important insights into the evolution of complex social behavior in a diversity of animal taxa.
MATERIALS AND METHODS
Study System and Species
A marked population of superb starlings was continuously monitored at the Mpala Research Centre, Laikipia, Kenya (0°17’ N, 37°52’ E) from
April 2001 through January 2011. Breeding activities of seven social groups were monitored over 10 long-rains and 10 short-rains breeding periods during this time. One additional group was added in January 2002, and another was added in January 2003; both were monitored through January 2011. Although birds have been recorded breeding during every month of the year, they typically only breed during both the long (April–May) and short (November) rains. Group size, which is a strong predictor of the number of available helpers (Rubenstein, 2007b; Rubenstein and Shen, 2009), was estimated annually for all groups from 2002 to 2005 during the long-rains breeding season. Active nests were checked every 1–3 days throughout the study during the hatching and nestling stages. Group reproductive success was quantified as the proportion of eggs laid that fledged young in each nest for each group in each breeding season. When nests were first encountered in the incubation stage and the number of eggs laid could not be determined, we used the mean clutch size of 3.5 eggs. When pairs had multiple clutches of eggs in a breeding period, the total number of eggs laid and fledged was summed before a proportion was calculated. Raw proportional data for each breeding pair were arcsine-square root transformed, and arithmetic means and SDs were used for all analyses, consistent with analyses of within-generation bet hedging (Gillespie, 1974, 1975; Crean and Marshall, 2009). Standardized variance in reproductive success was calculated as (variance in reproductive success)/(mean reproductive success)2 (Weatherhead and Boag, 1997; Rubenstein, 2007b). All statistical tests were conducted in JMP v9 (SAS Institute, 2010). We used nonparametric Wilcoxon tests to examine differences in reproductive success among years and among territories. Regressions were used to examine the relationships between reproductive success (mean and variance) and climatic uncertainty (breeding rainfall) and habitat heterogeneity (vegetation cover). When necessary, summary data were logarithm transformed to meet assumptions of normality. Data from the 2006 short rains were excluded from some analyses because only 1 of 40 nests (1 of 124 eggs) fledged young, which greatly skewed the standardized variance estimates.
Rainfall
Daily rainfall data were collected continuously from 1998 through 2009 using an automated Hydrological Services TB3 Tipping Bucket Rain Gauge located at the Centre. In 2010, rainfall data were collected using a manual gauge located at the same place. Because the two data-sets were highly correlated in previous years (F1,138 = 2577.84, P < 0.0001, R2= 0.97), we used the automated data from 2001 through 2009 supplemented with the 2009 manual data. Breeding rainfall was calculated
as the total monthly rainfall for all months in which nests were initiated during a breeding season. Prebreeding rainfall was calculated as the total rainfall during December, January, and February each year (Rubenstein, 2007a,b; Rubenstein et al., 2008). This period represents the primary dry season and the 3 months with the greatest coefficients of variation in mean monthly rainfall (Fig. 4.1).
Habitat Quality
Territory quality was previously quantified using vegetation transect surveys conducted on seven territories at a single time point [see Methods in Rubenstein (2007c)]. Two groups were not surveyed because their territories encompassed areas at the Centre where constant human habitation and building, continuous livestock rearing, and seasonal grass and tree cutting altered the landscape and provided year-round food and water for birds. Briefly, seven 100-m transects starting at a randomly chosen point and compass direction were conducted at each site. For each 100-m transect, a metal pin was dropped every 2 m (50 points per transect, and 350 points per territory), and whether the pin was touching vegetation or bare earth was recorded (Holmes, 1974; Stewart et al., 2001). Territory quality was quantified for each transect as the percentage vegetation cover, or the proportion of pins that were touching a grass or forb species over the total number of pin drops per transect. Vegetation cover on each territory was positively correlated with the proportion of Cynadon grass, the dominant grass species in glades and an indicator of nutrient-rich soils (Augustine, 2003), and the abundance of grasshoppers, a primary food source for nestlings (Rubenstein, 2007c). Although previous work on this landscape showed that relative habitat quality does not change even though vegetation cover varies seasonally (Rubenstein, 2007c), monthly vegetation transects were conducted on all seven territories from February 2008 through January 2011 (36 months) using the same methods detailed above to confirm that relative habitat quality of superb starling territories does not change. Raw proportional data were arcsine-square root transformed and analyzed using a generalized linear mixed model with year and transect as random effects to account for repeated sampling of territories, as well as month, territory, and their interaction as fixed effects.
ACKNOWLEDGMENTS
I thank F. Ayala, J. Avise, D. Queller, and J. Strassmann for organizing the Sackler Colloquium and inviting me to participate; J. Ekiru, F. Lomojo, G. Rana, J. Ronjore, B. McCleery, T. Davis, and especially W. Watetu and G.
Manyaas for their help in the field; and J. Avise, S. Emlen, P. Sherman, and especially C. Botero, S.-F. Shen, and D. I. Rubenstein for providing discussion and/or comments on earlier versions of the manuscript. The Kenyan Ministry of Education, Science and Technology, the National Council on Science and Technology, the National Museums of Kenya Ornithology Section, the Kenya Wildlife Service, and the Mpala Research Centre enabled this work. D.R.R.’s long-term research was supported by Columbia University, the Howard Hughes Medical Institute, the Smithsonian Institution, Cornell University, the Miller Institute for Basic Research at the University of California at Berkeley, and the Museum of Vertebrate Zoology, as well as by grants from the National Science Foundation (IBN-407713), the American Museum of Natural History Chapman Fund, the American Ornithologists’ Union, the Wilson Ornithological Society, the Society for Integrative and Comparative Biology, the Animal Behaviour Society, the Harvard Travellers Club, the Society of Sigma Xi, Cornell University, the Cornell Laboratory of Ornithology Benning Fund, and Cornell Sigma Xi.
This page intentionally left blank.


















