
CHAPTER TWO
Abrupt Changes of Primary Concern
The following section describes potential abrupt climate changes that are of primary concern, either because they are currently believed to be the most likely and the most impactful, because they are predicted to potentially cause severe impacts but with uncertain likelihood, or because they are considered to be unlikely to occur but have been widely discussed in the literature or media. As such, the Committee did not attempt to create a comprehensive catalog of potential abrupt changes. As described in the Introduction, this section examines both abrupt climate changes in the physical climate system itself and abrupt climate impacts in physical, biological, or human systems that are triggered by a steadily changing climate.
The Atlantic Meridional Overturning Circulation
The Atlantic Meridional Overturning Circulation (AMOC)—characterized by warm surface waters flowing northward and cold deep waters flowing southward throughout the Atlantic basin—is defined as the zonal integral of the northward mass flux at a particular latitude. The deep limb of this overturning circulation carries waters that are formed via convection in the Nordic and Labrador Seas (Figure 2.1). Collectively, these waters constitute North Atlantic Deep Water, which is exported to the global ocean at depths between about 1000 and 4000 m. The southward-flowing deep limb of the overturning circulation is compensated by an upper limb of northward-flowing surface waters, which head to the Nordic and Labrador Seas to replenish the regions of convection. Together, the upper and lower limbs of the overturning circulation produce a poleward flux of heat that has strong global and regional impacts. The AMOC also plays an important role in the transport of carbon in the Atlantic. Thus, variability in the AMOC’s strength is of much interest, as a diminishment or strengthening would impact the ocean’s effectiveness as a heat and carbon reservoir.
Examinations of paleoclimate temperatures and other variables recorded in both North Atlantic ocean sediments and Greenland ice cores (e.g., Lehman and Keigwin, 1992; Alley et al., 1993; Taylor et al., 1993) have led to suggestions that the AMOC
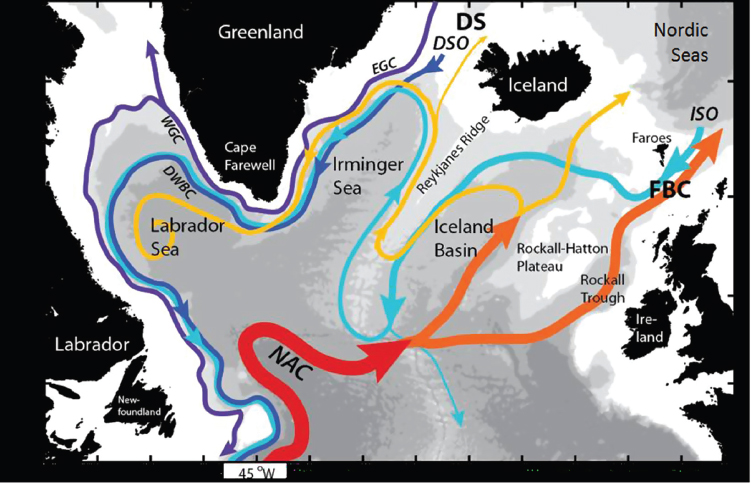
FIGURE 2.1 Schematic of the major warm (red to yellow) and cold (blue to purple) water pathways in the North Atlantic subpolar gyre. Acronyms not in the text: Denmark Strait (DS); Faroe Bank Channel (FBC); East and West Greenland Currents (EGC, WGC); North Atlantic Current (NAC); DSO (Denmark Straits Overflow); ISO (Iceland-Scotland Overflow). Figure courtesy of H. Furey (WHOI).
abruptly changed in the past. Following on this examination, questions have arisen as to the possible likelihood of an abrupt change in the future.
The Stability of the Atlantic Meridional Overturning Circulation
Climate and Earth system models are used to understand potential changes in the AMOC, including potential feedbacks in the system, although the representation of unresolved physics (such as the parameterization of ocean mixing) could potentially be of concern in long, centennial simulations. Because saltier water is denser and thus more likely to sink, the transport of salt poleward into the North Atlantic provides a potentially destabilizing advective feedback to the AMOC (Stommel, 1961); i.e., a reduction in the strength of the AMOC would lead to less salt being transported into the North Atlantic, and hence a further reduction in the AMOC would ensue. As noted
by Rahmstorf (1996), the presence of this slow salt-advection feedback is critical to the existence of stable multiple equilibria.
Climate and Earth system models have been used to investigate the stability of the AMOC, in particular the number of stable states that the system can exist in, which is an important characteristic to know for fully understanding the climate system. Carefully designed non-linear modeling experiments using Earth system Models of Intermediate Complexity (EMICs; and also the FAMOUS AOGCM; Hawkins et al., 2011) have revealed a model-dependent threshold beyond which an active AMOC cannot be sustained (Rahmstorf et al., 2005; see Figure 2.2). However, analysis of the AMOC in the models that submitted simulations in support of the third phase of the Community Model Intercomparison Project1 (CMIP3; Meehl et al., 2007a) suggested that the CMIP3 models were overly stable (Drijfhout et al., 2011; Hofmann and Rahmstorf, 2009), i.e., that an abrupt change in the AMOC was not likely to be simulated in the models even if it were to be likely in reality.
Several studies (de Vries and Weber, 2005; Dijkstra, 2007; Weber et al., 2007; Huisman et al., 2010; Drijfhout et al., 2011; and Hawkins et al., 2011) have suggested that the sign of the net freshwater flux into the Atlantic across its southern boundary via the overturning circulation determines whether or not the AMOC is in a monostable or bistable regime. Observations suggest that the present day ocean resides in a bistable regime, thereby allowing for multiple equilibria and a stable “off” state of the AMOC (Hawkins et al., 2011). By examining the preindustrial control climate of the CMIP3 models, Drijfhout et al. (2011) found that the salt flux was mostly negative (implying a positive freshwater flux), indicating that these models were mostly in a monostable regime. This was not the case in the CMIP5 models where Weaver et al. (2012) found that 40 percent of the models were in a bistable regime throughout their integrations. Although this question of the number of stable states of the system is important for a complete understanding of the climate system, it is important to emphasize that regardless of this stability question, the CMIP5 models also show no evidence of an abrupt collapse for the 21st century.
In addition to the main threshold for a complete breakdown of the circulation, other thresholds may exist that involve more-limited changes, such as a cessation or diminishment of Labrador Sea deep water formation (Wood et al., 1999). Rapid melting of the Greenland ice sheet causes increases in freshwater runoff, potentially weakening the AMOC. None of the CMIP5 simulations include an interactive ice sheet component. However, Jungclaus et al. (2006), with parameterized freshwater melt as high
____________________
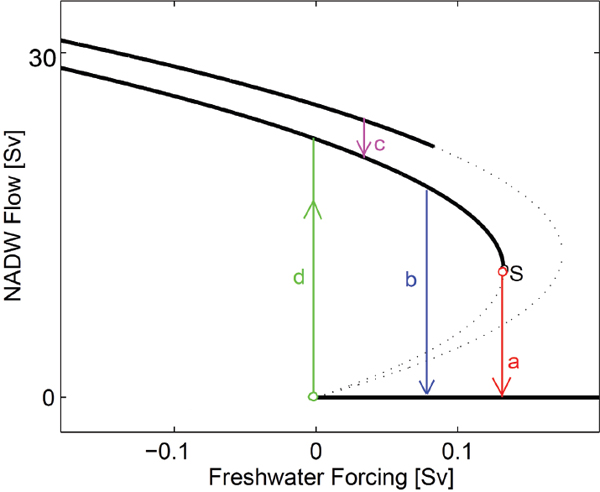
FIGURE 2.2 Schematic diagram illustrating the hysteresis behavior of the equilibrium strength of the AMOC in response to the addition of a North Atlantic surface freshwater perturbation of variable magnitude. Positive values indicate the sustained addition of freshwater to the surface; negative values indicate the sustained subtraction of freshwater from the surface; the zero value corresponds to the present-day situation. The two upper heavy branches indicate the possibility of multiple states with different convection sites. Transitions between stable equilibria of the AMOC with and without active deepwater formation are indicated by: (a) transition associated with slow advective instability, (b) transition associated with fast convective instability, and (d) initiation of convection and subsequent spin-up of North Atlantic Deep Water (NADW) formation. The S indicates the point beyond which a stable equilibrium with active NADW formation cannot exist. (c) indicates a possible transition between active modes of NADW formation with different location of convection.
Note: Hysteresis is defined as “a lag in response exhibited by a body in reacting to changes in forces” (Random House Kernerman Webster’s College Dictionary) and is used in many fields such as engineering, economics, biology, etc. to refer to a system that depends on the current but also past environmental conditions.
SOURCE: Rahmstorf, 1999.
as 0.09 Sv, and Hu et al. (2009), using the NCAR Community Climate System Model (CCSM) with year 2000 initial parameterized freshwater melt of 0.01 Sv increasing at a rate of 1 percent/year, 3 percent/year and 7 percent/year, as well as Mikolajewicz et al. (2007) and Driesschaert et al. (2007), using coupled ice-sheet/climate models, found only a slight temporary effect of increased melt water fluxes on the AMOC. The impact of these fluxes on the AMOC was generally small compared to the effect of enhanced poleward atmospheric moisture transport and ocean surface warming; or it was only noticeable in the most extreme scenarios. But this point needs to be further quantified.
While many more model simulations were conducted in support of the IPCC AR5 (Collins et al., 2012) under a wide range of forcing scenarios, projections of the behavior of the AMOC over the 21st century and beyond have changed little from what was reported in the IPCC AR4 (Meehl et al., 2007b). In the case of the CMIP5 models, Weaver et al. (2012) showed that the behavior of the AMOC was similar over the 21st century under four very different radiative forcing scenarios (RCP 2.6; RCP4.5; RCP 6.0; RCP8.5—these Representative Concentration Pathways [RCPs] are detailed in Moss et al., 2010). All models found a 21st century weakening of the AMOC with a multi-model average of 22 percent for RCP2.6, 26 percent for RCP4.5, 29 percent for RCP6.0 and 40 percent for RCP8.5. While two of the models eventually realized a slow shutdown of the AMOC under RCP8.5 (the scenario with the largest amount of warming), none exhibited an abrupt change of the AMOC.
The similarity of the model responses despite the widely varying transports of salt into the North Atlantic across its southern boundary (and hence sign and magnitude of the salt advection feedback) suggests that like the CMIP3 models (Gregory et al., 2005), the reduction of the AMOC in the global warming experiments performed by the CMIP5 models is mainly driven by local changes in surface thermal flux rather than surface freshwater flux. North Atlantic surface warming decreases water density there, thus reducing the rate of sinking. In addition, as noted above, none of the CMIP models incorporated the additional freshwater effects of ice sheet melting. This is an important caveat since asymmetric freshwater forcing is capable of initiating a fast, convective instability that could cause the AMOC to abruptly shut down if it were in a bistable regime and suitably close to its stability threshold. This would explain why abrupt changes of the AMOC appear to be pervasive features of the paleoclimate record when vast reservoirs of freshwater were available in the form of ice and proglacial lakes on land.
A question that needs to be further addressed is the extent to which projected changes in Greenland ice sheet melting could affect the amount and location of
freshwater release into the North Atlantic and hence the subsequent evolution of the AMOC. As noted in Meehl et al. (2007b) it is very unlikely that the AMOC will undergo an abrupt transition or collapse in the 21st century. Delworth et al. (2008) pointed out that for an abrupt transition of the AMOC to occur, the sensitivity of the AMOC to forcing would have to be far greater than that seen in current models. Alternatively, significant ablation of the Greenland ice sheet greatly exceeding even the most aggressive of current projections would be required. As noted in the ice sheet section later in this chapter, Greenland ice has about 7.3m equivalent of sea level rise, which, if melted over 1000 years, yields an annual rise rate of 7 mm/yr, about 2 times faster just from Greenland than today’s rate from all sources, and more than 10 times faster than the rate from Greenland over 2000–2011 (Shepherd et al., 2012). Although neither possibility can be excluded entirely, it is unlikely that the AMOC will collapse before the end of the 21st century because of global warming.
Observations of the Atlantic Meridional Overturning Circulation
Recent observational studies have focused on ascertaining two questions of relevance to the AMOC response to climate change: What is the impact of variable North Atlantic Deep Water production on the ocean’s meridional overturning? And, what is the current state of the AMOC and its variability? Studies relevant to both questions are briefly reviewed here (material drawn from Lozier, 2012).
Though many modeling studies have demonstrated the impact of deep water formation changes on the overturning circulation, the observational evidence for such a linkage has been hard to come by for two reasons: (1) Deep water formation is difficult to quantify because the time and locale of production are highly variable from winter to winter, and (2) overturning circulation measures require observations that span the basin, which have been limited in space and time. Because of this second difficulty, a measure of the Deep Western Boundary Current (DWBC) transport has traditionally been considered a shortcut to the measure of the AMOC: while the upper limb of the AMOC was considered inextricably linked to the much more energetic wind-driven circulation, the lower limb was considered to be “channeled” through the DWBC.
An opportunity to assess the linkage between deep water formation variability and DWBC changes was afforded by the deployment of a moored array east of the Grand Banks (Clarke et al., 1998; Meinen et al., 2000; Schott et al., 2006). In an extensive analysis of the time series from these two deployments, Schott et al. (2006) found that the transport rates of Labrador Sea Water (LSW) over these two time periods were remarkably similar despite the large differences in convective activity in the Labrador Sea
during the two time periods: the earlier time period was marked by strong convective activity, while LSW production was considerably weaker during the latter time period (Lazier et al., 2002). This result raised questions about the responsiveness of the AMOC to changes in deep water production; however, the linkage could not be conclusively ruled out because of increasing indications that the DWBC was not the sole conduit for the passage of deep waters to the lower latitudes (Schott et al., 2006). And in fact, recent observational (Lavender et al., 2000; Fischer and Schott, 2002; Bower et al., 2009) and modeling studies (Gary et al., 2011; Lozier et al., 2010) of subsurface floats have revealed that the DWBC is not the sole, and perhaps not even the dominant, conduit for the transport of the waters within the deep limb of the AMOC. Thus, a measure of the DWBC is no longer considered a sufficient monitor of AMOC changes.
For a full accounting of the AMOC and its variability, it is now understood that transbasin measurements of transport are necessary. Attempts to understand trans-basin AMOC variability over the modern observational record traditionally have had to rely on indirect estimates assessed from hydrography. Bryden et al. (2005) used five repeat surveys at 25°N from 1957 to 2004 to show that the overturning slowed by 30 percent over the period of the surveys, an astounding and unanticipated change over such a relatively short time. However, an assessment of transports at 48°N using five repeat World Ocean Circulation Experiment sections and air-sea heat and freshwater fluxes as input to an inverse box model yielded no significant trend in the meridional overturning at that latitude (Lumpkin et al., 2008), though the time period studied was relatively short (1993-2000).
In 2004 an observational system was put in place to provide the first continuous measure of the AMOC (Cunningham et al., 2007). The RAPID/MOCHA program (Rapid Climate Change/Meridional Overturning Circulation and Heatflux Array) comprises instruments deployed along a section at 25°N stretching from the North American continent to the west coast of Africa. After just one year of measurements, the conceptual understanding of overturning variability changed dramatically. As seen in Figure 2.3, the overturning strength changed six-fold from April of 2004 to April of 2005, from a minimum of ~5 Sv to a maximum of ~30 Sv. With the demonstrated intraseasonal variability, synoptic sections were now understood to be inadequate to capture measures of interannual transport variability. The continuation of the time series has revealed a strong seasonality (Rayner et al., 2011) that dominates the record, as well as strong intrannual variability (McCarthy et al., 2012).
Unfortunately, the strong intraseasonal variability of the AMOC revealed by the RAPID/MOCHA array seriously constrains our ability to recreate AMOC variability over the modern observational period, since synoptic hydrographic sections are the
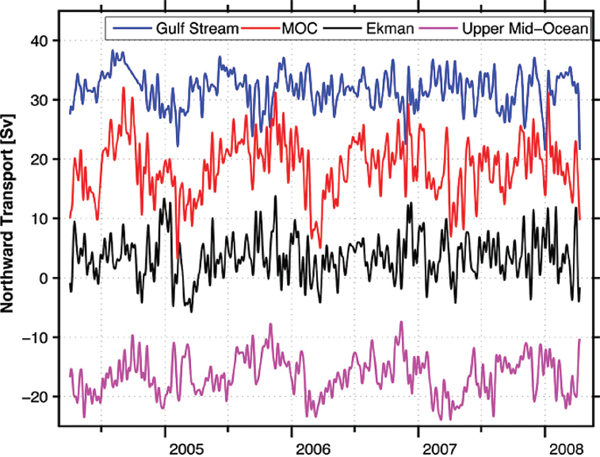
FIGURE 2.3 Time series of the meridional overturning transport at 26° N from the RAPID/MOCHA array. The meridional overturning circulation (MOC) is computed from the sum of the Gulf Stream transport through the Florida Straits, directly measured via electromagnetic cables; the Ekman transport, estimated from QuikSCAT winds; and the midocean geostrophic transport, estimated from the moored array instruments. Importantly, this time series demonstrates the significant interannual transport variability. SOURCE: Rayner et al., 2011.
only past trans-basin measurements. Furthermore, as detailed in a recent review by Cunningham and Marsh (2010), modeling estimates have been unable to help in this regard: there is currently no consensus on the strength of the AMOC in assimilation/ re-analysis products, and ocean general circulation models are in disagreement about the strength and variability of the AMOC. Indeed, an active area of research within the climate modeling community is focused on the cause for such wide ranges of AMOC estimates from state estimates that are drawing from the same observational databases (U.S. CLIVAR Project Office, 2011) and in ocean simulations forced with the same
atmospheric conditions (e.g., Danabasoglu et al., 2013). It is important to note that the models run for the IPCC discussed above also have varying AMOC strength and interannual variability, yet they are in agreement on a lack of abrupt change for the 21st century.
In lieu of consistent model estimates, proxy measures of the overturning derived from satellite altimetry and Argo float data are appealing, but to date these measures have been limited to latitudes of steep topography on the western boundary (Willis, 2010), and are of limited duration to provide a temporal context of decades. Thus, to date direct AMOC observations are limited to one latitude (26°N), and past measures of change remain elusive. Although the RAPID array is providing unprecedented measurements, recent modeling and data analysis studies (Bingham et al., 2007; Baehr et al., 2009; Lozier et al., 2010; Biastoch et al., 2008a; Biastoch et al., 2008b) reveal gyrespecific measures of the AMOC, suggesting that the AMOC variability measured by the RAPID array cannot safely be assumed representative of AMOC variability outside of the North Atlantic subtropical basin.
Summary and the Way Forward
Although models do not indicate that AMOC is likely to change abruptly in the coming decades, it is important to monitor the North Atlantic to confirm the understanding of how AMOC responds to a changing climate. Observational studies over the past decade or so reveal a meridional overturning circulation with a tenuous link to the production of deep water masses via local overturning at high latitudes in the North Atlantic. However, the deep ocean remains vastly undersampled, particularly so with respect to measures appropriate for the calculation of AMOC variability. To ascertain with confidence the extent to which deep water production impacts the ocean’s meridional circulation and hence the ocean’s contributions to the global poleward heat flux, continuous measures of trans-basin mass and heat transports are needed. Although such measurements are underway with the RAPID/MOCHA array, the studies cited above have made it increasingly clear that AMOC fluctuations are coherent over only limited meridional distances: break points in coherence occur at key latitudes, in particular at the subpolar/subtropical gyre boundary in the North Atlantic. Therefore, a transoceanic line in the subpolar North Atlantic, currently being planned by the international community, that measures the net contributions of the overflow waters from the Nordic Seas as well as those from the Labrador Sea, to the AMOC, would directly test the legitimacy of the decades-long supposition that variability in North Atlantic Deep Water production translates into meridional overturning variability (Figure 2.4). This measurement system would—in conjunction with the RAPID/
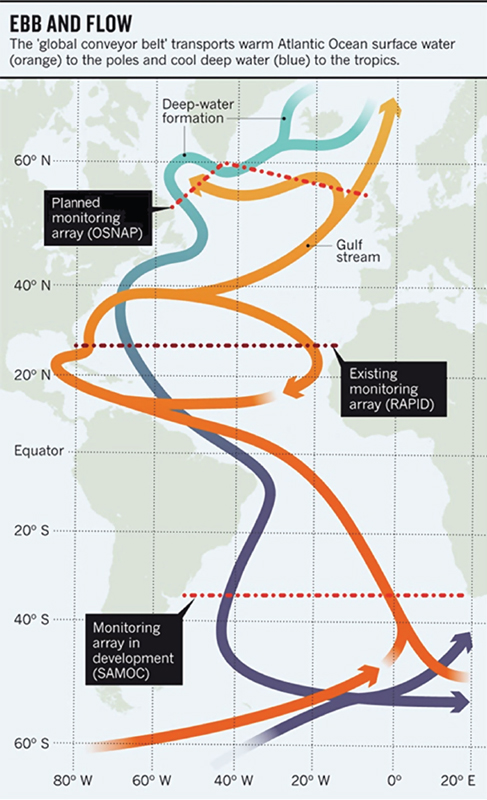
FIGURE 2.4 Existing and proposed monitoring locations for the Atlantic Ocean. Source: Adapted from Schiermeier, 2013.
MOCHA array—provide a means to evaluate intergyre connectivity within the North Atlantic and allow for a determination of how and whether deep water mass formation impacts overturning and poleward heat and freshwater transports throughout the North Atlantic. Additionally, such an observing system, by measuring the temporal and spatial variability of the AMOC for approximately a decade, would provide essential ground truth to AMOC model estimates and would also yield insight into whether AMOC changes or other atmospheric/oceanic variability have the dominant impact on interannual sea surface temperature (SST) variability. To make clear assessments of the AMOC’s response to anthropogenic climate change, it is expected that a multi-decadal observing system will be necessary. An observing system serving this purpose would be one where a few critical in situ observations, coupled with satellite observations and the Argo float array, provide a reliable and sustainable measure of the AMOC for decades to come.
Ice Sheets and Sea Level
Based on both simple physics and observations of the past, there is high confidence in the conclusion that sea level rises in response to warming. Sea-level rise can have large impacts (e.g., Nicholls et al., 2007), such as damage to or loss of infrastructure near coasts, loss of freshwater supplies, and displacement of people whose homes are lost to a rising ocean. Although sea-level rise typically is slow compared to many environmental changes, even this type of gradual sea-level rise may force other systems to cross thresholds and trigger abrupt impacts for natural or human systems unless adaptive measures are taken. For example, rising sea level increases the likelihood that a storm surge will overtop a levee or damage other coastal infrastructure, such as coastal roads, sewage treatment plants, or gas lines—all with potentially large, expensive, and immediate consequences (Nordhaus, 2010). (See Box 2.1 for discussion of vulnerabilities of US coastal infrastructure.)
A separate but key question is whether sea-level rise itself can be large, rapid and widespread. In this regard, rate of change is assessed relative to the rate of societal adaptation. Available scientific understanding does not answer this question fully, but observations and modeling studies do show that a much faster sea-level rise than that observed recently (~3 mm/yr over recent decades) is possible (Cronin, 2012). Rates peaked more than 10 times faster in Meltwater Pulse 1A during the warming from the most recent ice age, a time with more ice on the planet to contribute to the sealevel rise, but slower forcing than the human-caused rise in CO2 (Figure 2.5 and 2.6). One could term a rise “rapid” if the response or adaptation time is significantly longer than the rise time. For example, a rise rate of 15 mm/yr (within the range of projec-
BOX 2.1 VULNERABILITY OF U.S. COASTAL INFRASTRUCTURE
Thirty nine percent of the population lives in coastal shoreline counties. This population grew by 39 percent between 1970 and 2010, and is projected to grow by 8.3 percent by 2020. The population density of coastal counties is 446 people per sq mile, which is over 4 times that of inland counties.
Just under half of the annual GDP of the United States is generated in coastal shoreline counties, an annual contribution that was $6.6 trillion in 2011. If counted as their own country, these counties would rank as the world’s third largest economy, after the United States and China. Some portions of these counties are well above sea level and not vulnerable to flooding (e.g., Cadillac Mountain, Maine, in Acadia National Park, at 470 m). But, the interconnected nature of roads and other infrastructure within political divisions mean that sea-level rise would cause problems even for the higher parts of these counties. The following statistics, from NOAA’s State of the Coast,a highlight the wealth and infrastructure at risk from rising seas:
- $6.6 trillion: Contribution to GDP of the coastal shoreline counties, just under half of US GDP in 2011.b
- 51 million: Total number of jobs in the coastal shoreline counties of the US in 2011.c
- $2.8 trillion: Wages paid out to employees working at establishments in the coastal shoreline counties in 2011.d
- 3: Global GDP rank (behind the United States and China) of the coastal shoreline counties, if considered an individual country.e
- 39%: Percent of the nation’s total population that lived in coastal watershed counties in 2010 (less than 20 percent of the total land area excluding Alaska).f
- 34.8 million: Increase in US coastal watershed county population from 1970 to 2010 (or a 39 percent increase).g
- 446 persons/mi2: Average population density of the coastal watershed counties (excluding Alaska). Inland density averages 61 persons per square mile.h
- 37 persons/mi2: Expected increase in US coastal watershed county population by 2020 (or an 8.3 percent increase).i
Projections of sea-level rise remain notably uncertain even if the increase in greenhouse gases is specified accurately, but many recently published estimates include within their range of possibilities a rise of 1m by the end of this century (reviewed by Moore et al., 2013). For lowlying metropolitan areas, such as Miami and San Francisco, such a rise could lead to significant flooding (Figure A) (NRC, 2012e; Strauss et al., 2012; Tebaldi et al., 2012). In many cases, such areas would be difficult to defend by dikes and dams, and such a large sea level rise would require responses ranging from potentially large and expensive engineering projects to partial or nearcomplete abandonment of now-valuable areas as critical infrastructure such as sewer systems, gas lines, and roads are disrupted, perhaps crossing tipping points for adaptation (Kwadijk et al., 2010). Miami was founded little more than one century ago, and could face the possibility of sea level rise high enough to potentially threaten the city’s critical infrastructure in another century (Strauss et al., 2013). In terms of modern expectations for the lifetime of a city’s infrastructure, this is abrupt. If sometime in the coming centuries sea level should rise 20 to 25 m, as suggested
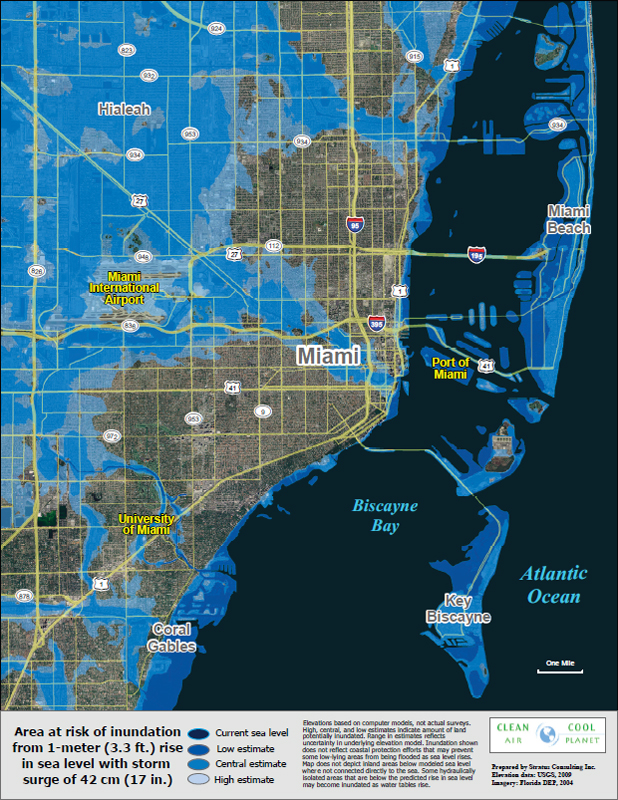
FIGURE A Elevation map of Miami, Florida. The low elevation of many parts of the city and surroundings, combined with a water-permeable sand and coral base, make it particularly vulnerable to sea-level rise. Areas at risk from a 1-meter rise in sea level are shown, where 1 meter is within the range of many recently published estimates for sea-level rise by the end of this century. SOURCE: Cool Air Clean Planet, http://cleanair-coolplanet.org/.
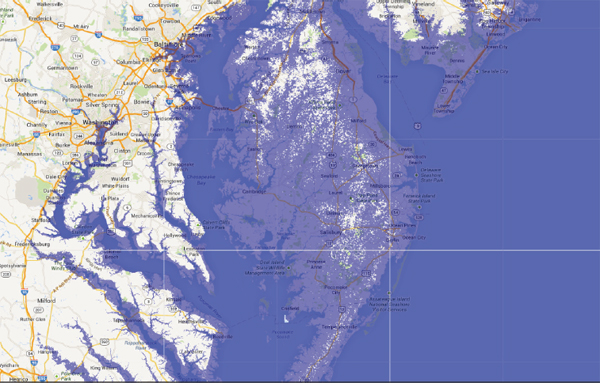
FIGURE B The long-term worst-case sea-level rise from ice sheets could be more than 60 m if all of Greenland and Antarctic ice melts. A 20 m rise, equivalent to loss of all of Greenland’s ice, all of the ice in West Antarctica, and some coastal parts of East Antarctica, is shown here. This may approximate the sea level during the Pliocene period (3–5 million years ago), the last time that CO2 levels are thought to have been 400 ppm. This figure emphasizes the large areas of coastal infrastructure that are potentially at risk if substantial ice sheet loss were to occur. SOURCE: http://geology.com/sea-level-rise/washington.shtml.
for the Pliocene Epoch, 3 to 5 million years ago (see Figure 2.5), when CO2 is estimated to have had levels similar to today of roughly 400 parts per million, most of Delaware, the first State in the Union, would be under water without very large engineering projects (Figure B). In terms of the expected lifetime of a State, this could also qualify as abrupt.
___________________
a http://stateofthecoast.noaa.gov/coastal_economy/.
e http://www.bls.gov/cew/; http://data.worldbank.org/indicator/NY.GDP.MKTP.CD.
f http://factfinder2.census.gov/faces/nav/jsf/pages/index.xhtml.
g http://factfinder2.census.gov/faces/nav/jsf/pages/index.xhtml.
h http://factfinder2.census.gov/faces/nav/jsf/pages/index.xhtml.
i Woods and Poole Economics Inc., 2011; http://coastalsocioeconomics.noaa.gov/.
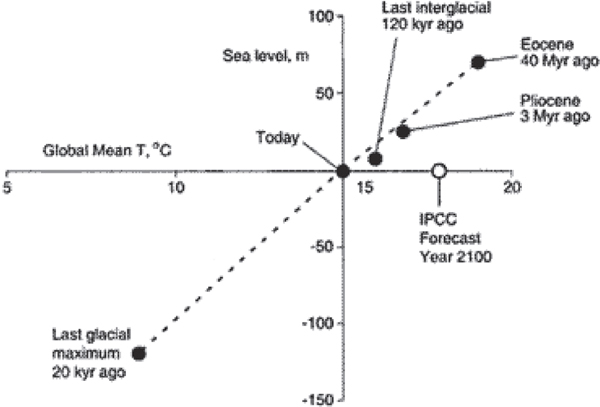
FIGURE 2.5 Co-variation of sea level with global average temperature in the geologic past, compared with the IPCC forecast for sea level rise by the year 2100. Figure from Archer, 2010. The straight line shown may be less accurate than a curve passing through each of the solid dots shown (e.g., Gasson et al., 2012), but sufficient warmth gives large sea-level rise in models and has done so in Earth’s history.
tions for this century, although on the high side; Moore et al., 2013) sustained over 33 years would be abrupt for an airport runway that is built to last longer than that (and financed over that time period), but becomes unusable with a half meter of sea-level rise (assuming no adaptive measures are taken).
Rise of the global average sea level over the time periods of most interest to human economies is controlled primarily by the mass or density of ocean water. Local relative sea level may be affected by several additional factors (Box 2.2).
For global average sea level, the main control on water density over these times is ocean temperature, with warming causing thermal expansion by roughly 0.4 m per degree C (Levermann et al., 2013). In response to atmospheric warming, the temperature of the bulk of the ocean will increase primarily through downward transport of water heated at the surface. Because the time for water to move through the deep
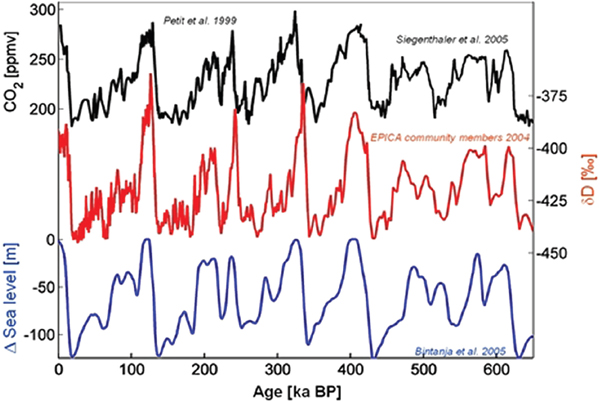
FIGURE 2.6 Sea level and climate over the last 600,000 years. Compilation of Vostok and EPICA Dome C CO2 concentrations (Petit et al., 1999; Siegenthaler et al., 2005) and δD (deuterium isotope record) as a proxy for local air temperature (Petit et al., 1999; Augustin et al., 2004) and the changes in global sea level relative to the present level (Bintanja et al., 2005). To a first approximation, sea-level changes reflect the volume of ocean water bound in continental ice sheets during the ice ages. CO2 minima were reached approximately when the sea level was at a minimum, hence, the extent of the continental ice sheets were at a maximum, and the highest CO2 levels were found during interglacials during the high stands of the sea level. The evolution of the local temperature (as deduced from δD) follows this overall picture and points to a strong coupling of the climate and the carbon cycle. Source: http://www.awi.de/en/research/research_divisions/geosciences/glaciology/techniques/high_precision_d13c_and_co2_analysis/
ocean is of the order of 1000 years, thermal expansion is expected to be relatively slow and predictable, although shifts in ocean circulation can influence the details of the warming and sea-level rise. Ocean mass over human time-scales is affected primarily by shifting water between ocean and land. Water may be stored on land in lakes or glaciers, and in spaces in the earth (whether the water is frozen in permafrost, or liquid as groundwater). Extraction of water from the ground for human use may exceed natural recharge, and that water quickly reaches the ocean (Lowe and Gregory, 2006; Headly and Severinghaus, 2007), raising sea level. However, increased storage in
BOX 2.2 LOCAL SEA-LEVEL CHANGE
Local sea-level change can differ notably from the global average for multiple reasons. Changes in land elevation may occur in response to many processes, including mountainbuilding (tectonic) processes, or flow or bending of rocks caused by ongoing or past changes in loading from ice, water or sediment (isostatic changes). In addition, compaction following removal of groundwater or fossil fuels, or possibly inflation from injection of fluids, may change land elevation (e.g., Bindoff et al., 2007; Sella et al., 2007). These changes can locally accelerate, reduce, or even reverse the global-mean trend, especially if the trend is not too much larger than that observed recently.
In addition, the ocean surface is not level, but exhibits topography caused by winds piling up water along some coast lines and pushing it offshore in other areas, and because different places have water with different local temperature and salinity, and thus density. For rising-CO2 model experiments tracking warming of the ocean with changing water density and winds, some sites exhibit simulated local sea-level rise that is twice the global-average rise, and other sites exhibit no rise (e.g., Lowe and Gregory, 2006).
Also important is the self-gravitation of ice sheets (e.g., Mitrovica et al., 2001). All masses are gravitationally attracted to other masses. The great bulks of the Greenland and Antarctic ice sheets actually have pulled ocean water toward them, so that their coastal sea levels are notably higher than they would be without that gravitational attraction. If the ice melts, adding water to the ocean, it is useful then to think of a two-step process (see Figure A), although the steps are coupled. First, the world ocean rises rapidly and nearly uniformly, within months or years, as the water from the melting ice spreads around the globe. Second, the ocean near the melting ice sheet drops because the smaller ice-sheet mass has less gravitational attraction for ocean water than before, and thus the water released from the former gravitational attraction of the ice sheet causes additional sea-level rise far from the ice sheet. Very near an ice sheet, the second effect may be larger than the first, with ice-sheet melting causing sea-level fall. Only a tiny fraction of people live near the world’s great ice sheets, and for most of the world’s coastlines the resulting local rise in sea level is larger than the global average, perhaps approaching 50 percent faster than the global average. Changes in Earth’s rotation from the redistribution of mass as the ice melts and ocean responds also contribute slightly to local deviations from the global average.
As discussed in the main text, however, if the large ice sheets were to begin to melt rapidly, the influence of this water being added to the ocean could greatly exceed all of these other effects except for self-gravitation for the vast majority of coastal sites. In such circumstances, almost all coastal sites removed from the immediate vicinity of the melting ice sheet would expect local sea-level rise proportional to the ice-sheet melting.
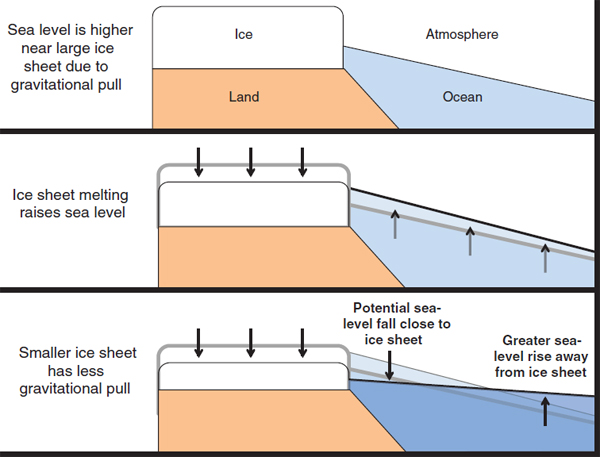
FIGURE A Effect of “self-gravitation” of ice sheets on sea-level rise during ice-sheet melting. The gravitational attraction from the mass in an ice sheet causes sea level to be higher near the ice than the global average. Melting of ice raises the global average sea level, and reduces the gravitational attraction from the ice, which allows the sea level near the ice to fall while sea level far from the ice rises more than the global average.
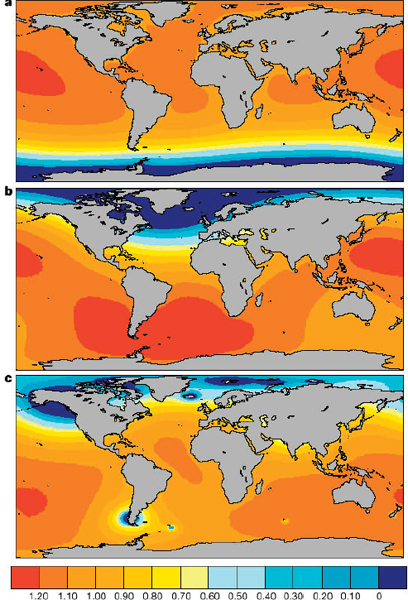
FIGURE B Predicted geometries of sea-level change due to continuing ice mass variations. Values shown are ratios of sea-level rise from enhanced analysis to those from a uniform sea-level rise. These normalized global sea-level variations were computed for the case of present-day ice mass variations in (a) Antarctica and (b) Greenland, as well as (c) melting of the mounting glaciers and ice sheets tabulated by Meier. In (a) and (b) it is assumed that the mass variation is uniform over the two polar regions. The results are normalized by the equivalent eustatic sea-level change for each mass flux event (see original reference). Departures from a contour value of 1.0 reflect departures from the assumption that the sea-level distribution accompanying these mass flux events is uniform. Predictions are based on a new sea-level theory solved using a pseudo-spectral algorithm with truncation at spherical harmonic degree and order 512. This truncation corresponds to a spatial resolution of 40 km. Source: Mitrovica et al., 2001.
new human-made impoundments or in groundwater fed by such impoundments will lower sea level. These partially offsetting effects lead to the expectation that direct human shifts in water storage on land will not have large effects on sea level in comparison to the effects of ocean warming and mountain-glacier and ice-sheet melting (Wada et al., 2012), although notable uncertainties remain in regards to future groundwater use and reservoir construction, and these effects vary considerably depending on the specific location (NRC, 2012e).
Most mountain glaciers worldwide are losing mass, contributing to sea-level rise. However, the amount of water stored in this ice is estimated to be less than 0.5 m of sea-level equivalent (Lemke et al., 2007), so the contribution to sea-level rise cannot be especially large before the reservoir is depleted. On the other hand, the reservoir in the polar ice sheets is sufficient to raise global sea level by more than 60 m (Lemke et al., 2007). Thus any large and rapid global sea-level rise, if it were to occur, would almost surely be sourced from the ice sheets.
Potential Abrupt Changes to Polar Ice Sheets
Ice-sheet volume is controlled by the balance between mass input and mass loss; mass input is almost entirely due to snowfall, and mass loss is from iceberg calving supplied by flow of the ice sheet, or runoff of melt water. As summarized in, for example, Meehl et al. (2007b), warming is expected to increase snowfall in the colder parts of Greenland and in Antarctica, and to increase melting in the warmer parts of Greenland. Beyond some threshold of a few degrees C warming, Greenland’s ice sheet will be almost completely removed. However, the timescale for this is expected to be many centuries to millennia, depending somewhat on the model used and more strongly on the emissions pathway (e.g., Meehl et al., 2007b). This still could result in a relatively rapid rate of sea-level rise. Greenland ice has about 7.3 m equivalent of sea-level rise (Lemke et al., 2007), which, if melted over 1000 years (a representative rather than limiting case), yields an annual rise rate of 7 mm/yr just from Greenland, slightly more than twice as fast as the recent rate of rise from all sources including melting of Greenland’s ice. Slower melting would obviously yield lower average rates, but the potential for rapid rise still exists. Surface melting removing the Antarctic ice sheet would require much more warming than in Greenland.
The loss of land ice by melting would be reversible if sufficient cooling were applied sufficiently rapidly (Ridley et al., 2010). For example, if the temperature increased across the threshold needed to remove the modern Greenland ice sheet, almost immediate cooling even slightly below the threshold would allow ice-sheet persistence.
Ice-sheet shrinkage with continuing melting lowers the surface into warmer parts of the atmosphere and warms the surroundings by replacing reflective snow and ice with dark rock, thus requiring greater cooling to allow regrowth.
Mass loss by flow of ice into the ocean is less well understood, and it is arguably the frontier of glaciological science where the most could be gained in terms of understanding the threat to humans of rapid sea-level rise. Increased ice-sheet flow can raise sea level by shifting non-floating ice into icebergs or into floating-but-still-attached ice shelves, which can melt both from beneath and on the surface. Rapid sea-level rise from these processes is limited to those regions where the bed of the ice sheet is well below sea level and thus capable of feeding ice shelves or directly calving icebergs rapidly, but this still represents notable potential contributions to sea-level rise, including the deep fjords in Greenland (roughly 0.5 m; Bindschadler et al., 2013), parts of the East Antarctic ice sheet (perhaps as much as 20 m; Fretwell et al., 2013), and especially parts of the West Antarctic ice sheet (just over 3 m; Bamber et al., 2009).
In understanding the behavior of ice sheets, attention is particularly focused on the boundary between the floating ice and grounded ice, which is usually called the grounding line, although in detail it is a zone with interesting but imperfectly understood properties (e.g., Schoof, 2007; Joughin et al., 2012a; Walker et al., 2013); see Figure 2.7. Large changes in ice mass are generally tightly coupled to grounding-line migration. If the ice-sheet bed deepens toward the center of the ice sheet, an instability exists, such that in the absence of additional stabilizers, the grounding line will advance with ice-sheet growth, or retreat with ice-sheet shrinkage, to a position where the bed rises towards the ice-sheet center. This instability can be overcome by a local reversal of the bed or narrowing of a fjord, and especially by friction between ice shelves and fjord walls or local highs in the sea floor.
The important role of ice shelves in stabilizing marine ice sheets introduces the potential for large and rapid ice-sheet shrinkage. The warmest upper surfaces of ice sheets are generally on ice shelves, because they are the lowest-elevation parts, and extend away from the cold central regions towards generally warmer oceans. Where meltwater forms on the ice-shelf surface, it can wedge open crevasses and cause ice-shelf disintegration, much like a line of balanced dominoes falling over, which has been observed to occur within weeks in the rapidly warming Antarctic Peninsula region (e.g., MacAyeal et al., 2003). Ice shelves are in contact with ocean water, and any warming of the water or increase in circulation of warm water under the shelves contributes to faster melting. Thinning or loss of ice shelves reduces friction, allows faster flow of the non-floating ice feeding the shelves, and thus contributes to sea-level rise. Furthermore, the recent behavior of ice shelves in Greenland (Nettles and Ekstrom, 2010),
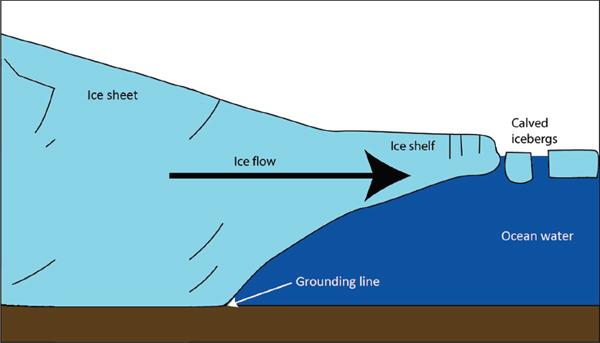
FIGURE 2.7 Schematic showing the grounding line for an ice sheet.
SOURCE: Adapted from www.AntarcticGlaciers.org by Bethan Davies. Used with permission.
the sedimentary record (Jakobsson et al., 2011), and our physical understanding (Alley et al., 2008) suggest that beyond some threshold ice-shelf reduction leads to complete loss as the ice shelf calves away, potentially in less than or much less than one year.
Model results, and the sedimentary record of deglaciated marine regions in both hemispheres, show that grounding lines tend to stabilize on local seafloor highs or fjord narrowings, and then to increase that stability through sedimentation (Anandakrishnan et al., 2007; Alley et al., 2007; Dowdeswell et al., 2008). In this stabilized position, the grounding line is little-affected by sufficiently small environmental forcings (e.g., Horgan and Anandakrishnan, 2006). However, for forcing beyond some threshold, the grounding line migrates rapidly to a new position of stability, which may be far away and involve an important contribution to sea-level change.
The possible rates of this migration are poorly quantified. The changes occurring now in drainages including Jakobshavn Glacier in Greenland (e.g., Joughin et al., 2012b) may be analogous to the events during retreat following the last ice age, but there at least are suggestions that past changes were sometimes faster (Jakobsson et al., 2011). The rate of discharge into deep water across a grounding line in general increases
with the thickness of the ice and the width of the fjord or channel through which the ice discharges. A retreat of Thwaites Glacier in West Antarctica could give a much wider and deeper calving front than any observed today, so the “speed limits” suggested by Pfeffer et al. (2008) may not apply (Parizek et al., 2013).
Because the full suite of physical processes at the grounding line (e.g., Walker et al., 2013) in general is not represented in modern models, the possibility exists that rates produced by extant models under strong simulated forcing may be greatly in error (Nowicki et al., 2013). Deglaciation of the marine portions of West Antarctica would raise sea level by ~3.3 m (Bamber et al., 2009), with additional marine ice in East Antarctica and Greenland, as noted above. Improved understanding of the retreat rates of WAIS and other marine based ice drainage zones is necessary to narrow the currently broad uncertainties and better quantify the potential worst-case scenarios.
Much process-based research coupling field work, remote sensing, and modeling is required to advance assessment of the likelihood of a threshold-crossing leading to abrupt sea-level rise from the ice sheets, as well as to improve projections of moregradual sea-level rise that could lead to threshold-crossing events in other systems. Great progress has been made recently in assessing the current rate of mass loss from the ice sheets (Shepherd et al., 2012), as well as monitoring the changing snowfall, surface melting, and temperature contributing to the changes. But maintenance and expansion of this effort are threatened, especially by possible loss of satellite observing capabilities (NRC, 2012d). The strong coupling of ice sheets and surrounding oceans (Joughin et al., 2012a) was not fully anticipated in early modeling efforts, and is not now fully represented in comprehensive Earth-system models. Key environmental information includes air temperatures and ocean temperatures in the upper kilometer of the ocean, sea ice, and related oceanic properties. Air temperature is relatively well monitored, although observations in remote polar areas are not dense, and that lack of data density can be problematic. More fixed monitoring sites as well as UAV-based observations are needed in the remote areas of both poles. Ocean temperatures are not well monitored, particularly in polar regions and particularly near the grounding lines and along the ice-ocean interface for marine-based ice. A concerted effort is needed to collect better data for constraining ocean conditions.
The second part of monitoring is to continually catalog those sensitive parts of coastal systems, human and natural, that are vulnerable to the impacts of a slowly or rapidly rising sea level and may exhibit abrupt behavior in response to a rising sea level, as well as the local changes in sea level. New surveys should be a routine part of environmental monitoring. Given the slow speed of sea-level rise, such new surveys need not
be annual, but regular surveys are needed at a frequency that takes into account local building rates as well as local rates of sea-level change.
Summary and the Way Forward
Sea level is rising, primarily in response to a warming planet, through thermal expansion of the oceans, and also via the loss of land ice as ocean and air temperatures increase, melting ice and speeding the flow of non-floating ice to form floating icebergs. Thermal expansion is expected to continue, and to be a slow but steady process. Loss of land ice by direct melting is also expected to be slow and steady. Both of these processes are likely to contribute to abrupt change via a slowly rising sea level forcing other systems to cross thresholds. Examples of such systems include human-built infrastructure at or near the coasts, natural ecosystems, freshwater in the ground, and natural dams or topographic choke points. Storm surges are exacerbated by rising sea level, amplifying the extent of the problem. Moving forward, tracking sea-level rise will require maintenance and expansion of the monitoring of sea level (tide gauges and satellite data), ocean temperatures at depth, and local coastal motions.
Sea level itself may also increase rapidly enough to be termed abrupt (O’Leary et al., 2013). The loss of land ice, particularly from marine-based ice sheets such as the West Antarctic Ice Sheet—possibly in response to gradual ocean warming—could trigger sea-level rise rates that are much higher than ongoing. Paleoclimatic rates at least 10 times larger than recent rates have been documented, and similar or possibly higher rates cannot be excluded in the future. This time scale is also roughly that of humanbuilt infrastructure such as roads, water treatment plants, tunnels, homes, etc. Deep uncertainty persists about the likelihood of a rapid ice-sheet “collapse” contributing to a major acceleration of sea-level rise; for the coming century, the probability of such an event is generally considered to be low but not zero (e.g., Bamber and Aspinall, 2013). To reduce this uncertainty moving forward, extensive effort is required, including the maintenance and expansion of satellite measurements, aerogeophysical monitoring, surface monitoring, process studies, and modeling.
Because air carries much less heat than an equivalent volume of water, physical understanding indicates that the most rapid melting of ice leading to abrupt sea-level rise is restricted to ice sheets flowing rapidly into deeper water capable of melting ice rapidly and carrying away large volumes of icebergs. In Greenland, such deep water in contact with ice is restricted to narrow bedrock troughs where friction between ice and fjord walls limits discharge. Thus, the Greenland ice sheet is not expected to destabilize rapidly within this century. However, a large part of the West Antarctic
Ice Sheet (WAIS), representing 3-4 m of potential sea-level rise, is capable of flowing rapidly into deep ocean basins. Because the full suite of physical processes occurring where ice meets ocean is not included in comprehensive ice-sheet models, it remains possible that future rates of sea-level rise from the WAIS are underestimated, perhaps substantially. Improved understanding of key physical processes and inclusion of them in models, together with improved projections of changes in the surrounding ocean, are required to notably reduce uncertainties and to better quantify worst-case scenarios. Because large uncertainties remain, the committee judges an abrupt change in the WAIS within this century to be plausible, with an unknown although probably low probability.
Changes in Ocean Chemistry and Associated Effects on Marine Ecosystems
Increasing carbon dioxide concentrations in the atmosphere, changing windstress, currents and mixing, and increasing temperatures are changing the chemistry of the world’s oceans. These changes are resulting in a decrease in pH, carbonate ion concentrations, and dissolved oxygen in the ocean.
Ocean Acidification
Carbon dioxide combines with water to form carbonic acid, which then dissociates to form bicarbonate ions and hydrogen ions (H+), so that increasing concentrations of CO2 in the atmosphere have been decreasing the pH (acidifying) of the surface ocean (NRC, 2010c). Since the preindustrial period, pH has declined by approximately 0.1 pH unit corresponding to a 30 percent increase in acidity. By 2100, the acidity is projected to increase by 100-150 percent compared to preindustrial values. Geologic records indicate that when the increase in atmospheric CO2 is gradual, oceanic pH and carbonate levels remain relatively stable due to processes that occur in equilibrium, such as dissolution of CaCO3 shells, weathering of terrestrial rock, and tectonic processes. However, the current rate of increase of atmospheric CO2 exceeds the rate at which natural processes can buffer these pH changes.
Although ocean acidification is not an abrupt climate change, i.e., the change occurs at the same rate as the forcing, the impacts of ocean acidification on ocean biology have the potential to cause rapid (over multiple decades) changes in ecosystems and to be irreversible when contributing to extinction events. Specifically, the increase in CO2 and HCO3– availability might increase photosynthetic rates in some photosynthetic marine organisms, and the decrease in CO32– availability for calcification makes
it increasingly difficult for calcifying organisms (such as some phytoplankton, corals, and bivalves) to build their calcareous shells and effects pH sensitive physiological processes (NRC, 2010c, 2013). As such, ocean acidification could represent an abrupt climate impact when thresholds are crossed below which organisms lose the ability to create their shells by calcification, or pH changes affect survival rates (see the Extinctions section below for more discussion of these issues).
Scientists are particularly concerned with the ability of reef-building corals to persist in the face of ocean acidification combined with the other stressors such as temperature increase, sea level rise, and changes in storm intensity all also associated with climate change. In addition, eutrophication and physical injuries inflicted from SCUBA diving and fishing further impact the health of coral ecosystems. Cascading events could irreversibly alter reef ecosystems on short time scales. However, the understanding of the effects on marine ecosystems is too limited to be able to draw any conclusions about the magnitude and rate of changes to come.
In addition, combined with the decline in oxygen availability, ocean acidification has the potential to impair aerobic respiration (see further discussion below). Changes in near-coastal circulation or biochemistry seem to be altering surface ocean pH more quickly than can be explained by an equilibrium response to the rising atmospheric CO2 concentration (Wootton and Pfister, 2012). This topic requires further research (see discussion below in this section).
Oxygen Content in the Global Ocean
The oxygen content in the surface ocean is projected to decline with warming because of the decrease in solubility of gases with increasing temperature, and changes in ventilation and biological consumption. A significant decrease in oxygen in the upper ocean between the 1970s and 1990s has already been observed at a global scale (Helm et al., 2011). Only approximately 15 percent of that decline can be attributed to a warmer mixed-layer, with the remainder being “consistent with an overall decrease in the exchange between surface waters and the ocean interior” (Helm et al., 2011). With a general weakening of ventilation rates as a result of climate change (Bryan et al., 2006), oxygen content of the global ocean is likely to further decrease (ventilation to the surface allows new input of oxygen from the atmosphere).
Of more immediate concern is the expansion of Oxygen Minimum Zones (OMZs). Photosynthesis in the sunlit upper ocean produces O2, which escapes to the atmosphere; it also produces particles of organic carbon that sink into deeper waters before they
decompose and consume O2. The net result is a subsurface oxygen minimum typically found from 200–1000 meters of water depth, called an Oxygen Minimum Zone.
Warming ocean temperatures lead to lower oxygen solubility. A warming surface ocean is also likely to increase the density stratification of the water column (i.e., Steinacher et al., 2010), altering the circulation and potentially increasing the isolation of waters in an OMZ from contact with the atmosphere, hence increasing the intensity of the OMZ. Thus, oxygen concentrations in OMZs fall to very low levels due to the consumption of organic matter (and associated respiration of oxygen) and weak replenishment of oxygen by ocean mixing and circulation. Furthermore, a hypothetical warming of 1°C would decrease the oxygen solubility by 5 μM (a few percent of the saturation value). This would result in the expansion of the hypoxic2 zone by 10 percent, and a tripling of the extent of the suboxic zone (Deutsch et al., 2011). With a 2°C warming, the solubility would decrease by 14 μM resulting in a large expansion of areas depleted of dissolved oxygen and turning large areas of the ocean into places where aerobic life disappears. In the tropical Atlantic, Pacific, and Indian Ocean, a decline in oxygen content in the subsurface waters has been confirmed with observations (Stramma et al., 2010).
The expansion and intensification of existing OMZs and the increase in CO2 are likely to pose a threat to aerobic marine life (Brewer and Peltzer, 2009). The amount of dissolved oxygen that marine life requires depends on how oxygen can diffuse across tissue boundaries, which is a function of oxygen content, temperature, and pressure. Some researchers have suggested that a respiration index (RI) be defined—based on the ratio of the partial pressures of oxygen and CO2—as a better metric for estimating the physiological limits of deep sea animals (Brewer and Peltzer, 2009). The use of this particular respiratory index has been disputed (Seibel and Childress, 2013), but it would be useful to develop a metric that could allow for a better assessment of the global extent of water masses where aerobic organisms could not survive. It could also contribute to improving early detection of thresholds for mass mortalities of aerobic organisms, which is of particular importance considering the economic value the fishing industry.
Limits to aerobic life in the sea are often defined as ~5 μM, below which it is inefficient for aerobic microbes to consume dissolved oxygen (Brewer and Peltzer, 2009). While some species adapted to lower-oxygen conditions, paleo records have shown the extinctions of many benthic species during past periods of hypoxia. These periods have
____________________
2 Hypoxia is the environmental condition when dissolved water column oxygen (DO) drops below concentrations that are considered the minimal requirement for animal life. Suboxia is even further depletion of oxygen and anoxia is the condition of no oxygen at all.
coincided with both a rise in temperature and sea level. Records also indicate long recovery times for ecosystems affected by hypoxic events (Danise et al., 2013).
In addition, when the oxygen in seawater is depleted, bacterial respiration of organic matter turns to alternate electron-acceptors with which to oxidize organic matter, such as dissolved nitrate (NO3–). A by-product of this “denitrification” reaction is the release of N2O, a powerful greenhouse gas with an atmospheric lifetime of about 150 years. Low-oxygen environments, in the water column and in the sediments, are the main removal mechanism for nitrate from the global ocean. An intensification of oxygen depletion in the ocean therefore also has the potential to alter the global ocean inventory of nitrate, affecting photosynthesis in the ocean. However, the lifetime of nitrate in the global ocean is thousands of years, so any change in the global nitrate inventory would also take place on this long time scale.
Likelihood of Abrupt Changes
Changes in global ocean oxygen concentrations have the potential to be abrupt because of the threshold to anoxic conditions, under which the region becomes uninhabitable for aerobic organisms including fish and benthic organisms. Once this tipping point is reached in an area, anaerobic processes would be expected to dominate resulting in a likely increase in the production of the greenhouse gas N2O. Some regions like the Bay of Bengal already have low oxygen concentrations today (Delaygue et al., 2001), but not quite low enough for denitrification to occur. Modest increases in the export of organic matter, or decreases in ventilation by the circulation, could decrease oxygen below the critical threshold for fixed nitrogen loss.
OMZs have also been intensified in many areas of the world’s coastal oceans by runoff of plant fertilizers from agriculture and incomplete wastewater treatment. These ‘dead zones’ have spread significantly since the middle of the last century and pose a threat to coastal marine ecosystems (Diaz and Rosenberg, 2008).This expansion of OMZs is due to nutrient runoff makes the ocean more vulnerable to decreasing solubility of O2 in a warmer ocean. Indeed, as warming of the ocean intensifies, the decrease in oxygen availability might become non-linear; particularly, as indicated by the expansion of the size of the oxygen minimum zone (Deutsch et al., 2011). The effect of temperature on oxygen solubility is well understood. However, it remains a major scientific challenge to model and project the changes of the magnitude and intensity of subsurface oxygen depletion because it depends on changes in ocean circulation, rates of de-nitrification, and nutrient runoff from land, and because global data coverage for chemical and biological parameters remains poor.
Summary and the Way Forward
In order to understand and possibly anticipate changes to the chemistry of the world’s oceans, the oxygen content, pH, and temperature of subsurface waters need to be monitored at the global, synoptic scale. The majority of the available oxygen data stem from the coastal oceans or from the World Ocean Circulation Experiment (WOCE) that took shipboard measurements across large portions of the ocean. However, the data remain too sparse in time and space (Stramma et al., 2010) to be able to detect longterm trends with confidence.
As oxygen sensors have become more sophisticated and accurate, they can be deployed more widely on buoys and floats. The current monitoring effort would ideally be expanded to equip Argo floats with oxygen sensors to achieve more global coverage in oxygen data. In order to better understand the effects of ocean chemistry on marine ecosystems, oxygen monitoring needs to be supplemented with biological observations at some select sites.
OMZs are not well represented in global climate models due to limited understanding of the physical and biological processes that affect them. In particular, the processes that lead deep water to be exchanged with the surface water remain poorly understood; for example, how rapidly a given parcel of ocean water is ventilated needs to be better resolved. Understanding such processes would enable models to be improved. Thus, physical processes such as vertical and isopycnal mixing that drive large scale circulations need to be better understood to improve the predictive capability and accuracy of the models.
Furthermore, research would benefit from new and standardized methods. For example, oxygen data need to be accompanied by contemporaneous pressure and temperature data as these variables combined give a better indication of how readily oxygen can diffuse across tissue boundaries. In addition, scientists could benefit by using common definitions for hypoxia, suboxia, and anoxia (Hofmann et al., 2011). While much research on the effects of shallow coastal dead zones has been published, little is understood on how this expansion will affect open ocean ecosystems.
Lastly, biological processes need to be better understood, including the microbial processes in OMZs, as well as how much larger organisms are affected and can adapt to the changes in OMZs. Resolving these questions would require a major effort given that OMZs represent relatively remote and under-sampled areas of the ocean.
ABRUPT CHANGES IN THE ATMOSPHERE
Atmospheric Circulation
The climate system exhibits variability on a range of spatial and temporal scales. On large (i.e., continental) scales, variability in the climate system tends to be organized into distinct spatial patterns of atmospheric and oceanic variability that are largely fixed in space but fluctuate in time. Such patterns are thought to owe their existence to internal feedbacks within the climate system.
Prominent patterns of large-scale climate variability include:
- the El-Nino/Southern Oscillation (ENSO),
- the Madden-Julian Oscillation (MJO),
- the stratospheric Quasi-Biennial Oscillation,
- the Pacific-North American pattern, and
- the Northern and Southern annular modes (the Northern annular mode is also known as the North Atlantic Oscillation).
All have a pronounced signature in atmospheric variability, and all owe their existence to internal climate dynamics. For example, ENSO is characterized by episodic warming and cooling of the eastern tropical Pacific, owes its existence to feedbacks between the tropical ocean and atmosphere, and fluctuates on timescales of ~2-7 years. The annular modes are characterized by north-south vacillations in the jetstream at middle latitudes, owe their existence to internal atmospheric dynamics, and fluctuate on timescales spanning weeks to decades. (In the discussion that follows, the middle latitude jetstreams are the eastward flowing air currents centered in middle latitudes near 6 to 12 km. The jetstreams are frequently collocated with wintertime storms.)
Abrupt climate change due to variations in the atmospheric circulation and its attendant patterns of climate variability can arise through two principal mechanisms: (1) through abrupt changes in the time-dependent behavior of the circulation; or (2) through slowly evolving changes in the circulation that project onto large horizontal gradients in surface weather. For example, a relatively slow shift in the distribution of precipitation could give rise to relatively rapid changes in precipitation patterns in regions that lie at the interface of dry and rainy regions (see Figure 2.8), potentially altering a location’s local climate with possible ramifications to water supplies and/or agriculture for example. The text below discusses the evidence for: (1) abrupt changes in the circulation and (2) steady changes in the circulation that may, in turn, trigger relatively abrupt changes in climate in regions of large spatial gradients in surface weather.
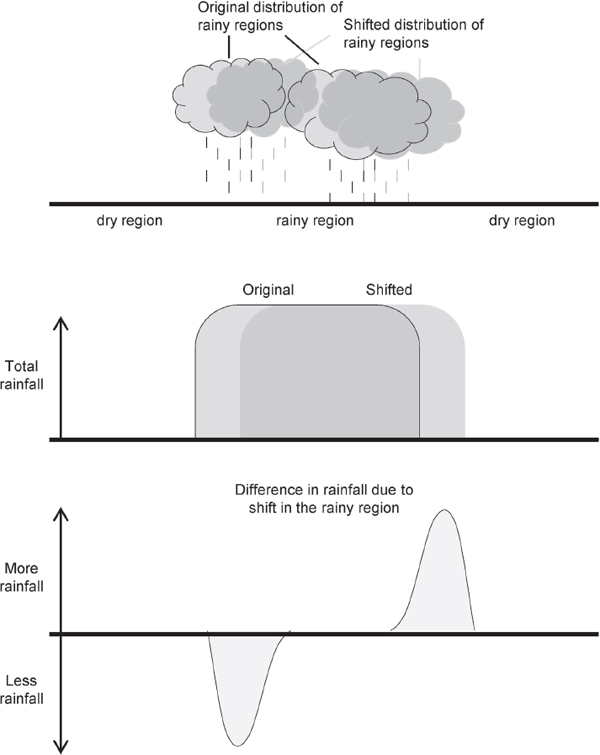
FIGURE 2.8 In the above example, the largest changes in rainfall due to the shift in the circulation are found on the flanks of the original precipitation regions. A slowly evolving change in the circulation may thus lead to seemingly abrupt changes in precipitation in regions where the existing spatial gradients in rainfall are largest.
Abrupt Changes in the Time-Dependent Behavior of the Atmospheric Circulation
Given the definition of abrupt change in this report (see Box 1.2), there is little evidence that the atmospheric circulation and its attendant large-scale patterns of variability have exhibited abrupt change, at least in the observations. The atmospheric circulation exhibits marked natural variability across a range of timescales, and this variability can readily mask the effects of climate change (e.g., Deser et al., 2012a, 2012b). As noted above, patterns of large-scale variability in the extratropical atmospheric wind field exhibit variations on timescales from weeks to decades (Hartmann and Lo, 1998; Feldstein, 2000). The time series of large-scale tropical climate variability—such as the MJO and ENSO—exhibit more quasi-periodic behavior (e.g., Rasmusson and Carpenter, 1982; Zhang, 2005). In both the tropics and extratropics, it is difficult to discern significant long-term trends in the patterns of climate variability from natural variability, never mind abrupt (threshold) changes in the atmospheric circulation.
The most widely studied and arguably most robust “regime shift” in the modern historical record (i.e., over the past century) is the relatively rapid change in North Pacific sea-surface temperatures circa 1976, referred to alternatively as ENSO-like decadal variability (Zhang et al., 1997) and the Pacific Decadal Oscillation (PDO; Mantua et al., 1997). Numerous physical mechanisms have been proposed to explain the apparent step-like jump in the extratropical atmosphere/ocean system. But it is unclear whether or not the 1976 regime shift in North Pacific climate reflects an abrupt change in the extratropical atmosphere-ocean system or simply the random superposition of different climate signals, e.g., similar regime-shifts are reproducible in simple stochastic models forced by atmospheric noise and ENSO (Newman et al., 2003).
One recent modeling study indicates that the atmospheric circulation may respond abruptly to future anthropogenic climate change, at least in a simple climate model. Wang et al. (2012b) force the dynamical core of an atmospheric general circulation model with warming in the tropical troposphere that mimics the effects of climate change there. Warmings up to ~5°C lead to steady changes in the atmospheric circulation consistent with those found in full IPCC-class simulations. When the warming is increased beyond 5°C (which is predicted to occur by the end of the 21st century in the IPCC A1B scenario [IPCC, 2007c]), the atmospheric circulation exhibits large and abrupt changes, including a sudden poleward jump in the middle latitude jetstream of roughly 10 degrees latitude. As of this writing, the result is derived from a full primitive equation model, albeit one with very idealized physics. The regime-like behavior found by Wang et al. (2012b) has not been reproduced in a full physics, IPCC-class model simulation.
Steady Changes in the Time-Dependent Behavior of the Atmospheric Circulation
Relatively abrupt changes in the climate of a particular location may be driven not by abrupt changes in the atmospheric circulation, but rather by otherwise slowly evolving changes in the circulation in regions of large horizontal gradients in surface weather. Steady changes in the atmospheric circulation (i.e., changes that scale linearly with the forcing) have been documented in both climate models and observations. The most robust evidence for steady changes in the large-scale atmospheric circulation include:
- Observational and numerical evidence of a poleward shift in the Southern Hemisphere middle latitude jetstream (a positive trend in the Southern Annular mode) in response to Antarctic ozone depletion (Gillett and Thompson, 2003; Arblaster and Meehl, 2006; Son et al., 2010; Polvani et al., 2011; McLandress et al., 2011; Thompson et al., 2011). The signature of the ozone hole in surface climate is most pronounced during the summer season (Thompson et al., 2011). Opposite signed trends in the Southern Hemisphere middle latitude jetstream are expected in response to the recovery of the Antarctic ozone hole (Son et al., 2010; Arblaster et al., 2011; Polvani et al., 2011). The circulation response to ozone recovery is expected to oppose the response to future increases in greenhouse gases (see 2 below).
- Numerical evidence of a poleward shift in the Southern Hemisphere and North Atlantic middle latitude jetstreams in response to increasing greenhouse gases (e.g., Fyfe et al., 1999; Kushner et al., 2001; Cai et al., 2003; Yin, 2005; Miller et al., 2006; Meehl et al., 2007b; Barnes and Polvani, 2013). The changes in the flow project strongly onto the Southern annular mode and North Atlantic Oscillation (NAO), respectively. The poleward shift of the Southern Hemisphere middle latitude jetstream in response to increasing carbon dioxide is one of the most robust circulation responses found in climate change experiments, and is predicted to occur during all seasons (IPCC, 2007c). The predicted changes in the Northern Hemisphere circulation are generally much less robust. This is particularly true for the North Pacific (Barnes and Polvani, 2013). Trends in the Northern Hemisphere atmospheric circulation generally do not occur in numerical models until the latter half of the 21st century. The evidence for changes in the circulation in response to increasing greenhouse gases derives primarily from numerical climate model experiments. Observed trends in the middle latitude jetstreams and annular modes are not robust across all months (IPCC, 2007c).
- Observational evidence and evidence from numerical models for changes in the northern and southern boundaries of the tropics (Fu et al., 2006; Previdi and
Liepert, 2007; Seidel et al., 2008; Lu et al., 2009; Lu et al., 2007; Allen et al., 2012). The observational evidence is on the margins of statistical significance (Davis and Rosenlof, 2012).
- Numerical evidence for an acceleration of the Brewer-Dobson circulation in response to increasing greenhouse gases (e.g., Butchart et al., 2010; Butchart et al., 2006; Garcia and Randel, 2008; McLandress and Shepherd, 2009; Shepherd and McLandress, 2011; Garny et al., 2011). The Brewer-Dobson circulation is a large-scale circulation cell with rising motion in the tropical stratosphere, poleward motion in the middle latitude stratosphere, and sinking motion in the polar stratosphere. It is the primary pathway for global-scale equator-pole transport in the stratosphere and has implications for ozone concentrations.
Long-term trends in ENSO are uncertain. Lau et al. (2008), Meehl and Teng (2007), and Power and Smith (2007) all argue that future climate change could lead to changes in the amplitude and/or structure of the meteorological anomalies associated with ENSO. But there is little consensus on the response of ENSO itself to climate change and no clear century-scale changes in observed character of ENSO (e.g., Collins et al., 2010; IPCC, 2013; Lenton et al., 2008; Vecchi and Wittenberg, 2010). Observations indicate changes in equatorial Pacific surface pressure (e.g., Bunge and Clarke, 2009; DiNezio et al., 2013; Karnauskas et al., 2009; L’Heureux et al., 2013; Power and Smith, 2007; Vecchi et al., 2006). But the observed trends vary notably depending on the time period being explored, and it is unclear to what extent they reflect internal variability or anthropogenic forcing (e.g., IPCC, 2013; Seager and Naik, 2012).
It is unclear to what extent the aforementioned steady changes in the circulation have given or will give rise to rapid changes in climate in regions marked by large spatial gradients in surface weather (e.g., regions that lie at the interface of dry and raining regions). Additionally, circulation trends that are robust on large spatial scales may be much more difficult to detect on regional spatial scales due to the competing effects of internal climate variability (e.g., Deser et al., 2012a, 2012b).
Summary and the Way Forward
It is difficult to detect steady trends in the atmospheric circulation, particularly on regional scales where the trends are superposed on marked internal variability. It is also difficult to detect statistically robust abrupt changes in the circulation. Detection of an abrupt climate change requires demonstrating that the system was stationary before and after the change occurred. Furthermore, a seemingly robust abrupt climate shift can readily arise due to the chance superposition of internal and forced climate
change. The steady changes in the circulation noted in this section are generally not abrupt, but rather scale with the timescale of the forcing (with the notable exception of the aforementioned Wang et al., 2012b study).
Nevertheless, even relatively steady changes in the atmospheric circulation may prove important for understanding past and future abrupt climate change if such changes are coincident with large horizontal gradients in surface climate. Modest and slowly evolving changes in the width of the Hadley Cell could force rapid changes in precipitation in transition regions that lie between the subtropical deserts and tropical rainforests (e.g., the Sahel). Similarly slowly evolving changes in the middle latitude jetstreams could potentially lead to marked changes in surface temperature and precipitation in regions that lie on the flanks of the storm tracks, such as southern Australia. But again, such changes in the circulation will be difficult to detect in the presence of internal climate variability, particularly on regional scales in the extratropics (Deser et al., 2012a, 2012b; Wallace et al., 2013). The ability of steady changes in the circulation to drive abrupt changes in surface climate has not been widely investigated and is a key topic for future research (Box 2.3).
BOX 2.3 COUPLING OF ATMOSPHERIC AND LAND SURFACE AS A CURRENT RESEARCH FRONTIER
The coupling between land surface vegetation and atmosphere could also potentially cause abrupt changes of atmospheric circulation at regional scales. For example, coupled atmospherevegetation models suggest that gradual changes in Earth’s orbit may lead to the collapse of the Sahara green vegetation and climatic drying (e.g., Bathiany et al., 2012; Claussen et al., 1999; Zeng and Neelin, 2000), although other mechanisms could also be responsible for the latter (e.g., Liu et al., 2006) and in general dynamic vegetation models are in the early stages of development. In a regional model experiment coupled with a simple coupled atmospheric-vegetation model, an abrupt northward jump of the West Africa monsoon circulation can result; when the regional model is initialized with the vegetation/desert border at about 21°N, the low-level westward jetstream over northern Africa and rainfall shift northward and lead to a vegetated central Sahara (Wang and Eltahir, 2000; Patricola and Cook, 2008). However, when the model is initialized by relatively small deviations of the vegetation/desert border from its location today (~10°N), the vegetation distribution tends to remain similar to that found today. Whether such abrupt changes can be reproduced by coupling an atmospheric model with a more comprehensive dynamic vegetation model remains to be seen.
The time-dependent behavior of the atmospheric circulation is generally well monitored by the current observing network of surface stations and radiosondes in the Northern Hemisphere. Circulation measurements in the tropics and Southern Hemisphere are less widespread, and the tropics in particular suffers from a lack of longterm in-situ observations of atmospheric temperatures and pressure. Maintaining and enhancing the current observational network of remotely sensed and in-situ measurements that can be used to infer changes in the atmospheric circulation is essential.
The likelihood of abrupt changes in the atmospheric circulation remains unclear, as does the potential for inducing abrupt climate change in regions of large gradients in surface weather. As such, understanding abrupt changes in—and due to—the atmospheric circulation remains a key topic for future research. Additional investigative work by individual scientists is required in a range of research areas. Interdisciplinary research is needed to assess the importance of changes in the circulation for regions of particular “vulnerability,” e.g., in terms of food-security or ecosystems habitat. Model studies are required to assess the mechanisms that drive trends in the circulation, and their amplitudes relative to internal climate variability. Observational studies are required to assess and monitor changes in the observed circulation.
Weather and Climate Extremes
Extreme weather and climate events include heat waves, droughts, floods, hurricanes, blizzards, and other events that occur rarely. In some cases, statistical probability is used to define these extremes, for example, heavy rainfall events or extremely hot or cold temperatures with a 1, 5, or 10 percent occurrence probability. The IPCC SREX report (Seneviratne et al., 2012) defines them as having a 5 percent or 1 percent or even lower chance of occurrence during the same period, and other examples include 10 percent coldest nights, 10 percent warmest daily maximum temperature, 5 percent heaviest rain rate, etc. In other cases, weather and climate extremes are defined by exceeding a threshold that typically results in significant impacts or costs. For example, hurricanes and typhoons defined by wind speeds exceeding specific thresholds are always considered extreme events.
Conditions considered extreme in one location (for example warm temperatures in Barrow, Alaska) may not be extreme in other locations (for example in Miami, Florida). Also, what is normal in one season, a snowfall of three feet in January in New Hampshire, for example, may be extreme at the same location but in another season. Other considerations include the impact of compounding events; for example, some climate extremes, such as droughts or floods, may be the result of an accumulation of moder-
ate weather or climate events. The individual events may not extreme, but the accumulation of them over a relatively short period of time leads to an extreme event, in which case the closely-spaced accumulation of the events is the extreme.
Extreme weather and climate events are among the most deadly and costly natural disasters. For example, tropical cyclone Bhola in 1970 caused about 300,000-500,000 deaths in East Pakistan (Bangladesh today) and West Bengal of India.3,4 Hurricane Katrina caused more than 1,800 deaths and $96-$125 billion in damages to the Southeast U.S. in 2005. Worldwide, more than 115 million people are affected and more than 9,000 people are killed annually by floods, most of them in Asia (Figure 2.9 or see, for example, the Emergency Events Database5). Heat waves contributed to more than 70,000 deaths in Europe in 2003 (e.g., Robine et al., 2008) and more than 730 deaths and thousands of hospitalizations in Chicago in 1995 (Chicago Tribune, July 31, 1995; Centers for Disease Control and Prevention, 1995). Heat waves are one of the largest weather-related sources of mortality in the United States annually.6
According to data collected by the National Climate Data Center, there were 134 weather or climate disaster events with losses exceeding $1 billion each in the United States between 1980 and 2011, an average of more than four per year (Table 2.1). Floods, droughts and wildfires—events that appear to be changing in frequency and severity due to climate change—make up about a third of these and slightly more than a third of the dollar damages (adjusted to 2012 dollars). Droughts are particularly costly, comprising about 12 percent of the events by number, but about double that (23.8 percent) by total cost.
Climate Change Is Affecting Extremes
Climate change is expected to shift frequency statistics for weather and climate events, as illustrated in Figure 2.10, in ways that affect the likelihood of extreme events on the tails of the distribution, either the high side (“extremely hot” for example) or the low side (“extremely cold”). Such shifts are already being observed. For example, Hansen et al. (2012) studied temperature anomalies over the past 6 decades and found that while anomalies greater than three standard deviations occurred over about 0.3 percent of the land area in their base period (1951-1980), they now
____________________
3http://www.wunderground.com/hurricane/deadlyworld.asp.
4http://www.nbcnews.com/id/24488385/ns/technology_and_science-science/t/deadliest-stormshistory/#.UX1b2KX2Wqx.
FIGURE 2.9 The projected increase in the number of people (in thousands) exposed to floods in 2030 compared to those in 1970. Only catchments bigger than 1,000k m2 were included in analysis; therefore, only the largest islands in the Caribbean are covered. Source: IPCC, 2012; Solterra Solutions, 2012.
TABLE 2.1 Billion-dollar weather and climate disasters in the United States from 1980 to 2011 by type. Total damages are in consumer-price-index-adjusted 2012 dollars. Note that the impacts of droughts are difficult to determine precisely, so those figures may be underestimated.
| Disaster Type |
# Events |
% Frequency |
CPI-adjusted Damages (billions of dollars) |
% Damage |
| Severe storm | 44 | 32.8 | 96.1 | 10.9 |
| Tropical cyclone | 31 | 23.1 | 417.9 | 47.4 |
| Flooding | 16 | 11.9 | 85.1 | 9.7 |
| Drought | 16 | 11.9 | 210.1 | 23.8 |
| Wildfire | 11 | 8.2 | 22.2 | 2.5 |
| Winter storm | 10 | 7.5 | 29.3 | 3.3 |
| Freeze | 6 | 4.5 | 20.5 | 2.3 |
| Total | 134 | 100% | 881.2 | 100% |
Source: Solterra Solutions, 2012.
(2006–2011) occur on 6-17 percent of the land (Figure 2.11; see also the comment on Hansen et al. [Rhines and Huybers, 2013], and Hansen et al.’s response [Hansen et al., 2013b]). A similar change has been observed in rainfall. While total precipitation in the United States increased by about 7 percent over the past century, the heaviest 1 percent of rain events increased by nearly 20 percent (Bull et al., 2007).
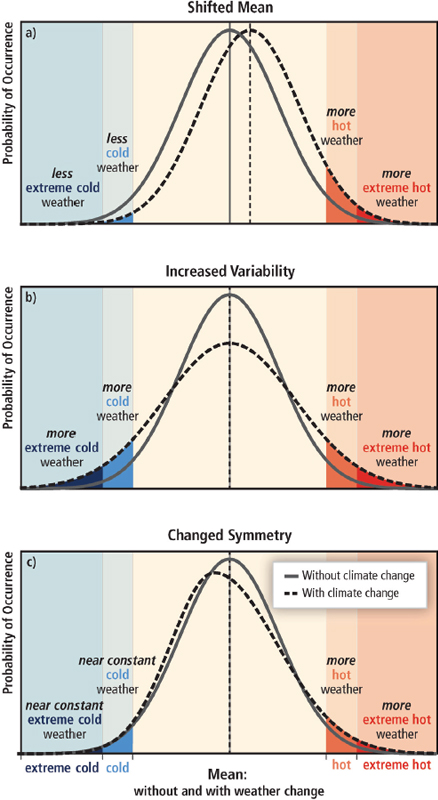
FIGURE 2.10 Potential effects of changes in temperature distribution on extremes: a) effects of a simple shift of the entire distribution toward a warmer climate; b) effects of an increased temperature variability with no shift of the mean; and c) effects of an altered shape of the distribution, in this example an increased asymmetry toward the hotter part of the distribution. SOURCE: Lavell et al., 2012.
FIGURE 2.11 Area of the world (in percent) covered by temperature anomalies (from Figure 5 in Hansen et al., 2012) in categories defined as hot (+0.43 σ), very hot (+2 σ), and extremely hot (+3 σ), with analogous divisions for cold anomalies. Dashed horizontal lines indicate areas of 33 percent and 67 percent, corresponding to climate dice with two and four sides colored red, respectively. Note: NH=Northern Hemisphere; SH=Southern Hemisphere. Source Hansen et al., 2012.
Climate change may also be affecting other weather and climate extremes, with impacts and trends that vary regionally. The Mediterranean and West Africa are likely experiencing more frequent and severe droughts, while the opposite is the case for central North America and northwest Australia (IPCC, 2013). Longer, hotter, and drier summers have contributed to an increase in the frequency and severity of wildfire in
the western United States (Westerling et al., 2006), a trend that is expected to continue as the climate warms (NRC, 2011a). The possibility of climate change leading to more intense tropical cyclone activity, particularly in the North Atlantic, continues to receive significant research attention.
Links Between Extreme Events and Abrupt Change
While extreme events per se are not abrupt climate changes as defined in this report, changes in extreme events could lead to abrupt changes in two ways: (1) an abrupt change in a weather or climate extremes regime, for example a sudden shift to persistent drought conditions; or (2) a gradual trend in the frequency or severity of extremes that causes abrupt impacts when societal or ecological thresholds are crossed, as illustrated in Figure 2.10. The potential for abrupt regime shifts was raised in NRC (2002), which highlighted the transitions into and out of the 1930s Dust Bowl as prime examples. Since NRC, 2002 was published, the potential for abrupt impacts associated with gradual changes in extreme events—such as abrupt changes in terrestrial ecosystems due to droughts and storms—has been studied extensively (e.g., Hutyra et al., 2005; Saatchi et al., 2013). Moreover, the vulnerabilities of the complex and globally connected socio-economic system to such extreme events have become more widely recognized (Mileti, 1999; NRC, 2010a; NRC, 2012c) and the impacts of extreme events on societal tipping points have been more clearly appreciated (Lenton et al., 2008; Nel and Righarts, 2008).
Most extreme events are temporary and their impacts are reversible. For example, the US Great Plains recovered from the severe drought in the 1930s when rainfall returned to normal and land management practices were improved and maintained. However, even temporary extreme climatic events can trigger abrupt and irreversible changes when their impacts exceed the threshold or resilience of the ecosystems. For example, the 1950s drought in New Mexico abruptly shifted the ecotone between semiarid ponderosa pine forest and piñon–juniper woodland (Allen and Breshears, 1998).
In addition to a changing climate causing changes in extreme events, extreme events themselves can accelerate abrupt changes in other parts of the climate and Earth system. For example, extreme transient sea-level rise due to tropical or extratropical storm surge can cause abrupt increases of flood risk (Nicholls et al., 2007), putting many coastal regions at risk for loss of human life before gradual sea-level rise inundates the region. Extreme warm temperatures in summer can greatly increase the risks of mega-fires in temperate forests, boreal forests, and savanna ecosystems, leading to abrupt changes in species dominance and vegetation type, regional water yield and
quality, and carbon emission (e.g., Adams, 2013), before the gradual increase of surface temperature crosses the threshold for abrupt ecosystem collapse (more discussion in the section on Ecosystem Collapse and Rapid State Change below).
Extreme events could lead to a tipping point in regional politics or social stability. In Africa, extreme droughts and high temperatures have been linked to an increase of risk of civil conflict and large-scale humanitarian crisis in Africa (Burke et al., 2009; Hsiang et al., 2011, 2013; Miguel et al., 2004; O’Loughlin et al., 2012; Zhang et al., 2011). Generally, extreme climate events alone do not cause conflict. However, they may act as an accelerant of instability or conflict, placing a burden to respond on civilian institutions and militaries around the world (NRC, 2012b). For example, the devastating tropical cyclone Bhola in 1970 heightened the dissatisfaction with the ruling government and strengthened the Bangladesh separatist movement. This led eventually to civil war and independence of Bangladesh in 1971 (Kolmannskog, 2008; NRC, 2012b). Historically, extreme climate events such as decadal mega-droughts may have triggered the collapse of civilizations, such as the Maya (Hodell et al., 1995; Kennett et al., 2012) or large scale civil unrest that ended the Ming dynasty (Shen et al., 2007). More extensive review of the extreme climate impacts is provided by the recent IPCC-SREX report (Field et al., 2012) and the 2012 NRC report on Climate and Social Stress (NRC, 2012b).
Summary and the Way Forward
The connection between extreme climate and related abrupt climate change is poorly understood, given the relatively poor understanding of both extreme climate events and abrupt changes. A number of reasons exist for this. First, because extreme climate phenomena represent rare events and modern climate records made by instruments are short, the modern record may capture only a few instances of these extreme events. Second, the statistical tools to which most climate researchers are accustomed are not applicable to this highly non-linear problem. Third, lack of quantitative understanding of the thresholds that trigger abrupt changes and causes of extreme climate events has limited our ability to provide process-based assessments of the risk of abrupt changes. Extreme events and the resultant abrupt changes are more likely unpredictable based on statistical models (Ditlevsen and Johnsen, 2010; Hastings and Wysham, 2010). Yet, it is prudent to assess the societal vulnerability and develop noregret mitigation policies for high-impact extreme events related to abrupt changes (NRC, 2012b). In this case, risk assessment based on a fundamental understanding of the climate dynamics may become a major tool for developing scenarios for stress
tests for the global and regional responding systems regarding their ability to manage potentially disruptive extreme and abrupt climate changes.
Coupled global climate models, such as those that participated in the Paleoclimate Modeling Intercomparison Project (PMIP) (Jansen et al., 2007), in combination with improved paleo-climate records have led to better appreciation of the extent of extreme events that have occurred in the past (e.g., Cook et al., 2010b). Finally, possible early warning for some of the abrupt climatic changes has begun to be explored. However, the understanding of the connections between climate change, climate and weather extremes, and abrupt change is still limited (e.g., Seneviratne et al., 2012). This report uses examples provided in the literature to illustrate the potential connections between extreme climate events and abrupt climate change, and highlights the need for a focused research effort to explore these climate events with high societal consequences but low probability.
In recent years, researchers, mainly in Europe, have begun to explore the feasibility of detecting early warning signs of abrupt climate changes. These studies have shown that an early warning signal may be detectable if an abrupt change is driven by gradual forcing and preceded by the critical slow down, increased variances, and skewness (e.g., Held and Kleinen, 2004; Livina and Lenton, 2007). However, extreme events are mainly a result of natural climate variability, making it hard to detect early warning signals within an otherwise noisy time series. The resultant abrupt changes are generally intrinsically unpredictable (e.g., Scheffer et al., 2009; Lenton, 2011). Given such a challenge, risk assessment would depend more on our predictive understanding and process-based probabilistic prediction than on statistical early warning signs of approaching a tipping point. This is central to the ability to improve the quantitative understanding of the thresholds that can trigger abrupt changes and the probability distribution changes of the extreme climate events with the slow varying climate states and forcings that can be monitored.
ABRUPT CHANGES AT HIGH LATITUDES
Potential Climate Surprises Due to High-Latitude Methane and Carbon Cycles
Interest in high-latitude methane and carbon cycles is motivated by the existence of very large stores of carbon (C), in potentially labile reservoirs of soil organic carbon in permafrost (frozen) soils and in methane-containing ices called methane hydrate or clathrate, especially offshore in ocean marginal sediments. Owing to their sheer size, these carbon stocks have potential to massively impact the Earth’s climate, should
they somehow be released to the atmosphere. An abrupt release of methane (CH4) is particularly worrisome as it is many times more potent as a greenhouse gas than carbon dioxide (CO2) over short time scales. Furthermore, methane is oxidized to CO2 in the atmosphere representing another CO2 pathway from the biosphere to the atmosphere in addition to direct release of CO2 from aerobic decomposition of carbon-rich soils.
Permafrost
Stocks Frozen northern soils contain enough carbon to drive a powerful carbon cycle feedback to a warming climate (Schuur et al., 2008). These stocks across large areas of Siberia comprise mainly yedoma (an ice-rich, loess-like deposit averaging ~25 m deep [Zimov et al., 2006b]), peatlands (i.e., histels and gelisols), and river delta deposits. Published estimates of permafrost soil carbon have tended to increase over time, as more field datasets are incorporated and deposits deeper than 1 m depth are considered. Estimates of the total soil-carbon stock in permafrost in the Arctic range from 1,700–1,850 Gt C (Gt C = gigatons of carbon; Tarnocai et al., 2009; Zimov et al., 2006a; McGuire et al., 2009). Figure 2.12 summarizes information on known stocks of highlatitude carbon.
To put the Arctic soil carbon reservoir into perspective, the carbon it contains exceeds current estimates of the total carbon content of all living vegetation on Earth (approximately 650 Gt C), the atmosphere (730 Gt C, up from ~360 Gt C during the last ice age and 560 Gt C prior to industrialization, Denman et al., 2007), proved reserves of recoverable conventional oil and coal (about 145 Gt C and 632 Gt C, respectively), and even approaches geological estimates of all fossil fuels contained within the Earth (~1,500 – 5,000 Gt C). It represents more than two and a half centuries of our current rate of carbon release through fossil fuel burning and the production of cement (nearly 9 Gt C per year, Friedlingstein et al., 2010).
These vast deposits exist largely because microbial breakdown of organic soil carbon is generally low in cold climates, and virtually halted when frozen in permafrost. Despite slow rates of plant growth in the Arctic and sub-Arctic latitudes, massive deposits of peat have accumulated there since the last glacial maximum (Smith et al., 2004; MacDonald et al., 2006).
Potential response to a warming climate Permafrost soils in the Arctic have been thawing for centuries, reflecting the rise of temperatures since the last glacial maximum (~21 kyr ago) and the Little Ice Age (1350-1750). However, this Holocene thawing
FIGURE 2.12 Top: Approximate inventories of carbon in various reservoirs (see text for references). Bottom: Provided for reference, estimated amount of carbon that would warm the planet approximately 2°C (Allen et al., 2009; uncertainty estimate in this value discussed in this reference) and estimated total amount of carbon to be released by the year 2100 under business-as-usual scenarios (IPCC, 2007c).
has accelerated in recent decades, and can be attributed to human-induced warming (Lemke et al., 2007). Under business-as-usual climate forcing scenarios, much of the upper permafrost is projected to thaw within a time scale of about a century (Camill, 2005, Lawrence and Slater, 2005). Exactly how this will proceed is uncertain. The rate of carbon degradation increases nonlinearly with temperatures above the freezing point of water. Furthermore, the spatial pattern of this degradation is spatially heterogeneous owing to small-scale geomorphic processes such as thermokarsting and slumping from ice-wedge melting (Jorgenson et al., 2006).
Wildfires have been spreading into some permafrost regions as local climatic changes promote increasingly dry conditions. (Lynch and Wu, 2000; Mack et al., 2011; Schuur et al., 2013). Charcoal records cored from 14 lakes in the Alaska interior suggest that recent fires burning there are unprecedented over the past 10,000 years (Kelly et al., 2013). The interaction of boreal fires with overall climate forcing is complex, because carbon release from burned plant material and soil carbon is at least partially countered by increased albedo of the fire scar (Goetz et al., 2007; Randerson et al., 2006). If the fire is sufficiently severe and burns the surface organic layer, heat flow through the active layer increases into the permafrost, and the active layer can increase to a thickness that does not completely refreeze the following winter. This results in formation of a talik, an unfrozen layer below the seasonally frozen soil and above the permafrost (Yoshikawa et al., 2002). Little is known about the potential of such burning to thaw and release stored carbon faster than would occur solely from broader, landscapescale decomposition, but the magnitude of carbon loss due to fire is significant and potentially offsets the increased carbon sequestration through Arctic greening (Mack et al., 2011). In sum, this known mechanism for rapid, unexpected carbon release demands further research.
The chemical fate of the decomposing carbon (i.e., methane versus CO2 emission) depends primarily on the availability of oxygen, which is controlled in these settings by how wet the soil is. Dry, well-aerated soils oxidize the carbon to produce CO2. Wet soils tend to be anoxic, leaving anaerobic fermentation as the degradation pathway. The maximum methane yield fraction is about 50 percent. However, methane can be oxidized to CO2 in the soil column (Reeburgh, 2007), so the methane fraction of the net carbon emissions to the atmosphere can be, and usually is, much lower than this.
Projecting the future water balance and moisture state of Arctic soils—and thus the ratio of CO2 to CH4 production—contributes the largest uncertainty in forecasting methane emissions from Arctic land surfaces. Because present-day permafrost landscapes generally support a greater abundance of lakes and wetlands than do thawed landscapes (Smith et al., 2005, 2007), a complete disappearance of permafrost would suggest an ultimately drier land surface and thus reduced methane production. However, such a transformation would require centuries to millennia, and numerous studies have shown that the initial stages of permafrost degradation lead to paludification (lake formation) of the land surface and increased methane emissions (Skre et al., 2002). Furthermore carbon-rich peatlands, when thawed, retain large volumes of liquid water and may have surfaces even moister than their frozen peatlands (Smith et al., 2012).
Climate-induced permafrost thaw is amenable to numerical modeling, because good theoretical frameworks of how heat propagates from the air-ground interface into the subsurface have been developed. While such models lack adequate observational datasets of subsurface soil properties and/or geology, it is clear that the time scale for deep permafrost thaw is measured in centuries, not years. Furthermore, unlike methane hydrates (see below), the very large stocks of permafrost soil carbon (i.e., the 1,672 Gt C of Tarnocai et al., 2009) must first undergo anaerobic microbial fermentation to produce methane, itself a gradual decomposition process. There are no currently proposed mechanisms that could liberate a climatically significant amount of methane or CO2 from frozen permafrost soils within an abrupt time scale of a few years, and it appears gradual increases in carbon release from warming soils can be at least partially offset, owing to rising vegetation net primary productivity (Beilman et al., 2009). Over a time scale of decades, however, a possible self-sustaining decomposition of Yedoma could occur before the end of this century (Khvorostyanov et al., 2008a, 2008b, 2008c). A related idea is the possibility of rising soil temperatures triggering a “compost bomb instability” (Wieczorek et al., 2011)—possibly including combustion—and a prime example of a rate-dependent tipping point (Ashwin et al., 2012). Such possibilities would represent a rapid breakdown of the Arctic’s very large soil carbon stocks and warrant further research. Even absent an abrupt or catastrophic mobilization of CO2 or methane from permafrost carbon stocks, it is important to recognize that Arctic emissions of these critical greenhouse gases are projected to increase gradually for many decades to centuries, thus helping to drive the global climate system more quickly towards other abrupt thresholds examined in this report.
Methane Hydrates in the Ocean
Stocks Under conditions of high pressure, high methane concentration, and low temperature, water and methane can combine to form icy solids known as methane hydrates or clathrates in ocean sediments. The methane derives from biological or thermal degradation of organic matter originally deposited on the sea floor. Although the overall rate of methane production in ocean sediments is fairly slow, over millions of years, substantial reservoirs of methane hydrate have accumulated in the world’s ocean margins.
Throughout most of the world ocean, a water depth of about 700 m is required for hydrate stability. In the Arctic, due to colder-than-average water temperatures, only about 200 m of water depth is required, which increases the vulnerability of those methane hydrates to a warming Arctic Ocean. The Arctic is also a focus of concern because of the wide expanse of continental shelf (25 percent of the world’s total), much
of which is still frozen owing to its exposure to the frigid atmosphere during lowered sea levels of the last glacial maximum (see above).
The inventory of methane in ocean margin sediments is large but not well constrained, with a generally agreed upon range of 1,000-10,000 Gt C (Archer, 2007; Boswell, 2007; Boswell et al., 2012). One inventory places the total Arctic Ocean hydrates at about 1,600 Gt C by extrapolation of an estimate from Shakhova et al. (2010a) to the entire Arctic shelf region (Isaksen et al., 2011) (see Figure 2.12). The geothermal increase in temperature with depth in the sediment column restricts methane hydrate to within a few hundred meters thickness near the upper surface of the sediments (e.g., Davie and Buffett, 2001). Beneath this stability zone, a layer rich in methane bubbles is often seen in seismic reflection data, called a “bottom simulating reflector.” The areal extent of methane-rich sediments is fairly well known from seismic observations of this feature, but uncertainty in the concentration of methane in those sediments is very large, thus resulting in the large uncertainty in the global inventory of ocean-floor methane.
Potential response to a warming climate Climate change has the potential to impact ocean methane hydrate deposits through changes in ocean water temperature near the sea bed, or variations in pressure associated with changing sea level. Of the two, temperature changes are thought to be most important, both during the last deglaciation (Mienert et al., 2005) and also in the future. Warming bottom waters in deeper parts of the ocean, where surface sediment is much colder than freezing and the hydrate stability zone is relatively thick, would not thaw hydrates near the sediment surface, but downward heat diffusion into the sediment column would thin the stability zone from below, causing basal hydrates to decompose, releasing gaseous methane. The time scale for this mechanism of hydrate thawing is on the order of centuries to millennia, limited by the rate of anthropogenic heat diffusion into the deep ocean and sediment column. Even on the Siberian continental margin, where water temperatures are colder than the global average, and where the sediment column retains the cold imprint from its exposure to the atmosphere during the last glacial time 20,000 years ago, any methane hydrate must be buried under at least 200 m of water or sediment. Bottom waters at depths of 50 or 100 m might warm relatively quickly with a collapse in sea ice cover, but it would take centuries for that heat to diffuse through the 100-150 m of sediment column to the hydrate stability zone. Thus the release of 50 Gt C from the Siberian continental shelf in 10 years as postulated by Whiteman et al. (2013) is unlikely.
The proportion of this gas production that will reach the atmosphere as CH4 is likely to be small. To reach the atmosphere, the CH4 would have to avoid oxidization within
the sediment column (a chemical trap) and re-freezing within the stability zone shallower in the sediment column (a cold trap). However, the hydrate stability zone thickness decreases to zero near the top of its depth range in the ocean, and an increase in water column temperature there could eliminate the stability zone entirely, potentially providing an easier pathway for methane to reach the sea floor. Episodic and explosive escapes of gaseous methane from the sediment column have been documented by kilometer-scale “wipeout zones” in seismic images (Riedel et al., 2002), and pockmarks on the sea floor, called eruption craters (Hill et al., 2004). However, the processes responsible for these observations are too poorly understood to predict what fraction of deeper CH4 might be released through them.
Most of the methane gas that emerges from the sea floor dissolves in the water column and oxidizes to CO2 instead of reaching the atmosphere. Bubble plumes tend to dissolve on a height scale of tens of meters (Rehder et al., 2002; Kessler et al., 2011), although larger plumes, consisting of larger bubbles, do rise farther. However, even in the cold Arctic Ocean, methane hydrate is only stable below about 200 m water depth, making for an inefficient pathway to the atmosphere at best. The highest oceanic methane fluxes to the atmosphere in the Arctic are probably in the coastal zone, associated with erosion of coastal permafrost (Shakhova et al., 2010b). In this region (and also in terrestrial lakes) the methane flux to the atmosphere is strongly impacted by ice formation on the water surface (Walter et al., 2007), providing another mechanism for climate feedback (He et al., 2013).
Another, more abrupt way to transfer methane hydrate from the sediment column to the atmosphere is by way of a submarine landslide. Methane hydrate floats in seawater just as water ice floats, and it also has greater potential to reach the atmosphere than methane bubbles (Brewer et al., 2002). The largest known submarine landslide (called Storegga) occurred ~8000 years ago, as documented in sediment deposits off Norway (Mienert et al., 2005). The volume of sliding material multiplied by a reasonable hydrate fraction in the pore space yields a possible methane source of about 1 Gt C. The climatic impact of this quantity of methane would be comparable to that of a volcanic eruption (although warming rather than cooling). As such it would have a significant climate impact, but one that is likely to be smaller than that of the anthropogenic CO2 rise (Archer, 2007).
Over time scales of centuries and millennia, the ocean hydrate pool has the potential to be a significant amplifier of the anthropogenic fossil fuel carbon release. Because the chemistry of the ocean equilibrates with that of the atmosphere (on time scales of decades to centuries), methane oxidized to CO2 in the water column will eventually increase the atmospheric CO2 burden (Archer and Buffett, 2005). As with decomposing
permafrost soils, such release of carbon from the ocean hydrate pool would represent a change to the Earth’s climate system that is irreversible over centuries to millennia.
Modeling the response of ocean hydrates to climate change is in its infancy. The largest uncertainty is the concentration of methane hydrate, especially in the shallow sediment column near the sediment water interface. Coupled atmosphere-ocean climate models can be used to simulate the thermal response of the ocean water column to climate change with a moderate degree of uncertainty and the subsequent penetration of heat into the sediment column. The response of an assumed column inventory of hydrate to warming can be simulated (Lamarque, 2008; Reagan and Moridis, 2009; Reagan et al., 2011), but the results depend strongly on the assumed hydrate concentrations. Another approach is to “grow” the sediment column through geologic time to obtain an initial condition for a climate change perturbation scenario (Archer et al., 2012), but uncertainties in various model parameters, such as the methane production rate and the fate of bubbles in the sediment column, prevent a well-constrained model forecast of the methane hydrate response to climate warming.
In summary, the ocean methane hydrate pool has strong potential to amplify the human CO2 release from fossil fuel combustion over times scales of decades to centuries. While anthropogenic warming should accelerate the thawing of offshore permafrost via warming of Arctic Ocean shelf waters, this impact should be considered additive to a broader thawing trend that has been underway for thousands of years.
Impacts of Arctic Methane on Global Climate
Although attention is often focused on methane when considering a potential Arctic carbon release, because methane is a short-lived gas in the atmosphere (CH4 oxidizes to CO2 within about a decade), ultimately a methane problem is a CO2 problem. It does matter how rapidly methane is released, and the impacts of a spike versus chronic emissions are discussed in Box 2.4. As methane emissions from permafrost degradation will also be accompanied by larger fluxes of CO2, Arctic carbon stores clearly have the potential to be a significant amplifier to the human release of carbon. The impact of Arctic carbon release on carbon policy thus should be considered. If anthropogenic carbon emissions are limited by law and/or economic means, yet CO2 and CH4 levels do not respond as expected because Arctic emissions are increasing, this could impact the willingness of countries to engage in limiting human carbon emissions.
Speculations about potential methane releases in the Arctic have ranged up to about 75 Gt C from the land (Isaksen et al., 2011) and 50 Gt C from the ocean (Shakhova et al., 2010a). A release of 50 Gt C methane from the Arctic to the atmosphere over 100 years
would increase Arctic CH4 emissions by about a factor of 25, and would make the present-day permafrost area about two times more productive of CH4 on average as comes from wetlands today. Postulating such a methane release over a more abrupt 10-year time scale, the emission rates from present-day permafrost would have to exceed that from wetlands by a seemingly implausible factor of 20, supporting a longer century timescale for this process, and making methane emission from polar regions an unlikely candidate for a tipping point in the climate system. Nonetheless, as can be seen in Box 2.4, releasing 50 Gt C of methane over 100 years would have a significant impact on Earth’s climate. The atmospheric CH4 concentration would roughly quadruple, with a resulting total radiative forcing from CH4 of about 3 Watts/m2. The magnitude of this forcing is comparable to that from doubling the atmospheric CO2 concentration, but the impact of the methane forcing would be strongly attenuated by its short duration (see Box 2.4).
The impact of the Arctic as a source of natural methane and CO2 can be monitored by measuring the concentrations of these gases in samples from around the world (in combination with models), as is being done by the NOAA Carbon Cycle Greenhouse Gas program7, although maintaining these networks in an era of budget cuts is an area of concern. As concluded above, an increase in Arctic CH4 emissions of more than a factor of 10 is required before it would begin to have a significant impact on Earth’s climate in the short term. Such a strong acceleration of methane degassing from the Arctic would result in measurably higher concentrations of methane in the high northern latitudes. Another avenue for monitoring is satellite measurements of column inventories of the gases, which provide much more detailed spatial coverage but no vertical resolution, in which air masses at different altitudes may carry gases that originated from different parts of the Earth’s surface. Measurements from aircraft, manned and unmanned, are the third potential monitoring approach, providing vertical resolution of the concentrations, which gives much tighter constraint on local-source fluxes.
Summary and the Way Forward
Arctic carbon stores are poised to play a significant amplifying role in the centurytimescale buildup of CO2 and methane in the atmosphere, but are unlikely to do so abruptly, on a time scale of one or a few decades. This conclusion is based on immature science, however, and a truly sparse monitoring capability. Marine hydrates are poorly mapped, and the possibility that they could even become a targeted fossilfuel resource for future economic development cannot be dismissed. Basic scientific research is required to assess the long-term stability of currently frozen Arctic and sub-
____________________
BOX 2.4 ARCTIC CARBON METHANE RELEASE: SPIKE OR CHRONIC?
The response of atmospheric concentration to a methane release depends on whether the release time scale is shorter or longer than the atmospheric lifetime of methane. An instantaneous release, for example, would cause the atmospheric methane concentration to spike immediately, then decay back toward the unperturbed value on a time scale of approximately one decade.
The climatic impact of a spike of methane would be shaped by the long time scale of the Earth’s temperature response to radiative (greenhouse gas) forcing, which in turn is set by the absorption of heat energy by the deep ocean on time scales of centuries to a millennium. The impact of the ocean uptake of heat would be to strongly attenuate the short-term climatic impact of such a spike in radiative forcing (see Figure). However, the ocean uptake of heat would also act to “bank” the heat, accumulating it through the spike period, prolonging the recovery of surface temperature beyond the demise of the methane spike itself. The spike therefore serves as a source of long-term ocean thermal pollution, which would be added to that from the anthropogenic atmospheric CO2 rise.
If, on the other hand, a methane release to the atmosphere continues for much longer than the methane lifetime, the concentration of methane in the atmosphere will rise to a new steadystate value. In general, the concentration of methane in the atmosphere is expected to scale roughly linearly with the global emission flux. The potential increase in chronic methane emissions from the Arctic must therefore be evaluated in the context of global methane emissions. These fluxes are compared in the Table below. Present-day methane emissions from the Arctic are much smaller than natural emissions, mostly from tropical wetlands, and human emissions (Denman et al., 2007). For this reason it would require a very large, prolonged relative increase in Arctic sources to significantly affect Earth’s climate.
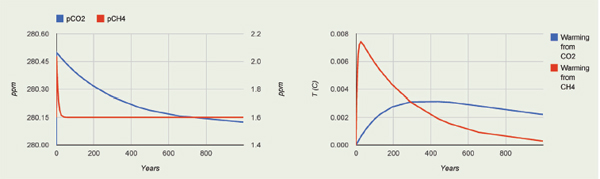
FIGURE Atmospheric chemistry and climatic impact of an abrupt 1 GtC methane release. (Left) methane concentration spikes after the release at the beginning of the simulation, then is oxidized to CO2. (Right) The temperature as driven by CH4 radiative forcing increases strongly during the methane spike, then subsides following the time scale of planetary (oceanic) cooling. The temperature change due to the oxidized CO2 grows but also subsides more slowly than that from CH4, due to the weaker greenhouse forcing but longer atmospheric lifetime of the CO2. From http://forecast.uchicago.edu/Projects/slugulator.html, based on data from Archer et al., 1997 and Schmidt and Shindell, 2003.
TABLE Summary of methane release scenarios compared with present-day methane fluxes and the radiative impact of business-as-usual CO2 rise.
| Scenario | CH4 emission rate, Gt C | Arctic Increase factor relative to today | Arctic CH4 flux/ Wetland flux, per m2 area | CH4 Conc. in Steady State | Radiative Forcing |
| Natural | 0.15 | ||||
| Anthropogenic | 0.25 | ||||
| Arctic lakes (Walter 2007) | 0.02 | ||||
| 50 Gt C over 10 years | 5 | 250 | 20 | 20 ppm | 5 Watts/m2 |
| 50 Gt C over 100 years | 0.5 | 25 | 2 | 6 ppm | 3 Watts/m2 |
| Business-as-usual CO2 in 2000 (500 Gt C released overall) | 1.5 Watts/m2 | ||||
| Business-as-usual CO2 in 2100 (~1500 Gt C) | 6 Watts/m2 | ||||
SOURCES: IPCC AR4 Ch 7, IPCC, 2000.
Arctic soil stocks, their future hydrologic status (i.e., moister or drier) that will largely drive their methane emissions, and the possibility of increasing methane gas bubble ebullition from currently frozen marine and terrestrial sediments as their temperatures rise.
If permafrost soils begin to release climatically significant amounts of methane, it should be detectable through monitoring atmospheric concentrations of methane using a network of monitoring stations around the world, but the current network is too sparse. Satellite observations of atmospheric chemistry would provide another means of detecting an Arctic methane feedback. Both types of sampling also provide constraint on sources and sinks of CO2, which play a significant part of the potential climate impact of the Arctic. It is therefore vital that the flask sample and satellite atmospheric chemistry monitoring efforts be continued and expanded.
While it is not possible to directly observe subsurface permafrost state from satellite remote sensing, surface freeze-thaw status is readily observed in radar scatterometer images, making this technology one of the best ways to infer frozen vs. thawed ground conditions over large, remote geographic areas. Below the ground surface, in-situ methods offer the most direct and effective way to monitor the state of permafrost health, through ongoing temperature measurements in boreholes. However, both the number and geographic extent of long-term borehole observing sites is small. Approximately 200 boreholes of varying depths have been identified for permafrost monitoring by International Permafrost Association (IPA) Global Terrestrial Network for Permafrost8 (GTN-P), which includes monitoring activities of the Geological Survey of Canada. The geographic coverage represented by these sites is greatest (by far) in Alaska, especially when shallow surface (<10m depth) boreholes are considered. Coverage is especially sparse in the continental interiors of Canada and Russia, and most notably so in the vast James Bay and West Siberian lowlands, as they contain very large stocks of frozen soil carbon in the form of peatland soils that have accumulated since the last glacial maximum.
A second key component for permafrost monitoring is measurements of active-layer depth (the thickness of seasonally thawed soil, measured downward from the soil surface). The dominant monitoring program in this respect is the IPA Circumpolar Active Layer Monitoring Network (CALM), which since its inception in 1991 has developed a network of more than two hundred monitoring sites in fifteen countries, mostly in the Arctic and sub-Arctic.9 Similar to the borehole monitoring sites (and indeed, often coincident with them), the geographic coverage of active-layer monitoring sites is
____________________
sparse, with greatest concentration in Alaska and a glaring absence of sites in carbonrich permafrost soils of interior Canada and Russia. Given that thawing permafrost also affects buildings, roads and other infrastructure, and thus society has a direct stake in its progression, there may also be an opportunity for citizen science in establishing additional permafrost monitoring sites.
Sea Ice
The Arctic Ocean has historically been largely covered in sea ice, which changes considerably with season and plays an integral role in the global climate system. Arctic sea ice has undergone rapid change since satellite records began in 1978. Significant decreases in sea ice have occurred during all months, but the most notable ice losses have occurred in summer. The linear trend in September sea ice from 1979-2012 was a loss of 13 percent per decade relative to the 1979-2000 mean (Fetterer et al., 2012; Stroeve et al., 2012a). The long-term decreases in summer sea ice are superposed on extreme record minima in 2007 and even less in 2012 (Stroeve et al., 2008),10 with the record low in Arctic sea ice extent on September 16, 2012 of only approximately 3.4 million square km.11 This September 2012 sea ice extent minimum was only 49 percent of its 1979-2000 mean. Thus, rapid Arctic sea ice loss is underway (Figures 2.13 and 2.14), and given the definitions used in this report, the changes already experienced qualify as an abrupt climate change. Projections from climate models suggest that ice loss will continue in the future, with a possibility of September ice-free conditions later this century (e.g., Stroeve et al., 2012b; Massonnet et al., 2012).
Scientific Understanding of Sea Ice Loss
Past climate models, as judged by the performance of the majority of Coupled Model Intercomparison Project 3 (CMIP3) simulations used in the IPCC Fourth Assessment Report, underestimated the observed linear trend in Arctic sea ice loss (Stroeve et al., 2007). The newer CMIP5 simulations that are being used in the upcoming IPCC Fifth Assessment Report are in better agreement with the observed sea ice loss (Stroeve et al., 2012a; Massonnet et al., 2012), but the reasons for the differences in sea ice trends between the CMIP3 and CMIP5 models remain unclear. They may result from more tuning of sea ice conditions within the models, improved model parameterizations and processes, or some combination of the two.
____________________
10 Also see http://nsidc.org/arcticseaicenews/2012/09/.
FIGURE 2.13 The time series of September Arctic sea ice extent from 1979-2013. SOURCE: National Snow and Ice Data Center, http://nsidc.org/arcticseaicenews.
A series of extremely low September sea ice conditions during the last decade, including the unprecedented declines in 2007 and 2012, suggests a recent acceleration in the long-term Arctic sea ice loss (e.g., Stroeve et al., 2012b). This bears resemblance to the so-called Rapid Ice Loss Events simulated in a number of climate models (Holland et al., 2006). These simulated events result when anthropogenic change is reinforced by natural variations. They appear to be triggered by increases in ocean heat transport from the North Atlantic to the Arctic and are amplified by the ice-albedo feedback. In the most dramatic of the simulated events, the September ice pack undergoes a 4 million square km loss (about 60 percent of the 1979-2000 ice cover) in only a decade, leading to near ice-free September conditions by 2040.
The rapid nature of observed and predicted changes in the Arctic suggests that Arctic sea ice could possibly undergo nonlinear threshold behavior as it retreats. Arctic climate change is strongly influenced by the surface albedo feedback, which acts to
FIGURE 2.14 Extent of Arctic sea ice in September 1979, 2000, 2007, and 2013. The magenta line shows the 1981 to 2010 median extent for September. The black cross indicates the geographic North Pole. Source: National Snow and Ice Data Center, http://nsidc.org/data/seaice_index/.
amplify warming as the reflective ice cover melts and exposes dark open ocean (see Figure 2.15). It has been proposed that this may lead to hysteresis in the Arctic sea ice (e.g., North, 1984), although other feedbacks may also play an important role (e.g., Abbot et al., 2011). To what extent such hysteresis has occurred and/or could occur in the Arctic climate system remains unclear. Some studies suggest a near-linear transition from perennial to seasonal sea ice cover to year-round ice-free conditions (Winton, 2006, 2008). Others suggest a bifurcation in the transition to a seasonally icefree Arctic (e.g., Abbot et al., 2011; Merryfield et al., 2008). And still others suggest a bifurcation in the transition to a year-round ice-free state (e.g., North, 1984, 1990; Ridley

FIGURE 2.15 Climate Feedback Loop: The melting of Arctic sea ice is an example of a positive feedback loop. As the ice melts, less sunlight is reflected back to space and more is absorbed into the dark ocean, causing further warming and further melting of ice. Source: NRC, 2010b.
et al., 2008; Winton, 2008). Eisenman (2012) discusses how differences in the strength of various climate feedbacks can lead to differences in the likelihood of hysteresis in the Arctic sea ice system.
The numerical evidence for irreversible change to a year-round ice-free state was first discussed in studies with simple diffusive climate models (e.g., North, 1984, 1990). In those studies, sea ice exhibits nonlinear behavior such that when it is reduced below a certain threshold (the “Small Ice Cap Instability” threshold), the model sea ice abruptly reverts to year-round ice-free conditions. The change in sea ice is largely irreversible, and substantial cooling is required to reestablish the original sea ice cover. Similar instability is found in numerous models that treat sea ice as a one-dimensional vertical column (Thorndike, 1992; Eisenman and Wettlaufer, 2009; Muller-Stoffels and Wackerbauer, 2011) and some but not all fully coupled IPCC-class climate models (e.g., Winton, 2006; Ridley et al., 2008). Based on these studies, it appears possible, but by no means certain, that a transition to year-round ice-free conditions could result from hysteresis.
Perhaps more relevant for the current Arctic climate is the possibility of a bifurcation to seasonally ice-free conditions. Although models generally simulate abrupt summer sea ice loss, the possibility that this might result from a lag in the system is mixed. Indeed, some simulated rapid ice loss events arise from the (random) superposition of large internal variability on the large greenhouse-gas forced trend (Holland et al., 2008). Some single-column model studies reveal bifurcation in the transition to seasonally ice-free conditions (Abbot et al., 2011; Merryfield et al., 2008; Flato and Brown, 1996) but others do not, due to the stabilizing effects of ice thickness on ice growth (Eisenman and Wettlaufer, 2009). A recent study argues that the bifurcation to a seasonally ice-free state in some single column models is an artifact of the model setup (Eisenman, 2012). In general, global climate model studies suggest that a bifurcation to seasonally ice-free conditions is unlikely (Tietsche et al., 2011; Ridley et al., 2008; Winton, 2006; Holland et al., 2008) and that sea ice loss is reversible if greenhouse gas concentrations decline (Armour et al., 2011; Ridley et al., 2012). Note, however, that this regrowth of sea ice would still require a return to the conditions under which sea ice existed; reversing sea ice loss requires reversing Arctic warming. Due to the long-timescales associated with the carbon cycle (e.g., Solomon et al., 2009), reversing sea ice loss would in practice require some type of active carbon removal from the atmosphere.
Regardless of whether hysteresis will occur, the transition to a nearly ice-free Arctic during summer is expected to happen rapidly with rising greenhouse gas forcing. Many methods have been used to predict the timing of near ice-free September conditions, here defined as an Arctic with less than 1 million km2 of ice extent for the
monthly average (see Overland and Wang, 2013 for a review). Extrapolating hindcast model-based historical sea ice volume trends results in September ice-free conditions prior to 2020 (e.g., Maslowski et al., 2012). However, this method does not account for the natural variability in the Arctic system that may be an important factor in trends over the recent past. Studies indicate that this natural variability is sizeable and can lead to a large range of trend magnitudes even on multi-decadal timescales (e.g., Wettstein and Deser, 2013; Kay et al., 2011). Simply extrapolating historical trends also does not account for feedbacks in the system, such as the negative ice thickness-ice growth rate feedback identified by Bitz and Roe (2004) that can slow the ice volume rate of loss. Other methods to predict the timing of September ice-free Arctic conditions have relied on projections from global climate models available through the Coupled Model Intercomparison Project (CMIP3 and CMIP5). A number of techniques have been employed to sub-set or recalibrate these projections based on different aspects of the observed ice cover, including the mean and/or seasonal cycle of ice extent (e.g., Stroeve et al., 2007, 2012a; Wang and Overland, 2009, 2012), historical ice cover trends (Boe et al., 2009), and ice volume and thin ice area (Massonnet et al., 2012). These different methods result in different timings for near-ice free September conditions within the Arctic, which range from the 2030s to 2100. Regardless of when icefree conditions are actually reached, it is clear that rapid Arctic sea ice loss is already underway and will continue for the foreseeable future.
The impacts of rapid decreases in Arctic sea ice are likely to be considerable (e.g., ACIA, 2005). Ice-free conditions during summer would have potentially large and irreversible effects on various components of the Arctic ecosystem (e.g., Arrigo et al., 2008; Bluhm and Gradinger, 2008; Andersen et al., 2009; Durner et al., 2009). They could have marked impacts on human society and economic development in the coastal polar regions (e.g., Jones et al., 2009; Huntington et al., 2012). Reductions in Arctic sea ice also have implications for Arctic shipping (Smith et al., 2012) and resource extraction (Prowse et al., 2009), which contribute to geopolitical concerns in the region. Additionally, ice-free Arctic summers would lead to large increases in the sensible and latent heat flux into the atmosphere during the fall season that may not only enhance large-scale high latitude terrestrial warming (e.g., Lawrence et al., 2008; Screen and Simmonds, 2010a, b) but also alter the large-scale atmospheric circulation and its variability (Alexander et al., 2004; Seierstad and Bader, 2009; Deser et al., 2010; Screen et al., 2012; Francis and Vavrus, 2012).
In contrast to the Arctic, the Antarctic has seen modest increases in sea ice. Climate models suggest that Antarctic sea ice will decline through the 21st century (e.g., Arzel et al., 2006). But very little work has been done on the stability characteristics of Southern Hemisphere sea ice. Note, however, that while relevant models have shown strong
agreement that the warming to date would have reduced Arctic sea ice, there is no such consensus for the Antarctic, with at least some models having simulated sea ice growth in response to moderate warming before switching to sea ice shrinkage from additional warming (e.g., Manabe et al., 1992; see also Liu and Curry, 2010).
Summary and the Way Forward
Arctic sea ice is already changing abruptly with numerous implications for ecosystems, the climate system, and socio-economic impacts. With continued warming, Arctic sea ice will continue to decline. There is considerably less consensus on future changes in Antarctic sea ice. For both poles, understanding future changes requires enhanced monitoring and research efforts.
Monitoring Studies suggest that rapid transitions in the Arctic sea ice are related to the ice thickness distribution (Holland et al., 2006; Lindsay et al., 2009). When the ice becomes thin enough, rapid transitions are likely as large areas of the ice pack can be effectively melted out. As such, monitoring Arctic ice thickness may be useful for predicting rapid changes in sea ice. Satellite-based altimetry measurements, often supplemented by similar measurements from aircraft, hold promise for obtaining basin-scale ice thickness information (e.g., Laxon et al., 2003; Giles et al., 2008; Kwok et al., 2009) and currently provide about a decade-long record. Altimetric freeboard measurements are converted into estimates of total ice thickness assuming isostatic balance. However, there are considerable uncertainties in these estimates due to limited information on the snow conditions on top of the sea ice, the ice density structure, and the high spatial variations in the ice pack. Upward-looking sonar measurements also provide estimates of ice thickness, but have limited spatial and temporal sampling. It remains unknown what uncertainties in ice thickness measurements are “acceptable” to realize any potential predictability for rapid sea ice change.
In addition to sea ice measurements, information on Arctic Ocean conditions may provide insight on the potential for rapid sea ice loss. For example, Holland et al. (2006) related simulated rapid ice loss events to anomalous ocean heat transport into the Arctic from the North Atlantic. Only limited observations of Arctic Ocean conditions currently exist. More research is needed to inform what specific ocean observations in what locations will prove the most useful and potentially enhance our ability to predict rapid ice loss events.
Research It is likely that a rapid transition to seasonally ice-free conditions in the Arctic will occur within the 21st century. However, the repercussions of this for climate, ecosystems, and societal impacts are still uncertain. Additional research is needed in these areas. There is also limited research on the potential predictability of rapid ice loss events. Most studies on ice predictability have used a perfect-model approach, in which a climate model is used to predict conditions simulated by that model, and have focused on seasonal to interannual predictability (e.g., Holland et al., 2011; Blanchard-Wrigglesworth et al., 2011; Chevallier and Salas-Melia, 2012). A recent study has assessed longer-lived ice loss events (Tietsche et al., 2013) and found little predictability in the onset of these events but some predictability in the magnitude of the events in simulations initialized after their onset. However, additional studies using different models and experiment design are needed to determine the robustness of these results.
Provided that rapid losses in sea ice may be predictable, there is additional uncertainty regarding what is required in terms of an observational network and modeling system to predict such events. Observing-network design studies, focused on the issue of abrupt sea ice loss, can be used to inform future observing needs. A recent Arctic Observing Network Design and Implementation Task Force report (AON Design and Implementation Task Force, 2012) provides more details.
A possible transition to year-round ice free conditions is still a distinct possibility. While this would only occur in the more distant future with continued and considerable increases in greenhouse gas concentrations, it would likely have dramatic impacts on the climate and ecosystems. More work is needed to determine why different models exhibit different behavior in this regard.
Finally, very little work has been done on the Antarctic sea ice system in terms of possible abrupt change. This is an additional research need. In particular, a better understanding of mechanisms of Antarctic sea ice variability and change, relevant feedbacks, and ice-ocean-atmosphere interactions is needed. This should be informed by both measurements and modeling of the Antarctic system. Currently, climate models struggle to accurately simulate even the mean conditions of Antarctic sea ice (Turner et al., 2013), and the utility of these models as a tool to study Antarctic ice needs to be critically assessed. It is likely that improvements in models, informed by observations, are needed to better understand the sea ice response to climate forcing and the potential for abrupt change.
Many different biological responses to climate change have been documented, both as an ongoing response to climatic change underway now and in the paleontological record. Thousands of species have reacted to a changing climatic regime by altering their geographic range, abundance, phenology (seasonal patterns), phenotype, or genotype, or in some cases recorded in the fossil record, have become extinct (Barnosky, 1986, 2009; Barnosky et al., 2003; Blois and Hadly, 2009; Brook and Barnosky, 2012; Hadly et al., 2004; Harnik et al., 2012; Pandolfi et al., 2011; Parmesan, 2006; Parmesan and Yohe, 2003; Root et al., 2003; Walther et al., 2002; Moritz et al., 2008). The ubiquity of biological response to climate indicates that climate changes underway will cause existing ecosystems to change noticeably. There is a possibility that at the ecosystem level the climatically-triggered changes will include abrupt state changes within the next few decades. This is supported by a large body of empirical and theoretical work that demonstrates when ecosystems change states, whatever the ultimate driver of change, they tend to do so abruptly once particular thresholds are crossed (for instance, see Barnosky et al., 2012; Bascompte and Sole, 1996; Carpenter et al., 2011; Peters et al., 2009; Scheffer et al., 2009; Swift and Hannon, 2010; and references therein).
Such abrupt state changes are well-documented for ecosystems at many scales, and can be triggered by a variety of forcing factors—including pollution, resource extraction, deforestation, and other land use changes—with climate change being only one of them (Scheffer et al., 2009; Lenton et al., 2008; Barnosky et al., 2012). In some cases ecosystems are known to have changed from the “old” to the “new” state within decades. For instance, in southern New Mexico, a changing dynamic between wind, water, and animals caused grasslands to transform into less productive shrublands over a total of about 70 years, with the shift from predominantly grassland to predominantly shrubland bracketed between 1980 and 1990 (Peters et al., 2009). More broadly, grazing and fire suppression have contributed to historic transitions from semi-arid grassland to desert shrub vegetation regimes in many parts of the southwestern United States and Africa (e.g., Schlesinger et al., 1990; Holdo et al., 2009), and prescribed fires as a land management practice can lead to abrupt transition from a savanna-desert plant regime to a savanna-grassland regime (e.g., Taylor et al., 2012).
Climate change has been shown to be an important component of abrupt ecosystem state-changes. A particularly instructive example is the Sahel region of Africa, which switched from vegetated land that supported cattle to unproductive desert within 5 years beginning about 1965, causing widespread famine and an international crisis in the region that continues to be a problem today. In the Sahel the onset of desertification involved an interplay between the position of the West African Monsoon
(Lenton et al., 2008) and the local climate feedbacks that are controlled by the amount of vegetative cover (Stewart, 2010). In the case of the 1960s desertification, a few bad drought years were caused by warm sea surface temperatures, which weakened the influence of the South African monsoon over the Sahel. At the same time, grazing pressure denuded the landscape. This, in turn, regionally increased albedo (caused more sunlight to be reflected from the land) and reduced evaporation, which further weakened the monsoon (Stewart, 2010). As a result, some regions of the Sahel that formerly supported grazing still remain unproductive desert today. Other empirical data and theoretical models indicate that such interplays between land use and climate change are likely to cause desertification in many other dryland environments that currently support 2 billion people (D’Odorico et al., 2013; Schlesinger et al., 1990).
While much of the work on whole-ecosystem regime shifts has focused on drylands, a growing body of evidence indicates abrupt state shifts will plausibly affect many other ecosystems as climate continues to change over the next several decades. Boreal forests appear susceptible to rapid transition to sparse woodland or treeless landscapes as temperature and precipitation patterns shift (Scheffer et al., 2012b). Climatic shifts would be expected to exacerbate the large-scale ecosystem changes in boreal regions that human induced changes from grazing or fires can also trigger (e.g., Chapin et al., 2004, Randerson et al., 2006).
At the global scale, observations show that the transitions from forests to savanna and from savanna to grassland tend to be abrupt when annual rainfall ranges from 1,000 to 2,500 mm and from 750 to 1,500 mm, respectively (Hirota et al., 2011; Mayer and Khalyani, 2011; Staver et al., 2011). Such rainfall regimes cover nearly half of the global land, where either a gradual climate change across the ecosystem thresholds or a strong perturbation due to either extreme climate events, land use, or diseases could trigger abrupt ecosystem changes. The latter could in turn amplify the original climate change in the areas where land surface feedback is important to climate (e.g., Friedlingstein et al., 2006; Scheffer et al., 2006).
Amazon forests represent the world’s largest terrestrial biome and potentially the tropical ecosystem most vulnerable to abrupt change in response to future climate change in concert with agricultural development (e.g., Cox et al., 2000; Lenton et al., 2008; Zelazowski et al., 2011). Thus, the rest of this section explores the risk of collapse of the Amazon forests as an example of a potentially vulnerable ecosystem.
Abrupt Transformation of the Amazon Forest by Climate Change and Deforestation
The closed-canopy equatorial forests of Amazonia are iconic in public perception: lush, highly productive, richly diverse ecosystems. The forests are characterized by a tall canopy of broadleaved trees, 30-40m high, sometimes with impressive emergent trees up to 55 m or taller. The Brazilian portion of the Amazon comprises 4 × 106 km2,12 less than 1 percent of global land area, but disproportionally important in terms of aboveground terrestrial biomass (15 percent of global terrestrial photosynthesis [Field et al., 1998]) and number of species (~25 percent, Dirzo and Raven, 2003). Direct human intervention via deforestation represents an existential threat to this forest: despite recent moderation of rates of deforestation, the Amazon forest is on track to be 50 percent deforested within 30 years—arguably by itself an abrupt change of global importance (Fearnside, 1983; Gloor et al., 2012).
Climate change represents yet another source of stress on an already distressed system. In particular, seasonal and multi-year drought frequency and intensity may have increased, and such increase could in part be attributable to anthropogenic forcing (e.g., Dai, 2011; Li et al., 2008). The projected radiative-forced increase of extreme surface temperatures and stronger spring barrier for wet season onset (Cook et al., 2010a; Seth et al., 2011) would increase risk of forest fires (Golding and Betts, 2008), although how changes of ENSO, AMO, and aerosols loadings will influence future droughts remain unclear (e.g., Andreae et al., 2005).
Biophysical Mechanisms and Feedbacks Defining the Boundaries of the Closed-Canopy Equatorial Forest At the continental scale, nonlinear feedbacks between the equatorial forest and the atmosphere have been recognized for decades. These forests receive enormous inputs of radiant energy and moisture. A significant fraction (25-35 percent) of regional rainfall represents water recycled between the forest and the atmosphere (Salati et al., 1979; Eltahir and Bras, 1994; Zeng et al., 1996; da Rocha et al., 2009), providing a strong homeostatic mechanism, i.e., the forest can regenerate rainfall that waters itself. The local water recycling provides nearly 100 percent of the regional rainfall during dry season (Li and Fu, 2004). Thus, forest clearing would reduce dry season rainfall, increase fire risk, and possibly delay wet season onset (Gash and Nobre, 1997; Fu and Li, 2004, Costa and Pires, 2010). This, in turn, increases the ecosystem’s vulnerability to forest clearing via reduction in water recycling as forest cover is removed (Salati and Nobre, 1991), or via changes in rain formation processes (e.g.,
____________________
12 Approximately 40% of the total area of the United States.
FIGURE 2.16 Frequency of tropical forests and savannas, plotted vs. mean annual precipitation (Hirota et al., 2011).
Andreae et al., 2004) and monsoon circulation transition (Zhang et al., 2009, Bevan et al., 2009) due to inputs of smoke from agricultural burning.
Nonlinear feedbacks, thresholds, and bi-stable hysteresis13 operate at the ecosystem scale. Classic biogeographical studies have shown that shifts in the balance between potential evaporation and precipitation (“potential evaporation ratio”) can give rise to threshold behavior in these forests (Holdridge, 1947, 1964). With current mean temperatures in the range 26-29°C in Amazonia, the lower limit of rainfall to sustain a closed canopy forest is about 1,600 mm/yr (Nix, 1983; Hirota et al., 2011; see Figure 2.16). The length of the dry season and rainfall variability are also important, however. Tropical or subtropical areas with higher rainfall, even with seasonally flooded landscapes, typically have fire-adapted, low-stature ecosystems if the dry season is longer than ~5 months (e.g., Pantanal in Brazil, northern Australia).
Biophysical Mechanisms and Feedbacks of Nonlinear Feedbacks, Thresholds, and Hysteresis Trees that make up the main canopy, or emerge above it, access moisture
____________________
13 A bi-stable hysteresis refers to a system that can be in one of two stable states and which state it occupies depends on the history of the system. An example would be a landscape that has been stable in the past as either a rain forest or a dry savanna depending on the pervading climate conditions.
stored deep in the soil and use it to harvest the intense sunlight available in the dry season. At least in eastern Amazonia, where deep soil columns are accessible to roots (see, for example, Zeng et al., 1996; Kleidon and Heimann, 2000; Nobre and Borma, 2009), transpiration rates in tropical forests are sustained at peak levels throughout dry periods of four to five months. This persistence of very high latent heat flux is opposite to the seasonal trends of transpiration and evaporation in savannas, even with similar total rainfall (Saleska et al., 2003; da Rocha et al., 2009). The forest is resilient, with rapid growth of canopy trees. Proliferation of diverse flora and fauna is enabled by the moderated microclimate, made possible by conversion of solar energy to latent heat.
Two processes can disrupt this system and cause a transition to a less dense transitional forest or to fire-adapted vegetation: (1) damage to canopy trees by fires fueled by dry understory vegetation; or (2) depletion of deep stores of soil moisture. These processes can potentially lead to abrupt changes in Amazon forest structure and extent.
1. Fires—Natural fires are rare in closed-canopy forests, but human-set fires are common today, and may have been widespread in the past when indigenous populations were much higher than today (Roosevelt et al., 1996; Bush et al., 2000; Shepard et al., 2012). Fires that enter the closed-canopy forest from agricultural or forest clearing activities are normally of low intensity, but may kill large trees during droughts or strong dry seasons. Smoldering ground fires may also be lethal if the fires return too frequently (Cochrane et al., 1999; Numata et al., 2010). Hence the damage to canopy trees by fires depends on the factors that control intensities and return intervals: temperature, humidity, duration of the dry season, occurrence of dry season rainfall, sources of ignition, and fuel loads (Adams, 2013). Positive feedbacks operate for some time after ground fire, because the killing of undergrowth vegetation creates more dry fuel, higher temperatures, and lower humidity for the next fire (Cochrane et al., 1999). Pueyo et al. (2010) found that strong positive feedbacks between drought and deforestation caused the very large fires in Roraima, Brazil, in 1997.
Although apparently irreversible shifts in ecosystem composition may occur after fires, the underlying causes are often changes in dry season balances of energy and water (means), the frequency of extremely hot or dry periods (variances), edge effects at forest-agriculture boundaries, and the occurrence of sources of ignition (human associated, e.g., agriculture). Vulnerable forest may persist for extended periods until an “event” leads to an actual transition, which may appear to be irreversible.
2. Deep water depletion and variability of rainfall in vulnerable (nearthreshold) forests—Nepstad et al. (2007) carried out a manipulation experiment on a central Amazon forest by reducing rainfall in the wet season by 60 percent. Initially the forest was unaffected, but after the third year the largest trees began to die. Detailed simulations support Nepstad’s hypothesis that the key factor was incomplete recharge of deep soil moisture, which supports prodigious rates of photosynthesis and transpiration by the largest trees during the dry season (Figure 2.17; Ivanov et al., 2012). Hutyra et al. (2005) used the 100-year, monthly reconstruction of rainfall by New et al. (1999) to refine the Holdridge plot (Holdridge, 1947, 1964) and explore the role of
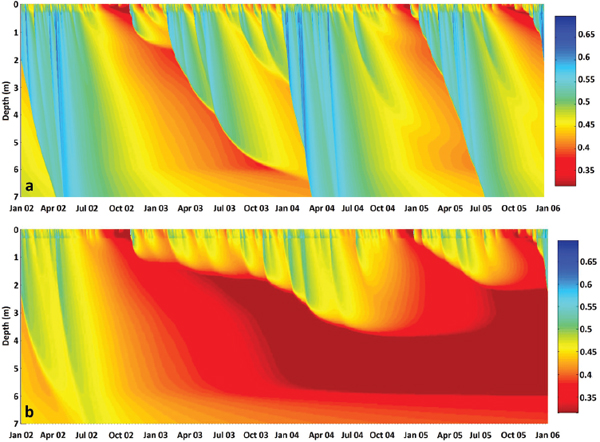
FIGURE 2.17 Colors show volumetric soil moisture content, where blue is more moist and red is drier. Soil moisture shown as a function of depth for two model simulations over a four-year period. (a). Soil moisture profile in a central Amazon forest, simulated using observed meteorology. Trees in the upper canopy had a higher fraction of deep roots, conversely for understory plants. (b) Simulation with 60 percent reduction in precipitation or increase in evaporation; upper canopy trees cannot survive. Source: Ivanov et al., 2012 (model) using data from Nepstad et al., 2007.
FIGURE 2.18 (left) Drought probability (soil moisture < 25 percent of capacity) for 100 years of CRU monthly climate reconstruction, based on evaporation parameterized from eddy flux data and plant available water capacity from biogeographical information. The boundary of the closed canopy forest with transitional forest or savanna (right) corresponds closely to 50 percent drought probability, consistent with the requirement to recharge deep soil reservoirs every 2 years as discussed by Nepstad et al. (2007).
Source: Hutyra et al., 2005.
variance in determining the stability of tropical forests, finding that the natural forestsavanna boundary corresponded to a 50 percent annual probability of incomplete recharge (Figure 2.18). Thus observed biogeographical boundaries give independent support for Nepstad’s hypothesis.
It appears that forests in the Amazon, at least in the central and eastern regions, may be rendered vulnerable to collapse either by increases of Potential Evaporation (PE, by increasing temperature or sunlight) or decreasing precipitation (Pc). In areas close to the biogeographic boundary, increasing variability of rainfall or longer dry seasons can shift forests to savannas, without changes in mean PE or Pc. In general, vulnerable forests may persist for extended periods until events, such as a series of strong droughts or repeated fire occurrences, lead to ecosystem collapse.
It is unclear how much hysteresis would attach to the re-establishment of tall trees in a closed canopy forest that had collapsed due to climatic shifts, since little is known about how this process proceeds. Amazonian forests appear to have expanded during moist periods of the Holocene and contracted in dry periods (Oliveira and Marquis, 2002), but in at least one case, the forest did not re-occupy its previous extent for some time after wetter conditions returned (Ledru et al., 1998). This “hysteresis” could have
been associated with any or all of a number of mechanisms, including persistent flammability, inhibited recruitment of big trees in locations where their early growth was not sheltered so that their roots could reach deep soil water, persistent shrub vegetation types, etc.
Summary and the Way Forward
Lenton et al. (2008) and Nobre and Borma (2009) have summarized current understanding of “tipping points” in Amazonian forests. Global and regional models do indeed simulate hysteresis and collapse of Amazonia forests. Models exhibit these shifts for a range of perturbations: temperature increases of 2-4°C, precipitation decreases by ~40 percent (1100 mm, according to Lenton et al., 2008), and/or deforestation that replaces large swathes of the forest with agriculture (e.g., Betts et al., 2004).
It is noted, however, that large-scale models do not have the detailed representation of subsurface hydrological processes that the Nepstad et al. (2007) data would seem require (see Ivanov et al., 2012). Thresholds may occur much closer to current conditions, for example, if precipitation falls below 1,600-1,700 mm (Nobre and Borma, 2009). Indeed, long-lasting damage to Amazonian forests may have occurred after the single severe drought in 2005 (Saatchi et al., 2013). In addition, the large-scale climate models have large errors in representing the processes that control rainfall variability and changes (e.g., Li et al., 2006; Yin et al., 2012). This is also a leading source of uncertainty in determining the risk of the Amazon ecosystem collapse (Friedlingstein et al., 2006; Good et al., 2013). A recent study shows that the dry season length over part of the Amazonia has increased much faster than that represented by climate models for both the current and future climate (Fu et al. 2013). Thus, the risk of rainforest collapse due to climatic drying is likely significantly higher than that represented by the current climate models. The committee concludes that credible possibilities of thresholds, hysteresis, indirect effects, and interactions amplifying deforestation, make abrupt (50 year) change plausible in this globally important system. Rather modest shifts in climate and/or land cover may be sufficient to initiate significant migration of the ecotone defining the limit of equatorial closed-canopy forests in Amazonia, potentially affecting large areas.
Monitoring for early warning The very strong 2005 drought gave rise to intensive studies of forest resilience and vulnerability using multi-spectral imaging from the MODIS satellite instrument (Marengo et al., 2008; Saleska et al., 2007), from the Quick-Scat active microwave sensor (Saatchi et al., 2013), and from distributed forest plots
(Phillips et al., 2009). The forest initially “greened up” during the dry period, as anticipated from seasonal changes under normal conditions (da Rocha et al., 2009), but subsequently experienced productivity decline and mortality (Phillips et al., 2009). Notably, the microwave data indicated that the forest did not fully recover for at least three years (Saatchi et al., 2013). The combined all-weather canopy surface temperature provided by passive microwave sensor, such as AMSR-E and hyperspectral data are also important for monitoring plants water stress for early warning.
Hence it appears that the tools required for monitoring and provision of early warning are at hand. Multi-spectral and active microwave data from satellites, plus an effective network of ecological plots, appear capable of monitoring response to climate change. Landsat combined with multi-spectral satellite sensing and LIDAR (e.g., Asner et al., 2010, 2012), can detect forest clearing and chart regrowth. It is not clear, however, that these tools will actually be available in the future to provide the required data at high spatial and temporal resolution with the necessary continuity. Commitment to carrying forward the satellite sensors is in doubt.
Long-term networks of ecological plots should be the foundation of global change studies of forest ecosystems. Current networks in the tropics (e.g., the Rainfor network14), represent diverse collections of scientists from many countries pursuing a range of questions, funded by a patchwork of sources. There is a very strong need for a “global service” network that makes comprehensive monitoring and early detection its main focus in the near future. This outcome would require a framework that does not now exist: a well-structured organization with long-term funding, broad international participation, and quality controlled data that enter the public domain. The lack of such an effort today undermines efforts to detect and respond to ecological changes in tropical systems, both forest and non-forest.
Extinctions: Marine and Terrestrial
Extinction is an irreversible biological change that can fundamentally alter the ecosystem of which a lost species was a part, contributing to ecological state shifts as described in the last section and to depleting ecosystem services as described below (see Chapter 3, Boxes 3.1 and 3.2). In the context of this report, extinction is recognized as “abrupt” in two respects. First, the numbers of individuals and populations that ultimately compose a species may fall below critical thresholds such that the likelihood for species survival becomes very low. This kind of abrupt change is often cryptic, in
____________________
that the species at face value remains alive for some time after the extinction threshold is crossed, but becomes in effect a “dead clade walking” (Jablonski, 2001). Such losses of individuals that take species towards critical viability thresholds can be very fast—within three decades or less, as already evidenced by many species now considered at risk of extinction due to causes other than climate change by the International Union for the Conservation of Nature.15 The second kind of abrupt change is simply the terminal event in the extinction process—the loss of the last individual of a species. While this is what most people recognize as extinction, it generally postdates by decades the dropping of numbers of individuals below species-viability thresholds. The abrupt impact of climate change on causing extinctions of key concern, therefore, is its potential to deplete population sizes below viable thresholds within just the next few decades, whether or not the last individual of a species actually dies.
The possibility that ongoing anthropogenic climate change will push many species past extinction thresholds is increasingly cited (Barnosky et al., 2011; Foden et al., 2013; Harnik et al., 2012; NRC, 2011a; Pimm, 2009; Cahill et al., 2012) and is rooted in considerations about both rate and amount of projected change. The rate of global climate change now underway is at least an order of magnitude faster than any warming event in the last 65 million years (Barnosky et al., 2003; Blois and Hadly, 2009; Blois et al., 2013; Diffenbaugh and Field, 2013).16 From the late 20th to the end of the 21st century, climate has been and is expected to continue changing faster than many living species, including humans and most other vertebrate animals, have experienced since they originated. Consequently, the predicted “velocity” of climate change—that is, how fast populations of a species would have to shift in geographic space in order to keep pace with the shift of the organisms’ current local climate envelope across the Earth’s surface—is also unprecedented (Diffenbaugh and Field, 2013; Loarie et al.,
____________________
15http://www.iucnredlist.org/.
16 In this context we refer to global-scale warming rates, not regional or local. At the global scale, glacialinterglacial transitions exhibit the most rapid and highest-magnitude warming rates documented in Earth history (Barnosky et al., 2003; Blois and Hadly, 2009; Diffenbaugh and Field, 2013). Note that regional proxies, such as the oxygen-isotope temperature reconstructions from the Greenland Ice Core Project that record Dansgaard-Oeschger events, often indicate faster regional rates of climate change than the overall global average for glacial-interglacial transitions, just as today warming is more pronounced in Arctic regions than in equatorial regions (Barnosky et al., 2003; Diffenbaugh and Field, 2013). Therefore in comparing rates of global warming today with past rates of global warming, it is essential to use global averages, rather than comparing a global average with a regional proxy. In so doing, Diffenbaugh and Field (2013) estimate that climate change now is proceeding at “at a rate that is at least an order of magnitude and potentially several orders of magnitude more rapid than the changes to which terrestrial ecosystems have been exposed during the past 65 million years.” Blois and Hadly (2009) and Barnosky et al. (2003) discuss how even when standardizing for higher rates that are apt to be a statistical artifact of measuring rates over shorter time intervals, the current global average rate of warming is above the global average that species have experienced over the past 65 million years.
2009). Moreover, the overall temperature of the planet is rapidly rising to levels higher than most living species have experienced (Figure 2.19). Consequently all the populations in some species, and many populations in others, will be exposed to local climatic conditions they have never experienced (so-called “novel climates”), or will see the climatic conditions that have been an integral part of their local habitats disappear (“disappearing climates”) (Williams et al., 2007). Models suggest that by the year 2100, novel and disappearing climates will affect up to a third and a half of Earth’s land surface, respectively (Williams et al., 2007), as well as a large percentage of the oceans (see, for example, the Ocean Acidification section of this report; NRC, 2011a; Ricke et al., 2013). Thus, many species will experience unprecedented climatic conditions across their geographic range. If those conditions exceed the tolerances of local populations, and those populations cannot migrate or evolve fast enough to keep up with climate change, extinction will be likely. These impacts of rapid climate change will moreover occur within the context of an ongoing major extinction event that has up to now been driven primarily by anthropogenic habitat destruction.
Most projections of future climate-driven extinctions rest upon the assumption that potential geographic distribution of each species is ultimately determined by the climatic tolerances of the populations that make up that species. These tolerances define a species “climate envelope” which moves in space as the global climate changes,

FIGURE 2.19 Global climatic conditions (here exemplified by temperature rise) by 2050-2100 are expected to be outside the range that most living species have ever experienced (figure from Moritz and Agudo, 2013).
causing the decline of populations at the trailing edge. If a species’ populations cannot adapt fast enough to tolerate local climate change, or migrate fast enough to track the changing geographic location of suitable climate space at the leading edge of the species range, that species will go extinct (Aitken et al., 2008; Corlett and Westcott, 2013). Species distributions have shifted across the landscape in response to past climate change without evidence of climate-driven elevated extinction rates (Moritz and Agudo, 2013; Jackson and Weng, 1999; Sandel et al., 2011). However, those past climate changes were considerably slower and less intense than what species are expected to experience over the next 30 to 80 years, projections which lead to forecasts of significant future extinctions (Moritz and Agudo, 2013). For example, recent work suggests that up to 41 percent of bird species, 66 percent of amphibian species, and between 61 percent and 100 percent of corals that are not now considered threatened with extinction will become threatened due to climate change sometime between now and 2100 (Foden et al., 2013; Ricke et al., 2013), and that in Africa, 10-40 percent of mammal species now considered not to be at risk of extinction will move into the critically endangered or extinct categories by 2080, possibly as early as 2050 (Thuiller et al., 2006).
An important consideration for such projections is the spatial velocity of climate change (Diffenbaugh and Field, 2013; Loarie et al., 2009). At the last glacial-interglacial transition (the most rapid global climatic transition known prior to today’s), the range of plant dispersal velocities was between 0.1 and 1.0 km/yr (Loarie et al., 2009), with some species lagging behind their moving climate envelope (Normand et al., 2011; Ordonez, 2013). Predicted climate velocities for the next century are considerably faster. Loarie et al. (2009) calculated velocities of climate change in terms of relative changes in temperature gradients using three different emissions scenarios (A2, AB, and B1) and concluded that between 2050 to 2100, organisms now living in areas that cover about 29 percent of the planet’s land will have to disperse faster than observed post-glacial velocities. Diffenbaugh and Field (2013), using the more recent RCP8.5 scenario, expressed velocities in terms of nearest equivalent temperatures. Their calculations indicated that by 2081-2100, most terrestrial species on the globe would need to disperse at a rate that exceeds 4 km/yr, and that nearly half of the land surface would require dispersal velocities that exceed 8 km/yr. Over roughly a third of Earth’s lands, dispersal velocities would need to exceed 16 km/yr, with velocities in high-latitude regions reaching more than 128 km/yr (Figure 2.20). All of these estimates assume no inhibition of dispersal, with transient trajectories following unobstructed “climate paths” to the predicted future climate space. However, short-term climate fluctuations may result in transient loss of suitable climate for certain species, thus preventing those species from migrating to track suitable future conditions (Early and Sax, 2011).
FIGURE 2.20 As temperatures rise, populations of many species will have to move to new habitats to find suitable food, water, and shelter. The colors on these maps show how fast individuals in a species will have to move across the landscape in order to track the mean temperature that now characterizes the places where they live. The figure shows two methods of calculating the velocity of climate change for different time periods at the end of this century. The top panel shows the velocity in terms of nearest equivalent temperature, i.e., the climate change velocity in the CMIP5 RCP8.5 ensemble, calculated by identifying the closest location (to each grid point) with a future annual temperature that is similar to the baseline annual temperature. The lower panel expresses velocity as change in present temperature gradients calculated by using the present temperature gradient at each location and the trend in temperature projected by the CMIP3 ensemble in the SRES A1B scenario. Source: Diffenbaugh and Field, 2013.
Will species be able to track future rapid climate change? Species range shifts in response to the past 50 years or so of warming climate have already been observed (Chen et al., 2011; Parmesan, 2006; Parmesan and Yohe, 2003; Poloczanska et al., 2013; Root et al., 2003). Recent meta-analyses indicate that on average, examined terrestrial species have been moving poleward about 1.76 km/yr (reported as 17.6 + 2.9 km/ decade), apparently keeping pace with regional temperature change, although species range shifts to higher elevations have on average lagged behind climate (Chen et al., 2011). However, individual species vary widely in observed dispersal velocity (Chen et al., 2011), and several studies report many plant populations lagging behind recent warming (Zhu et al., 2012, Corlett and Westcott, 2013). Marine species have been moving poleward at about 7.2 km/yr (reported as 72 + 13.5 km/decade) (Poloczanska et al., 2013). The faster rates in marine organisms may occur because dispersal is enhanced by ocean currents. It is unknown whether the species that have been exhibiting a range-shift response (Chen et al., 2011; Parmesan, 2006; Parmesan and Yohe, 2003; Poloczanska et al., 2013; Root et al., 2003) will be able to accelerate their dispersal velocities to keep pace with the climate change expected over the next few decades under business-as-usual scenarios. This is an area that requires more investigation. We also need to know more about the role of evolutionary adaptation in shaping future species range shifts (Hoffmann and Sgro, 2011).
It is an open question whether the climatic tolerances of local populations can evolve fast enough to keep up with rapid climate change (Aitken et al., 2008; Hoffmann and Sgro, 2011; Moritz and Agudo, 2013). Rapid phenotypic evolution may be required to track changing conditions. For example, adaptation of Sitka spruce to climate change projected for 2080 under the A2 scenario (using the Canadian and Hadley GCMs) would require advancement of annual bud set date within each local population by over 50 days (Aitken et al., 2008). Such rapid change can only occur if there is sufficient genetic variation in the selected population. In some cases, extensive gene flow or assisted migration from populations in warmer parts of the range may enhance the genetic potential for rapid evolutionary response of poleward populations (e.g., Kuparinen et al., 2010). However, adaptive evolution of populations at the warmer range limit of a species will be limited by the amount of genetic variation and covariation within populations for traits affecting climate tolerance (Hoffmann and Sgro, 2011; Shaw and Etterson, 2012). In general, “evolutionary rescue” from extinction pressures induced by climate change requires large populations and high levels of genetic variation for natural selection to act upon (Alberto et al., 2013; Moritz and Agudo, 2013), and may thus be unlikely to occur in many species, particularly rare endemics, and species whose genetic variation has already been severely decimated by other extinction pressures (for instance, species like tigers and black rhinos). If adap-
tive evolution lags behind the rate of environmental change, population viability will decline, increasing the risk of local and global extinction (Aitken et al., 2008; Maurer, 1999; Stephens et al., 1999).
A critical consideration is that the biotic pressures induced by climate change will interact with other well-known anthropogenic drivers of extinction to amplify what are already elevated extinction rates. Even without putting climate change into the mix, recent extinction has proceeded at least 3-80 times above long-term background rates (Barnosky et al., 2011) and possibly much more (Pimm and Brooks, 1997; Pimm et al., 1995; WRI, 2005),17 primarily from human-caused habitat destruction and overexploitation of species. The minimally estimated current extinction rate (3 times above background rate), if unchecked, would in as little as three centuries result in a mass extinction equivalent in magnitude to the one that wiped out the dinosaurs (Barnosky et al., 2011) (see Box 2.4). Importantly, this baseline estimate assumes no effect from climate change. A key concern is whether the added pressure of climate change would substantially increase overall extinction rates such that a major extinction episode would become a fait accompli within the next few decades, rather than something that potentially would play out over centuries.
Known mechanisms by which climate change can cause extinction include the following.
- Direct impact of an abrupt climatic event—for example, flooding of a coastal ecosystem by storm surges as by seas rise to levels discussed earlier in this report.
- Gradually changing a climatic parameter until some biological threshold is exceeded for most individuals and populations of a species across its geographic range—for example, increasing ambient temperature past the limit at which an animal can dissipate metabolic heat, as is happening with pikas at higher elevations in several mountain ranges (Grayson, 2005). Populations of ocean corals (Hoegh-Guldberg, 1999; Mumby et al., 2007; Pandolfi et al., 2011; Ricke et al., 2013) and tropical forest ectotherms (Huey et al., 2012) also inhabit environments close to their physiological thermal limits and may thus be vulnerable to climate warming. Another potential threshold phenomenon is decreasing ocean pH to the point that the developmental pathways of many invertebrates (NRC, 2011a; Ricke et al., 2013) and vertebrate species are disrupted, as is already beginning to happen (see examples below).
____________________
17 The wide range of estimates for exactly how much extinction rates are now elevated is because there is much statistical uncertainty in estimating the background rate from fossils, even in the best cases where the fossil record is reasonably good (as for mammals).
- Interaction of pressures induced directly by climate change with non-climatic anthropogenic factors, such as habitat fragmentation, overharvesting, or eutrophication, that magnify the extinction risk for a given species—for example, the checkerspot butterfly subspecies Euphydryas editha bayensis became extinct in the San Francisco Bay area as housing developments destroyed most of their habitat, followed by a few years of locally unfavorable climate conditions in their last refuge at Jasper Ridge, California (McLaughlin et al., 2002).
- Climate-induced change in biotic interactions, such as loss of mutualist partner species, increases in disease or pest incidence, phenological mismatches, or trophic cascades through food webs after decline of a keystone species. Such effects can be intertwined with the intersection of extinction pressures noted in mechanism 3 above. In fact, the disappearance of checkerspot butterflies from Jasper Ridge was because unusual precipitation events altered the timing of overlap of the butterfly larvae and their host plants (McLaughlin et al., 2002).
BOX 2.4 MASS EXTINCTIONS
Mass extinctions are generally defined as times when more than 75 percent of the known species of animals with fossilizable hard parts (shells, scales, bones, teeth, and so on) become extinct in a geologically short period of time (Barnosky et al., 2011; Harnik et al., 2012; Raup and Sepkoski, 1982). Several authors suggest that the extinction crisis is already so severe, even without climate change included as a driver, that a mass extinction of species is plausible within decades to centuries. This possible extinction event is commonly called the “Sixth Mass Extinction,” because biodiversity crashes of similar magnitude have happened previously only five times in the 550 million years that multi-cellular life has been abundant on Earth: near the end of the Ordovician (~443 million years ago), Devonian (~359 million years ago), Permian (251 million years ago), Triassic (~200 million years ago), and Cretaceous (~66 million years ago) Periods. Only one of the past “Big Five” mass extinctions (the dinosaur extinction event at the end of the Cretaceous) is thought to have occurred as rapidly as would be the case if currently observed extinctions rates were to continue at their present high rate (Alvarez et al., 1980; Barnosky et al., 2011; Robertson et al., 2004; Schulte et al., 2010), but the minimal span of time over which past mass extinctions actually took place is impossible to determine, because geological dating typically has error bars of tens of thousands to hundreds of thousands of years. After each mass extinction, it took hundreds of thousands to millions of years for biodiversity to build back up to pre-crash levels.
These dangers of extinction from climate change are well documented for mammals, birds, reptiles, amphibians (Foden et al., 2013; Pimm, 2009; Sinervo et al., 2010), and corals (Hoegh-Guldberg, 1999; Mumby et al., 2007; Pandolfi et al., 2011; Ricke et al., 2013). Theoretical considerations and some empirical data also indicate that continued climate change at its present pace would be detrimental to many species of marine clams and snails, fish, tropical ectotherms, and some species of plants (examples and citations below). For such species, continuing the present trajectory of climate change would very likely result in extinction of most, if not all, of their populations by the end of the 21st century. The likelihood of extinction from climate change is low for species that have short generation times, produce prodigious numbers of offspring, and have very large geographic ranges. However, even for such species, the interaction of climate change with habitat fragmentation may cause the extirpation of many populations. Even local extinctions of keystone species may have major ecological and economic impacts.
The interaction of climate change with habitat fragmentation has high potential for causing extinctions of many populations and species within decades (before the year 2100 if not sooner). The paleontological record and historical observations of species indicate that in the past species have survived climate change by their constituent populations moving to a climatically suitable area, or, if they cannot move, by evolving adaptations to the new climate. The present condition of habitat fragmentation limits both responses under today’s shifting climatic regime. More than 43 percent of Earth’s currently ice-free lands have been changed into farms, rangelands, cities, factories, and roads (Barnosky et al., 2012; Foley et al., 2011; Vitousek et al., 1986, 1997), and in the oceans many continental-shelf areas have been transformed by bottom trawling (Halpern et al., 2008; Jackson, 2008; Hoekstra et al., 2010). This extent of habitat destruction and fragmentation means that even if individuals of a species can move fast enough to cope with ongoing climate change, they will have difficulty dispersing into suitable areas because adequate dispersal corridors no longer exist. If individuals are confined to climatically unsuitable areas, the likelihood of population decline is enhanced, resulting in high likelihood of extinction if population size falls below critical values, from processes such as random fluctuations in population size (Maurer, 1999) or Allee effects (Stephens et al., 1999).
These considerations make it very likely that at least some populations and species would likely go extinct, and even more will likely drop below viable numbers of individuals within the next few decades simply because they could not disperse across fragmented landscapes fast enough to keep pace with movement of their required climate zones. Concerted efforts of human-mediated translocation of species could help mitigate this, but the practice is still regarded as controversial and experimental
(McLachlan et al., 2007; Ricciardi and Simberloff, 2009; Hoegh-Guldberg et al., 2008; Sax et al., 2009; Schwartz et al., 2012).
Vulnerabilities of Species to Extinction
The demonstrable vulnerabilities of populations and species to extinction by climate change fall into three categories.
- Those whose physiological tolerances to various climatic parameters will be exceeded by climate change throughout their geographic range.
- Those that will have their growth, development, reproduction, or survival detrimentally impacted by climate change or consequent changes in biotic interactions, resulting in population decline.
- Those that are effectively trapped by habitat fragmentation in areas where climate changes detrimentally, even though suitable climatic habitat may exist for them elsewhere in the world.
Examples of species in Category 1 are: polar bears, which require sea ice in order to thrive, as their primary hunting strategy to maintain adequate fat reserves is waiting for seals to emerge from openings in the ice (Derocher et al., 2004); mountain species such as pikas (Grayson, 2005; Beever et al., 2011), which cannot survive sustained temperatures above ~27°C (80°F); endemic Hawaiian silverswords, which are restricted to cool temperatures at high altitudes and die from moisture stress (Krushelnycky et al., 2013); and some coral species, which are known to die at ocean temperatures that are only 0.5-1°C above the maxima experienced prior to 1998 (Hoegh-Guldberg, 1999; Mumby et al., 2007; Pandolfi et al., 2011).
In Category 2 are many marine species whose growth and development are affected by calcium and aragonite concentrations in ocean water, which vary with increasing acidification caused by adding CO2 to the atmosphere. Already exhibiting detrimental effects are the oyster Crassostrea gigas (Barton et al., 2012; Gazeau et al., 2011) in the US Pacific Northwest, where warmer, more acidic waters cause the oyster eggs to die after a few days of apparently normal development. Experimental work, where organisms are reared in waters simulating ocean chemistry expected by the year 2100, also reveals fatal or potentially detrimental effects on other species, including the oysters Crassostrea virginica (Miller et al., 2009) and Pinctada fucata (Liu et al., 2012), inland silverside fish Menidia beryllina (Baumann et al., 2011), Atlantic cod Gadus morhua (Frommel et al., 2011), sea bass Atractoscion nobilis (Checkley et al., 2009), orange clown fish Amphiprion percula (Munday et al., 2009), and damsel fish (Pomacentrus
amboinensis) (Ferrari et al., 2012). This effect is predicted to be particularly severe for coral-forming species (Ricke et al., 2013). Similarly, many plant populations are stressed by climate change. For example, earlier snowmelt in the Rocky Mountains exposes plants to increased frost damage, (e.g., Inouye, 2008), and declining summer fog causes stress to coastal redwoods (Johnstone and Dawson, 2010). Climate change also causes indirect impacts on plants via outbreaks of pests such as pine bark (Kurz et al., 2008) and spruce bark beetles (National Climate Assessment and Development Advisory Committee, 2013; Bentz et al., 2010).
In Category 3 are many species that will probably experience lethal effects in large parts, but not all, of their geographic ranges. For example, warmer river temperatures could reduce habitat for trout in the Rocky Mountain West up to 50 percent, and locally up to 70 percent, by 2100 (Kinsella et al., 2008). Survival for such species will depend on whether or not viable population sizes will remain in areas where climate does not change unsuitably, and on the potential of surviving individuals to disperse from climatically unsuitable areas into regions with favorable climate. Of particular concern are species now much reduced in numbers of individuals and restricted to protected habitat islands, such as national parks, that are surrounded by humandominated landscapes where survival of the affected species is not possible without changing societal norms (Early and Sax, 2011).
Plausible vulnerabilities are potentially more severe than the demonstrable vulnerabilities. Of primary concern are probabilities of novel and disappearing combinations of climatic parameters (Williams and Jackson, 2007). Novel climates are those that are created by combinations of temperature, precipitation, seasonality, weather extremes, etc., that exist nowhere on Earth today. Disappearing climates are combinations of climate parameters that will no longer be found anywhere on the planet. Modeling studies suggest that by the year 2100, between 12 percent and 39 percent of the planet will have developed novel climates, and current climates will have disappeared from 10 percent to 48 percent of Earth’s surface (Williams et al., 2007). These changes will be most prominent in what are today’s most important reservoirs of biodiversity (including the Amazon, discussed in more detail in the “Abrupt Changes in Ecosystems” section above) and if they result in loss of critical aspects of species’ ecological niches, a large number of extinctions would result. Other circumstances that have high plausibility of accelerating extinctions include climatically induced loss of keystone species, collateral loss of species not necessarily affected by climate directly but dependent on species removed by climate change (for example, the myriad species dependent on coral-building species, see below), and phenology mismatches (disruption of the links between a species’ yearly cycle and the seasons) (Dawson et al., 2011; NRC, 2011a).
Likelihood of Abrupt Changes
It presently is not possible to place exact probabilities on the added contribution of climate change to extinction, but the observations noted above indicate substantial risk that impacts from climate change could, within just a few decades, drop the populations in many species below sustainable levels, which in turn would commit the species to extinction. Thus, even though such species might not totally disappear as a result of climate change within the next two or three decades, climate impacts emplaced during that time would seal the species’ fate of extinction over the slightly longer term. On the other hand, the risks of abrupt extinction (within 30 to 80 years) are high for many species that live within two kinds of highly biodiverse ecoystems—tropical and subtropical rainforests such as the Amazon, and coral reefs. Although rainforests presently cover only about 2 percent of Earth’s land, they harbor about half of the planet’s terrestrial species,18 and the tropics as a whole contain about two-thirds of all terrestrial animal and plant species (Pimm, 2001). It is these areas that are among those expected to experience the greatest relative difference between 20th century and late 21st century climates, including a large proportion of “disappearing” and “novel” climates (Williams et al., 2007). Coral reefs, which plausibly as a result of climate change could disappear entirely by 2100 and almost certainly will be reduced much in areal extent within the next few decades (Hoegh-Guldberg, 1999; Mumby et al., 2007; Pandolfi et al., 2011; Ricke et al., 2013), are essentially the “rainforests of the sea” (Knowlton and Jackson, 2008) in terms of biodiversity. Coral reefs support 800 hard coral species, over 4,000 fish species, over 25 percent of the world’s fish biodiversity, and between 9-12 percent of the world’s total fisheries.19 Species in high-elevation and high-latitude regions may also be especially vulnerable to extinction as their current climate zones disappear.
It is possible to gain some qualitative insights from natural experiments afforded by the fossil record to bound the worst-case scenarios. A 4oC increase in mean global temperature, which is plausible by the year 2100,20 would make mean global temperature similar to what it was 14 to 15 million years ago (Barnosky et al., 2003). Then, areas that are now at the top of the Continental Divide in Idaho and Montana were occupied by large tortoises that could not withstand freezing temperatures in winters
____________________
18http://www.nature.org/ourinitiatives/urgentissues/rainforests/rainforests-facts.xml.
19http://coralreef.noaa.gov/aboutcorals/values/biodiversity/.
20 The IPCC AR5 RCP8.5 scenario suggests that exceeding 4.0°C of warming is “about as likely as not and the AR4 suggests warming of 4.0°C by 2100 (relative to 1980-1999) as the ‘best estimate’ for the A1F1 scenario (IPCC, 2007c; NRC, 2011a). Society may be closer to this trajectory than to the IPCC AR4 A2 scenario, or the AR5 RCP4.5 or 6.0 scenarios. Davis et al., 2013 note that “actual annual emissions have exceeded A2 projections for more than a decade,” citing Houghton, 2008 and Boden et al., 2011.
(Barnosky et al., 2007). At the same time, what are now arid lands in Idaho and Oregon supported forests of warm-temperate trees like those in the mahogany family, presently characteristic of central and South America, and in the deserts of Nevada, forests were composed of trees that now are native to the southeastern United States and eastern Asia, for instance, maple, alder, ash, yellowwood, birch, beech, and poplars (Graham, 1999). Given current emissions trajectories, there is a chance that the temperature increase by 2100 could be near 6oC.21 The last time Earth exhibited a global mean temperature that high, what are now sagebrush grasslands in the southwestern Wyoming and Utah were covered by subtropical, closed canopy forests interspersed with open woodlands (Townsend et al., 2010), reminiscent of subtropical areas in Central America today.
While different continental configurations, elevations, and atmospheric circulation patterns now prevail on Earth, precluding a return to those exact past conditions, the underlying message is that warming of 4o-7o will result in a biotically very different world. At best, changes of such magnitude would trigger dramatic re-organization of ecosystems across the globe that would play out over the next few centuries; at worst, extinction rates would elevate considerably for the many species adapted to pre-global warming conditions, via mechanisms described above (inability to disperse or evolve fast enough to keep pace with the extremely rapid rate of climate change, and disruption of ecological interactions within communities as species respond individualistically).
In the oceans, some insights can be gained by tracking how pH values and relative change in pH values correlated with the most severe past mass extinction event, the end-Permian extinction. At current emissions trends, average pH of the oceans would drop from about 8.1 (current levels) to at least 7.9 in about 100 years (NRC, 2011a).22 A similar change occurred over the 200,000 years leading up to the end-Permian mass extinction, which resulted in loss of an estimated ~90 percent or more of known species (Chen and Benton, 2012; Knoll et al., 2007). The actual extinction event may have been considerably less than 200,000 years in duration, but the vagaries of geological dating preclude defining a tighter time span. While there may well have been multiple stressors that contributed to end-Permian extinctions, hitting critical thresholds of equatorial warming and acidification are now thought to be major contributors
____________________
21 The IPCC AR4 scenario A1F1 also yields a 66% chance of warming as much as 6.4°C (IPCC, 2007c; NRC, 2011a) and the AR5 scenario a similar chance of warming 5.8°C (IPCC, 2013).
22 This estimate holds for both the IPCC AR4 A2 Scenario, in which CO2 concentrations rise to approximately 850 ppm in 2100, or for the the A1F1 Scenario with CO2 concentrations of around 940 ppm in 2100 (IPCC, 2001), and for the RCP8.5 scenario (IPCC, 2013). NRC (2011a) notes that at 830 ppm, tropical ocean pH would be expected to drop .3 pH units; the A1F1 Scenario would result in a larger decrease in pH.
(Hönisch et al., 2012; Payne and Clapham, 2012; Sun et al., 2012). The end-Permian extinction started from a different continental configuration and global climate, so an exact reproduction is not to be expected, but the potential for a very large number of extinctions in the next few decades, as a result of elevated CO2 levels that warm the atmosphere and oceans and acidify ocean waters, is analogous (Hönisch et al., 2012; Payne and Clapham, 2012; Sun et al., 2012).
More recently in geological time, the climatic warming at the last glacial-interglacial transition was coincident with the extinction of 72 percent of the large-bodied mammals in North America, and 83 percent of the large-bodied mammals in South America—in total, 76 genera including more than 125 species for the two continents (Barnosky and Lindsey, 2010; Brook and Barnosky, 2012; Koch and Barnosky, 2006). Many of these extinctions occur within and just following the Younger Dryas, and generally they are attributed to an interaction between climatic warming and human impacts (Barnosky et al., 2004; Brook and Barnosky, 2012; Koch and Barnosky, 2006). The magnitude of climatic warming, about 5oC, was about the same as currently-living species are expected to experience within this century, although the end-Pleistocene rate of warming was much slower. Also similar to today, the end-Pleistocene extinction event played out on a landscape where human population sizes began to grow rapidly, and when people began to exert extinction pressures on other large animals (Barnosky, 2008; Brook and Barnosky, 2012; Koch and Barnosky, 2006). The main differences today, with respect to extinction potentials, are that anthropogenic climate change is much more rapid and moving global climate outside the bounds living species evolved in, and the global human population, and the pressures people place on other species, are orders of magnitude higher than was the case at the last glacialinterglacial transition (Barnosky et al., 2012).
Summary and the Way Forward
The current state of scientific knowledge is that there is a plausible risk for climate change to accelerate already-elevated extinction rates, which would result in loss of many more species over the next few decades than would be the case in the absence of climate change. Many of the extinction impacts in the next few decades could be cryptic, that is, reducing populations to below-viable levels, destining the species to extinction even though extinction does not take place until later in the 21st or following century. The losses would have high potential for changing the function of existing ecosystems and degrading ecosystem services (see Chapter 3). The risk of widespread extinctions over the next three to eight decades is high in at least two critically important ecosystems where much of the world’s biodiversity is concentrated, tropical/
sub-tropical areas, especially rainforests and coral reefs. The risk of climate-triggered extinctions of species adapted to high, cool elevations and high-latitude conditions also is high.
There are several questions that are still at a nascent stage of discovery:
- Exactly which species in which ecosystems are most at risk?
- Which species extinctions would precipitate inordinately large ecological cascades that would lead to further extinctions?
- What is the impact of climate-induced changes in seasonal timing and species interactions on extinction rates?
Likewise, much remains to be learned about whether loss of biodiversity in all cases means loss of ecosystem services (see Chapter 3, section on Ecosystem Services), and what loss of diversity through extinctions would actually cost humanity.
What can be monitored to see abrupt changes coming? Evaluating trends of species decline/persistence in uniform ways, such as using techniques developed and in place by the International Union for the Conservation of Nature (IUCN), is especially important for non-charismatic species that may be essential in controlling ecosystem function, and for marine invertebrates. In general, it would be useful to monitor species composition, abundance, phenotype, genetic diversity, nutrient cycling, etc. in uniform ways in many different ecosystems, especially those thought to have little impact by humans or otherwise set aside as protected areas (national parks, remote regions, etc.) (Barnosky et al., 2012).
Currently, monitoring is taking place in a variety of contexts, and some data-sharing and uniformity of data sharing is emerging with efforts such as the National Ecological Observatory Network (NEON).23 But in general different things are being monitored in different ecosystems, and there is little coordination among different groups. Overall, a more uniform, worldwide system of ecosystem/species monitoring is needed (e.g., Pereira et al., 2013). Additionally, a longer time perspective is needed to develop ways to separate the ecological “noise” from the significant ecological signals that would presage biodiversity collapse. This requires comparing changes observed over decades and centuries to long-term ecological baselines of change interpreted from relevant prehistoric records—much as the climate community has done with comparing recent changes with prehistoric proxy data (Barnosky et al., 2012; Hadly and Barnosky, 2009).
Further research is also needed in several key areas:
____________________
- Developing metrics to set short-term changes observed over decades or centuries in the context of long-term (several hundreds to thousands of years or more) variation in specific ecosystems
- Better monitoring and modeling of population parameters that would predict extinction risk in a wide variety of species
- Better understanding of what species are most imperiled by climate change—of those IUCN species in the vulnerable categories, for example, which would be substantially affected by climate change and which would be more resilient
- Better understanding of which species are true keystones, and which of those are actually at risk from climate
- Better understanding of how particular life history traits of species predict vulnerability
- Better predictive models of spatial and demographic responses of species to changes in specific climate parameters
- Better understanding of the role of species interactions in affecting resilience to climate change
- Better understanding of the costs—in ecosystem services, economics, and aesthetic/emotional value—of losing species through extinction
With improved understanding of these issues, society can make more informed decisions about potential intervention actions (Figure 2.21).
FIGURE 2.21 Improved understanding of adaptive capacity, sensitivity, and exposure to climate change can allow for more informed policy decisions. Potential actions are shown as a function of these variables. Source: Dawson et al., 2011.
























































































