
2
Major Structures and Functions of the Brain
Outside the specialized world of neuroanatomy and for most of the uses of daily life, the brain is more or less an abstract entity. We do not experience our brain as an assembly of physical structures (nor would we wish to, perhaps); if we envision it at all, we are likely to see it as a large, rounded walnut, grayish in color.
This schematic image refers mainly to the cerebral cortex, the outermost layer that overlies most of the other brain structures like a fantastically wrinkled tissue wrapped around an orange. The preponderance of the cerebral cortex (which, with its supporting structures, makes up approximately 80 percent of the brain's total volume) is actually a recent development in the course of evolution. The cortex contains the physical structures responsible for most of what we call “brainwork ”: cognition, mental imagery, the highly sophisticated processing of visual information, and the ability to produce and understand language. But underneath this layer reside many other specialized structures that are essential for movement, consciousness, sexuality, the action of our five senses, and more—all equally valuable to human existence. Indeed, in strictly biological terms, these structures can claim priority over the cere-
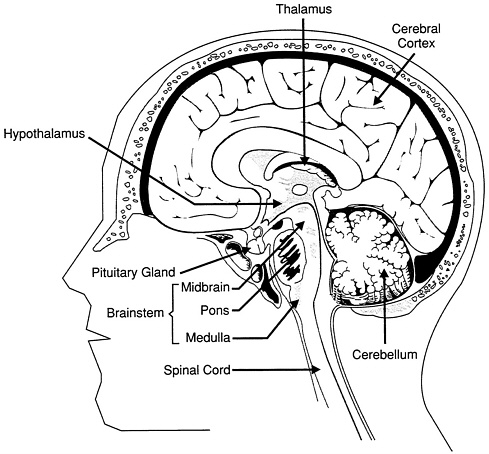
FIGURE 2.1. The brain owes its outer appearance of a walnut to the wrinkled and deeply folded cerebral cortex, which handles the innumerable signals responsible for perception and movement and also for mental processes. Below the surface of the cortex are packed a number of other specialized structures: the thalamus, an important relay station for the senses, and the hypothalamus, a meeting point between the nervous system and the endocrine system and between emotion and physical feeling. The pituitary gland, acting on signals from the hypothalamus, produces hormones that regulate many functions from growth to reproduction. The pons and the medulla, two major elements of the brainstem, channel nerve signals between the brain and other parts of the body, controlling vital functions such as breathing and deliberate movement. (The extension of this signal pathway throughout the trunk and abdomen is, of course, the spinal cord.) At the back of the brain is the cerebellum, which coordinates the brain's instructions for skilled repetitive movements and for maintaining posture and balance. Source: Adapted from G. J. Torbra, Principles of Human Anatomy, 3rd ed. Harper and Row (1983).
bral cortex. In the growth of the individual embryo, as well as in evolutionary history, the brain develops roughly from the base of the skull up and outward. The human brain actually has its beginnings, in the four-week-old embryo, as a simple series of bulges at one end of the neural tube.
VENTRICLES
The bulges in the neural tube of the embryo develop into the hindbrain, midbrain, and forebrain—divisions common to all vertebrates, from sharks to squirrels to humans. The original hollow structure is commemorated in the form of the ventricles, which are cavities containing cerebrospinal fluid. During the course of development, the three bulges become four ventricles. In the hindbrain is the fourth ventricle, continuous with the central canal of the spinal cord. A cavity in the forebrain becomes the third ventricle, which leads further forward into the two lateral ventricles, one in each cerebral hemisphere.
BRAINSTEM
The hindbrain contains several structures that regulate autonomic functions, which are essential to survival and not under our conscious control. The brainstem, at the top of the spinal cord, controls breathing, the beating of the heart, and the diameter of blood vessels. This region is also an important junction for the control of deliberate movement. Through the medulla, at the lower end of the brainstem, pass all the nerves running between the spinal cord and the brain; in the pyramids of the medulla, many of these nerve tracts for motor signals cross over from one side of the body to the other. Thus, the left brain controls movement of the right side of the body, and the right brain controls movement of the left side.
In addition to being the major site of crossover for nerve tracts running to and from the brain, the medulla is the seat of several pairs of nerves for organs of the chest and abdomen, for movements of the shoulder and head, for swallowing, salivation, and taste, and for hearing and equilibrium.
At the top of the brainstem is the pons—literally, a bridge— between the lower brainstem and the midbrain. Nerve impuls-
es traversing the pons pass on to the cerebellum (or “little brain”), which is concerned primarily with the coordination of complex muscular movement. In addition, nerve fibers running through the pons relay sensations of touch from the spinal cord to the upper brain centers.
Many nerves for the face and head have their origin in the pons, and these nerves regulate some movements of the eye-ball, facial expression, salivation, and taste. Together with nerves of the medulla, nerves from the pons also control breathing and the body's sense of equilibrium.
What had been the middle bulge in the neural tube develops into the midbrain, which functions mainly as a relay center for sensory and motor nerve impulses between the pons and spinal cord and the thalamus and cerebral cortex. Nerves in the midbrain also control some movements of the eyeball, pupil, and lens and reflexes of the eyes, head, and trunk.
THALAMUS AND HYPOTHALAMUS
Deep in the core area of the brain, just above the top of the brainstem, are structures that have a great deal to do with perception, movement, and the body's vital functions. The thalamus consists of two oval masses, each embedded in a cerebral hemisphere, that are joined by a bridge. The masses contain nerve cell bodies that sort information from four of the senses—sight, hearing, taste, and touch—and relay it to the cerebral cortex. (Only the sense of smell sends signals directly to the cortex, bypassing the thalamus.) Sensations of pain, temperature, and pressure are also relayed through the thalamus, as are the nerve impulses from the cerebral hemispheres that initiate voluntary movement.
The hypothalamus, despite its relatively small size (roughly that of a thumbnail), controls a number of drives essential for the functioning of a wide-ranging omnivorous social mammal. At the autonomic level, the hypothalamus stimulates smooth muscle (which lines the blood vessels, stomach, and intestines) and receives sensory impulses from these areas. Thus it controls the rate of the heart, the passage of food through the alimentary canal, and contraction of the bladder.
The hypothalamus is the main point of interaction for the
body's two physical control systems: the nervous system, which transmits information in the form of minute electrical impulses, and the endocrine system, which brings about changes of state through the release of chemical factors. It is the hypothalamus that first detects crucial changes in the body and responds by stimulating various glands and organs to release hormones.
The hypothalamus is also the brain's intermediary for translating emotion into physical response. When strong feelings (rage, fear, pleasure, excitement) are generated in the mind, whether by external stimuli or by the action of thoughts, the cerebral cortex transmits impulses to the hypothalamus; the hypothalamus may then send signals for physiological changes through the autonomic nervous system and through the release of hormones from the pituitary. Physical signs of fear or excitement, such as a racing heartbeat, shallow breathing, and perhaps a clenched “gut feeling,” all originate here.
Also in the hypothalamus are neurons that monitor body temperature at the surface through nerve endings in the skin, and other neurons that monitor the blood flowing through this part of the brain itself, as an indicator of core body temperature. The front part of the hypothalamus contains neurons that act to lower body temperature by relaxing smooth muscle in the blood vessels, which causes them to dilate and increases the rate of heat loss from the skin. Through its neurons associated with the sweat glands of the skin, the hypothalamus can also promote heat loss by increasing the rate of perspiration. In opposite conditions, when the body's temperature falls below the (rather narrow) ideal range, a portion of the hypothalamus directs the contraction of blood vessels, slows the rate of heat loss, and causes the onset of shivering (which produces a small amount of heat).
The hypothalamus is the control center for the stimuli that underlie eating and drinking. The sensations that we interpret as hunger arise partly from a degree of emptiness in the stomach and partly from a drop in the level of two substances: glucose circulating in the blood and a hormone that the intestine produces shortly after the intake of food. (Receptors for this hormone gauge how far digestion has proceeded since the last meal.) This system is not a simple “on” switch for hunger,
however: another portion of the hypothalamus, when stimulated, actively inhibits eating by promoting a feeling of satiety. In experimental animals, damage to this portion of the brain is associated with continued excessive eating, eventually leading to obesity.
In addition to these numerous functions, there is evidence that the hypothalamus plays a role in the induction of sleep. For one thing, it forms part of the reticular activating system, the physical basis for that hard-to-define state known as consciousness (about which more later); for another, electrical stimulation of a portion of the hypothalamus has been shown to induce sleep in experimental animals, although the mechanism by which this works is not yet known. In all, the hypothalamus is a richly complex cubic centimeter of vital connections, which will continue to reward close study for some time to come. Because of its unique position as a midpost between thought and feeling and between conscious act and autonomic function, a thorough understanding of its workings should tell us much about the earliest history and development of the human animal.
PITUITARY AND PINEAL GLANDS
The pituitary and the pineal glands function in close association with the hypothalamus. The pituitary responds to signals from the hypothalamus by producing an array of hormones, many of which regulate the activities of other glands: thyroid-stimulating hormone, adrenocorticotropic hormone (which stimulates an outpouring of epinephrine in response to stress), prolactin (involved in the production of milk), and the sex hormones follicle-stimulating hormone and luteinizing hormone, which promote the development of eggs and sperm and regulate the timing of ovulation. The pituitary gland also produces several hormones with more general effects: human growth hormone, melanocyte-stimulating hormone (which plays a role in the pigmentation of skin), and dopamine, which inhibits the release of prolactin but is better known as a neurotransmitter (see Chapter 5 ).
The pineal gland produces melatonin, the hormone associated with skin pigmentation. The secretion of melatonin var-
ies significantly over a 24-hour cycle, from low levels during the day to a peak at night, and the pineal gland has been called a “ third eye” because it is controlled by neurons sensitive to light, which originate in the retina of each eye and end in the hypothalamus. In animals with a clear-cut breeding season, the pineal gland is a link between the shifting hours of daylight and the hormonal responses of the hypothalamus, which in turn guide reproductive functions. In humans, who can conceive and give birth throughout the year, the pineal gland plays no known role in reproduction, although there is evidence that melatonin has a share in regulating ovulation.
THE “LITTLE BRAIN” AT THE BACK OF THE HEAD
While autonomic and endocrine functions are being maintained by structures deep inside the brain, another specialized area is sorting and processing the signals required to maintain balance and posture and to carry out coordinated movement. The cerebellum (the term in Latin means “little brain”) is actually a derived form of the hindbrain—as suggested by its position at the back of the head, partly tucked under the cerebral hemispheres. In humans, with our almost unlimited repertoire of movement, the cerebellum is accordingly large; in fact, it is the second-largest portion of the brain, exceeded only by the cerebral cortex. Its great surface area is accommodated within the skull by elaborate folding, which gives it an irregular, pleated look. In relative terms, the cerebellum is actually largest in the brain of birds, where it is responsible for the constant streams of information between brain and body that are required for flight.
In humans, the cerebellum relays impulses for movement from the motor area of the cerebral cortex to the spinal cord; from there, they pass to their designated muscle groups. At the same time, the cerebellum receives impulses from the muscles and joints that are being activated and in some sense compares them with the instructions issued from the motor cortex, so that adjustments can be made (this time by way of the thalamus). The cerebellum thus is neither the sole initiator of movement nor a simple link in the chain of nerve impulses, but a site for the rerouting and in some cases refining of instructions for movement. There is evidence, too, that the cerebellum can
store a sequence of instructions for frequently performed movements and for skilled repetitive movements—those that we think of as learned “by rote.”
The right and left hemispheres of the cerebellum each connect with the nerve tracts from the spinal cord on the same side of the body, and with the opposite cerebral hemisphere. For example, nerve impulses concerned with movement of the left arm originate in the right cerebral hemisphere, and information about the orientation, speed, and force of the movement is fed back to the right cerebral hemisphere, through the left half of the cerebellum. The nerves responsible for movement at the ends of the arms and legs tend to have their origin near the outer edges of the cerebellum. By contrast, nerves that have their origin near the center of the cerebellum serve to monitor the body's overall orientation in space and to maintain upright posture, in response to information about balance that is transmitted by nerve impulses from the inner ear, among other sources.
RETICULAR NETWORK
Some nerve fibers from the cerebellum also contribute to the reticular formation, a widespread network of neurons (“reticular” is derived from the Latin word for “net”). This formation and some neurons in the thalamus, together with others from various sensory systems of the brain, make up the reticular activating system—the means by which we maintain consciousness. The reticular activating system also comes into play when we deliberately focus our attention, “tuning out” distractions to some degree. At the midline of the brainstem are the raphe nuclei, whose axons extend down into the spinal cord and up to the cerebral cortex —a reach that makes it possible for many areas of the nervous system to be contacted simultaneously. The reticular formation plays a role in movement, particularly those forms of movement that do not call for conscious attention: it is also involved in transmitting or inhibiting sensations of pain, temperature, and touch. Less tangibly, the reticular activating system appears to work as a filter for the countless stimuli that can act on the nervous system both from within and from outside the body. It is this filtering of signals that allows a passenger on an airplane, for
example, to doze off undisturbed by sounds of nearby conversation and steady jet engines, but to awake and become alert when the pitch of the engines changes and the plane tilts into its descent.
THE “EMOTIONAL BRAIN”
The limbic system (from the Latin limbus, for “hem” or “border”) is another assembly of linked structures that form a loose circuit throughout the brain. This system is a fairly old part of the brain and one that humans share with many other vertebrates; in reptiles, it is known as the rhinencephalon, or “smellbrain,” because it reacts primarily to signals of odor. In humans, of course, the stimuli that can affect the emotional brain are just about limitless in their variety.
The limbic system is responsible for most of the basic drives and emotions and the associated involuntary behavior that are important for an animal's survival: pain and pleasure, fear, anger, sexual feelings, and even docility and affection. As with the rhinencephalon, the sense of smell is a powerful factor. Nerves from the olfactory bulb, by which all odor is perceived, track directly into the limbic system at several points and are then connected through it to other parts of the brain; hence the ability of pheromones, and perhaps of other odors as well, to influence behavior in quite complex ways without necessarily reaching our conscious awareness.
Also feeding into the limbic system are the thalamus and hypothalamus, as well as the amygdala, a small, almond-shaped complex of nerve cells that receive input from both the olfactory system and the cerebral cortex. These connections are illustrated in an unusual way in the context of epilepsy. Perhaps because the amygdala is located near a common site of origin of epileptic seizures—that is, in the temporal lobe of the cerebral hemispheres—epileptics sometimes experience unidentifiable or unpleasant odors or changes of mood as part of the aura preceding a seizure. The limbic system is not thought to be involved in the causes of epilepsy, but it is indirectly stimulated by the electric discharge in the brain that sets off a seizure and gives evidence of the stimulation in its own characteristic ways.
HIPPOCAMPUS
The hippocampus is another major structure of the limbic system. Named for its fanciful resemblance to the shape of a sea horse, the hippocampus is located at the base of the temporal lobe near several sets of association fibers. These are bundles of nerve fibers that connect one region of the cerebral cortex with another, so that the hippocampus, as well as other parts of the limbic system, exchanges signals with the entire cerebral cortex. The hippocampus has been shown to be important for the consolidation of recently acquired information. (In contrast, long-term memory is thought to be stored throughout the cerebral cortex. The means by which short-term memory is converted into long-term memory has posed a particularly challenging riddle that only now is beginning to yield to investigation; see Chapter 8 .)
Recent work with a variety of animals has found dense clusters of receptor sites for tetrahydrocannabinol, the active ingredient of marijuana and related drugs, in the hippocampus and other nearby structures of the limbic system. This localization helps explain the effects of marijuana, which range from mild euphoria to wavering attention to temporarily weakened short-term memory. A loss of short-term memory is also seen in certain syndromes of alcoholism and in Alzheimer 's disease, which involves some degeneration of the hippocampus and other limbic structures.
CEREBRAL CORTEX
The cerebral cortex occupies by far the greatest surface area of the human brain and presents its most striking aspect. Also known as the neocortex, this is the most recently evolved area of the brain. In fact, the enormous expansion in the area of the cerebral cortex is hypothesized to have begun only about 2 million years ago, in the earliest members of the genus Homo; the result today is a brain weighing approximately three times more than would be expected for a mammal our size. The cortex is named for its resemblance to the bark of a tree, because it covers the surface of the cerebral hemispheres in a similar way. Its wrinkled convoluted appearance is due to a
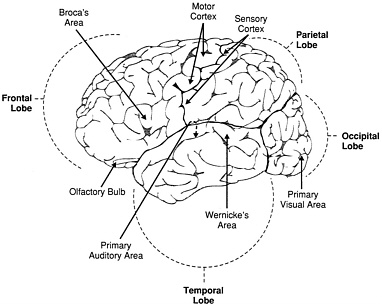
FIGURE 2.2. The brain is divided into a left and a right hemisphere by a deep groove that runs from the front of the head (at left) to the back (at right). In each hemisphere, the cerebral cortex falls into four main divisions, or lobes, set off from one another by noticeable folds in the surface. Although there is some overlap of tasks among the lobes, each is best known for one or two specialized functions. The frontal lobes house the motor area (responsible for instructions of movement) and Broca's area, which handles the production of speech. The faculties of planning and mental representation of the outside world are also attributed to the frontal lobes. In the parietal lobes, the cerebral cortex processes the signals that come from sensation; the temporal lobes are concerned with memory, hearing, and, in Wernicke 's area, with the ability to understand language. The occipital lobes are specialized to manage the intricate processing of vision. The olfactory bulb, one of the older parts of the brain in vertebrates, is tucked just under the frontal lobes.
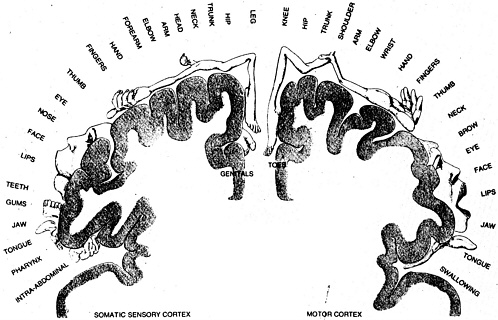
FIGURE 2.3. Two miniature “maps” represent the body on the cerebral cortex. One of these, in the motor area, assigns a specific portion of the cortex to each part of the body that calls for muscular control; the portions assigned to the fingers, lips, and tongue are surprisingly large, reflecting the demands of highly precise instructions needed for speech and for manual skills. The other map, in the sensory area, devotes a specific area to each part of the body that receives sensations. Here, again, the portion for the face and hands is relatively large —but so, too, is the portion that processes signals from the abdomen and intestines. Source: W. H. Freeman, 1979. The Brain: A Scientific American Book.
growth spurt during the fourth and fifth months of embryonic development, when the gray matter of the cortex is expanding greatly as its cells grow in size. The supporting white matter, meanwhile, grows less rapidly; as a result, the brain takes on the dense folds and fissures characteristic of an object with great surface area crowded into a small space.
Although the folds in the cerebral cortex appear at first to be random, they include several prominent bulges, or gyri, and grooves, or sulci, that act as landmarks in what is in fact a highly ordered structure (the finer details of which are still not completely known). The deepest groove extends from the front to the back of the head, dividing the brain into the left and right hemispheres. The central sulcus, which runs from the middle of the brain outward to both left and right, and the lateral sulcus, another left-to-right groove somewhat lower on the hemispheres and toward the back of the head, further divide each hemisphere into four lobes: frontal, parietal, temporal, and occipital. A fifth lobe, known as the insula, is located deep within the parietal and temporal lobes and is not apparent as a separate structure on the outer surface of the cerebral hemispheres.
Two noticeable bulges, the precentral gyrus and the post-central gyrus, are named for their positions just in front of and just behind the central sulcus, respectively. The precentral gyrus is the site of the primary motor area, responsible for conscious movement. From eyebrows to toes, the movable parts of the body are “mapped” on this area of the cortex, with each muscle group or limb represented here by a population of neurons. In complementary fashion, the job of receiving sensations from all parts of the body is managed by the primary somatosensory area, which is located in the postcentral gyrus. Here, too, the human form is mapped, and, as with the precen-
tral gyrus, the areas devoted to the hand and the mouth are disproportionately large. Their size reflects the elaborate brain circuitry that makes possible the precision grip of the human hand, the fine motor and sensory signals required for striking up a violin arpeggio or sharpening a tool, and the coordination of the lips, tongue, and vocal apparatus to produce the highly arbitrary and significant sounds of human language.
Close observations of animals and humans after injury to particular sites of the brain indicate that many areas of the cortex control quite specific functions. Additional findings have come from stimulating sites on the cortex with a small electrical charge in experimental procedures or during surgery; the result might be an action in some part of the body (if the motor cortex is involved) or (for a sensory function) a pattern of electrical discharges in other parts of the cortex. Careful exploration has established, for example, that the auditory area in the temporal lobe is made up of smaller regions, each attuned to different sound frequencies.
But for much of the cortex, no such direct functions have been found, and for a time these areas were known as “silent” cortex. It is now clear that “association” cortex is a better name for them because they fill the crucial role of making sense of received stimuli, piecing together the signals from various sensory pathways and making the synthesis available as felt experience. For instance, if there is to be not merely perception but conscious understanding of sounds, the auditory association area (just behind the auditory area proper) must be active. In the hemisphere that houses speech and other verbal abilities—the left hemisphere, for most people—the auditory association area blends into the receptive language area (which also receives signals from the visual association area, thereby providing a neural basis for reading as well as for the comprehension of speech in most languages).
A large portion of the association cortex is found in the frontal lobes, which have expanded most rapidly over the past 20,000 or so generations (about 500,000 years) of human evolution. Medical imaging shows increased activity in the association cortex after other areas of the brain have received electrical stimulation and also before the initiation of movement. On present evidence, it is in the association cortex that we locate
long-term planning, interpretation, and the organization of ideas — perhaps the most recently developed elements of the modern human brain.
Visual functions occupy the occipital lobe, the bulge at the back end of the brain. The primary area for visual perception is almost surrounded by the much larger visual association area. Nearby, extending into the lower part of the temporal lobe, is the association area for visual memory—a specialized area in the cortex. Clearly, this function has been important for an omnivorous foraging primate that probably spent a long evolutionary period ranging among scattered food sources. (For an account of the intricate mechanisms that underlie depth perception and color vision, see Chapter 7 .)
A less specific kind of function has been attributed to the prefrontal cortex, located on the forward-facing part of the frontal lobes. This area is connected by association fibers with all other regions of the cortex and also with the amygdala and the thalamus, which means that it, too, makes up part of the “emotional brain,” the limbic system. Injury to the prefrontal cortex or its underlying white matter results in a curious disability: the patient suffers from a reduced intensity of emotion and can no longer foretell the consequences of things that are said or done. (The injury must be bilateral to produce such an effect; if only one hemisphere is injured, the other can compensate and avert this strange, potentially crippling social deficit.) Among its other functions, the prefrontal cortex is responsible for inhibiting inappropriate behavior, for keeping the mind focused on goals, and for providing continuity in the thought process.
Long-term memory has not yet been found to reside in any exclusive part of the brain, but experimental findings indicate that the temporal lobes contribute to this function. Electrical stimulation of the cerebral cortex in this area gives rise to sensations of déjà vu (“already seen”) and its opposite, jamais vu (“never seen”); it also conjures up images of scenes witnessed or speech heard in the past. That the association areas for vision and hearing and the language areas are all nearby may suggest pathways for the storage and retrieval of memories that include several types of stimuli.
The function of language itself is housed in the left hemisphere (in most cases), in several discrete sites on the cortex.
The expressive language area, responsible for the production of speech, is found toward the center of the frontal lobe; this is also called Broca's area, after the French anatomist and anthropologist of the mid-1800s who was among the first to observe differences in function between the left and right hemispheres. The receptive language area, which is located near the junction of the parietal and temporal lobes, allows us to comprehend both spoken and written language, as described above. This is often called Wernicke's area, after the German neurologist Karl Wernicke, who in the late 1800s laid the basis for much of our current understanding of how the brain encodes and decodes language. A bundle of nerve fibers connects Wernicke's area directly to Broca 's area. This tight linkage is important, since before any speech at all can be uttered, its form and appropriate words must first be assembled in Wernicke's area and then relayed to Broca's area to be mentally translated into the requisite sounds; only then can it pass to the supplementary motor cortex for vocal production.
For nine of ten right-handed people and almost two-thirds of all left-handers, language abilities are sited in the left hemisphere. No one knows why there should be this asymmetrical distribution rather than an even balance or, for that matter, a consistent location of language in the left brain. What is clear is that in all cases, the hemisphere that does not contain language abilities holds the key to other functions of a less distinct, more holistic nature. The appreciation of forms and textures, the recognition of the timbre of a voice, and the ability to orient oneself in space all appear to lodge here, as do musical talent and appreciation—a host of perceptions that do not lend themselves well to analysis in words.
The limited specialization of the two hemispheres is efficient in terms of the use of space: it increases the functional abilities of the brain without adding to its volume. (The skull of the human infant, it is calculated, is already as large as can be accommodated through the birth canal, which in turn is constrained by the skeletal requirements for upright walking.) Moreover, the bilateral arrangement allows for some flexibility if one hemisphere is injured; often the other hemisphere can compensate to some degree, depending on the age at which injury occurs (a young, still-developing brain readjusts more readily).
The two hemispheres are connected mainly by a thick bundle of nerve fibers called the corpus callosum, or “hard body,” because of its tough consistency. A smaller bundle, the anterior commissure, connects just the two temporal lobes. Although the corpus callosum is a good landmark for students of brain anatomy, its contribution to behavior has been difficult to pin down. Patients in whom the corpus callosum has been severed (a way of ameliorating epilepsy by restricting seizures to one side of the brain) go about their everyday business without impairment. Careful testing does turn up a gap between sensations processed by the right brain and the language centers of the left brain—for instance, a person with a severed corpus callosum is unable to name an object placed unseen in the left hand (because stimuli perceived by the left half of the body are processed in the right hemisphere). On the whole, though, it appears that the massive crossing-over of nerve fibers that takes place in the brainstem is quite adequate for most purposes, at least those related to survival.
Although the cerebral cortex is quite thin, ranging from 1.5 to 4 millimeters deep (less than 3/8 inch), it contains no fewer than six layers. From the outer surface inward, these are the molecular layer, made up for the most part of junctures between neurons for the exchange of signals; the external granular layer, mainly interneurons, which serve as communicating nerve bodies within a region; an external pyramidal layer, with large-bodied “principal” cells whose axons extend into other regions; an internal granular layer, the main termination point for fibers from the thalamus; a second, internal pyramidal layer, whose cells project their axons mostly to structures below the cortex; and a multiform layer, again containing principal cells, which in this case project to the thalamus. The layers vary in thickness at different sites on the cortex; for example, the granular layers (layers 2 and 4) are more prominent in the primary sensory area and less so in the primary motor area.
BUILDING BLOCKS OF THE BRAIN
Extensive and intricate as the human brain is, and with the almost limitless variation of which it is capable, it is built from relatively few basic units. The fundamental building block of
the human brain, like that of nervous systems throughout the animal kingdom, is the neuron, or nerve cell. The neuron conducts signals by means of an axon, which extends outward from the soma, or body of the cell, like a single long arm. Numerous shorter arms, the dendrites (“little branches”), conduct signals back to the soma.
The ability of the axon to conduct nerve impulses is greatly enhanced by the myelin sheath that surrounds it, interrupted at intervals by nodes. Myelin is a fatty substance, a natural electrical insulator, that protects the axon from interference by other nearby nerve impulses. The arrangement of nodes increases the speed of conductivity, so that an electrical impulse sent along the axon can literally jump from node to node, reaching velocities as high as 120 meters per second.
The site of communication between any two neurons—actually not a physical contact but an infinitesimal cleft across which signals are transmitted—is called a synapse, from the Greek word for “conjunction.” An axon may extend over a variable distance to make contact with other neurons at a synapse. The end of an axon near a synapse widens out into a bouton, or button; the bouton contains mitochondria, which supply energy, and a number of synaptic vesicles. It is these vesicles, each less than 200 billionths of a meter in diameter, that contain the chemical neurotransmitters to be released into the synaptic cleft. On the other side of the synapse is usually a dendrite, sometimes with a dendritic spine—a small protuberance that expands the surface area of the dendrite and provides a receptive site for incoming signals.
A completely different arangement for transmitting signals is the electrical synapse, at which the cell membranes of two neurons are extremely close together and are linked by a bridge of tubular protein molecules. This bridge allows passage of water and electrically charged small molecules; any change in electrical charge in one neuron is instantaneously transmitted to the other. Hence this mechanism for relaying signals relies entirely on direct electrical coupling; an electrical synapse is about 3 nanometers (nm), or billionths of a meter, wide, as compared with the 25-nm gap of a chemical synapse. Outside of nervous tissue, electrical synapses (and other, similar gap junctions) are the messengers of choice.
The brain is sometimes said to be full of “gray matter,” which is supposed to be the stuff of intelligence. The material referred to is actually grayish pink in living brain, and only gray in specimens that have been chemically preserved; it consists of nerve cell bodies and dendrites and the origins and boutons of axons. It is gray matter that forms sheets of cortex on the surface of the cerebral hemispheres. White matter receives its name from the appearance of the myelin enclosing the elongated region of axons. The third main form of matter in the brain is the neuroglia, or “glue” cells. These cells do not connect the neurons, as their name implies; connections are already far from scarce, with the vast system of neural soma, axons, and dendrites packed so densely into the brain. Rather, the neuroglia provide structural support and a source of metabolic energy for the roughly 100 billion nerve cells of the human brain.
CHEMICAL AND ELECTRICAL SIGNALS
The actual signals transmitted throughout the brain come in two forms, electrical and chemical. The two forms are interdependent and meet at the synapse, where chemical substances can alter the electrical conditions within and outside the cell membrane.
A nerve cell at rest holds a slight negative charge (about –70 millivolts, or thousandths of a volt, mV) with respect to the exterior; the cell membrane is said to be polarized. The negative charge, the resting potential of the membrane, arises from a very slight excess of negatively charged molecules inside the cell.
A membrane at rest is more or less impermeable to positively charged sodium ions (Na+), but when stimulated it is transiently open to their passage. The Na+ ions thus flow in, attracted by the negative charge inside, and the membrane temporarily reverses its polarity, with a higher positive charge inside than out. This stage lasts less than a millisecond, and then the sodium channels close again. Potassium channels (K+) open, and K+ ions move out through the membrane, reversing the flow of positively charged ions. (Both these channels are known as voltage-gated, meaning that they open or close in response
to changes in electrical charge occurring across the membrane.) Over the next 3 milliseconds, the membrane becomes slightly hyperpolarized, with a charge of about –80 mV, and then returns to its resting potential. During this time the sodium channels remain closed; the membrane is in a refractory phase.
An action potential—the very brief pulse of positive membrane voltage—is transmitted forward along the axon; it is prevented from propagating backward as long as the sodium channels remain closed. After the membrane has returned to its resting potential, however, a new impulse may arrive to evoke an action potential, and the cycle can begin again.
Gated channels, and the concomitant movement of ions in and out of the cell membrane, are widespread throughout the nervous system, with sodium, potassium, and chlorine being the most common ions involved. Calcium channels are also important, particularly at the presynaptic boutons of axons. When the membrane is at its resting potential, positively charged calcium ions (Ca2+) outside the cell far outnumber those inside. With the advent of an action potential, however, calcium ions rush into the cell. The influx of calcium ions leads to the release of neurotransmitter into the synaptic cleft; this passes the signal to a neighboring nerve cell.
Having taken a close look at the electrical side of the picture, we are in a better position to see where the chemistry comes in. Molecules of neurotransmitter are released into a synaptic cleft and bind to specific receptor sites on the postsynaptic side (the dendrite or dendritic spine), thereby altering the ion channels in the postsynaptic membrane. Some neurotransmitters cause sodium channels to open, allowing the influx of Na+ ions and thus a lessening of negative charge inside the cell membrane. If a considerable number of these potentials are received within a short interval, they can depolarize the membrane enough to trigger an action potential; the result is the transmission of a nerve impulse. The substances that can cause this to occur are the excitatory neurotransmitters. By contrast, other chemical compounds cause potassium channels to open, increasing the outflow of K+ ions from the cell and making excitation less likely; the neurotransmitters that bring about this state are considered inhibitory.
A given neuron has a great quantity of sites available on its
dendrites and cell body and receives signals from many synapses simultaneously, both excitatory and inhibitory. These signals often amount to a rough balance; it is only when the net potential of the membrane in one region shifts significantly up or down from the resting level that a particular neurotransmitter can be said to be exerting an effect. Interestingly, in the membrane's overall balance sheet, the importance of a particular synapse varies with its proximity to where the axon leaves the nerve cell body, so that numerous excitatory potentials out at the ends of the dendrites may be overruled by several inhibitory potentials closer to the soma. Other kinds of synapse regulate the release of neurotransmitters into the synaptic cleft, where they go on to affect the postsynaptic channels as described above.
The list of known neurotransmitters, once thought to be quite short, continues to grow as more substances are found to be synthesized by neurons, contained in presynaptic boutons, and bound on the postsynaptic membrane by specific receptors. Despite stringent requirements for identifying a substance as a neurotransmitter (see Chapter 5 ), well over two dozen have been so named, and another several dozen strong candidates are under review.
The most cursory look at the human brain can excite awe at its complex functions, the intricacy of its structure, and the innumerable connections all maintained on microscopic fibers a few millionths of a meter in diameter. But a slightly more intimate acquaintance with this 3-pound organ inside our heads, an acquaintance that builds on observation of the brain in action and discovery of the principles by which it works, can yield something more satisfying than awe: the sense of mastery and of rewarded curiosity that comes with understanding. With the rewarding of curiosity as our goal, let us take a closer look at a few aspects of the functioning brain.





















