
The constraints on how human and animal minds process these different quantities are similar (Gallistel and Gelman, 1992). For example, all of these quantities show cognitive processing limitations that can be predicted by Weber’s law. Weber’s law states that quantity discrimination is determined by the objective ratio between their values. This ratio-based psychological and neural signature of quantity processing indicates that many quantities are represented in an analog format, akin to the way in which a machine represents intensities in currents or voltages (Gallistel and Gelman, 1992). I discuss the types of constraints that influence quantity discrimination, using “number” as the initial example, and then consider the psychological and neural relationship between “number” and other quantitative dimensions. Similar constraints on processing across different quantities have been interpreted as evidence that they have a common evolutionary and/or developmental origin and a common foundation in the mind and brain (Zorzi et al., 2002; Walsh, 2003; Pinel et al., 2004; Feigenson, 2007; Ansari, 2008; Cohen Kadosh et al., 2008; Cantlon et al., 2009c; de Hevia and Spelke, 2009; Lourenco and Longo, 2011; Bonn and Cantlon, 2012). The resolution of these issues is important for understanding the inherent organization of our most basic conceptual faculties. The issue is also important for understanding how our formal mathematical abilities originated.
Primitive quantitative abilities play a role in how modern humans learn culture-specific, formal mathematical concepts (Gallistel and Gelman, 1992). Preverbal children and nonhuman animals possess a primitive ability to appreciate quantities, such as the approximate number of objects in a set, without counting them verbally. Instead of counting, children and animals can mentally represent quantities approximately, in an analog format. Studies from our group and others have shown that human adults, children, and nonhuman primates share cognitive algorithms for encoding numerical values as analogs, comparing numerical values, and arithmetic (Meck and Church, 1983; Gallistel, 1989; Feigenson et al., 2004; Cantlon et al., 2009c). Developmental studies indicate that these analog numerical representations interact with children’s developing symbolic knowledge of numbers and mathematics (Gelman and Gallistel, 1978; Feigenson et al., 2004). Furthermore, the brain regions recruited during approximate number representations are shared by adult humans, nonhuman primates, and young children who cannot yet count to 30 (Dehaene et al., 2003; Nieder, 2005; Ansari, 2008). Finally, it has recently been demonstrated that neural regions involved in analog numerical processing are related to the development of math IQ (Halberda et al., 2008). Taken together, current findings implicate continuity in the primitive numerical abilities that are shared by humans and nonhumans, as well as a degree of continuity in human numerical abilities ranging from primitive approximation to complex and sophisticated math.
The fact that humans have been recording tallies with sticks and bones for 30,000 years is impressive, but the critical issue is this: what cognitive abilities enabled them to encode quantities in the first place? To identify the inherent constraints on humans’ ability to process numerical information, it is helpful to consider the evolutionary history of numerical thought. We can look for clues to the evolutionary precursors of numerical cognition by comparing human cognition with nonhuman primate cognition. The degree to which humans and nonhuman primates share numerical abilities is evidence that those abilities might derive from a common ancestor, in the same way that common morphology like the presence of 10 fingers and toes in two different primate species points to a common morphological heritage.
So far, there is evidence that nonhuman primates share three essential numerical processing mechanisms with modern humans: an ability to represent numerical values (Brannon and Terrace, 1998; Nieder, 2005; Cantlon and Brannon, 2006, 2007b), a general mechanism for mental comparison (Cantlon and Brannon, 2005), and arithmetic algorithms for performing addition and subtraction (Beran and Beran, 2004; Cantlon and Brannon, 2007a). These findings compliment and extend a long history of research on the numerical abilities of nonhuman animals [see Emmerton (2001) for review].
Representation
When adult humans and monkeys are given a task in which they have to rapidly compare two visual arrays and touch the array with the smaller numerical value (without counting the dots), their performance reliably yields the pattern shown in Fig. 16.1: accuracy decreases as the ratio between the numerical values in the two arrays approaches 1 [Cantlon and Brannon (2006); see Dehaene (1992) and Gallistel and Gelman (1992) for review]. The explanation of this performance pattern is that both groups are representing the numerical values in an analog format (Fig. 16.2).
In an analog format, number is represented only approximately, and it is systematically noisy (Dehaene, 1992; Gallistel and Gelman, 1992]. More precisely, the probability of noise (i.e., the spread of the distributions) in the subjective representation of a number increases with the objective number of items that are coded by that representation. Consequently, the probability of confusion (i.e., the overlap between distributions) between any two objective numbers increases as their value increases. This means that the probability of having an accurate subjective representation of a numerical value decreases with its objective value. This relationship can be succinctly quantified by the ratio between the numerical values being
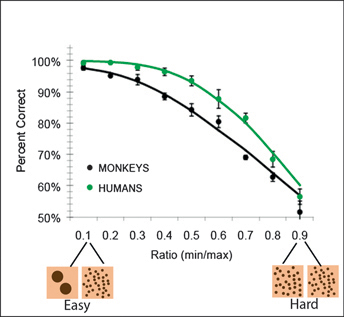
FIGURE 16.1 Accuracy on a numerical discrimination task for monkeys and humans plotted by the numerical ratio between the stimuli. From Cantlon and Brannon (2006).
compared. Two different pairs of numerical values that have the same ratio (e.g., 2 and 4, 4 and 8) have the same amount of overlap, or the same probability of confusion. As numerical pairs get larger and closer together, their ratio increases and so does the probability that they will be confused (leading to more errors). For example, one might be 80% accurate at choosing the larger number when the numerical choices are 45 vs. 70 (45/70 = a 0.64 ratio) but might perform at chance when the choices are 45 vs. 50 (45/50 = a 0.9 ratio). This effect is known as Weber’s law. The curves in Fig. 16.1 [from Cantlon et al. (2009c)] represent predicted data from a model of number representation under Weber’s law (Pica et al., 2004), and they show that the predictions of this analog numerical model fit the data well.
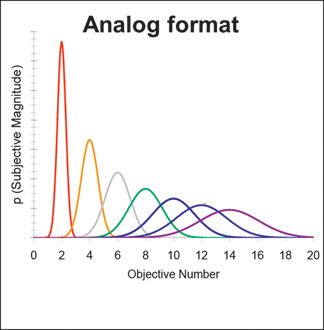
FIGURE 16.2 An analog representation of numerical value represents an objective numerical value with a probability distribution that scales with the size of the objective numerical value. From Cantlon et al. (2009a). Reprinted with permission from the American Association for the Advancement of Science.
The empirical data from monkeys and humans and the fit of the analog model demonstrate that although humans have a means of representing numerical values precisely using words and Arabic numerals, they still have an approximate, analog numerical system that functions essentially in the same way as in monkeys.
Comparison
The ratio effect, described by Weber’s law, indicates that numerical values can be represented in an analog format. However, that does not tell us anything about the process by which two numerical values are compared. We have identified a signature of mental comparison in monkeys that is commonly observed when adult humans make judgments of magnitudes: the semantic congruity effect (Cantlon and Brannon, 2005; Holyoak, 1977). The semantic congruity effect is a response time effect that is observed in adult humans’ response times whenever they have to compare things along a single dimension. For instance, when people are presented with pairs of animal names and asked to identify the larger or smaller animal from memory, they show a semantic congruity effect in their response time: people are faster to choose the smaller of a small pair of items (e.g., ant vs. rat) than they are to choose the larger of that pair. However, for pairs of large items (e.g., horse vs. cow), people are faster to choose the larger item than the smaller item. This effect suggests that the physical size of the animal interacts with the “size” of the question (whether “Which is larger?” or “Which is smaller?”) in subjects’ judgments. In humans, the semantic congruity effect is observed for judgments of many dimensions, including judgments of numerical values, from Arabic numerals. We found that this effect is also observed in monkeys when they compare numerical values from arrays of dots. Monkeys performed a task in which they had to choose the larger numerical value from two visual arrays when the background color of the computer screen was blue, but when the screen background was red, they had to choose the smaller numerical value of the two arrays. As shown in Fig. 16.3 [from Cantlon and Brannon (2007a)], both monkeys showed a crossover pattern of faster response times when choosing the smaller of two small values compared with the larger of two small values, and the opposite pattern for large values. The semantic congruity effect is the signature of a mental comparison process wherein context-dependent mental reference points are established (e.g., 1 for “choose smaller” and 9 for “choose larger”), and reaction time is determined by the distance of the test items from the reference points; this has been modeled as the time it takes for evidence to accrue in the comparison of each item to the reference point (Holyoak, 1977). In humans the semantic congruity effect is observed for a variety
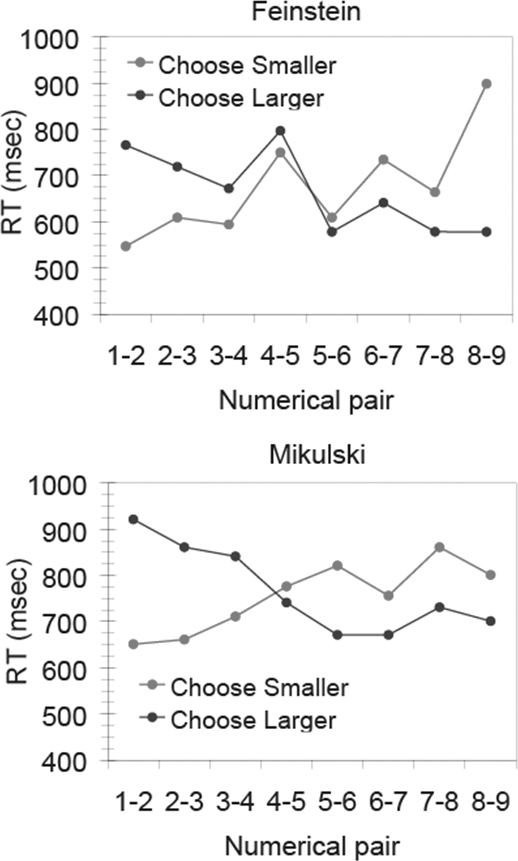
FIGURE 16.3 The semantic congruity effect in the response times of two different monkeys (Feinstein and Mikulski) on a numerical comparison task where they sometimes chose the larger numerical value from two arrays (dark line) and other times chose the smaller value (light line). The cross-over pattern reflects the effect of semantic congruity. From Cantlon and Brannon (2005).
of mental comparisons from both perceptual and conceptual stimuli: brightness, size, distance, temperature, ferocity, numerals, etc. Our data from nonhuman primates indicate that the mental comparison process that yields the semantic congruity effect is a primitive, generalized, nonverbal mental comparison process for judging quantities and other one-dimensional properties.
In fact, the ability to compare quantities, and the proposed algorithm underlying that ability, could be so primitive that it extends to nonprimate animals. A recent study by Scarf et al. (2011) showed that pigeons can compare numerical values, and in doing so they represent an abstract numerical rule that can be applied to novel numerical values. Pigeons’ accuracy on that ordinal numerical task is comparable to that of monkeys tested on an identical task (Brannon and Terrace, 1998).
Arithmetic
Arithmetic is the ability to mentally combine values together to create a new value without having directly observed that new value. We have found that monkeys possess a capacity for basic, nonverbal addition that parallels human nonverbal arithmetic in a few key ways (Cantlon and Brannon, 2007a). First, monkeys and humans show a ratio effect when performing rapid nonverbal addition, similar to the ratio effect described earlier. Monkeys’ and humans’ accuracy during arithmetic depends on the ratio between the values of the choice stimuli. We also observed a classic signature of human arithmetic in monkeys’ performance: the problem size effect. Adult humans typically exhibit a problem size effect wherein performance worsens as the problem outcome value increases (Campbell, 2005). Like humans, monkeys exhibited a problem size effect in their addition accuracy (even when controlling for the ratio effect).
However, there are also important and potentially informative differences between the performance of humans and monkeys. Adult humans and young children show a practice effect in their arithmetic performance wherein performance on a specific problem improves the more that it is practiced (Campbell, 2005). Monkeys do not show a practice effect for specific problems. This was the case even over 3 years of practice on a specific problem (Fig. 16.4 shows performance for two monkeys, over 3 years of testing on 1 + 1, 2 + 2, and 4 + 4). Nonhuman primate arithmetic thus parallels human nonverbal arithmetic in the ratio and problem size effects but not the practice effect, which has been observed primarily in symbolic arithmetic performance in humans. Presumably, discrete symbols are necessary for humans to encode arithmetic problems in a format that is amenable to memorization, which is why monkeys do not show a practice effect.
The overarching conclusion from this line of research is that the abilities to represent, compare, and perform arithmetic computations reflect a cognitive system for numerical reasoning that is primitive and based on analog magnitude representations. However, if analog numerical cognition is truly “primitive” and homologous across primate species, then it should be rooted in the same physical (neural) system in monkeys and
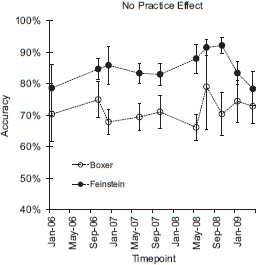
FIGURE 16.4 The lack of a practice effect in monkeys’ addition performance over 3 years. Data from Cantlon and Brannon (2007a).
humans. In fact, there is evidence from multiple sources that analog numerical processing recruits a common neural substrate in monkeys, adult humans, and young children (Fig. 16.5).
In monkeys who are trained to match visual arrays of dots according to number, single neurons along the intraparietal sulcus (IPS) will
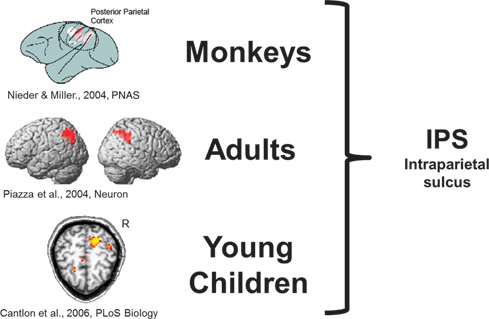
FIGURE 16.5 Monkeys, human adults, and human children exhibit similar activation in the IPS during analog numerical processing. Redrawn from Nieder and Miller (2004), Piazza et al. (2004) (reprinted with permission from Elsevier, Copyright 2004), and Cantlon et al. (2006).
respond maximally to a preferred numerical value, and their firing rate decreases as the number that is presented gets numerically farther from that preferred value (Nieder and Miller, 2004). This neural firing pattern has been linked to the behavioral ratio effect and is thought to reflect analog numerical tuning in the IPS. A similar pattern of numerical tuning has been observed with functional MRI in the human IPS. Manuela Piazza et al. (2004) found a neural adaptation effect for numerical values in the IPS that depended on the ratio between the adapted numerical value and a deviant numerical value. Our group also observed neural adaptation in the IPS for numerical values ranging from 8 to 64 in preschool children who could not yet verbally count to 30 (Cantlon et al., 2006). Together, these studies reflect a common neural source for analog numerical representation that bridges species as well as stages of human development and is thus independent of language and formal mathematics experience. These neural data support the conclusion derived from the behavioral data that there is continuity between humans and nonhuman animals in the mechanisms underlying analog numerical representations.
THEN THERE WERE SYMBOLS
A long history of studies with preverbal human infants has shown that they too possess an ability to quantify objects with approximate, analog representations (Feigenson et al., 2004). Thus, there is general agreement that the analog system for numerical reasoning is primitive in human development. A fundamental question is how a child’s developing understanding of numerical symbols interfaces with preverbal analog representations of number. Of particular interest is how children initially map numerical meanings to the first few symbolic number words (Gelman and Gallistel, 1978; Wynn, 1990; Gelman and Butterworth, 2005; Le Corre and Carey, 2007; Piazza, 2010). There is currently a debate over the types of preverbal numerical representations that form the initial basis of children’s verbal counting. However, regardless of how this initial mapping transpires, behavioral evidence suggests that as children learn words in the counting sequence, they map them to approximate, analog representations of number (Wynn, 1992; Lipton and Spelke, 2005; Gilmore et al., 2007). Lipton and Spelke (2005) found that 4-year-old children could look at a briefly presented array of 20 dots and, if they could count to 20, they could verbally report (without counting) that there were 20 dots in the array, and their errors were systematically distributed around 20 (i.e., their errors exhibited a numerical ratio effect). If they could not yet count to 20, however, they responded with random number labels. Thus, as soon as children learn a particular verbal count word in the sequence, they know the approximate quantity to which it corresponds without counting, suggesting
that number words are attached to the analog numerical code as soon as they are learned. These data have been taken to indicate that analog numerical representations are used to assign semantic meanings to numerical symbols over human development. There is also evidence that children who have learned to count verbally, but have not yet learned to add and subtract, psychologically “piggyback” on analog arithmetic representations as they transition to an understanding of exact symbolic arithmetic (Gilmore et al., 2007). The general conclusion that then emerges is that the cognitive faculties that children initially use for nonsymbolic, analog numerical operations (and which they share with nonhuman animals) provide a scaffolding for verbal counting in early childhood.
IS “NUMBER” ALONE?
The data from the development of counting in early childhood make the case that a primitive numerical system is conceptually transformed into a system for symbolic numbers. However, how do we know that analog numerical representations are the sole precursors of formal, symbolic numerical cognition? Currently, we do not. Although numerical reasoning seems to be primitive in the sense that it is shared among primate species, other quantitative abilities are just as widespread. For instance, the abilities to judge nonnumerical intensities such as size, time, brightness, height, weight, velocity, pitch, and loudness are as common among animal species as the ability to judge numerical values. Furthermore, all of these quantities can be discriminated by human infants, and discriminations among instances from those continua bear many of the same properties and signatures as numerical discrimination [e.g., ordinality, Weber’s law, the semantic congruity effect, arithmetic transformations; see Feigenson (2007) for review]. In adults, all of these dimensions are effortlessly mapped to numerals. For example, adult humans can represent loudness, handgrip pressure, time, size, and brightness as numerical values. Finally, evidence from the semantic congruity effect (described earlier) suggests that many different quantitative dimensions are mentally compared by a common process. The modularity and taxonomy of analog numerical representations is a central issue for understanding the development and origins of numerical and mathematical cognition. Here I discuss relations between numerical cognition and other quantitative dimensions, such as size, length, duration, brightness, pitch, and loudness.
Until recently, the cognitive and neural mechanisms of numerical cognition were considered to be specialized processes. Neuropsychological and neuroimaging studies of adult humans have shown that numerical knowledge dissociates from other forms of semantic knowledge, and it has been argued from those data that the processes subserving numerical
knowledge are domain specific [see Dehaene et al. (2003) for review]. For example, individuals with semantic dementia, resulting from left temporal lobe atrophy, exhibit severe impairments on picture and word naming tasks but can be spared for number tasks (Cappelletti et al., 2001). The opposite disorder of impaired numerical cognition but spared semantic and linguistic knowledge has also been demonstrated (Warrington, 1982; Cipolotti et al., 1991). Moreover, in cases of developmental dyscalculia, mathematical reasoning can become selectively impaired over development (without impairments to other aspects of reasoning). Furthermore, developmental dyscalculia is coupled with atypical anatomy and functional responses in the IPS (Molko et al., 2003; Price et al., 2007). The fact that focal brain injuries and developmental impairments, perhaps especially to the IPS, specifically impair numerical reasoning indicates that at some level of cognitive and neural processing, numerical computation is independent. However, it remains unclear what aspects of numerical processing operate independently of other psychophysical and conceptual domains. Most previous neuropsychological and neuroimaging studies controlled for many nonnumerical abilities (eye movements, spatial attention, memory, semantic knowledge), but they did not test performance on continuous dimensions other than number (length, area, brightness, etc.). Thus we cannot know whether other quantitative abilities were simultaneously impaired in many of those neuropsychological patients.
Recently, Marco Zorzi et al. (2002) found that representations of spatial and numerical continua can be jointly impaired in patients with right parietal lesions and hemispatial neglect; patients not only neglect the left visual field and place the midpoint of a line right of center in a line bisection task, but they also overestimate the middle value of two numbers in a numerical bisection task. The patients thus neglect both the left side of a line and the left side of their mental representation of the numerical continuum. This finding and several others have led to proposals that concepts of “space” and “number” are interrelated (Walsh, 2003; Pinel et al., 2004).
The degree to which “space” (e.g., size, height, or length) interacts with numerical information is currently being investigated with a range of methods [see Walsh (2003), Cantlon et al. (2009c), and Lourenco and Longo (2011) for reviews]. One view is that space and number have a biologically privileged psychological relationship (Dehaene et al., 2008; de Hevia and Spelke, 2009, 2010). Evidence for this view comes from developmental studies of number and space representation (de Hevia and Spelke, 2009, 2010). In line-bisection tasks, incidental displays of dot arrays presented at the endpoints of the line systematically distort preschoolers’ perception of the line’s midpoint; subjects bisect the line asymmetrically toward the larger number of dots (de Hevia and Spelke, 2009). In addition, infants spontaneously map number onto space when habituated to positively
correlated number/line-length pairs (de Hevia and Spelke, 2010). The fact that infants map number onto space within the first months of life has been used to argue for an innate bias to relate space and number.
Biologically privileged relations between space and number are also indicated by the universality of their association (Dehaene et al., 2008). The ability to map numbers onto space (number lines) is widespread among human cultures. The Mundurucu, an Amazonian people who lack a rich linguistic system for discrete number words or symbols, can place sets of objects that vary in numerical value onto horizontal lines in numerical order (just as Western subjects do). That finding supports the conclusion that mapping between space and number is not culturally determined by reading and reciting numerical symbols, because Mundurucu do not generally use such symbols. However, this finding does not necessarily indicate the presence of an innate bias to map numbers to space in humans, but may represent an analogical relation between the ordinal properties of the stimuli or the primacy of “space” alone (Cantlon et al., 2009a). In support of those alternatives, there is evidence that a similar mapping to space is made with representations of pitch in typical adults from Western cultures (Rusconi et al., 2006). If pitch shows the same kind of relation to space as number does, then a biologically “privileged” relation between space and number seems less likely. One possibility is that the relationship is ubiquitous among any of a number of dimensions (e.g.., pitch, number, length, loudness, etc.). Alternatively, number and space and pitch and space could be related because of a privileged representation of space alone, which grounds a number of quantitative representations.
Several researchers have suggested deep psychological interactions not just between number and space but among many quantitative dimensions. In their review of behavioral data from humans and other animals, Gallistel and Gelman (2000) argued that although number is objectively a discrete property, it should be represented with an analog magnitude code. They argued that animals must combine discrete number with continuous quantities in making decisions. For example, they observed that animals need to combine estimated time and amount of potential food in making foraging decisions (i.e., for “rate”). Because natural numbers are discrete and time is continuous, combining information from these incompatible formats necessitates conversion to a common analog format. The same argument could be applied to “density,” which integrates information about number and surface area. This idea implicates the possibility of common representations and shared computations for multiple quantities.
Studies in young children provide evidence that different quantitative representations have a common foundation, in the sense that they develop together. As described earlier, numerical discriminations are modulated by the ratio between the values, as per Weber’s law. In human infants,
the ratio effects for judgments of size, time, and number are refined at a similar rate of development (Brannon et al., 2006; vanMarle and Wynn, 2006; Feigenson, 2007). Infants’ discriminations of size, time, and number improve by approximately 30% between 6 and 9 mo of age. Similarly, in children, the precision of numerical discrimination improves from ages 6 to 8 years, and the discrimination of luminance, duration, and length systematically follow the same developmental trajectory (Holloway and Ansari, 2008; Volet et al., 2008). Because they develop at the same rate, it is likely that either the same mechanism underlies the different abilities or that different mechanisms are subject to the same constraints. The developmental trajectories of the discrimination of other quantities, such as loudness, pitch, pressure, temperature, density, motion, and saturation, have not been tested. However, there is evidence that young children and even infants can form compatible representations across many of these different dimensions (Smith and Sera, 1992; Gentner and Medina, 1998; Mondloch and Maurer, 2004; Walker et al., 2010).
As mentioned earlier, the dimensions of space and number can be related to one another already in infancy (de Hevia and Spelke, 2010). One recent study showed that 9-mo-olds were equally likely to transfer an arbitrary, experimentally learned magnitude-to-texture association from one dimension (e.g., number) to another dimension (size or duration) (Lourenco and Longo, 2010). In addition, 9-mo-olds can readily learn pairs of positively (but not negatively) correlated line lengths and tone durations (Srinivasan and Carey, 2010), suggesting that infants at least can represent an abstract “more-than” and “less-than” representation that applies to both dimensions. However, 9-mo-old infants do not show equal sensitivity to monotonic pairings between the dimensions of loudness and space as they do for pairing of space and time (Srinivasan and Carey, 2010). Those findings suggest that there may be an asymmetry between magnitudes in their intrinsic ordinal associations. It is important to note, however, that asymmetries in relations between magnitudes could arise either through a biologically privileged psychological mapping (de Hevia and Spelke, 2009) or through correlational and statistical learning [see Bonn and Cantlon (2012) for discussion].
Perhaps the best evidence for early-developing psychological relations among quantities is that infants at 4 mo of age spontaneously prefer to look at a ball that is bouncing congruently with the pitch of an auditory stimulus (the ball goes up when the pitch goes up) compared with a ball that is bouncing incongruently with pitch. In addition, they prefer to look at a shape that is getting sharper as the pitch of the auditory stimulus gets higher than the reverse (Walker et al., 2010). Infants are thus capable of aligning the dimensions of pitch and space (height) as well as pitch and shape (sharpness) early in development. Similarly, 3-year-olds reliably
match high-pitched sounds to smaller and brighter balls in a categorization task (Mondloch and Maurer, 2004). Those data show that magnitude dimensions beyond the canonical “privileged” dimensions of space and number can be mapped onto each other early in development.
Relations among different quantities also have been found at the neural level in adult humans and nonhuman primates. As mentioned above, individuals with spatial neglect resulting from damage to parietal cortex can exhibit impaired numerical processing. Single-neuron data from neurophysiology studies of monkeys broadly indicate that regions of parietal cortex represent space, time, and number (Tudusciuc and Nieder, 2007). Moreover, some data even suggest that a single parietal neuron can represent more than one type of magnitude. In one study (Tudusciuc and Nieder, 2007), monkeys were trained to perform a line-length matching task and a numerical matching task. During stimulus presentation as well as a subsequent delay, single neurons in the IPS responded selectively to visual stimuli according to their numerosity or length. Although some neurons responded only to numerosity and others only to line length, a subset of cells (~20%) responded to both magnitudes of line length and numerical value. These and other studies, including functional MRI studies of adults, have led some researchers to argue for a “distributed but overlapping” representation of different magnitudes at the neural level (Pinel et al., 2004; Tudusciuc and Nieder, 2007; Cantlon et al., 2009c). Simply put, different types of magnitude representation, including size, number, and time (and possibly others such as brightness), share some neural resources in parietal cortex but not others. The next section discusses some possible explanations of the origin of the relationship between number and other quantitative dimensions.
HOW IS NUMBER LINKED TO OTHER QUANTITIES?
How do different quantitative dimensions become related in the mind and brain in the first place? We have recently reviewed existing theoretical frameworks for how quantitative relations might originate (Bonn and Cantlon, 2012). Here, I briefly sketch five mechanisms for how different quantities could become related in the mind. These hypotheses are not mutually exclusive and may even be complementary.
Correlational and Statistical Associations
Learning via association and correlation is the classic developmental account of the origins of abstract percepts and concepts [e.g., Piaget (1952)]. On this view, integrated representations of information coming from separate senses, modalities, or cognitive domains arise from exposure
to correlations in the environment. Under this account, relations among magnitudes would arise from the strength of their correlations in the natural environment. For example, it takes a long time to walk a great distance (time and space are correlated), and a large number of a particular object tends to take up more surface area than a small number of that object (number and space are correlated). In this way, empirical correlations between different quantities can be absorbed through experience.
Analogical Reasoning
Another possibility is that conceptual alignment of relational information, termed “structural similarity,” mediates mapping among magnitude dimensions (Gentner and Medina, 1998). On this view, cross-dimensional mapping could be a form of analogy. Relations between magnitudes could develop through conceptual knowledge of how those dimensions are structured (Srinivasan and Carey, 2010). For example, knowledge of the conceptual fact that time and number are ordinal and monotonic dimensions (they are organized from small/short to large/long) could serve as the cognitive basis for identifying relations among those dimensions.
Amodal Representations
A third conceptual framework that could be useful for understanding relations among magnitudes derives from the literature on cross-modal sensory perception. Gibson (1969) argued that an abstract, amodal representation of intensity or amount of stimulation is present from birth or very early in infancy. On her view, amodal representations can take one of two forms: (i) intersensory redundancy (e.g., timing information about hammer strikes can be sampled from both the auditory and visual modalities), and (ii) relative intensity [e.g., “sharpness, bluntness, and jerkiness”; Gibson (1969, p. 219)]. Under a conceptualization of magnitude representation within this framework, redundancy of information would be the main source of representational overlap. For example, a bright light could be mapped to a loud tone because they both evoke an amodal representation of relatively high intensity.
Automatic Cross-Activation
A fourth hypothesis is suggested by evidence that infants experience something akin to synesthesia of sensory representations near birth [reviewed in Spector and Maurer (2009)]. A strong version of this hypothesis claims that a percept experienced in one modality automatically
stimulates a percept in another modality. Over the course of the first year of life, these associated percepts become weaker as overabundant neural connections between different functional areas of the brain become pruned or inhibited. Magnitudes, under a similar conceptualization, might be related via automatic cross-activation of dimension representations. This could imply that patterns of associations (mappings) between many magnitudes are initially strong in infancy, then get weaker during the first year(s), and then return to a strong state later in development. Generally speaking, the developmental data from cross-modal perception indicate that patterns of associations among magnitudes might not strengthen straightforwardly over development.
Evolutionary History
A final possibility is that relations among magnitudes derive from their evolutionary history rather than solely from developmental processes that unfold within an individual lifespan. On this view, one quantitative dimension evolved from another, inheriting functional similarities and potentially mutual dependencies in neural and computational operations. For example, many magnitude representations could have emerged from descent with modification of the functional substrates that code for space, resulting in a common psychological and neural code for dimensions such as space, number, time, loudness, brightness, and pitch (Bonn and Cantlon, 2012).
Clearly there is a dense set of possibilities for how different quantities could come to be related in the mind and brain. The five hypotheses sketched above address different levels of influence ranging from ontogeny to phylogeny. They also address different levels of psychological functioning ranging from basic representations of psychophysical values to abstract perceptual and conceptual relations. Different levels of analysis will be important for understanding the full taxonomy of numerical cognition in humans. However, although questions remain as to how primitive numerical representations are organized with respect to other types of quantities (e.g., size, time, loudness), it is clear that human children use those primitive numerical representations to learn the process of verbal counting early in development. Verbal counting (discussed earlier) is the first formal cognitive step toward acquiring the uniquely human capacity for complex symbolic math. In the next section we discuss how the “primitive” analog numerical abilities are related to symbolic math in humans.
ORIGINS OF MATH IQ
A further issue central to understanding the taxonomy of primitive numerical cognition is the extent to which analog numerical abilities bear
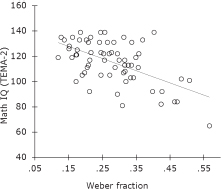
FIGURE 16.6 Childhood math IQ (measured by the TEMA-2) is correlated with the precision of analog numerical discrimination (measured by subjects’ Weber fractions). A higher Weber fraction reflects worse discrimination. Redrawn from Halberda et al. (2008).
a neural relationship with full-blown formal mathematics IQ. Researchers have begun to examine, in humans, how formal math intelligence may be modulated by developments in the “primitive” analog numerical system that is shared by nonhuman primates, adult humans, and children. These studies have largely hinged on analyses of individual differences in numerical and mathematical abilities.
Individual differences in math IQ are predicted by differences in analog numerical sensitivity (Bull and Scerif, 2001; Halberda et al., 2008; Holloway and Ansari, 2009). Studies with children indicate that analog numerical ability correlates with performance on math IQ tests and that formal math ability is more closely correlated with analog numerical abilities than it is with other formal abilities, such as reading. For example, in Fig. 16.6, adolescents’ analog numerical ability (measured by the Numerical Weber Fraction) correlates with their math IQ from early childhood [measured by the Test of Early Mathematics Ability (TEMA)-2 test score]. This and similar findings indicate that the “primitive” ability to estimate numerical values from sets of objects is related to the development of full-blown math skills. Other studies highlight the role of executive function and working memory in the development of formal mathematical reasoning (Bull and Scerif, 2001; Mazzocco et al., 2006; Mazzocco and Kover, 2007). Together, these studies indicate a need to understand the relative contributions of domain-specific and domain-general processes to formal mathematical skill.
Behavioral data, like those described earlier, provide evidence of a relationship between the skills required for analog numerical processing and those that are used in formal mathematics by children. Neuroimaging studies of children can provide an independent source of data on whether there is a common foundation for analog numerical abilities and formal math by testing whether a common neural substrate underlies both faculties. As described above, analog quantity judgments recruit regions of
the IPS in adult humans, human children, and nonhuman primates. One issue is whether the same neural patterns that are evoked during analog numerical processing are observed when children and adults process the symbolic numbers that are unique to human culture (e.g., numerals, number words). Several studies suggest that they do: regions of the IPS exhibit activity that is greater for numerical symbols compared with control stimuli, and those IPS regions also exhibit the numerical distance and ratio effects in their neural responses (Cohen Kadosh et al., 2007; Piazza et al., 2007; Ansari, 2008; Cantlon et al., 2009b; Holloway and Ansari, 2010). Research further suggests that the same neural response patterns are elicited for both symbolic and nonsymbolic (analog) numbers in the same subjects (Piazza et al., 2007). Together, these results implicate neural overlap in the substrates underlying symbolic and nonsymbolic (analog) numerical representations in humans.
In humans, a second brain region is often recruited during symbolic numerical tasks: the prefrontal cortex, particularly the inferior frontal gyrus, bordering insular cortex (Ansari et al., 2005; Piazza et al., 2007; Cantlon et al., 2009b; Emerson and Cantlon, 2012). Structurally, the prefrontal cortex is thought to be unique in primates compared with other mammals (Preuss, 2007). In humans the prefrontal cortex responds during many types of abstract judgments (Miller et al., 2002), and several studies have noted a unique involvement of the prefrontal cortex in the development of semantic representations, symbols, and rules [see Nieder (2009) for review]. A pattern of greater activation of prefrontal sites in children compared with adults has also been observed for numerical and basic mathematical tasks (Ansari et al., 2005; Rivera et al., 2005; Cantlon et al., 2009b). The role of prefrontal cortex in children’s symbolic numerical processing is related to performance factors such as response time, or “time on task” [Emerson and Cantlon (2012); see also Schlaggar et al. (2002)], which could reflect the nascent state of children’s abstract, symbolic numerical representations. Studies with nonhuman primates have suggested that they too engage prefrontal cortex during numerical processing [see Nieder (2009) for review] and that prefrontal regions play a unique role in associating analog numerical values with arbitrary symbols at the level of single neurons in monkeys (Diester and Nieder, 2007).
Findings that highlight mutual involvement of the IPS and prefrontal cortex in basic numerical tasks have led to the hypothesis that interactions between frontal and parietal regions are important for the development of uniquely human numerical cognition, such as symbolic coding. Specifically, it has been proposed that the IPS computes “primitive” analog numerical representations and the prefrontal cortex facilitates links between those analog numerical computations and symbolic number representations in humans (Cantlon et al., 2009b; Nieder, 2009). If this
hypothesis is correct then network-level neural synchrony between frontal and parietal regions should predict formal mathematics development in humans. That is, individual variability in the strength of correlations between neural responses in frontal and parietal regions, or “functional connectivity,” should be related to individual variability in mathematics performance. We have recently tested this hypothesis and found that number-specific functional connectivity of the fronto-parietal network does predict children’s math IQ test scores (independently of their verbal IQ test scores) (Emerson and Cantlon, 2012). The implication is that number-specific changes in the interactions between frontal and parietal regions are related to the development of symbolic, formal math concepts in children. This general conclusion is in line with the hypothesis that interactions between the “primitive” numerical operations of the IPS and the abstract, symbolic operations of frontal cortex give rise to formal mathematics concepts in humans.
CONCLUSION
The goal of this review has been to examine the origins and organization of numerical abilities ranging from analog quantification to formal arithmetic. The general hypothesis is that the uniquely human ability to perform complex and sophisticated mathematics can be traced back to a simpler computational system that is shared among many animals: the analog numerical system. Humans and nonhuman animals possess a common system for making numerical judgments via analog representations. Throughout development, analog numerical representations interact with the uniquely human ability to represent numerical values symbolically, suggesting a relationship between “primitive” and modern numerical systems in humans. Data from neural analyses of numerical processing support this conclusion and provide independent confirmation that these are in fact related systems. Questions remain regarding the precise taxonomy of the development and organization of numerical information, and its relationship to other domains, such as “space.” However, the general nature of the relationship between “primitive” and modern numbers seems to derive from evolutionary constraints on the structure of numerical concepts in the mind and brain as well as the conceptual and neural foundation that evolution has provided for the development of numerical thinking in humans.
ACKNOWLEDGMENTS
I thank Brad Mahon and Vy Vo for comments. Support was received from National Institute of Child Health and Human Development Grant R01HD064636 and the James S. McDonnell Foundation.
A Hierarchical Model of the Evolution of Human Brain Specializations
![]()
The study of information-processing adaptations in the brain is controversial, in part because of disputes about the form such adaptations might take. Many psychologists assume that adaptations come in two kinds, specialized and general-purpose. Specialized mechanisms are typically thought of as innate, domain-specific, and isolated from other brain systems, whereas generalized mechanisms are developmentally plastic, domain-general, and interactive. However, if brain mechanisms evolve through processes of descent with modification, they are likely to be heterogeneous, rather than coming in just two kinds. They are likely to be hierarchically organized, with some design features widely shared across brain systems and others specific to particular processes. Also, they are likely to be largely developmentally plastic and interactive with other brain systems, rather than canalized and isolated. This chapter presents a hierarchical model of brain specialization, reviewing evidence for the model from evolutionary developmental biology, genetics, brain mapping, and comparative studies. Implications for the search for uniquely human traits are discussed, along with ways in which conventional views of modularity in psychology may need to be revised.
__________
Center for Behavior, Evolution, and Culture and FPR-UCLA Center for Culture, Brain and Development, Department of Anthropology, University of California, Los Angeles, CA 90095. E-mail: barrett@anthro.ucla.edu.




















