
Ecosystem Trajectories
Affected by Water Quality and Quantity
Previous National Research Council (NRC) reports on Everglades restoration noted that progress had not yet occurred (NRC, 2007) or that it was slow (NRC, 2008, 2010) and emphasized that tangible restoration progress is needed to prevent irreversible ecosystem declines. Such declines result from disruptions in hydrologic and water quality conditions that have been so altered from their natural states that the ecological conditions in the remnant Everglades have departed ever further from the target conditions envisioned in the restoration plan. As noted in Chapter 3, restoration initiatives have focused mainly on the perimeter of the Everglades with little benefit to the remnant Everglades. The latest (August 2011) Integrated Delivery Schedule (IDS; see Figure 3-1) gives little cause for optimism, because the bulk of the Water Conservation Area (WCA)-3 Decompartmentalization and Sheet Flow Enhancement (Decomp) project has been delayed beyond 2020. Recent state budget cuts threaten to slow restoration progress even further. In light of the ongoing declines and the slow pace of restoration progress, NRC (2010) recommended “a rigorous scientific analysis of the short- and long-term tradeoffs between water quality and quantity for the Everglades ecosystem.”
In this chapter, the committee explores recent trends, possible future trajectories, and timescales for recovery of 10 ecosystem attributes of the remnant Everglades to better understand the implications of the current slow pace of progress and the potential consequences of focusing on water quality at the expense of water quantity, or vice versa. The chapter is organized in four main sections. First, the committee examines the context for water quality and quantity issues and discusses instances when water quality concerns have delayed restoration progress or have the potential to impact the future pace of implementation. Second, the committee synthesizes its analysis of current conditions and trajectories for 10 ecosystem attributes under three generic restoration scenarios to provide insights on how ecosystem attributes might respond differentially to management efforts. Furthermore, the committee explains the tradeoffs involved
when restoration efforts focus on only water quality or water quantity. Third, the committee discusses each of the 10 attributes considered in the condition and trajectory analysis, including supporting evidence for the synthesis section. Finally, the committee identifies key conclusions.
CONTEXT FOR WATER QUALITY AND QUANTITY ISSUES
The problems in the central Everglades result from hydrological conditions that make some areas often too dry, while other areas are often too wet. The sheet flow that characterized the original ecosystem occurs only in some areas when sufficient water is available. Moreover, compartmentalization has limited areas that can sustain flow velocities necessary to support the historic landscape features, such as the ridge and slough. As a result, topography and interconnected biological communities have changed.
Issues with water quality present additional challenges to future restoration progress. Additional stormwater treatment areas (STAs) and/or source controls (e.g., best management practices) are needed to address elevated concentrations and loads of nutrients, most notably phosphorus, in current sources of inflow to the central Everglades (EPA, 2010). Thus, hydrologic restoration involving additional flow volumes or even redistribution of existing flows cannot proceed as planned without introducing levels of contaminants that would substantially affect the ecosystem and likely lead to potential violations of the 1992 Consent Decree. This difficulty was discussed in detail in the committee’s previous report (NRC, 2010). Restoration challenges are exacerbated because the original Comprehensive Everglades Restoration Plan (CERP) assumed that water quality would be largely addressed outside of the CERP by the state’s Everglades Construction Project. Additionally, the natural system was likely sustained by large pulses of wet season flows, but STA performance depends upon dampening such flows (e.g., through the construction of flow equalization basins) to maximize phosphorus removal. Thus, new planning is essential to determine how to support substantive flow restoration while simultaneously protecting the ecosystem from adverse water quality impacts.
Attempts to restore flows in WCA-3 provide a clear example of the challenges stemming from the interplay between water quality and quantity. The hydropattern restoration project component in WCA-3 has been delayed by water quality concerns, and there are additional concerns about the ability to operate Decomp Part 1 under anticipated water quality conditions. The hydropattern restoration component, designed to spread treated water from the STAs along the northern boundary of WCA-3A to better replicate pre-drainage flows, was originally part of the Everglades Construction Project, begun in the 1990s (FDEP, 1999). However, concerns about distributing water with high phos-
phorus concentrations into unimpacted areas have delayed this effort, which has now been moved into Decomp Part 1 (Baisden et al., 2010). The project implementation report for Decomp Part 1, which includes plugging or filling the Miami Canal and hydropattern restoration in northern WCA-3A, has been delayed, largely because of concerns that the project (currently scheduled in the IDS to be completed by 2020) would not be operational because of water quality issues (USACE, 2012b).
One of the key features of Decomp is sheet flow through WCA-3. The Modified Water Deliveries to Everglades National Park (Mod Waters) project included preliminary steps toward that goal via plans for conveyance features in the L-67 levees that would enable water to flow from WCA-3A into WCA-3B and into Northeast Shark River Slough. The Florida Department of Environmental Protection (FDEP) has raised concerns that this restoration component could compromise water quality in WCA-3B, which is currently a rainfall-driven system (E. Marks, FDEP, personal communication, 2012). Additionally, during wet periods, stage constraints in WCA-3B and in the L-29 canal limit the flow of water through WCA-3B and into Northeast Shark River Slough. Instead, water will continue to flow from WCA-3A into the L-67 and L-29 canals, bypassing WCA-3B, and then under the new 1-mile bridge into Northeast Shark River Slough. This flow pattern, which likely will remain the only option until water quality and stage issues are resolved, is hardly the vision of restoration. Even the small adjustments in flow of existing water from WCA-3A to the south represented by the switch from the Interim Operational Plan (IOP) to the Everglades Restoration Transition Plan (ERTP; see Chapter 3) has raised concerns about a decrease in the quality of the water delivered to Everglades National Park (Surratt, 2010). With anticipated new water to increase flows, these concerns about where and when water can flow will be further magnified.
SYNTHESIS OF THE CURRENT STATUS AND TRAJECTORIES OF ECOSYSTEM ATTRIBUTES UNDER VARIOUS SCENARIOS
The following sections discuss the current state, trajectories, and timeframes of recovering ecosystem declines for 10 ecosystem attributes of the remnant Everglades. These ecosystem attributes include total phosphorus (TP) loads, interior TP concentrations, soil phosphorus, cattail (Typha domingensis), periphyton, fish mercury concentrations, peat depth, ridge-and-slough topography, tree islands, and snail kites. These attributes are thought to be good measures of changes in structure and functioning that have occurred because of disruptions in the quantity, distribution, and quality of water inflows. The committee also selected these attributes because they reflect important and valued system characteristics and because there is considerable information on their status from past
monitoring and research. The spread of nonnative species, most notably Burmese pythons, reflects a significant change in the Everglades ecosystem (Dorcas et al., 2012), but abundance, effects, and potential for control of these species are not obviously related to water quality or flow, and so they are not included in this analysis. The attributes selected for this committee’s evaluation are both similar to and different from those selected for other assessment reports. For example, except for periphyton these attributes do not overlap with the stoplight indicators of the South Florida Ecosystem Restoration Task Force (SFERTF, 2010b), which largely focus on organism response. In contrast, there is considerable overlap with the System Status Report (RECOVER, 2010) and CERP performance measures (RECOVER, 2007).
Table 4-1 summarizes the committee’s assessment of status, current trends, and trajectories of all 10 ecosystem attributes under three generic restoration scenarios involving water quality and hydrology. More detailed discussions of each attribute that support the committee’s assessment appear later in the chapter. By necessity, the table simplifies the complex ecosystem responses to management actions (much like a doctor’s health checkup), but the chapter sections attempt to capture some sense of the underlying dynamics and complex interactions among the various ecosystem attributes. This analysis provides a realistic qualitative assessment that underscores the increasingly degraded condition of the remnant Everglades and illuminates the consequences of various restoration scenarios.
The current conditions of the 10 ecosystem attributes in varying states of decline are highlighted in Table 4-1. These conditions are driven by decades of diminished flow volumes and velocities, compartmentalization with associated distortions of water depths, altered hydroperiods,1 and poor water quality. The committee summarizes the condition of each attribute by providing “grades” of the current state relative to the pre-drainage system: “A” no significant degradation, “B” evidence of degradation, “C” degraded, “D” seriously degraded, and “F” near irreversible2 degradation. For most attributes, these grades range from B to C (evidence of degradation to degraded; e.g., interior TP concentrations, TP load, soil P, cattails, periphyton) to D (seriously degraded; e.g., peat, tree islands, ridge and slough, fish mercury). For the snail kite, whose population has declined to near extirpation, the conditions are dire (grade of F or near irreversible damage). The overall grade (or condition) for the 10 attributes is seriously degraded. Clearly the Everglades is in need of an aggressive and sustained restoration effort, beyond what is currently under construction (see Chapter 3), if
______________________
1“Hydroperiod describes the depth, duration and timing of inundation,” (Sklar and van der Valk, 2002). The term is sometimes also defined as the length of time (usually within a year) that a feature or an area is flooded (Bedford et al., 2012).
2The committee considers irreversible degradation to represent ecosystem loss that cannot be restored over many centuries. Extinction is one example of irreversible degradation.
its structure and functioning are to improve. The grades are not intended to be used to prioritize restoration of a single attribute to the detriment of others, but to highlight the urgency for ecosystem restoration actions that could benefit a wide range of attributes, as well as the cost of inaction.
Table 4-1 also summarizes the current trajectories of the attributes (improving, stabilizing, or degrading), which are discussed in more detail in the following sections. The current trajectories in Table 4-1 can be characterized as those largely driven by hydrology (i.e., peat, tree islands, ridge and slough, snail kite); those largely affected by phosphorus concentrations (i.e., interior TP concentrations, periphyton) or load (i.e., TP load, soil phosphorus, cattail); and those largely responding to other mechanisms, although with linkages to hydrology (i.e., fish mercury). As shown in Table 4-1, the attributes most affected by hydrology, in general, are described as degrading, while those affected by phosphorus concentrations show a range of responses. Phosphorus-related stressors are stabilizing or stable to improving, because of the construction of STAs and implementation of source controls since the mid-1990s. Ecosystem conditions affected by phosphorus concentration are stable (periphyton), and those affected by loads are degrading but at slowing rates in some areas (cattails).
Using available science, monitoring, and modeling, the committee also considered how the current trajectories of the 10 attributes might change in response to three hypothetical scenarios of management actions: 1) improved water quality (with no increase in flow), 2) improved hydrology (with no additional water quality treatment), and 3) the combination of improved water quality and hydrology (see Box 4-1 for details on each scenario). Scenario 3 is the preferred scenario because it reflects the original CERP objective, but when it is achieved depends on the implementation schedule of restoration projects addressing water quality and quantity. These scenarios are simplifications of management alternatives. In Table 4-1, the effects of the three restoration scenarios on each ecosystem attribute are evaluated relative to the attribute’s current trend. Thus, a 0 rating for an attribute that is currently degrading means it will continue to degrade under that scenario.
Estimates of the timescales for recovery are also described. These timescales reflect the committee’s qualitative estimates of the time required after substantial degradation has occurred to recover the losses in that ecosystem attribute (i.e., snail kites, tree islands, ridge-and-slough topography, periphyton, peat, cattail) or to attain established restoration criteria (i.e., phosphorus concentrations and loads in the water and soil, fish mercury concentrations). The importance of providing estimates of the timescales for recovery is to emphasize that some attributes will take longer to recover than others. The outcome of this analysis is an understanding that near-term progress that addresses only water quality or water quantity leads to continued system declines of many components. The
TABLE 4-1 Summary of Trajectories of Different Ecosystem Attributes in the Current System and under Three Restoration Scenarios
| Attribute | Current “Grade” of System (A to F) | Current System Trend | Effects of Restoration Scenarios on Current Trends | Timescales of Recovery3 | ||
| (1) Effect of Improved Water Quality1 | (2) Effect of Improved Hydrology1,2 | (3) Effect of Improvements in BOTH Water Quality and Hydrology1 | ||||
| Stressors | ||||||
| TP load | C | Stable to Improving | ++ | – – | + | Years |
| Interior TP conc. | B to C4 | Stable to Improving | ++ | – | + | Decades |
| Soil P | C | Stabilizing | + | – – | + | Decades to centuries |
| Ecosystem condition | ||||||
| Cattail | C | Degrading, but degradation slowing in some areas | + | – – | + | Decades to centuries (years if actively managed) |
| Periphyton | C | Stable | ++ | – – | + | Years. Recovered communities may not be the same as prior to disturbance |
| Peat | D | Degrading in dry areas | 0 | + + | + + | Centuries |
| Tree islands | D | Degrading | 0 | +5 | +5 | Decades to centuries; may require active restoration |
| Ridge and slough | D | Degrading | 0 | + | + + | Centuries; could involve adaptive management |
| Snail kite | F | Degrading | 0 | + | + | Years to irreversible |
| Fish mercury | D | Stable | – | + | + | Years to decades |
analysis also helps to prioritize the focus: stabilizing and ultimately reversing declines of attributes that would take a long time to recover merit higher priority than attributes that would recover more quickly, all other things being equal, especially if other aspects of the restoration depend on them.
Observations
The committee’s qualitative analysis (explained in more detail in the attribute-specific sections later in the chapter) led to several overarching observations. If only system hydrology were to be addressed in restoration projects over
1 The three scenarios considered are detailed in Box 4-1.
2 Hydrologic improvements are assumed to address flow volumes, flow velocity and direction, flow variability and frequency, and water depths and their spatial distribution, timing, and duration.
3 Timescales of recovery reflect the committee’s qualitative estimates of the time required after sub-stantial degradation has occurred to recover the losses in that ecosystem attribute (i.e., snail kites, tree islands, ridge-and-slough topography, periphyton, peat, and cattail) or to attain established restoration criteria (i.e., phosphorus concentrations and loads in the water and soil, fish mercury concentrations).
4 The grade of B applies to Everglades National Park and WCA-2, while a grade of C applies to WCA-3 and LNWR.
5 The “+” for scenario 2 for tree islands reflects minor improvement given the potential negative impacts of increased phosphorus on low elevation islands, whereas “+” for scenario 3 reflects moderate improvement (see the tree island section later in this chapter for more detail).
NOTES:
The following reflect responses to the three scenarios relative to the current system trend:
++ Major improvement in trend
+ Minor to moderate improvement in trend
0 No change
- Minor to moderate decline in trend
- - Major degradation in decline in trend
“Grades” are based on an assessment of the current level of impairment of that ecosystem attribute relative to a pre-drainage state:
A No significant degradation
B Evidence of degradation
C Degraded
D Seriously degraded
F Near irreversible degradation
The trajectories presented in this table do not consider climate change and sea level rise effects, because the analysis was intended to highlight implications of decision-making alternatives over the next 1-2 decades. Climate change and sea level rise could certainly impact long-term trajectories of recovery and timescales of recovery, but these effects were not analyzed for this report.
the next decade, then minor to moderate improvements could be expected for the trajectories of ridge and slough, tree islands, fish mercury, and snail kites, and major improvements for peat. However, these improvements would be accompanied by major expansion of cattails and continued accumulation of soil phosphorus. Soil phosphorus and dense cattail stands, if not actively managed, may persist for decades to centuries because soil phosphorus will continue to impact vegetation—even as phosphorus concentrations in inflow waters improve—until the soils are buried by less contaminated organic matter. However, the timescale for recovery for periphyton is anticipated to be relatively short.
In contrast, if restoration priorities in the central Everglades focus only on
BOX 4-1
Three Scenarios of Management Action Used in the Committee’s Analysis of Ecosystem Attribute Trajectories
The committee developed the following three scenarios for its analysis of likely changes to current ecosystem trajectories under different management actions:
1. Improved water quality (with no increase in flow). For this scenario the committee assumed a decrease in TP concentrations supplied to the Everglades Protection Areas from the STAs to meet the 18 parts per billion (ppb) TP annual flow-weighted mean, which was identified in the Amended Determination as one of two parts of an enforceable framework necessary to meet the 10 ppb geometric mean water quality criterion in the Everglades Protection Area. The second part was a requirement that STA discharge concentrations not exceed 10 ppb as an annual geometric mean (equal to approximately 12 ppb TP as a flow-weighted mean) in more than two consecutive years (EPA, 2010). An STA discharge of 18 ppb TP represents a 28 percent decrease in current flow-weighted mean TP concentrations and loads without any change in flow (compared to the flow-weighted mean of 25 ppb TP across all STAs; Pietro et al., 2010). Meeting both parts of the Amended Determination framework would require lower long-term TP averages than the short-term 18 ppb annual limit considered in this scenario.
2. Improved hydrology (with no additional water quality features). The committee considered improved hydrology to address flow volumes, flow velocity and direction, flow variability and frequency, and water depths and their spatial distribution, timing, and duration. For this scenario, the committee assumed an increase in flow volumes into the northern end of the Everglades Protection Area from the current annual average of 1.4 million acre-feet (MAF) to the CERP-proposed discharge of 1.7 MAF. Nevertheless, based on recent science suggesting a wetter pre-drainage system (~2.1 MAF; Wilcox, 2012), higher total flow volumes could be considered, as was done in the River of Grass planning process. An average annual discharge of 1.7 MAF represents a 21 percent increase in flow. Given that the current extent of STAs do not have capacity to accommodate this additional flow, such a scenario would involve 0.3 MAF of untreated water from Lake Okeechobee (at an assumed concentration of 100 ppb, see Figure 4-2) reflecting an additional 37 metric tons (mt) TP/year load. This represents an approximate 30 to 50 percent increase in the total TP load to WCAs -1, -2, and -3 (considering the five-year moving averages for 2009-2011; see Figure 4-3). The actual load increase could be even greater if the Lake Okeechobee water were distributed to only a single WCA. Additionally, the scenario assumes restoration features, including decompartmentalization, to address the currently altered water distribution and depths in the central Everglades, and releases that generate a flow velocity in the ridge and slough of at least 2.5 cm/s for a few weeks per year.
3. Improvements in both hydrology and water quality. The third scenario assumes the same hydrologic improvements of scenario 2, but it also assumes additional water quality features to reach the water quality objectives outlined in scenario 1. The combination of a 28 percent decrease in phosphorus concentration and a 21 percent increase in flow results in an assumed 13 percent decrease in phosphorus load to the Everglades Protection Area.
As noted previously, these are hypothetical scenarios with postulated endpoints, primarily to illuminate the different trajectories that ecosystem attributes could take under different scenarios. The committee has not analyzed what (or whether) specific project features could create these results.
the water quality of existing flows, then the ecosystem should see some recovery in periphyton and slow improvement in soil phosphorus and cattails. However, peat loss would continue in over-drained areas (e.g., northern WCA-3), and trajectories of deteriorating condition would continue for characteristic landscape features such as tree islands and ridge and slough. Most of these losses would require decades to centuries to recover under ideal conditions. The reality is that these optimal conditions might never occur, and opportunities for restoration could be lost. Meanwhile, the Everglade snail kite faces a serious threat of extirpation. Attributes most directly influenced by hydrology are continuing to decline and are the most difficult to recover (e.g., peat, tree islands, ridge and slough), making addressing them a high priority. The areas of the Everglades where these hydrology-driven attributes are relatively intact and functioning therefore merit priority for protection and management.
The benefits of restoration are not as simple and clear-cut as a tradeoff between water quantity and water quality. In many ways, improvements in water quality are linked with improvements in water quantity, and vice versa. For example, increases in water depth and duration will decrease the decomposition rates of peat and the associated release and transport of phosphorus, sulfur, and other nutrients associated with soil organic matter and therefore improve water quality. Likewise decreases in TP loads will likely encourage the development of native vegetation and the peat, landforms, and hydrology associated with that vegetation. Thus, benefits associated with management actions that improve water quality and water quantity are interconnected. Therefore, this qualitative analysis should be viewed only as a first step toward an integrated analysis of water quantity and water quality management actions. It points to the need for a more critical and comprehensive quantitative analysis using models to evaluate management issues in an integrated manner (see Chapter 5).
Nevertheless, based on this qualitative assessment of the central Everglades system components’ status and trajectories, the committee concludes that near-term progress is needed in the central Everglades to address both water quality and quantity to prevent continued degradation that will take decades or longer to recover under optimal conditions. The committee is encouraged by the Central Everglades Planning Project, which intends to expedite the planning of the next increment of projects focused on the “core” rather than the periphery of the Everglades. This effort represents a significant step forward, although many details remain unresolved. The committee has not reviewed specific project plans, because the planning process was only in the early stages when this report was being finalized. But the Central Everglades Planning Project conceptually offers an opportunity to make significant steps toward reversing the declines in the remnant Everglades.
ANALYSIS OF ECOSYSTEM ATTRIBUTE TRAJECTORIES
The following sections summarize the current state of the science of key ecosystem attributes of the remnant Everglades and provide the basis for the committee’s analysis of current status and trajectories under various restoration scenarios, as summarized in Table 4-1. These ecosystem attributes include: phosphorus loads and concentrations, soil phosphorus, cattails (Typha), periphyton, fish mercury concentrations, peat, tree islands, ridge-and-slough topography, and snail kites.
Phosphorus
The wetlands of the historical Everglades were primarily low-nutrient, phosphorus-limited systems. These biotic communities, including microbes, algae, and aquatic plants, are efficient in utilizing and conserving nutrients through reallocation and uptake of nutrients at very low concentrations. However, phosphorus loading from agricultural and urban lands has converted some of these areas from low-nutrient to high-nutrient systems, particularly near the source areas and along canals. The phosphorus inputs have led to substantial alterations to the indigenous system, including large incursions of cattail and disappearance of periphyton (discussed later in the chapter; McCormick et al., 2002; Noe et al., 2001, 2002; Richardson, 2008; Scheidt and Kalla, 2007).
Phosphorus Concentrations and Loads
This section describes trends in phosphorus loads and concentrations in Lake Okeechobee and in the Everglades Protection Area. Because a substantial quantity of “new water” for the CERP will be delivered from Lake Okeechobee, water quality trends in the lake have important implications for Everglades restoration plans. Five-year trailing moving averages (5YrTMA) of total phosphorus (TP) loads to Lake Okeechobee increased from 1994 until about 2006 to a maximum of about 700 metric tons (mt) per year, peaking after two consecutive years of heavy hurricane activity, and since 2006 the trend has been downward after several dry years (Figure 4-1). Even at the level of 500 mt in 2010, the average TP load is still far above the annual target of 140 mt. Phosphorus concentrations in the lake have seemingly returned to pre-hurricane levels following the sharp increases starting in 2005, although the current values (~100 parts per billion [ppb]) remain far above the target concentration of 40 ppb TP (Figure 4-2). It is too early to discern from the data whether the long-term increasing trends in loads and concentrations are, in fact, beginning to level off. Nevertheless, if increased amounts of Lake Okeechobee water are to be conveyed in the near term to the
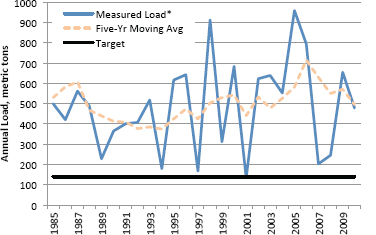
FIGURE 4-1 Annual total phosphorus loads and five-year trailing moving average loads to Lake Okeechobee. Reported loads include atmospheric deposition.
SOURCE: Data from Zhang and Sharfstein (2012).
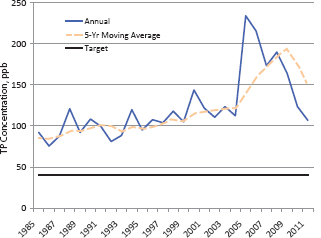
FIGURE 4-2 Annual average concentrations of phosphorus in Lake Okeechobee.
SOURCE: Adapted from Zhang and Sharfstein (2012), Figure 8-12.
remnant Everglades through the CERP, significant additional water quality treatment will be needed.
Trends in TP loads in the Everglades Protection Area are far more encouraging. Annual and 5YrTMA loads for the Arthur R. Marshall Loxahatchee National Wildlife Refuge (LNWR, also called WCA-1), WCA-3, and Everglades National Park have declined sharply since the mid-1990s. In WCA-2 loading rates have been relatively stable since 2005 (Figure 4-3).
Inflow concentrations to LNWR have varied over a wide range (30 to 90 ppb) since 1994, while those for WCA-2 and -3 have steadily declined (Figure 4-4). Concentrations entering Everglades National Park (ENP) have fluctuated in a narrow range around the 10 ppb level since 2000.
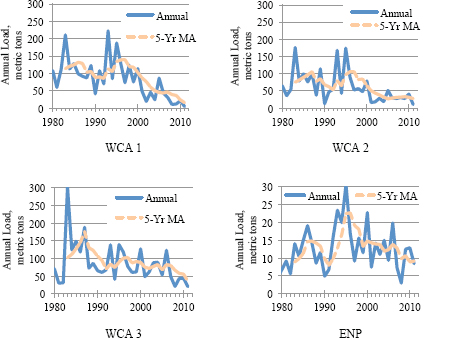
FIGURE 4-3 Annual total phosphorus loads and five-year trailing moving averages of annual loads on individual components of the Everglades Protection Area. NOTE: Please note the different scale used for Everglades National Park.
SOURCE: Payne et al. (2011).
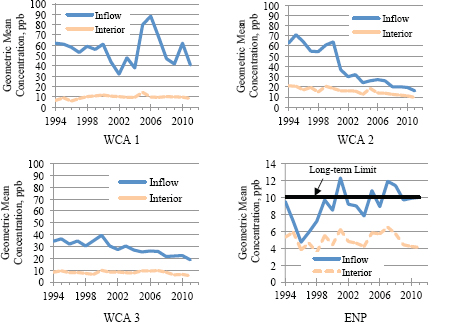
FIGURE 4-4 Annual geometric mean TP concentrations in the Everglades Protection Area. Concentrations at interior locations are geometric means over all monitoring locations within each component of the Everglades Protection Area for which sufficient samples were collected in an annual period. NOTE: Please note the different scale used for Everglades National Park.
SOURCE: Payne et al. (2011).
Since 2000, trends in geometric mean TP concentrations of all interior locations have been relatively steady (in LNWR and ENP) or declining (in WCA-2 and -3). However, temporary increases in interior concentrations have been observed when intense rainy periods are followed drought years (see 2005 data in Figure 4-4).
The concentrations shown in Figure 4-4 are averaged over all interior sites. LNWR, WCA-2, WCA-3, and Everglades National Park (ENP) are protected by the water quality standards established in Florida’s Administrative Code Chapter 62-302.540, but compliance with the phosphorus limits in the Everglades Protection Area is determined by two complex rules:
• In LNWR, WCA-2, and WCA-3, compliance with these standards is determined by a four-part test3 applied separately to impacted areas (soil phosphorus greater than 500 mg P/kg) and unimpacted areas;
• In Everglades National Park, compliance with the standards is determined using the methods as set forth in Appendices A and B of the Settlement Agreement of 1991 entered as a Consent Decree in 1992 and modified in 1995. Appendices A and B of the Settlement Agreement provide an additional level of water quality protection for LNWR and Everglades National Park (Mo et al., 2012).
Unimpacted areas in the WCAs have consistently passed all parts of the four-part test since 2007. Impacted areas have consistently failed the annual all-site geometric mean limit of 11 ppb and the five-year annual geometric mean limit of 10 ppb. Annual geometric means for many individual stations have been below the 15 ppb limit (Payne and Xue, 2012). LNWR has been in compliance with the Consent Decree since June 2009, although “exceedances” occurred in November 2008 and June 2009 (SFWMD, 2009b). Compliance tests for Everglades National Park are based on flow-weighted mean concentrations in inflows to Shark River and to Taylor Slough and Coastal Basins. Shark River inflows just satisfied applicable criteria in 2008-2010. Concentrations in inflows to Taylor Slough and coastal basins have been well below applicable criteria in each of the past three years (Mo et al., 2012).
Given these past trends, the committee anticipates that a hypothetical future scenario of improved phosphorus treatment with no change in flow (scenario 1, Table 4-1) would lead to a 28 percent decrease in the phosphorus load to the Everglades Protection Area (see Box 4-1 for assumptions) and continued decrease in interior concentrations of total phosphorus particularly in LNWR and WCAs-2 and -3, which are most impacted by STA inflows (Table 4-1). Under the scenario of increases in discharge with no additional phosphorus treatment, it is anticipated that there would be a major increase in total phosphorus load to the Everglades Protection Area and, as a result, deterioration in interior phosphorus concentrations (although a lesser effect than what would be observed at inflow locations).4 Finally under the scenario of both increases in flow and additional phosphorus treatment (scenario 3), the committee expects an overall decrease in phosphorus load (13 percent decrease), a lesser improvement than scenario 1
______________________
3The four-part test is used to assess compliance according to the following four provisions: (1) five-year geometric mean is less than or equal to 10 ppb, (2) annual geometric mean averaged across all stations is less than or equal to 11 ppb, (3) annual geometric mean averaged across all stations is less than or equal to 10 ppb for three of five years, and (4) annual geometric mean at individual stations is less than or equal to 15 ppb (FAC §§ 62.302.540).
4The committee has neither evaluated nor is providing any opinion on whether violations of the four-part test or the Consent Decree Appendices A or B will increase.
(see Box 4-1). Because the rates of response of phosphorus loads and interior TP concentrations have been relatively rapid, a reasonable timescale for recovery of total phosphorus loads and associated interior TP concentrations is anticipated to be years to decades (Table 4-1).
Soil Phosphorus
The primary cause of soil phosphorus enrichment in the Everglades is external loading from surface water inflows, although peat oxidation during prolonged drought or fire can contribute to phosphorus enrichment (Bruland et al., 2007; Scheidt and Kalla, 2007). Phosphorus from surface-water inflows is readily retained by sorption to soil or taken up by periphyton and vegetation; thus, soils are integrators of the long-term nutrient supply and indicators of surface-water quality. Soil phosphorus concentrations are generally highest in areas near inflow structures and lowest in interior areas (Figure 4-5). Between these two conditions there is a gradient in quality and quantity of organic matter, nutrient accumulation, and biogeochemical cycles. Cattail encroachment (described in more detail in the next section) is closely linked with increasing levels of soil phosphorus, and restoration goals aim to decrease or maintain long-term average soil phosphorus concentrations below 400 mg/kg to inhibit cattail expansion (Newman et al., 1996; Osborne et al., 2011b; Payne et al., 2003). The state considers soils in the Everglades to be phosphorus-enriched if the soil phosphorus exceeds 500 mg/kg.
Results from the U.S. Environmental Protection Agency (EPA) Regional Environmental Monitoring and Assessment Program (R-EMAP; Scheidt and Kalla, 2007) and the University of Florida Everglades Soil Mapping (ESM) project (Reddy et al., 2005) showed similar spatial patterns in soil phosphorus concentrations in surface (0-10 cm) soils (e.g., Figure 4-5). However, the ESM data suggest that the area of phosphorus enrichment may be smaller than that shown by the R-EMAP data. Scheidt and Kalla (2007) reported that in 2005 the soil phosphorus content exceeded 500 mg/kg in 25 ± 6 percent of the Everglades and 400 mg/kg in 49 ± 7 percent. These values are greater than those observed by EPA in 1995-1996 (16 ± 4 percent and 34 ± 5 percent, respectively). Bruland et al. (2007) also assessed rates of changes in soil phosphorus, focusing on WCA-3A between 1992 and 2003. In 2003, 30 percent of the surface soils were considered enriched (>500 mg/kg) in contrast to 21 percent in 1992.
The majority of phosphorus entering the Everglades is retained in various components of wetlands—either as plant or periphyton biomass or sorbed to peat or particulate matter (Figure 4-6). A lesser amount exists as dissolved phosphorus in the water column, because soluble phosphorus is usually quickly taken
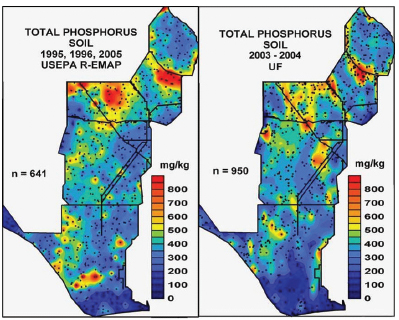
FIGURE 4-5 Spatial distribution of soil total phosphorus for the 0-10 cm soil profile for both R-EMAP (Scheidt and Kalla, 2007) and ESM/UF (Reddy et al., 2005) datasets. Maps by D.J. Scheidt.
SOURCE: Osborne et al. (2011a).
up by biotic communities. Storage of phosphorus in wetland vegetation and other biotic communities is generally small and short-term. When the vegetation dies and decomposes, through the processes of peat accretion, the detrital material accumulates and the plant phosphorus is cycled back into the soil. Soil pore water phosphorus concentrations in the nutrient-enriched areas are approximately 10 times higher than water column phosphorus, creating steep concentration gradients (Koch and Reddy, 1992). Long-term phosphorus storage occurs through burial of stable organic and mineral matter into soils (Craft and Richardson, 1993; Reddy et al., 1993). Vertical soil profiles from WCA-2A show nutrient-enriched material in surface soils (<15 cm depth) but not below 15 cm depth, suggesting minimal redistribution of buried soil phosphorus
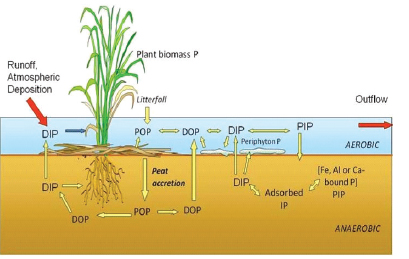
FIGURE 4-6 Phosphorus cycle in Everglades wetlands.
NOTE: Dissolved organic phosphorus (DOP), dissolved inorganic phosphorus (DIP), particulate organic phosphorus (POP), particulate inorganic phosphorus (PIP), and inorganic phosphorus (IP).
SOURCE: Reddy and DeLaune (2008).
(Figure 4-7). Dating techniques (Cs-137 and Pb-210) confirm that the accumulation of phosphorus-enriched soil occurred over the past 20 to 50 years (Craft and Richardson, 1993; Reddy et al., 1993).
Implementation of best management practices (BMPs) in the Everglades Agricultural Area (EAA) and establishment and optimization of STAs, however, have helped to significantly reduce phosphorus loads to the WCAs, particularly during the past decade (Figure 4-3). Based on this information, the committee judges that soil phosphorus may now be stabilizing, although more recent soil phosphorus analyses across a broad spatial scale would be needed to confirm this trend. When enriched soils are exposed to water with low phosphorus concentrations, they release phosphorus until a new equilibrium is reached. Fisher and Reddy (2001) showed high phosphorus flux (after external loads are curtailed) from nutrient-enriched soils that were exposed to surface water with total phosphorus concentration of <10 ppb. If it is assumed that approximately
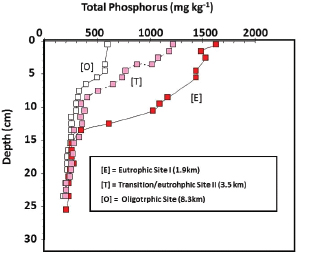
FIGURE 4-7 Vertical distribution of soil phosphorus in WCA-2A. The distance noted in the legend refers to the location of the site samples from the inflow point where agricultural drainage waters are discharged into WCA-2A.
SOURCE: Reddy et al. (1993).
25 percent of the total phosphorus in the top 30 cm of soil (Reddy et al., 1998) is potentially mobile and can diffuse at a rate of approximately 2 mg P m-2 day-1 into the overlying water column, this release of phosphorus from sediments would be sustained for a period of approximately five years. However, because the demand for soluble phosphorus is high among biotic communities, this release of phosphorus from soil will have minimal effect on the overlying water column phosphorus concentrations.
Further reduction of phosphorus concentrations in the inflow water (scenarios 1 and 3, Table 4-1) will help to reduce soil phosphorus enrichment in soils. However, cattails are extremely efficient at recycling phosphorus from aging and dying plant materials, inhibiting the export of phosphorus from dense cattail stands, even under conditions of reduced phosphorus loads. Thus, the recovery of soil phosphorus-enriched areas may take several decades to centuries (Walker and Kadlec, 2011), particularly in cattail-enriched areas. Reducing the impact of soil phosphorus on the Everglades landscape will more likely be
driven by the rate of burial of soil phosphorus than by leaching the phosphorus out of the ecosystem.
Cattail
Species of cattails are among the most widespread and competitive emergent plants in freshwater wetland ecosystems. Their rapid expansion into wetlands that historically were not dominated by cattail has occurred across the globe, mostly in response to various natural and anthropogenic disturbances (Osland et al., 2011; Richardson, 2008). Prior to human impact, cattail had been a part of the Everglades ecosystem, although its extent was minor as evidenced by pollen records from peat cores (Willard and Weimer, 1997). However, since the 1970s, cattail has been spreading in phosphorus-enriched areas of the oligotrophic Everglades at the expense of sawgrass and other less competitive species (Richardson, 2008). This spread is associated with elevated phosphorus loads and altered hydroperiods (Newman et al., 1996). In areas that have been overdrained, oxidation of soil organic matter can release phosphorus, resulting in enhanced cattail growth in absence of external loads. Cattail expansion, resulting from years to decades of sediment phosphorus enrichment, is generally preceded by changes in more sensitive components of the ecosystem such as periphyton (Surratt et al., 2012; see next section). The spread of cattail greatly impacts ecosystem processes, including an increase in primary production, replacement of water column autotrophy with heterotrophy,5 and sediment accretion (Hagerthey et al., 2010; Miao et al., 2000; Richardson, 2008). Other ecological implications of cattail expansion include decreases in dissolved oxygen concentration, degradation of fish and wading bird habitat, accelerated biogeochemical cycling of nutrients and metals, such as mercury (Osborne et al., 2011b), and marked changes to the calcareous periphyton and microbial communities (Gaiser et al., 2005; Ogram et al., 2011; Reddy et al., 1999).
Recent Trends in Cattail Expansion
Temporal changes in the extent of cattail in WCA-2A and WCA-3 have been well documented in nutrient gradient research, transect sampling, and remote sensing. In the 1940s, WCA-2A included nearly monospecific sawgrass plains in addition to sawgrass mosaics, wet prairies, and sloughs, but by 1991, cattail monocultures and larger areas of sparse cattails appeared in the eastern portion of the impoundment and along the southwestern boundary, with continued
______________________
5Autotrophs use energy from sunlight or inorganic chemicals, whereas heterotrophs derive energy from organic carbon.
expansion through 2003 (Figure 4-8). The average rate of cattail expansion, however, has decreased from 961 hectares/year (between 1991-1995) to 312 hectares/year (between 1995-2003; Rutchey et al., 2008), likely because of the decrease in phosphorus loads into WCA-2A after 1995 (Figure 4-3b). The southward expansion of cattail could also have been decreased by the presence of dense sawgrass in the central parts of WCA-2A, which is more resistant to cattail invasion than open water sloughs (Richardson, 2008). Recent data and analysis indicating that the spread of cattail in central Everglades marshes has slowed somewhat in the past 5-10 years also have been reported by RECOVER (2010). However, considerable evidence (reviewed in Osborne et al. [2011b]) shows that existing high phosphorus concentrations in the soils of cattail marshes represent a source of phosphorus that will continue to impact downstream marshes even if canal phosphorus loading decreases substantially (see also previous section on soil phosphorus).
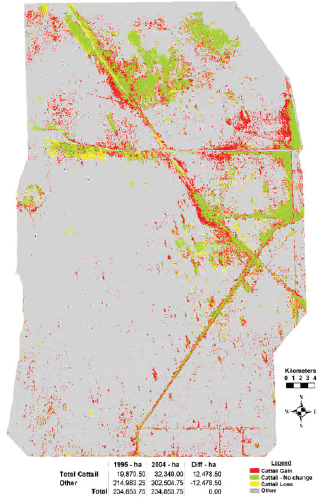
Vegetation mapping of WCA-3 shows a 63 percent increase in cattail acreage (nearly 12,500 ha) between 1995 and 2004 (Figure 4-9; Sklar et al., 2011). The emergent cattail mapped in the 2004 survey occurred both near canals and in interior locations. This rapid cattail expansion has been explained by hydrologic alterations of WCA-3 combined with large inflows of phosphorus, although the relative importance of these factors in cattail establishment has not been determined.
The emergence of cattail has also been recently observed in Upper Taylor Slough, where vegetation transects have been monitored since 1979.
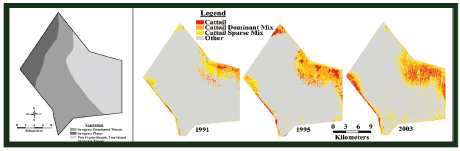
FIGURE 4-8 WCA-2A: (A) Map of three physiographic landscape categories within the WCA-2A impoundment based on 1940’s aerial photography; (B) cattail spread 1991-2003.
SOURCE: Rutchey et al. (2008).
Between 2003 and 2007, sawgrass along one transect was replaced by cattail (Figure 4-10). The combination of increased hydroperiods associated with the operation of detention areas constructed for the C-111 South Dade project and increased nutrient loading likely allowed cattail to outcompete other species acclimated to shorter hydroperiods and oligotrophic conditions (e.g., muhly grass [ Muhlenbergia capillaris]). This resulted in cattail spread of more than 0.2 km2 in Taylor Slough. Although the cattail stand there is not as dense as tall monocultures that can be seen in WCA-2A, it is still denser than natural coverage of cattail in unimpacted areas, which is typically well below 5 percent with the exception of naturally enriched spots such as bird rookeries and alligator holes. According to Surratt et al. (2012), upstream surface water quality monitoring showed concentrations that were below phosphorus targets established for Taylor Slough. The recent discovery of cattail stands suggests that Taylor Slough “has been experiencing impacts for years to decades and that surface-water quality alone did not serve as an early warning indicator” (Surratt et al., 2012).
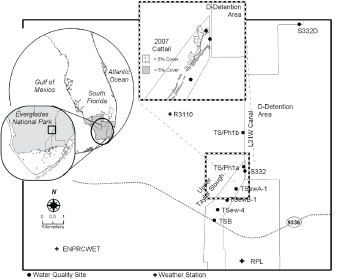
FIGURE 4-10 Upper Taylor Slough surface water quality and weather monitoring sites. Upper inset shows the cattail expansion in 2007. Stage gages are located at the TSB and S332.
SOURCE: Surratt et al. (2012).
To prevent dense cattail expansion, areas with newly emerging cattails should be treated before they become well established (Sklar et al., 2012).6
Predictions under Various Scenarios
Cattail expansion after the 1970s has resulted from increased phosphorus input combined with changed hydrology. Based on the trend data in WCA-2 and -3, cattail expansion is likely to continue in the future if the status quo is maintained. Improvement in water quality (scenario 1; Table 4-1) should slow cattail expansion overall, although altered hydrology would continue to foster cattail growth in overdrained areas. If no attempt is made to remove the cattail, then existing areas of cattail-dominated marshes are likely to persist for a long time—probably decades—even if the input of phosphorus-enriched water stops. This condition is due to the existing phosphorus-enriched soil in cattail-dominated areas and to an efficient internal recycling of nutrients from older to newer plant parts (see Soil Phosphorus section). Aggressive cost-effective cattail management strategies tested in small field-scale plots have been successful in removing dense cattail stands and rehabilitating the nutrient-enriched marsh using a combination of fire and herbicides (see Box 4-2). If hydrology is improved without additional water quality treatment (scenario 2; Table 4-1), then significant cattail expansion would be expected because of the substantial increase in phosphorus loads.
If both water quality and hydrology are improved according to the assumptions of scenario 3 (see Box 4-1), then the net phosphorus loads should decrease by 13 percent. Thus, the overall improvements to both water quality and hydrology should reduce the spread of cattail. However, the committee estimated that the large internal reservoirs of soil phosphorus would lead to minor to moderate (rather than major) improvements in current trends. In specific areas where much more water is being delivered than the 21 percent increase assumed in scenario 3, an expansion of cattail would be expected if overall phosphorus loads to those area increase. As noted previously for Taylor Slough, higher phosphorus loads and longer hydroperiods are suitable conditions for cattail expansion (Newman et al., 1996).
Periphyton
A complex entity called periphyton has been recognized as a suitable indicator of water quality deterioration in the Everglades, as well as a performance
______________________
6Recent results from a SFWMD study indicate that a single aerial application of imazamox at a rate of 0.28 kg/ha provided excellent control of cattails in marginally invaded marsh ridge-and-slough habitat with only minimal damage to desirable emergent macrophytes.
BOX 4-2
Aggressive Cattail Management Strategies
Two large ecosystem-scale experimental manipulations of cattail have been conducted in WCA-2 to test the efficacy of cattail removal strategies (Sklar et al., 2010, 2011) with promising results, and there are records on cattail suppression from other ecosystems such as Palo Verde National Park in Costa Rica or the upper St. Lawrence River wetlands (Farrell et al., 2010; Osland et al., 2011). Of the two Everglades experiments focused on removing cattail as a restoration method, the Fire Project aims to assess whether repeated fire can be used as an effective tool to manage cattail expansion. The project has been conducted in WCA-2A and has considered water levels, fuel loads, and fire intensity to maximize phosphorus loss from highly enriched habitats. Project results have been summarized in a process-based biogeochemical model that simulates plant growth and phosphorus dynamics in water to evaluate the effects of prescribed fire (Tian et al., 2010). The objectives of the Cattail Habitat Improvement Project (CHIP) are to accelerate the ecological rehabilitation of the phosphorus- enriched, emergent macrophyte Everglades marsh. Using a combination of herbicides and fire, open areas were created in enriched and moderately enriched areas of WCA-2A in July 2006. The most recent aerial photographs clearly show that for more than 925 days since the last herbicide application, and 1,406 days since the burn, an alternative regime of submerged aquatic vegetation (SAV) has been sustained in an otherwise cattail-dominated region of the Everglades. With minimal further active management, these plots could be sustained and dramatic shifts in phosphorus storage would likely be observed (Sklar et al., 2011).
Either fire alone or a herbicide-fire combination is a very cost-effective management approach. The cost estimate of removal ranges from approximately $40/acre using burning to $125/acre using herbicide (S. Newman, SFWMD, personal communication). Given the spatial extent of cattail, this type of management can be readily conducted. At the same time, the risk of burning or otherwise trying to eliminate most of the cattail still needs to be evaluated. It has been assumed that a large burn or herbicide treatment could cause a large downstream release of soil phosphorus. This effect must be considered as active marsh improvement is scaled up. Sufficient buffer zones will be needed to prevent any downstream nutrient transport (S. Newman and F. Sklar, SFWMD, personal communication, 2012).
measure of restoration success. Periphyton is defined as diverse communities of microorganisms, including cyanobacteria and algae, attached to the bottom sediments or stems of aquatic plants, or freely floating on the water surface (Figure 4-11; McCormick and Stevenson, 1998; Gaiser et al., 2011). Periphyton provides important functions in the Everglades: it contributes significantly to primary production and influences soil quality, nutrients, and dissolved gases (Gaiser, 2009; Liao and Inglett, 2012; Ogram et al., 2011). Periphyton was once found abundantly in the Everglades ecosystem, with the largest expanses in WCA-3 and Everglades National Park (Gleason and Spackman, 1974). However, throughout much of the Everglades, periphyton communities have been either

FIGURE 4-11 Photographs showing different types of periphyton-substrate associations: (A) calcareous floating (metaphyton), (B) calcareous epiphytic (attached to stems of aquatic plants), (C) calcareous epipelic (attached to bottom sediments), and (D) green filamentous periphyton
SOURCE: Gaiser et al. (2011).
reduced or even completely eliminated because of exposure to high phosphorus loads and, in some areas, replacement by dense cattails (McCormick and O’Dell, 1996; McCormick et al., 1996).
Currently, periphyton is most abundant in oligotrophic sloughs and wet prairie habitats (Richardson, 2008). Periphyton is known to respond quickly (days to weeks) across large spatial scales (meters to tens of kilometers) to changes in environmental conditions (Gaiser et al., 2004). Several metrics serve as reliable measures of the response of periphyton to water quality changes— abundance (total biomass), quality (TP concentration in periphyton tissue), and species composition—which can be used collectively to assess periphyton’s condition. Of these metrics, the TP concentration in periphyton tissue has been identified as one of the best measures of phosphorus load history (Gaiser, 2009; Gaiser et al., 2006; McCormick and Stevenson, 1998). Although increases in water and
soil phosphorus are only detectable after years of enhanced phosphorus loading, effects upon periphyton TP concentration are immediate (Gaiser et al., 2004).
Recent Trends
Large areas of periphyton mat have been lost in WCA-2 and WCA-3 because of nutrient impacts, and species composition and metabolism have been altered (Gaiser et al., 2004; McCormick and O’Dell, 1996). Analysis of periphyton metrics in 2005 and 2006 demonstrated a general north to south trend of increasing periphyton biomass and decreasing periphyton TP concentration over the Everglades Protection Area (Figure 4-12a). WCA-2 and WCA-3 showed a slight improvement in periphyton TP concentration between 2005 and 2006 followed by some increases in 2007 (Figures 4-12b and Figure 4-13; RECOVER, 2010).
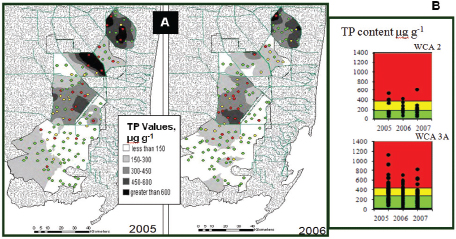
FIGURE 4-12 Examples of system assessment based on periphyton. (A) Pattern of distribution of TP concentration in periphyton across the Greater Everglades in fall 2005 and 2006. (B) Distribution of values for periphyton phosphorus, TP, for surveys in 2005 and 2006 in WCA 3A.
NOTE: Sites are coded as: green = within 1 standard error [SE] of mean in natural system, yellow = > 1 SE of natural system mean, and red = >2 SE of natural system mean.
SOURCES: Gaiser (2009) and RECOVER (2010).
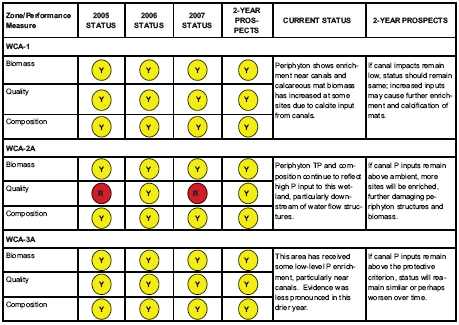
FIGURE 4-13 Stoplight indicator assessment of each WCA. The data are scored based on the proportion of sites falling within each categories. Red indicates failure, yellow indicates caution and green indicates success.
SOURCES: SFERTF (2010b); Gaiser, (2009).
However, it is difficult to discern temporal trends with only three years of data, particularly in the context of natural hydrologic variation.
Predictions under Various Scenarios
Continued or increased input of above-ambient phosphorus concentrations will both increase severity of enrichment effects near canals and cause periphyton deterioration effects to cascade downstream. In contrast, enhanced water treatment will promote periphyton recovery. Periphyton can recover in areas recently dominated by cattails only if the cattails are first eliminated (see
Box 4-2). Increased input of water treated for phosphorus removal (i.e., scenario 3 in Box 4-1) will likely increase periphyton development, particularly in areas that have been overly dry. However, because the overall load reductions are greatest in scenario 1 (see Box 4-1), the committee estimates larger improvement for this scenario.
No long-term observations exist to provide reliable predictions for time-frames for recovery of periphyton once it is degraded or eliminated from an area. However, there is anecdotal support for a relatively rapid recovery of periphyton calcareous mats after the completion of the phosphorus dosing experiment in Everglades National Park (Gaiser, FIU, personal communication, 2011). After a complete collapse of the calcareous mats following dosing of phosphorus above-ambient levels for five years, the periphyton seemingly recovered about a year after dosing was terminated. Periphyton re-appeared quickly where cattails were not present, but the periphyton recovery was never fully documented. A second example comes from the CHIP project (Box 4-2; Sklar et al., 2011), for which higher periphyton productivity was observed in the plots after cattails were removed compared to the cattail-dominated control. However, the taxonomic composition of this re-emergent periphyton is not known (S. Newman, SFWMD, personal communication, 2011). The establishment of periphyton in STAs could also provide information on the potential for recovery of this important ecosystem component, although little data are available on STA periphyton. Because periphyton is such a diverse and complex community, better characterization is needed. To obtain a better understanding of under what conditions and how quickly periphyton can be restored, performance measures such as biomass, species composition, and nutrient content, as measured in the Everglades Protection Area by Restoration, Coordination, and Verification (RECOVER), should be monitored in STAs, CHIP, and other manipulated settings.
Fish Mercury
Mercury contamination is a chronic environmental problem in the South Florida ecosystem. Elevated concentrations of mercury have been observed in fish and other animals such as the American alligator, softshell turtles, and the Florida panther (Gu et al., 2012). The source of ionic mercury inputs to South Florida is overwhelmingly from atmospheric deposition, and these inputs have remained relatively constant to the Everglades since the early 2000s. However, the formation of methyl mercury—which strongly bioaccumulates up the aquatic food chain and results in high concentrations in fish—varies across the Everglades landscape based on hydrology and the supply of sulfate, phosphorus and other contaminants. Elevated concentrations of sulfate, primarily derived from agricultural lands, are processed in downstream wetlands by
sulfate-reducing bacteria, which also convert ionic mercury to methyl mercury. Human exposures to mercury are largely from consumption of fish. Methyl mercury is a neurotoxin, and to limit human exposure to mercury, Florida has issued fish consumption advisories.
The spatial pattern of sulfate across the Everglades Protection Area reflects the source of sulfate largely from the Everglades Agricultural Area (Figure 4-14). The highest sulfate concentrations generally occur in WCA-2 and decrease southward. High sulfate concentrations are also noted along canals because of preferential flow along these conduits. Sulfate concentrations have generally held steady or declined between 1979 and 2010 in inflows to and outflows from the major regions of the Everglades Protection Area (Payne et al., 2011). These long-term declines in sulfate are linked to long-term declines in fish mercury concentrations.
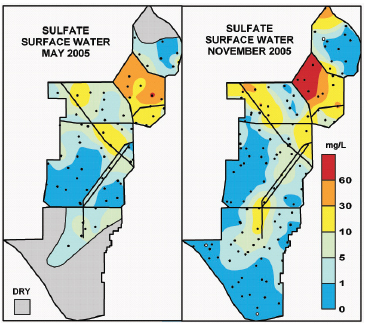
FIGURE 4-14 Spatial pattern of concentrations of sulfate in the Everglades Protection Area.
SOURCE: Scheidt and Kalla (2007).
A complex relationship exists between sulfate concentrations and the formation of methyl mercury. Maximum formation of methyl mercury appears to occur around sulfate concentrations of 10-20 mg/L. At sulfate concentrations below this range, increases in sulfate will result in increases in methyl mercury formation. At sulfate concentrations above this value, increases in sulfate will result in decreases in methyl mercury formation. This response results in a “hotspot” of elevated methyl mercury concentrations and fish mercury concentrations in the Everglades (see NRC, 2010). The location of this hotspot would likely shift with variations in water discharge and transport of sulfate.
Monitoring data show clear spatial patterns of fish mercury that are linked to the spatial patterns of sulfate (Figure 4-14) and nutrients (Figure 4-4) in the Everglades. Recent monitoring data (2009-2010) for largemouth bass show low methyl mercury concentrations in the STAs (~0.1 ?g/g), high concentrations in the WCAs (~0.5 ?g/g), and very high concentrations in Shark River Slough in the Everglades National Park (~1.2 ?g/g; much higher than in other portions of the Park) (Gu et al., 2012). For reference the EPA-recommended criterion for fish mercury is 0.3 ?g/g. This spatial pattern reflects variations in the processes controlling fish mercury concentrations. Under high sulfate concentrations in waters adjacent to the EAA, as in the STAs, microbes produce high sulfide concentrations that inhibit the production and bioavailability of methyl mercury (Benoit et al., 2003). As EAA drainage moves south into the WCAs and ultimately into Everglades National Park, sulfate concentrations and the production of sulfide generally decrease, thereby allowing for more formation of methyl mercury by reducing inhibition effects from sulfide. Nutrients potentially also play an important role. High inputs of nutrients from the EAA support high biomass production, which decreases the mercury concentration in biota via a process known as biodilution.7 With decreases in phosphorus concentrations with distance from the EAA, decreases in net aquatic production decrease the biodilution phenomenon and concentrations of mercury in fish and other biota increase.
Long-term observations show that concentrations of mercury in largemouth bass have significantly declined in the WCAs since measurements were initiated in the late 1980s (Figure 4-15). Mercury concentrations in largemouth bass were very high in the early to mid-1990s in the WCAs. Indeed the “hotspot” of fish mercury at that time was located in WCA-3A. However, the decreases in fish mercury concentrations ceased by 1998, and concentrations have remained relatively constant since that time. These decreases in mercury concentrations in largemouth bass are thought to result from declines in sulfate inputs (Kalla et
______________________
7Biodilution is a phenomenon through which concentrations of a contaminant (e.g., mercury) in organisms decrease because of increases in nutrient supply and associated increases in biomass (Chen and Folt, 2005; Pickhardt et al., 2002).
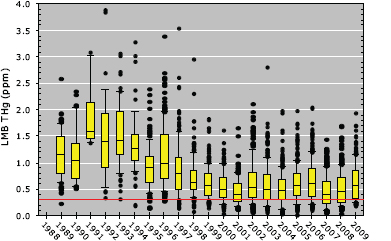
FIGURE 4-15 Annual summaries of mercury concentrations in largemouth bass collected from canal and marsh sites in WCAs -1, -2, and -3 from 1989 to 2010. Note that mercury concentrations are normalized to a standard fish length of 356 mm. The red line indicates the EPA methyl mercury criterion of 0.3 ppm.
SOURCE: Axelrad et al. (2011).
al., 2010). At Shark River Slough, long-term measurements of mercury concentrations in largemouth bass show considerable year-to-year variability, with no significant trends (Gu et al., 2012).
Using the current understanding of the patterns and mechanisms driving fish mercury concentrations in the Everglades, one can speculate on the trajectories that fish mercury concentrations might take under various future management strategies. The two major drivers of fish mercury concentrations that might be affected by restoration management changes are: (1) agricultural sulfate inputs that control the production of methyl mercury and (2) phosphorus inputs that control fish mercury concentrations. Based on the available monitoring data, it appears that fish mercury concentrations are in quasi steady-state with respect to these drivers and that they respond relatively quickly to environmental change (~ years to a decade). If water and phosphorus inputs to the Everglades remain steady, then fish mercury concentrations should remain relatively constant through time. With improved hydrology (i.e., increased water discharge, decom-
partmentalization) but no additional water quality features (i.e., scenario 2 in Box 4-1), it is anticipated that fish mercury concentrations would decrease. With the restoration of sheet flow the interaction of water with wetlands will likely facilitate the removal of sulfate, thereby reducing methyl mercury formation and fish mercury concentrations. Additionally, increased phosphorus concentrations associated with scenario 2 lead to greater biodilution of mercury.
This projected outcome of hydrologic change is based on considerable speculation about the driver of system response (i.e., biodilution). Other outcomes may occur. Alternating drying and wetting cycles can facilitate mineralization of organic sulfur in peat deposition, releasing sulfate followed by the methylation associated with the subsequent sulfate reduction. Restoration of a more normal hydroperiod to the Everglades would likely diminish this phenomenon and could decrease fish mercury concentrations. Also the committee has assumed that elimination of channelized flow with decompartmentalization would decrease sulfate transport southward and decrease fish mercury concentrations. However, a more distributed transport of sulfate, which would be a by-product of decompartmentalization, would likely spread out mercury contamination in fish. The committee believes this action would result in an overall decrease in fish mercury concentrations, but this management action could increase fish mercury concentrations in areas of the Everglades that previously have not experienced high concentrations.
If improved controls on phosphorus supply decrease phosphorus loading to the Everglades, fish mercury concentrations could increase. This response would be due to decreases in biomass production associated with decreases in nutrient loading and a resulting decrease in the biodilution of fish mercury. Finally, management measures that involve simultaneous increases in discharge and decreases in phosphorus would likely decrease fish mercury because of the effectiveness of decompartmentalization in the immobilization of sulfate (but see above discussion). These effects are summarized in Table 4-1.
Peat
Most of the historical Everglades was underlain by organic-rich peat soils (Figure 4-16), approximately 2 to 10 feet thick. The peat soil’s thickness decreased toward the southern Everglades, where it formed a thin, sometimes patchy layer over marl soils (McVoy et al., 2011).8 In addition to providing the substrate for the sawgrass plains and ridge-and-slough landscapes, peat soils in the Everglades provided the critical elevation differences that hydrologically dif-
_________________
8Marl soils are comprised of calcitic mud deposited from calcareous periphyton and have lower organic content.

FIGURE 4-16 Peat and marl soils of the Everglades.
SOURCE: Scheidt and Kalla (2007).
ferentiated ridges from sloughs, and, in many cases, tree islands from sloughs. In the pre-drainage system, the decreasing thickness of the peat soils with distance downstream from Lake Okeechobee was responsible for much of the regional land slope that drove sheet flow.
Peat accumulates when detrital plant biomass partially decomposes under anaerobic conditions and is buried and compacted, creating a soil with approximately 90 percent organic matter content (Figure 4-17). Rates of peat accumulation in the Everglades are a function of the balance between net primary productivity—the transformation of inorganic carbon (CO2) into organic carbon through photosynthesis—and abiotic and biotic decomposition processes. In areas unimpacted by phosphorus, peat accretion rates are extremely low, in the range of 0.2 to 2 mm per year (see Table 4-2; Box 4-3). Phosphorus-enriched areas have been shown to accrete organic matter at higher rates (approximately 5 to 12 mm per year), although peat produced in cattail-dominated areas is of poor quality and easily decomposed, which releases nutrients into the water column.
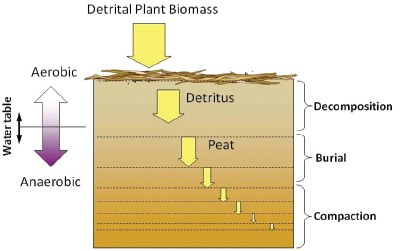
FIGURE 4-17 Decomposition and burial of organic matter and genesis of organic soil.
SOURCE: Reddy and DeLaune (2008).
Altered hydrologic regimes have led to significant losses of peat, particularly in overdrained areas. Rates of peat loss (i.e., loss of peat soil mass and the associated critical loss of peat-based elevation or subsidence) are directly related to the position of the water surface relative to the ground (peat) surface. As long as surface water covers the peat, anaerobic conditions prevail within the soil profile, and peat accumulation outpaces peat oxidation. If the water table drops below the surface, air enters the portion of the profile above the water table and allows aerobic microbial oxidation of the organic matter to occur. Peat decomposition also releases phosphorus, sulfur, and other nutrients, impacting water quality once the area is rehydrated. Peat decomposition rates under aerobic (drained) soil conditions have been shown to be approximately 3 to 5 times higher than under anaerobic (flooded) soil conditions (DeBusk and Reddy, 1998; McLatchey and Reddy, 1998; Wright and Reddy, 2001). Additionally, as the water table drops, a larger fraction of the profile is subject to oxidation, and the overall rate of subsidence increases. In a field study using controlled water tables, Stephens and Johnson (1951) found a linear relationship between subsidence rate and depth of the water table. Similarly, Volk (1973) found that the decomposition
TABLE 4-2 Peat Accretion Rates in Select Hydrologic Units of South Florida Wetlands
| Location | Method Used | Peat Accretion Rates (mm yr-1 ) | Reference |
| Loxahatchee NWR | 137Cs | 0.8 | Craft and Richardson (1993) |
| 137Cs | 1.1 – 4.2 | Robbins et al. (1999) | |
| 210Pb | 1.1 | Craft and Richardson (1993) | |
| 14C | 0.9 | Craft and Richardson (1993) | |
| WCA-2A | 137Cs | 1 – 12 | Reddy et al. (1993) |
| WCA-2A enriched | 137Cs | 5.3 ± 0.9 | Craft and Richardson (1993) |
| 137Cs | 2.6-5,1 | Robbins et al. (1999) | |
| 210Pb | 5.8 ± 1.4 | Craft and Richardson (1993) | |
| WCA-2A unenriched | 137Cs | 2.0 ± 0.6 | Craft and Richardson (1993) |
| 137Cs | 1.1 – 1.2 | Robbins et al. (1999) | |
| 210Pb | 2.0 ± 0.1 | Craft and Richardson (1993) | |
| 14C | 0.6 | Craft and Richardson (1993) | |
| WCA-2B | 137Cs | 2.4 ± 0.4 | Craft and Richardson (1993) |
| WCA-3A | 137Cs | 1.7 ± 0.3 | Craft and Richardson (1993) |
| WCA-3A north | 137Cs | 0.7-2.8 | Robbins et al. (1999) |
| WCA-3A north | 137Cs | 0.4-1.4 | Robbins et al. (1999) |
| 210Pb | 1.4 | Craft and Richardson (1993) | |
| 14C | 0.2 | Craft and Richardson (1993) | |
| Shark River Slough-Ridge | 137Cs | 2.0-3,5 | Clark and Reddy (2007) |
| Shark River Slough-Slough | 137Cs | 1.3-5.4 | Clark and Reddy (2007) |
| Taylor Slough | 210Pb | 3.0 | Meeder et al. (1996) |
| Mangroves | 210Pb | 1.0 | Meeder et al. (1996) |
| STA-1W | Soil properties-bulk density, TP, δ15N, and δ13C | 10 ± 3 | Bhomia et al. (2012) |
| STA-1W (Cell 5) | Soil properties-bulk density, TP, δ15N, and δ13C | 12 ± 6 | Bhomia et al. (2012) |
| STA-2 | Soil properties-bulk density, TP, δ15N, and δ13C | 11 ± 3 | Bhomia et al. (2012) |
| STA-3/4 | Soil properties-bulk density, TP, δ15N, and δ13C | 17 ± 8 | Bhomia et al. (2012) |
NOTES: Accretion rates measured using the Cs-137 technique represent < 40 years and using Pb-210 techniques represent <100 years. Values shown (at limited sites) show peat accretion in inundated areas.
rates of Everglades peat soils were significantly lower when the water table was raised from 25 cm to 5 cm below the soil surface.
Other factors can also impact rates of peat loss. Nutrient loading can increase peat decomposition rates (DeBusk and Reddy, 1998). If the peat soil actually dries, then it becomes at risk for fires that can cause substantial losses, as in the May 1981 muck fires in northern WCA-3A when 9 to 29 cm of peat were lost (Wetzel, 2002).
One of the most well-known cases of drainage of peatlands occurred in the EAA, which was drained for agricultural production beginning in the early
BOX 4-3
Estimated Historic Rates of Peat Accretion
Based upon 14C dating, McDowell et al. (1969) estimated that peat soil in the EAA first formed about 4,400 years ago. From these studies it was estimated that it took approximately from 500 to 1,000 years to form the first 7.6 cm of a marl/organic soil on top of the bedrock, while peat developed at about 7.3 cm per century from about 3,500 to 1,200 years before present. By 1914, approximate peat depth was 3.65 m, which represents an average accretion rate of about 8.4 cm/century (0.84 mm/yr). Scholl et al. (1969) estimated soil accretion using 14C dating in sediment cores obtained from several locations in the freshwater Everglades, coastal mangroves, Florida Bay, and Rodriquez key (Atlantic Ocean). During the past 4,000 years, coastal sedimentation has occurred at a rate of 3 cm/100 years (0.3 mm/yr). Calcitic mud formation in nearby coastal freshwater swamps has averaged 1.6 cm/100 years (0.16 mm/yr). Soil accretion rates in these ecosystems were approximately equal to the rate of sea-level rise.
1900s. These soils subsided at a rate of approximately 2.5 cm/year, which then declined to a current average rate of about 1.5 cm/yr (Shih et al., 1998; Snyder, 2005). Over the course of less than 100 years in the EAA, a significant portion of the peat, which took more than 5,000 years to form, was lost to biological oxidation and fire (Stephens et al., 1984) (Figure 4-18).
A substantial fraction of Everglades peat soils have already been lost, leaving about 25 percent of the remnant Everglades with a peat thickness of less than 1 foot (Scheidt and Kalla, 2007). Sklar et al. (2010) estimated that 7 billion cubic meters of peat have been lost in the remnant Everglades since drainage began. Scheidt et al. (2000) estimated that between 1946 and 1996, water depths lowered by drainage and reduced inflows have caused the Everglades Protection Area to lose up to 28 percent of its organic soil volume (Figure 4-19), with soil oxidation, subsidence, and peat fires as the causes. Upper and lower limits of peat loss between 1946 and 1996 show significant loss in northern WCA-3A, -3B, -2A and Northeast Shark River Slough in Everglades National Park (Figure 4-20; Scheidt et al., 2000). McVoy et al. (2011) state that the historical record suggests that peat soil has also been lost in Shark River Slough and from what are now called the marl prairies flanking Shark River Slough to the east and west. This loss likely occurred before 1946 and therefore would not be reflected in the Scheidt et al. (2000) study.
Equally important as the overall loss of peat is the spatial distribution of that loss. The spatially uneven rates of peat accretion and loss within the impounded system hinder the slope-generating role for peat, thereby affecting the capacity to support the water flow and depths that ecological attributes such as ridge and

FIGURE 4-18 Peat loss as a result of soil subsidence in the Everglades Agricultural Area. The concrete post was buried in 1927 at the Everglades Research and Education Center, University of Florida, Belle Glade, Florida. In 1927, the top of the post was flush with the soil surface, and more than 6 feet of soil subsidence has occurred since that time.
SOURCE: K. R. Reddy, University of Florida.
slough and tree islands depend upon. Within the impounded WCAs, a portion of the original peat-based land surface slope remains, yet the water surface, rather than paralleling the ground surface as it did originally under sheet flow, now tends toward level. As a result, the higher upstream areas (e.g., northern WCA-3a) have shorter hydroperiods and endure aerobic conditions for longer
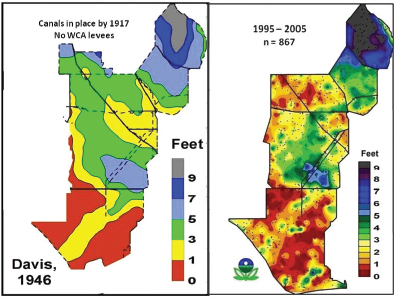
FIGURE 4-19 Maps of peat thickness in the Everglades in 1946 (left) and 1995-2005 (right).
SOURCES: Davis (1946); Scheidt and Kalla (2007).
durations, accelerating peat decomposition. The net effect is to undo the original downward slope of the ground surface, driving the peat surface within the impoundment area toward becoming level, that is, parallel to the water surface.
Predictions under Various Scenarios
Overall, accretion of organic matter in the central Everglades is very slow, and it takes centuries to accumulate significant amounts of organic matter under oligotrophic conditions (Table 4-2). Because decomposition exceeds primary productivity when water level is below the soil surface for extended periods, overly dry areas of the Everglades continue to lose peat. Water quality improvement alone will not alter the trend of peat loss in dry areas. Hydrologic restoration to increase hydroperiods in the Everglades, particularly in currently overly drained areas, is key to reversing ongoing peat loss.
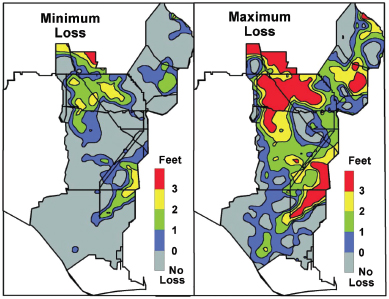
FIGURE 4-20 Difference in peat thickness between 1946 and the R-EMAP studies.
SOURCE: Scheidt et al. (2000).
The unusual nature of the Everglades as a “hillslope wetland” makes protection even more challenging. If impoundments are not removed, and if a sloped water surface parallel to the sloped ground surface is not restored (i.e., sheet flow), then the impounded portions of the remnant Everglades will remain on a trajectory to become a series of disjointed flat steps, without the slope necessary to sustain the ridge-and-slough landscape. Protection of peat soils in the remnant Everglades would require removal of impounding impediments to flow as well as simultaneous restoration of upstream inflows.
Ridge and Slough
The ridge-and-slough landscape in the historical Everglades consisted of patterned peatland surfaces with hundreds of alternating ridges and linear sloughs, aligned parallel to the direction of regional water flow. In its original form, the
ridge-and-slough landscape was the essence of the Everglades—half land and half water. Tree islands were scattered irregularly throughout the landscape, slightly higher and drier than the ridges. The ridges were covered by sawgrass, while the sloughs, typically 1 to 2 feet lower in elevation under pre-drainage conditions, were populated by aquatic species such as water lilies. In the pre-drainage system, both ridges and sloughs were inundated annually, with the ridges submerged for as much as 10 months per year, while the sloughs were nearly always below the water table (McVoy et al., 2011). The ridge-and-slough terrain extended hundreds of miles from the sawgrass plains south of Lake Okeechobee to the end of Shark River Slough, covering about 1.5 million acres (McVoy et al., 2011). Because of its unusual geometry, the landscape sustained long-lived fish, alligators, and otters, and the landscape represented “a principle center for primary and secondary production and interannual survival of aquatic organisms” in South Florida’s freshwater wetlands (Ogden, 2005).
Beginning in the late 1800s, construction of canals and the lowering of Lake Okeechobee stages lowered water depths within the Everglades. Later, in the 1950s and 1960s, construction of the WCAs partially reversed some of the lowering of water depths, but at the same time distorted the spatial distribution of water depths and greatly reduced, if not eliminated, sheet flow. These water management activities disrupted important controls on ridge-and-slough landscape processes, and by the late twentieth century the degradation of ridge-and-slough patterning became widely recognized. Throughout the system, the elevation differences between ridges and sloughs are now significantly reduced relative to the pre-drainage system; the maximum elevation difference currently measured is about 0.7 feet, and the minimum is zero (McVoy et al., 2011). Sawgrass or wet prairie vegetation has expanded into some aquatic sloughs, and in some areas, the ridges and sloughs have lost their linear geometry (Figure 4-21). As of 2005, 28 percent of the original ridge-and-slough landscape was considered degraded, and another 27 percent had been drained and lost to urban or agricultural land uses (McVoy et al., 2011). Harvey et al. (2012) identified a smaller surviving percentage—about 22 percent—in those areas bounded by WCA-2, WCA-3, and Everglades National Park.
Recent scientific investigations have improved understanding of the dynamics of this complex system. Research reported by Harvey et al. (2011; summarized by Harvey et al., 2012), Larsen et al. (2009), Larsen and Harvey (2010), and McVoy et al. (2011) leads to a general model for ridge-and-slough processes, which highlights the system requirements to avert further declines and improve the condition of the remnant ridge and slough. A series of drivers are thought to contribute to the maintenance of the Everglades ridge-and-slough landscape. First, the prevailing type of water flow must be a broad, shallow distribution of water (many miles wide; i.e., sheet flow) that does not include a

FIGURE 4-21 Two aerial views of ridge-and-slough topography in WCA-3A. Above, an example of a functional ridge-and-slough system from the central part of WCA-3A showing features with distinctive linear orientation reflecting flows. Below, an example from the northern portion of WCA-3A with severely degraded ridge-and-slough topography showing lack of elongated and oriented forms and with extensive sawgrass cover.
SOURCES: Images by Christopher McVoy and SFWMD; Fling et al. (2009).
centralized and confined conveyance channel. Second, this sheet flow generally needs to be from a definable and little-changing direction. This directionality of flow is critical in creating and maintaining the linear characteristics of the ridges and sloughs so that directional flows control the system’s horizontal geometry. Third, flow velocity must be at least greater than 2.5 cm/s in order to entrain fine organic particles, or floc, from the sloughs and to redeposit them on the ridges. The ridge height then becomes limited by the depth of these relatively fast-moving sheet flows. Fourth, the depth of flow needs to fluctuate annually so that the ridges are not continuously inundated with the concomitant loss of sawgrass.
Additionally, the form of the vegetation in the sloughs exerts control over these physical processes. In pre-drainage sloughs, water depths were great enough to permit aquatic species such as water lilies to survive but not other intrusive species that have dense stem networks (McVoy et al., 2011). If the vegetation in the sloughs is too dense, then the flows are not able to entrain and redistribute the floc to the ridges, which disrupts the system dynamics. The growth of sawgrass or wet prairie vegetation in the sloughs as a result of consistently shallow water depths thus disrupts the entire ridge-and-slough process. Woody vegetation may invade ridges that are subject to long-term dry conditions. High levels of phosphorus may also stimulate the growth of invasive vegetation in sloughs, creating greater stem density and influencing the mobility of floc from sloughs to ridges. Thus, hydrologic restoration combined with water quality restoration offers the greatest prospects to improve conditions of the ridge and slough. Yet water quality restoration alone has little effect on the current downward trajectory.
Disruption of these drivers leads to continuing degradation of the ridge-and-slough landscape in the remnant Everglades (Figure 4-22). Although the difference in elevation between ridge and slough is significantly degraded throughout the system, the present condition of the characteristic ridge-and-slough patterning (plan view) within the remnant Everglades ranges from quite similar to pre-drainage patterning (e.g., south of I-75 and west of the Miami Canal), to partial disintegration of the pattern (e.g., east of Miami Canal), to complete loss of pattern (i.e., conversion to uniform stands of sawgrass, as in northern WCA-2A) (McVoy et al., 2011). Also, recent trends in the ridge-and-slough landscape are variable according to location, and the characteristic patterning can undergo significant degradation or enhancement on decadal timescales as a result of flow modifications (NRC, 2010; Sklar et al., 2009). In those cases where ridge-and-slough terrain has lost its directional alignment, resulting in irregularly shaped ridges rising above surrounding pools, the change from aligned to unaligned forms took only a few decades, essentially since the completion of the WCAs (McVoy et al., 2011).
The length of time required to restore areas of ridge and slough that are presently disappearing is unclear although losses are likely to occur more quickly than restoration. Researchers do not completely agree on how long restoration of ridge and slough might require, but computer simulation models that account for some of the system drivers suggest that many centuries would be needed (Harvey et al., 2012). Other researchers, however, including some at the public session where Harvey et al. (2012) presented their results, have indicated that more rapid restoration may be possible. In some instances where dense wet prairie vegetation now clogs the sloughs in extremely degraded areas, additional efforts, such as vegetation removal, could be required to facilitate restoration.
Tree Islands
Tree islands are “small, slightly elevated forested wetlands within a ridge-slough matrix” (Sklar et al., in review). Two major types of tree islands occur in the Everglades. Pop-up tree islands (also known as floating or barrier tree islands) originate when a large portion of peat detaches from the substrate and are colonized with shrubs and trees; these occupy Loxahatchee National Wildlife Refuge and WCA-2A. Fixed teardrop-shaped tree islands are associated with topographical variations in the mineral substrate and extend from WCA-3 to Shark River Slough in Everglades National Park (van der Valk and Sklar, 2002). Tree islands play a crucial ecological role in the Everglades, providing habitat for a range of fauna, sequestering and cycling nutrients, and contributing to the spatial heterogeneity and landscape complexity of the ecosystem (Sklar et al., 2011; van der Valk and Sklar, 2002; Wetzel et al., 2005, 2011). Tree islands also have long-established and deep-rooted cultural heritage and societal importance. Hence, maintenance and restoration of tree islands are key components of Everglades restoration.
Drainage, compartmentalization, and subsequent changes in hydrology resulted in a loss of 67 percent of tree island area from 1940 to 1995 (Figure 4-22; Sklar et al., 2005). Further declines of 6 percent in total acreage of tree islands continued between 1995 and 2004 (Figure 4-23) (Sklar et al., 2011). Although some tree islands have experienced gains in acreage, many more have declined (Figure 4-23). The conditions causing tree island degradation and decline have not abated and therefore continued declines can be expected. The greatest ongoing threats to tree islands are altered hydrological regime and altered fire regime.
Four components of hydrology—depth, hydroperiod, flow, and quality—can impact tree islands if extreme conditions outside the normal historic range are experienced for extended periods (van der Valk and Sklar, 2002). Because tree islands are nutrient hotspots and serve as nutrient sinks, water quality is regarded as the least important stressor among these components. However, water quality
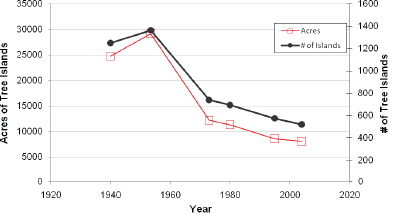
FIGURE 4-23 Tree island trends between 1940 and 2004 in WCA-3. The apparent increase in acreage and number of islands between 1940 and the mid-1950s is likely a combination of differences in mapping techniques and increases in areas covered by trees and shrubs (e.g., colonization of ridges) after extensive drydowns and shortened hydroperiods.
SOURCE: F. Sklar, SFWMD, personal communication, 2010, 2012.
is an important factor in the accumulation and distribution of phosphorus on tree islands (Wetzel et al., 2005, 2011).
High water depths and long periods of inundation or flooding cause plant stress, failure of seed germination, and diminished wading bird nest habitat (Sklar and van der Valk, 2002). Simulations with the Everglades Landscape Vegetation Model (ELVM; a simulation model that links fire, nutrient dynamics, hydrological regimes, and vegetation succession on tree islands to analyze management alternatives) suggested that in WCA-2A tree island water depths of 30 cm for longer than 150 days result in loss of tree island species, and these results align well with historical records (Wu et al., 2002). Nevertheless, for Shark River Slough tree islands, water levels deep enough to protect the thin peat covering from both microbial oxidation and fires were critical to the elevated areas’ ability to support woody species.
Shallow water depths and reduced hydroperiods result in peat oxidation and muck fires, which lower the elevations of peat-based tree islands. The overall effect and legacy of the 1915 to 1950s period of uncontrolled Everglades drainage was a flattening of the landscape: lowering the tree islands and the ridges
relative to the sloughs and moving the elevations of tree islands closer to those of ridges (McVoy et al., 2011). Subsided tree islands with reduced peat become more vulnerable to flooding even under normal water depths and particularly under restored conditions. Peat oxidation and loss of up to 1 cm per year can occur under extended periods of shallow water depth (F. Sklar, SFWMD, personal communication, 2011), with greater rates in a muck fire. The May 1981 muck fires in the northern portion of WCA-3A resulted in 9 to 28 cm of peat loss (Wetzel, 2002). In Shark River Slough 55 percent of islands and 58 percent of tree island hectares have been lost because of peat oxidation caused by fires and lack of water (Sklar, 2012). Fire frequency in the Everglades is estimated to be approximately 10-14 years (Gunderson and Snyder, 1994), but drier conditions increase the frequency, size, and intensity of fires. Under wetter conditions fire is a natural part of the disturbance regime and does not have the devastating effects evident with the large, frequent, and high-intensity fires under drier conditions.
WCA-3A currently experiences hydrologic extremes to differing degrees. In general, the northern part of WCA-3A has become drier, whereas tree islands in the southern areas experience higher water depths, ponding, and longer hydroperiods. Wetzel (2002) reported that tree island peat depths are generally shallower in northern WCA-3A (0.62-1.08 m) compared to southern WCA-3A (1.08-1.22m) as a result of peat oxidization. The northern tree islands in WCA-3A have also become more vulnerable to fires than their southern counterparts.
If water depths are substantially reduced for extended periods of time, then peat will oxidize, lowering the elevation of the island. The island then becomes more susceptible to inundation and drowns, reducing the diversity of floral species on the islands to those that are flood tolerant (Wetzel, 2002). However, even woody species that are flood tolerant can perish under sustained extreme flooding events, resulting in “ghost islands” that are lacking in floral diversity; it is estimated that the drowning process can take 20 years (Sklar, SFWMD, personal communication, 2011). Eventually, if tree islands subside to the extent that they lose their elevation above the surrounding ridges, then they can no longer support woody vegetation, and the vegetation is replaced by sawgrass and cattails.
If existing conditions are maintained, then the decline of tree islands will continue. Unless decompartmentalization occurs, tree islands in the southern portions of WCA-3A will continue to experience inundation and ponding and to lose species diversity (shifting to more flood-tolerant species). Hydrologic restoration (e.g., increased flow volumes, more natural hydroperiods and water depths) offers opportunities for recovery of tree islands, particularly in the southern and central portions of WCA-3A, where the greatest number of higher elevation tree islands remains. Much of the tree island acreage in northern WCA-3A has already been lost because of peat subsidence, and some remaining subsided islands may experience greater inundation, reduction in floral diversity,
and ultimate loss with hydrologic restoration. Nevertheless, there is substantial variability in the current elevations of tree islands, which will result in different responses to restored water depths. With hydrologic restoration, many tree islands that are currently on a trajectory of drowning can recover, particularly if their elevation differences remain, although active restoration in the form of tree planting may be needed in some cases (van der Valk and Sklar, 2002). Natural recolonization of degraded islands can occur through seed dispersal if there are nearby islands with sufficient diversity and abundance of species (van der Valk and Sklar, 2002; Wetzel et al., 2005; Wu et al., 2002). Nevertheless, with long-term flooding and associated declines in plant diversity, opportunities for natural recovery through natural seed dispersal and recolonization will be lost over time. It may be possible to restore severely subsided tree islands by raising the elevation of their heads, although such efforts would be expensive and labor intensive (van der Valk and Sklar, 2002).
The restoration of hydrologic flow, water depths, and duration will benefit many tree islands, but high phosphorous levels may promote cattail and willow encroachment on islands with elevations that are low relative to the surrounding marshes. However, as phosphorus increases, vegetation and peat increase on tree islands that sequester and redistribute phosphorus (Wetzel et al., 2011). Hence, the detrimental effects of altered water depths and duration are expected to exceed the effects of water quality on tree islands. Improved hydrology and water quality offer the best opportunity for tree island restoration across the landscape (Bedford et al., 2012), although some subsided islands may become inundated and lose floral diversity because of variations in tree island elevation across the landscape. When considering restoration alternatives, the choice does not appear to be one of causing harm versus not, but instead one of causing the least overall harm while promoting the most improvement.
ELVM simulations suggest that restoration of 60 percent of tree islands known to be lost could occur within 50 years (Wu et al., 2002). However, there are reasons to believe that the time to recover tree islands in WCA-3A is substantially underestimated. The predictions were based on flows that had been increased to pre-drainage levels and not the lower flows proposed under the CERP. Furthermore, the model was designed for, and calibrated well with, tree island dynamics in WCA-2A, but it did not calibrate well with islands in WCA-3, in part because WCA-3 has experienced substantial fire-induced peat losses that were not explicitly modeled.
Snail Kite
Snail kites in the Everglades are part of a subspecies (Rostrhamus sociabilis plumbeus) that includes other populations in Cuba and northwestern Honduras
(Sykes et al., 1995). The two other subspecies of snail kite extend through Mexico, Central America, and all of South America. The Everglade kites are the only snail kites in the United States, and the population has been designated as endangered because of its limited distribution and declining numbers.
The decline of the snail kite in South Florida reflects the degradation of the ecosystem on which it depends. The committee’s previous report (NRC, 2010) discussed the decline of the snail kite over the past decade, which has reduced the population to an extremely low level (Figure 4-24). Kite populations have fluctuated historically in response to drought cycles, with prior low points in the 1960s (Takekawa and Beissinger, 1989) and late 1980s (Beissinger, 1995), but the current decline differs in being driven by degradation of habitat in previously productive areas (e.g., Lake Okeechobee, WCA-3A) as well as by climate (Reichert et al., 2011). Kites are highly mobile, and they move throughout the system to find conditions favorable for foraging and breeding (Bennetts and Kitchens, 1997; Takekawa and Beissinger, 1989). However, in recent years con-
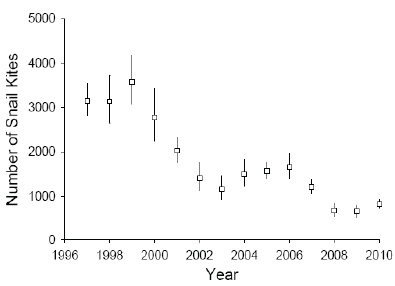
FIGURE 4-24 Population size and associated 95 percent confidence intervals of snail kites, 1997-2010.
SOURCE: Reichert et al. (2011).
ditions have generally been unfavorable everywhere, and the kite population in the Everglades has declined precipitously. One can argue that the current trajectory of the kite population, which has brought it to the brink of extirpation, mirrors the current trajectory of the ecosystem and reflects the fact that every part of the Everglades has been altered, such that the kites have increasing difficulty finding suitable conditions anywhere (Kitchens et al., 2002).
The most recent decline of the snail kite has been specifically linked to unfavorable conditions in southwestern WCA-3A, their primary nesting area during the past decade (see Endangered Species Issues in Chapter 3). Prolonged high water levels in southern WCA-3A have well-documented, adverse effects on kites (NRC, 2010). However, the problem is more complicated than wet season water levels that are too high and last too long. These conditions and the accompanying loss of tree islands that serve as nesting sites might explain the lack of kite nesting in some former nesting areas, such as eastern WCA-3A (Figure 4-25). However, kites also suffer from dry season lows that are too low and rates of recession that are too fast (FWS, 2010). Kites are highly specialized feeders, relying on apple snails (Pomacea paludosa) to feed themselves and their young. Not coincidentally, these snails are also adversely impacted by these same hydrological problems, that is, prolonged wet season high water, prolonged dry downs in the dry season, and rates of recession that are too fast (FWS, 2010).
It is actually rapid rates of recession and low water levels in the dry season, not prolonged high water during the wet season that explain poor nesting success in southwestern WCA-3A over the past decade (NRC, 2010). It is the minimum stage, not the wet season high water maximum, that is most highly (and positively) correlated with kite nesting success (Cattau et al., 2008; FWS, 2010). Historically kite numbers have decreased during droughts and increased during wet periods (Takekawa and Beissinger, 1989). This explains the seemingly paradoxical pattern that kite nesting, which is adversely affected by prolonged high water, is concentrated in southern WCA-3A, where water levels are the highest, rather than in central or northern WCA-3A. When the kites shifted away from the ponded areas in eastern WCA-3A in the 1980s, they initially moved to central WCA-3A (Figure 4-25). However, these areas now dry out too much and too fast to support kite nesting. Thus the kites have shifted to the seemingly unsuitable southwestern portion of WCA-3A, not because conditions there are ideal, but because conditions everywhere else in WCA-3A are even worse. In some places prolonged high water has converted the wet prairies and emergent marshes that the kites use for foraging to other habitat types (Holling et al., 1994; Sklar et al., 2001; Zweig and Kitchens, 2008). These habitat types still occur in abundance; however, the more pervasive problem is that the historical wet season/dry season water cycles that support kite nesting and large apple snail populations (SEI, 2007) no longer reliably occur in these habitats.
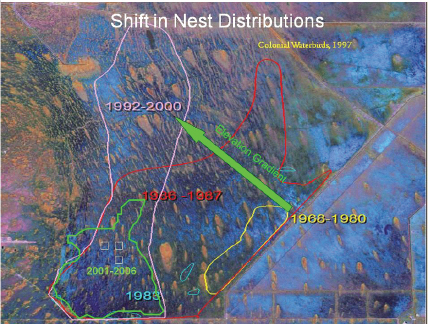
FIGURE 4-25 Shifts in the distribution of nesting snail kites in WCA-3A, 1968-2006.
SOURCE: FWS (2010).
Looking beyond WCA-3A, the picture is much the same. The kites formerly nested in large numbers in the Kissimmee Chain of Lakes, Lake Okeechobee (Cattau et al., 2009), and WCA-3B and WCA-2 (Bennetts et al., 1994; Sykes, 1983). Thus, the dependence of kite reproduction on WCA-3A is a relatively recent phenomenon (Figure 4-26), and the dependence on the southwestern portion of WCA-3A is even more recent (Figure 4-25). In most of these former nesting areas, hydrology or habitat (or both) are altered in ways that make it unlikely that kites will return in significant numbers without restoration. Lake Okeechobee continues to be unproductive under current water management, although there were a few nests there in 2010 following three years with no nests (Figure 4-26). WCA-2 has experienced extensive loss of the tree islands (Sklar et al., 2009), which kites use as nesting sites, and no nesting has occurred there
FIGURE 4-26 Number of young fledged from kite nests in the Water Conservation Areas (WCA), Kissimmee Chain of Lakes (KCOL), and Lake Okeechobee (Okeechobee), 1992-2009.
SOURCE: Cattau et al. (2009).
in recent years. The ridge-and-slough landscape of WCA-3B is highly degraded (SCT, 2003), and there have been a few nests there in some years and none in others. Some nesting has occurred in Everglades National Park, but the dry season water levels there tend to be too low. Interestingly, kites have resumed nesting in the Kissimmee Chain of Lakes, coincident with their increasing ability to forage on an invasive apple snail species (Pomacea insularum) found in abundance there (see below). This area has been the primary nesting area during the past few years because productivity in WCA-3A has declined to near zero (Figure 4-26).
The kites are not as directly impacted by deterioration of water quality as are many other fauna and flora of the Everglades, although they can be indirectly impacted by changes in habitat mediated by water quality, such as cattail invasion. Instead, the problems that have plagued the kites in various areas of the central Everglades result from altered flow regimes; that is, they are problems of water quantity and distribution, especially seasonal cycles, rather than water
quality. Restoration of historical seasonal cycles of water levels and recession is necessary not only to create suitable nesting conditions for kites, but also to support the life cycle of their apple snail prey. Snail kites are very successful in other areas with extensive, shallow wetlands (e.g., the Llanos of Venezuela and the Pantanal of Brazil), and although little is known about where snail kites were most successful in the pre-drainage Everglades and to what extent their distribution changed during wet and dry cycles, the committee judges that conditions for the kite should improve in the Everglades with system-wide hydrologic restoration. Two independent panels of ornithologists and wetland experts have reached the same conclusion (SEI, 2003, 2007). The strength of the kites’ response will be complicated by the fact that restoration might reduce the amount of preferred wet prairie habitat while increasing the quality (due to restored hydrological cycling) of remaining habitat.
Conservation increasingly focuses on those few areas that remain potentially suitable for the snail kite. Recent changes in water management in WCA-3A focus on improving conditions for kites and apple snails in the area on which kites have become most dependent, that is, southwestern WCA-3A (see Chapter 3). This likely will improve the kites’ nesting success in the target area (southern WCA-3A) but at the expense of making conditions even worse for them (and other system components) in other areas (central and northern WCA-3A). Until more substantial progress is made with all that the CERP is designed to accomplish in the central Everglades—increased inputs of water, a shift in the distribution of water from west to east, restoration of sheet flow and historic seasonal cycles of water levels and recession—kite conservation likely will remain in crisis as the system continues to degrade. Local actions, such as in WCA-3A, may ward off extirpation, but having a viable population is likely contingent on system-wide restoration. Not until then will the kites’ mobility and resiliency become the assets they once were.
The kites’ adaptability may enable them to persist despite continuing system degradation. Specifically the kites appear to be increasingly able to sustain themselves on exotic apple snails: recent increases in nesting in the Kissimmee Chain of Lakes (Cattau et al., 2009) and in the STAs (see Chapter 3) involve use of this prey. The kites may be adapting to these large snails by foraging for juveniles (Cattau et al., 2009) and by adults feeding snails to nestlings (Williams, 2011), and even by behavioral changes in prey handling that enable the kites to extract the large exotic snails more efficiently (H. Tipton, FWS, personal communication, 2011).
CONCLUSIONS
An assessment of the status and trajectories of 10 ecosystem attributes reveals that conditions for tree islands, ridge-and-slough landscape, snail
kites, and peat continue to degrade and that cattail coverage continues to expand 12 years after the initiation of the CERP. These declines can be attributed to altered hydrology and/or the elevated supply of phosphorus in the remnant Everglades. Despite its ability to search throughout the Everglades ecosystem for suitable conditions, the Everglade snail kite has experienced a precipitous decline in numbers over the past 15 years and is in danger of extirpation.
The state’s extensive phosphorus control efforts over the past two decades appear to be stabilizing or improving the current trends for several ecosystem components driven by phosphorus (e.g., periphyton, soil P). Cattail expansion, however, is continuing but at a decreasing rate in some areas (e.g., WCA-2). Implementation of STAs and best management practices has markedly decreased phosphorus loads to the WCAs, and interior phosphorus concentrations have decreased in WCA-2 and -3 in response to decreases in the concentrations of inflowing waters. Despite this progress, impacted areas of the WCAs consistently fail the four-part test for compliance with Florida’s water quality standards. Thus, it is widely recognized that additional water quality improvements are needed to prevent further degradation and reverse ongoing adverse impacts to the ecosystem caused by elevated phosphorus.
In contrast, the restoration of flows in the central Everglades has been limited, and the ecosystem attributes most directly influenced by hydrologic factors continue to decline. In many cases these ongoing losses can only be recovered over long time scales. The velocity, depth, and duration of water in the Everglades are important controlling factors for the distinctive terrain of the Everglades: tree islands, ridge-and-slough topography, and peat accumulations. These landscape components have been severely degraded by flow alterations during past decades. Recovering additional losses will require decades if not centuries. Of the many projects under construction, only Mod Waters (a non-CERP project) and the C-111 Spreader Canal (a CERP project) offer promise of direct, significant effects in the central Everglades.
Substantial near-term progress to address both water quality and hydrology in the central Everglades is needed to prevent further declines. Near-term progress that addresses only water quality or water quantity leads to continued system declines of many components. Additionally, many improvements in water quality are linked with improvements in water quantity. Thus, decisions on restoration project design and scheduling should not be viewed as simple tradeoffs between water quantity and water quality. Instead, this qualitative analysis points to the need for a more critical and comprehensive quantitative analysis using models and field data to evaluate management alternatives in an integrated manner (see Chapter 5). Also, it highlights the importance of stabiliz-
ing and ultimately reversing declines of attributes that would take a long time to recover, particularly if other aspects of the restoration depend on them. Because of its focus on the remnant Everglades and accelerated planning, the Central Everglades Planning Project conceptually provides promise for rehabilitating the remnant Everglades.






















































