
7
Pathways of Survival and Social Structure During Human Transitions from the Darwinian World
Caleb Finch and Burton Singer
Pathways of survival within and between social structures have undergone remarkable change in recent centuries, largely through minimization of infection as the main cause of morbidity and mortality at all ages. Within the past two centuries, the human life expectancy (LE) has doubled, whether measured at birth (Oeppen and Vaupel, 2002 ) or at age 70 (Finch and Crimmins, 2005). As lifespans lengthened, vascular disease and cancer became dominant causes of morbidity and mortality at later ages. Social status and gender continued to be, as in centuries past (Benedictow, 2004), major influences on morbidity and mortality that in the United States separate socioeconomic status (SES) and gender groups by up to 20 years of LE (Crimmins et al., 2009).
Since the early 1800s, the increasingly rigorous demographic data show inverse relationships between SES and rates of overall mortality, age-specific mortality, and a variety of measures of disease morbidity. These mortality-morbidity relationships to SES hierarchies can be characterized in various ways (Humphreys, 1887; Sorokin, 1959). There is an analogous, but more limited, literature for nonhuman primates. Eusocial insects also have hierarchical communities—e.g., bees (Al-Khafaji et al., 2009; Seeley, 2010) and ants (Hölldobler and Wilson, 2009)—but their structure and modes of operation differ qualitatively from primates and humans (Fewell, 2003).
The recently identified and pervasive inverse association between SES to mortality and morbidity in human populations raises a major question: Has inequality among human groups always been present? A minimal answer at least requires some understanding of the origin and character of social
inequality in humans. We ask the same question for wild animal populations that have well-defined hierarchies, but with evolutionary pathways that differ importantly from social and cultural evolution in humans.
We also move beyond associations in human populations to seek understanding of individual-specific pathways from position in a hierarchy to disease and mortality outcomes. Of necessity, this topic requires longitudinal data on different time scales—daily, weekly, monthly, and over multiple years. For example, a person’s relative position in a wealth or household income hierarchy can be fixed over most of his/her adult life, whereas on a daily or weekly basis the same individual may spend time in different hierarchies—for example, church groups or voluntary organizations, the workplace, military reserve—where the negative impact of low position on income in the workplace may have strong positive countervailing effects from high position and influence in a local church group. The study of the psychological consequences for individuals of having distinct positions in multiple hierarchies simultaneously and in moving among hierarchies on both short and long time scales is a topic in its infancy. The reality of this dynamic, however, indicates the need to move from perception to signatures in the brain to physiological impact and feedback to break new ground on mechanisms that underlie the quite coarse-level associations that occupy most of the contemporary literature on inequality and disease, mortality, and health. Over the long sweep of history, few have had the opportunity to occupy multiple hierarchies. However, contemporary technological advances, particularly in communications, are adding considerable complexity to relationships between hierarchical positions and disease, mortality, and positive health outcomes.
Associations between position in a single social hierarchy and LE have points of commonality and differentiation when comparing humans and animals. The change from primarily infectious disease causes of death to noncommunicable chronic diseases in humans leads to different mechanisms associated with pathways to death and a major deviation from the Darwinian background where mortality from predation and infection was dominant. Extending the current focus in studies of LE from single hierarchies to the more nuanced consideration of multiple hierarchies and movement of individuals among them will require longitudinal data collection on human populations with this particular kind of emphasis. A particularly valuable shift of emphasis would be the collection of information on dyadic (network) relationships among individuals in diverse communities. This would facilitate consideration of more subtle, and important, notions of hierarchy than heretofore. Apicella et al. (2012) describe an interesting example from hunter-gatherers in Tanzania.
A central point about hierarchy that social network data would illuminate is that the word “hierarchy” itself has multiple meanings, only one
of which is encapsulated in the vast literature on SES and its association with morbidity and mortality. As discussed above, a hierarchy means a set of ordered categories into which individuals are classified. If dyadic relationships among individuals were assessed, then an idealized notion of hierarchy is a tree-structure (directed graph) in which the vertices (nodes) represent individuals, with the directed arrows signifying an ordering, or dominance, based on criteria such as wealth, power, position in an organization, control, and others. Few real-world communities that we will consider have a structure of this type. A common theme is that at intermediate levels in a tree structure, a subgroup of the community that are essentially unorderable among themselves may be encountered, but as a group, they dominate individuals or other subgroups lower down in a tree-like structure.
For example, a pair of individuals at the top of a hierarchy may dominate the overall community, but cannot be ranked between themselves. Lower down in the tree-like structure, there may be other unorderable groups that, as collectivities, dominate individuals or other unorderable groups lower down. The combination of individual vertices and subsets of unorderable individuals constitute a tree-like structure that is a cruder approximation of the idealized tree structure, the more subgroups there are containing unorderable individuals. In contrast to the branching from the top description just provided, a community may be top-heavy with many individuals dominating fewer and fewer individuals as one moves down a tree that narrows and appears as a cone with apex at the bottom. With no intermediate unorderable groups in such a top-heavy tree, an idealized reverse shape of the idealized tree with single vertex at the bottom can be envisioned. Scoring these extreme trees as +1 for the standard idealized tree and –1 for the inverted idealized tree would score approximations to each of these according to a degree of treeness taking values between –1 and +1. The standard hierarchies in the SES–morbidity mortality literature have no branching structure at all, and thus score 0 in terms of degree of treeness. For a rigorous discussion of measures of treeness, see Corominas-Mutra et al. (2011).
In addition to treeness, it is useful to bring in the notion of feedforwardness, meaning the extent to which there is a flow of order relations from the top of a structure to the bottom. An idealized tree structure as described above would have full feedforwardness. However, if there are intermediate groups in a graph that are unorderable, but as a collectivity if each forms a subgraph of a larger graph that is orderable, degrees of feedforwardness in a community can be represented as a directed graph. A score of 0 corresponds to a cyclic graph—no orderability—and a score of 1 corresponds to a full-ordered structure with no intermediate unorderable subgroups. Feedforwardness and degree of orderability (with scores ranging between 0 and 1) are also fully developed quantitatively in the insightful papers of Corominas-Mutra et al. (2011, 2013).
Assuming dyadic data corresponding to relationships between individuals in a community, the community can be scored as a point in a 3-dimensional morphospace with coordinates T (= treeness, taking values between –1 and +1), F (= feedforwardness, taking values between 0 and +1), and O (= orderability, taking values between 0 and +1). This kind of representation adds considerable nuance to the notion of hierarchy. A substantial research agenda for the future concerns investigation of the dynamics of communities of diverse types and their trajectories in the morphospace with coordinates (T,F,O).
With this background at hand, the purposes of this paper are to (1) review and compare hierarchy–morbidity/mortality associations in animals and humans and identify their place in a more rigorous quantitative formulation of “hierarchy”; (2) embed current evidential bases about the origin of inequality in humans in a quantitative caricature of the changing structure of hierarchies; (3) indicate the shift from a Darwinian view of human lifespan to present and future complex settings where survival extends well beyond reproductive ages; and (4) specify a research agenda focused on a quantification of hierarchy involving multiple kinds of ties simultaneously. The latter topic is a critical missing link in the current studies of associations between membership in stratification systems and morbidity–mortality outcomes.
SOCIAL HIERARCHIES, PAST AND PRESENT
On the Origin of Hierarchies in Human Populations
Hunting and gathering societies that possessed no grouping larger than the extended family were, as seen from archaeological evidence (Flannery and Marcus, 2012), to be nonhierarchical. The networks of extended families approximated the location (0,0,0) in the morphospace. They tended to be small, almost completely connected networks governed by reciprocal friendship and work relationships. Cooperation was the order of the day. At the most basic level, neither individual nor family units were orderable on the basis of ties involving interpersonal interactions. Furthermore, in the arctic and in prehistoric communities residing in current Colorado, there is considerable archaeological evidence for wide networks of nonrelatives cooperating in hunting on a scale not achievable with extended families alone (Flannery, 1972; Flannery and Marcus, 2012). These are also unorderable networks located at (0,0,0), or close to it, in the morphospace.
The formation of clans in a distant Paleolithic horizon ushered in hierarchy in human communities. Despite a seeming lack of power differential, clans have an “us vs. them” mentality, and violence between clans became commonplace. The simplest version of a two-clan society, one of which
dominates the other, with extended family structure within a clan would have location approximately (1,0.5,0.5). The two clans are unorderable within themselves, but the dominance of one clan over another provides a feedforward structure and partial ordering, with the overall community described by two large collapsed nodes—the clans—and a directed arrow pointing from the dominant to the subordinate clan.
With the onset of agriculture, stable communities had village chiefs, thereby building a small power hierarchy within the village. With the household as the unit of analysis, such communities consisted of an unorderable set of households dominated by the household of the chief. Such a village was located at (1,1,1) for a two-node network consisting of the chief’s household and a collapsed network comprising all other households. The farming communities comprising the Jewish Pale of Settlement in Eastern Europe (Poland, Lithuania, parts of Russia, and other areas), had a two-tiered system ordered by wealth. The majority in these communities was quite poor, with minimal ordering by wealth. Among the rich, there was also limited ordering by wealth, but this group totally dominated the lower class, with rabbis playing a central role in maintaining hierarchies (Wienryb, 1972; Risch et al., 1995). In the morphospace, with the wealthy and the poor collapsed into distinct classes, the overall structure was located at (1,1,1).
The onset of the Industrial Revolution introduced greater within-population differentiation based on wealth. In the language of the morphospace, this meant that populations were tree-like, but that there were multiple tiers on the tree corresponding to essentially unorderable wealth groups, and with a strict hierarchy from one tier to the next moving down from the wealthiest to the poorest groups. The wealth groups reflected an ordering of persons by occupational skills with accompanying wage differentials in the occupational hierarchy. Basically, the organization of production induced the hierarchies, and they have persisted to the present time. Company towns were located at (1,1,1) in the morphospace. This structure broke down and became somewhat more complex when industrial cities grew up with diverse types of companies within them. If individuals are the unit of analysis, select subgroups of a population can be unorderable by wealth but nevertheless fit into a hierarchy with location (1,0.5,0).
Hierarchies and Disease, Illness, and Death
Simple Hierarchies
An early example of a hierarchy–mortality association is available from 19th-century England and Ireland, with ordering based on occupational categories (Humphreys, 1887). The ordering of mortality rates as deaths
per 1,000 under age 5 is consistent across five different attributed causes of under-age-5 mortality. In the language of the morphospace, the nodes of a graph based on these data would be the occupational categories for one or both parents, and the arrows moving downward from “Professional” at the top to “General Service” at the bottom would have the interpretation “is a riskier environment than.” The vertical graph has morphospace coordinates (T,F,O) = (0,1,1), as it is not at all tree-like but is nonetheless orderable and has feedforward structure.
For a more contemporary example of occupational hierarchy, we consider the all-cause 10-year (1969-1979) mortality experience of British Civil Service workers in the Whitehall I study (Marmot et al., 1984) (see Table 7-1).
The key point is that the top three levels in the British Civil Service would qualify as ”Professional” in the classification scheme used above for 19th-century England and Ireland. Thus a refinement in the broad category “Professional“ into the top three Civil Service Grades produces an ordering of mortality rates within the broad category, as well as an ordering relative to a subset of “Middle Class,” represented by “Other“ in Table 7-1. A more nuanced analysis of the Whitehall II population is given in Case and Paxson (2011). For a more comprehensive discussion of hierarchy in work environments, see Spilerman (2000).
Hierarchies of Trajectories
Using longitudinal data from the Wisconsin Longitudinal Study ([WLS]; University of Wisconsin–Madison, 2013), Singer and Ryff (1999) constructed coarse-grained economic status histories and personal relationship pathways and investigated their association with a biomarker risk index. The Economic Pathways are based on classification of individuals according to (1) whether or not the household income of the family in which the person grew up was in the upper (+) or lower (–) half of the income distri-
TABLE 7-1 Mortality in Age Group by Civil Service Grade
| Age | Administrators | Professional and Executive | Clerical | Other |
| 10-Year Mortality: % Died Within 10 Years Beyond Baseline (# of deaths) | ||||
| 40-49 years | 2.1 (10) | 3.7 (211) | 3.6 (29) | 9.6 (27) |
| 50-59 years | 6.4 (27) | 9.3 (493) | 16.0 (216) | 19.3 (155) |
| 60-64 years | 5.6 (4) | 17.2 (188) | 23.9 (148) | 26.7 (144) |
NOTE: Hierarchy rank order: Administrators > Professional and Executive > Clerical > Other.
SOURCE: From Marmot et al. (1984).
bution for the state of Wisconsin when he/she was age 18; and (2) whether or not the individual’s household income at age 59 was in the upper (+) or lower (–) half of the income distribution for the state of Wisconsin at that time. The Economic Pathways were defined as Early life category and Later life, with the inclusive four possibilities: (–, –), persistently negative; (+,–) = downwardly mobile; (–,+) = upwardly mobile; (+,+) = persistently positive. The Relationship Pathways are (1) Negative [N] if the person had an uncaring/abusive parent when growing up or a poor relationship with a significant other in adulthood, or both; and (2) Positive [P] if at least one parent was very caring when the person was growing up, and in adulthood there was a good relationship with a significant other.
Besides the economic and relationship measures, a set of 10 biomarkers for morbidity were assessed on the same individuals at age 59 (see Table 7-2). An individual was classified as having elevated morbidity risk for a given
TABLE 7-2 Economic-Relationship Pathwaysa X Risk Index in the Wisconsin Longitudinal Study
| A. Economic-Relationship Pathway | Fraction with Biomarker Risk Indexb ≥ 3 |
| (–,–); N | 0.69(13)c |
| (+,–): N | 0.65 (8) |
| (–,+); N | 0.55 (9) |
| (+,+); N | 0.36 (11) |
| (–,–); P | 0.22 (9) |
| (+,–); P | 0.31 (13) |
| (–,+); P | 0.20 (10) |
| (+,+) P | 0.36 (11) |
| B. Aggreged Pathways | Fraction with Biomarker Risk Index ≥ 3 |
| At least one economic (–), N | 0.63 (30) |
| (+,+) | 0.36 (22) |
aEconomic pathways are defined as Early life and Later life, with four possibilities: (–,–) persistently negative; (+,–) downwardly mobile; (–,+) upwardly mobile; (+,+) persistently positive. Relationship pathways are (1) Negative [N] if the person had an uncaring/abusive parent when growing up or a poor relationship with a significant other in adulthood, or both; (2) Positive [P] if at least one parent was very caring when the person was growing up and there were good relationships with a significant other in adulthood. For detailed specification of the scales and their scoring, see Singer and Ryff (1999).
bThe biomarkers and thresholds defining elevated risk zones are systolic blood pressure (> 147 mm Hg); diastolic blood pressure (> 82 mm Hg); Waist-hip ratio (> 0.93); Ratio of total cholesterol/HDL (5.9); glycosylated hemoglobin (> 7.0%); urinary cortisol (> 25.6 ug/g creatinine); urinary norepinephrine ( > 47 ug/g creatinine); urinary epinephrine (> 5 ug/g creatinine); HDL cholesterol (< 38 mg/dl); DHEA-S (< 351 ng/ml).
cNumber of persons on the designated pathway.
biomarker if he/she was in the upper (or, in some cases, lower) quartile of values for that measure in the WLS population. The risk index simply counts for each individual the total of biomarker scores with elevated morbidity risk. A threshold of risk index ≥ 3 was interpreted as “elevated risk” based on prior studies that showed that such levels were predictive of earlier mortality, incident cardiovascular disease, and decline in physical functioning (Seeman et al., 2001). For those on a negative relationship pathway, there is a basis for an ordering of economic pathways from persistently positive at the top to persistently negative at the bottom. An alternative order would put the relationship pathways from P at the top to N at the bottom. Table 7-2 shows the associations between socioeconomic pathway structure and the morbidity risk index on a pilot sample from WLS.
Among the more interesting results in Panel A is the fact that if an individual is persistently positive on the economic measures, then the relationship history does not matter to morbidity risk. However, if economic measures are persistently negative, then being on a positive relationship (+,+) pathway tends to maintain lower risk index. Further, the lowest morbidity risk index is for persons who are both upwardly mobile economically and on the positive relationship pathway. The WLS thus reveals a richness of possible findings if two or more types of SES measures are considered simultaneously. This analysis also challenges a large literature that discusses biomarkers, disease, or mortality in association with but one socioeconomic measure.
A useful hierarchy based on an aggregation of the pathways in panel A is shown in panel B with larger sample sizes. This hierarchy implies that persistent economic advantage is protective against physiological wear and tear, regardless of the relationship history. In contrast, at least one period of economic disadvantage combined with negative relational experience lead to elevated morbidity risk.
The above pair of hierarchies is very coarse-grained in time and limited in context. However, this unique dataset (University of Wisconsin–Madison, 2013) also contains information about membership in voluntary organizations, church groups, military experience, and other alternatives to the workplace where hierarchies are salient and where possibly negative disease and disability influences of low position in a work environment can be compensated by positive experience at high rank in other hierarchies. Exploring this kind of question in WLS, which has longitudinal follow-up data on individuals from age 18 to age 74, as well as a current wealth of biomarker data, should be an important sequel to the discussion herein.
We note that the hierarchies discussed here for WLS are not interesting in the context of the morphospace introduced above. Little can be done about this because WLS does not have the requisite network data that would allow us to capture the full impact of social connections on health. For example, Berkman et al. (2000) give a nice theoretical treatment of
the cascade of factors involved in translating social ties of various types to health outcomes. This analysis gives a useful template for network-based surveys that would allow a population, such as in WLS, to be situated in the morphospace with interesting dynamics as the people age.
Having indicated this data analytic opportunity, we caution that person-centered strategies using the full individual history as the unit of analysis should be employed if the nuances of movement between distinct hierarchies at several time scales and the health consequences of this dynamic are to be identified. Such an analysis with a dataset like WLS requires that the information for a single individual with survey responses on over 1,000 variables be distilled into a life history narrative. Then, narratives from a small subsample of the population must be compared and contrasted via text analysis, and sets of composite variables focused on different life domains and varieties of experience need specification for quantitative analysis of the full population for ascertainment of multiple hierarchies.
Details of such a person-centered analysis on WLS data are presented with a focus on mental health outcomes in Singer et al. (1998) and Singer and Ryff (2001), but without the multiple hierarchy focus we are advertising here. An especially informative example of person-centered analysis is given by Zhao et al. (1999), who focused on resilience in young children ascertained in the Child Supplement of the National Longitudinal Study of Youth (NLSY-CS).
Social Hierarchy and Life Expectancy
The Evolution of Lifespans
In current evolutionary theory, the LE is an indirect outcome of natural selection to balance mortality against fecundity. This balance, which must be steady-state neutral or positive for species survival, can be achieved via a vast range of reproductive strategies. At one extreme are mayflies and Pacific salmon with short adult lifespans and death soon after laying thousands of eggs (Finch, 1990; National Research Council, 1997). Species with single event “big-bang” reproduction have negligible parental investment in feeding or protection from predation. Humans, at another extreme, have singleton births, prolonged maturation, and lifespans of many decades.
Relative to the great apes, human reproductive schedules are uniquely evolved in three synergizing features: faster weaning, greater maternal support from fathers and others in the community, and a definitive postmenopausal life phase. Great apes typically are not weaned before age 4 to 5, and may still be suckling until puberty, whereas humans under conditions of natural fertility usually wean by age 2 to 3. Consequently, human pregnancies can occur twice or more frequently than in great apes. The young
child requires considerable support for feeding and care, which is uniquely provided by the father and other members of the social group. Rarely do great ape fathers give any direct child care. Humans are also unique in multigenerational social support that reduces mortality of mothers and their children (Hooper et al., this volume). The grandmother hypothesis proposes that the uniquely human post-menopausal phase was important to the evolution of humans’ long lifespans by enabling maternal support (Hawkes and Coxworth, 2013). More broadly, resource transfer and alloparenting are known to involve many individuals in a community besides grandmothers (Hooper et al., this volume).
Social Hierarchy in Darwinian Outcomes
Social status is also a major factor in morbidity, mortality, and lifespan across the animal kingdom. By morbidity, we mean organ damage, infections, and chronic disease. We present selected examples of the character of social hierarchies in social insects and primates to help calibrate the even greater complexities of human social status, disease, disability, and lifespan.
Honey Bees. The honey bee (Apis mellifera) is an archetypal example of genomic programming for social classes with major differences of mortality and lifespan. Alternate life histories of honey bees include short-lived workers with lifespans of weeks to months and queens that live several years (Rueppell et al., 2007; Finch, 1990; Flatt et al., 2013). Worker bees maintain the queen by providing housekeeping for hygiene, caregiving, and nest repair. As hive bees, their mortality is low. However, in their foraging phase as field bees, worker bee mortality increases rapidly with lifespans of only days (Omholt, 1988; Omholt and Amdam, 2004; Rueppell et al., 2007). The timing of foraging flights varies widely. If born in the late summer, worker bees over-winter and delay foraging until the following spring. If born in the later spring, foraging begins much sooner. Mortality of field bees is attributed to external risk factors of predation and “wear-and-tear” on irreplaceable brittle wing parts and nutrient reserves. Resistance to oxidative stress, starvation, and heat also decline (Remolina et al., 2007), while aging pigment also rapidly increases in brain neurons (Flatt et al., 2013). The relationship of foraging flights to mortality was shown by experimental manipulation (Rueppell et al., 2007; Münch et al., 2013).
The plasticity of honey bee life histories joins numerous examples of polyphenisms, or phenotypes with different lengths of developmental and adult stages that are adaptive for particular environments, as discussed in a recent synthesis by Flatt et al. (2013). The broader role of social class in life history polyphenisms, however, merits further inquiry. In the case of social insects, the social castes are determined from genetically identical eggs by
micronutrients in royal jelly fed during development. However, even within these developmentally defined castes, workers with identical genes show individual variations in their preferences for tending larvae destined to be workers or queens (Robinson et al., 1994). Thus even the most rigidly defined castes show some degree of developmental plasticity (Finch and Kirkwood, 2000).
When considering caste systems in the context of the morphospace, it is natural to think of the castes as being in an order relationship, somewhat like human occupational hierarchies. However, this is decidedly not the case, as communication within and between castes, differing according to a particular task being executed, is a fundamental feature of honey bee and other social insect communities. To illustrate, imagine a network with five tasks (pollen dancing, pollen foraging, pollen storage, feeding pollen, and brood care) as nodes and directed arrows representing types of workers (Forager, Recruit, Nurse, Brood) transmitting information. Four of the tasks (pollen foraging, pollen storage, feeding pollen, and brood care) are not orderable. However, they can be collapsed into a single large node, giving a condensed graph with pollen dancing as the top task with recruits signaling to foragers in the condensed set of nodes via the task, pollen foraging. The condensed graph would be T = 0, F = 0, and O = 1 in morphospace. If individual bees were used as nodes rather than tasks, the result would be a very complex set of interconnections (i.e., communication paths) whose overall position in the morphospace is unclear due to the current unavailability of a technology to measure such a network in detail.
Ants. The caste system in ants is specialized to tasks, and the activities of all individuals are in service of the needs of the entire colony. There is no chain of command, and any ant can communicate with any other ant. Most importantly, ants in worker castes communicate in a feedback fashion with a queen, who also sends direct messages to them. In a word, an ant community is replete with multiple channels of communication in the form of a heterarchy, in which each type of individual can communicate with every other type of individual. This is a vast oversimplification of the caste structure and modes of communication in ant colonies. However, a heterarchical structure with dense communication interconnections between ants is generic (Wilson and Hölldobler, 1988). The communication system for diverse ant species at the individual and community levels is discussed by Hölldobler and Wilson (2009).
Baboons and Chimpanzees. Primate social status is strongly associated with morbidity and mortality in field studies of baboons and chimpanzees. Because of ecological variation, we note the location of each population. Female chacma baboons from Moremi have a range of lifespans that differ
based on the stability of social bonds (Silk et al., 2010). High-ranking females live about 35 percent longer than the bottom third, mainly due to social bond stability. Comparisons with human populations are not appropriate because field studies offer small samples and because field mortality rates are much above modern humans (Hill et al., 2001; Bronikowski et al., 2011).
The very coarse description of female chacma baboons needs to be supplemented by network data over time to show bond formation and dissolution at the level of individual animals. This would facilitate an analysis of interaction structure over time where, for example, detection of block model representations (White et al., 1976) of the full troop would allow placement of baboons in a more behaviorally driven set of positions in the morphospace, and over time. This would give a more nuanced association between mortality and troop hierarchy than is possible with current data.
Female high-ranking chimpanzees from Gombe also tend to live longer (Pusey et al., 1997) and also show robust associations of maternal rank with their offspring. High maternal rank increased survival of offspring ages over a 2-fold range; their daughters also grow faster, mature earlier, and have more offspring (Pusey et al., 1997). High rank also shortened intervals between births (Jones et al., 2010). These reproductive advantages are attributed to better access to food or protection from aggression by the higher ranking. Most deaths at Gombe and elsewhere are attributed to infectious disease (Lonsdorf et al., 2006). Infanticide is another cause: several infants of the lower ranked mothers were killed by higher rank females and their daughters (Pusey et al., 1997).
Higher social status was associated with the parasite load in several populations, based on fecal counts of protozoans and eggs of parasitic worms. In Amboseli baboons, worm egg numbers varied inversely with rank in both sexes, with a greater load in females (Hausfater and Watson, 1976). However, infections in males showed the opposite relationship: As indicated by coughing and diarrhea, infections were more common in lower-ranking males, including older individuals that tend to be low ranking (Archie et al., 2012). While female baboons from the Namib Desert also had more worms and protozoans than males, neither sex showed association of parasite load with social status (Benavides et al., 2012). The Kibale alpha male chimpanzees had more worms, ranging 2-fold across rank and positively associated with cortisol (Muehlenbein, 2006; Muelhenbein and Watts, 2010). Alpha males may receive more grooming than other males (Newton-Fisher, 2002), thus acquiring health benefits through removal of ectoparasites and stress reduction (Sapolsky, 2005).
These associations of rank and parasite load suggest rank-differences in immunity. A large literature documents that resistance to infections and the capacity for wound-healing are influenced by steroids, particularly cortisol,
which can be immunosuppressive. Cortisol has been shown to be elevated in high-ranking baboon males (Muller and Wrangham, 2004a, 2004b). However, within the complex social hierarchy, cortisol varies in relation to degrees of social challenge and stress (Sapolsky, 2005; Gesquiere et al., 2011), a topic that would be better investigated by monitoring dyadic interactions among members of a troop over time. Further analyses of baboon female associations with rank and disability and mortality are forthcoming (Archie and Altmann, personal communication).
Intriguingly, skin wound healing was 25 percent faster in high-ranking male baboons than in lower ranks (Archie et al., 2012). Within the highest ranks, the alpha and beta males did not differ in healing rates, which was unexpected because alphas have lower cortisol than betas. Evolutionary flexibility in hormones and immune function may arise through genetic variation in steroid influences on specific immune functions that include pathogen clearance and tissue regeneration. Evidence for these pathways is the association of allele-specific differences of gene expression in white blood cells with social connectedness (Runcie et al., 2013).
Humans. Despite the huge advances in public health and social services, social status remains a strong influence on morbidity and mortality (Crimmins et al., 2009; Marmot and Sapolsky, this volume). SES effects are strongly shown for both genders in age-related morbidities and survival. Analysis of the multiethnic U.S.-wide National Health and Nutrition Examination Survey, III (NHANES III) Sample (Crimmins et al., 2009) showed that poverty is consistently associated with shorter lifespan, mean and maximum, across a range of vascular disease risk factors. Similar social rank differences are present across the world, from the least to the most developed countries (Marmot and Sapolsky, this volume). The same phenomenon was also shown in the 19th-century study of Humphreys (1887) noted above. Despite the stark differences in operationalization of SES, the basic qualitative findings are quite robust. The dichotomy of poor versus non-poor in Crimmins et al. (2009) was not intended to fully represent the gradations of LE across the complex continuum of hierarchies of human societies.
Social and behavioral factors are the major focus in analyzing these associations with morbidity and mortality. Figure 7-1 outlines some of the pathways by which sociobehavioral traits are transduced into outcomes of morbidity and mortality (Crimmins and Finch, 2006).
Boxes labeled “Infection(s)” and “Diet” also include variable exposure within an SES hierarchy. The subsidiary boxes are associated with consequences of infection(s) and dietary quantity/quality, and drugs and medications. Before antibiotics became generally available after 1950, exposure to infections and co-infections was greater in the lower classes. Moreover, concurrent malnutrition exacerbated the cross-class differences, with associated
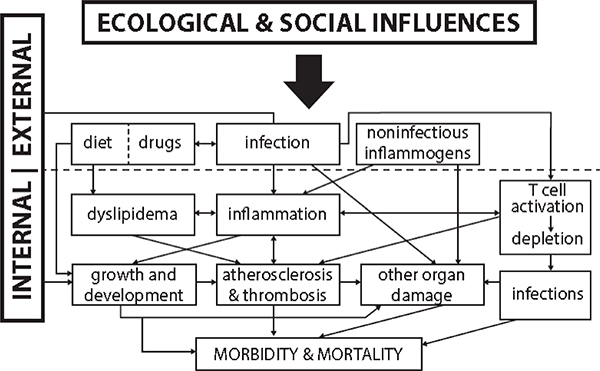
FIGURE 7-1 External and internal pathways to morbidity and mortality.
SOURCE: Expanded from Crimmins and Finch (2006) to include ecological and social influences.
morbidity and mortality via the pathways shown in the figure. Indeed, increased inflammation, dyslipidemia, atherosclerosis and thrombosis, and other organ damage, together with retarded growth and development, were a direct consequence of increased mono- and co-infection coupled to increasingly inadequate diets in lower SES.
As a specific example, in the Jewish Pale of Settlement in Eastern Europe from the 1500s into the early 20th century, infant and child mortality were very high in the lower tier of wealth, essentially due to infectious diseases and malnutrition (Weinryb, 1972). An interesting manifestation of this was the rapid increase in gene frequency of a mutation for idiopathic torsion dystonia in the higher wealth tier (Risch et al., 1995, 2003). The origin of the mutation is traceable into the 1600s, and it has been propagated into the present by genetic drift almost entirely via the elites, who had much larger families with greater survival than the lower classes.
When did SES differences in LE, mortality, and morbidity emerge? It is not possible to know, because there were no statistically valid data for mortality of populations before the 17th century (Harlow and Laurence, 2002; Hoppa and Vaupel, 2002; Scheidel, 2007; Woods, 2007; Bengtsson and van Poppell, 2011). Only Sweden has national comprehensive birth and death records from the mid-18th century. Moreover, there are deep
uncertainties in the comprehensiveness and reliability of public and private records, particularly for lower status groups. Skeletal age estimates of adults are arguably limited to +/– 5 years after age 30, while samples are subject to bone loss from acidic soils and by burial and funerary practices, particularly for infants (Finch, 2010, notes 42-45). Records for the poorest are even more elusive, particularly for criminals sentenced to work in mines or row in galleys, and for the urban homeless. The mortality of the dispraised lowest tier could be extremely high. Two millennia ago, Pliny the Younger told of varying mortality, comparing his Tuscany summer estate to pestilent Rome: “… My servants are healthier here…. I have not lost a single one I brought with me…. You can see grandfathers and great-grandfathers of living men and hear stories of the past” (Pliny the Younger, Letter to Domitius Appolinaris, B. Radice trans., 1969). Caveats granted, records of a few special groups before the 17th century suggest that most elites incurred the same high mortality as other groups in the preindustrial era, from rampant infections (Bengtsson and van Poppell, 2011).
The British peerage may have the deepest historical record for a self-defined society, for some families extending back to 740 CE. Two analyses show gradual increase of aristocrats’ lifespans after 1700, presaging improvements in the general population (Harris, 2004, based on Hollingsworth, 1964; Westendorp and Kirkwood, 1998).
By the mid-1600s, stable SES differences in mortality were emerging for those of sufficient status to be recognized as permanent residents. A strong case is the city of Geneva, which has kept comprehensive records since the 1550s. By 1625-1644, the upper class LE was about 10 years longer for men and women than the middle and lower classes (Schumacher and Oris, 2011, from Perrenoud, 1975). Even so, these Geneva elites’ life expectancies at birth were 37 years (male) to 38 (female), that is, slightly below pre-industrial Sweden and 20th century forager-farmers with limited access to modern medicine (Gurven et al., 2007).
Since the mid-17th century, SES differences in LE have emerged across Europe, with much SES heterogeneity and many fluctuations (Bengtsson and van Poppell 2011; Schumacher and Oris, 2011). Although elites had better diets than the general populace (McKeown, 1976; Fogel, 2004; Harris, 2004), health and good diet may not be sufficiently protective in highly infectious environments. A recent example is the influenza pandemic of 1918, where the healthiest adult group of men and women, ages 20 to 40, had the highest mortality (Morens and Fauci, 2009). We provisionally conclude that SES differences in LE were sporadic in the preindustrial world and did not appear to have stabilized across populations until the 20th century when infectious mortality was greatly reduced.
Since the mid-1800s, the increasing LE began to grow faster for women than men in the most developed countries, from historical baseline of
< 1 year difference, to > 3 years by 1970 (Barford et al., 2006; Thorslund et al., 2013). Although the differences have narrowed in the richer countries, all countries of the world now have excess female LE. The sex differences in lifespan are largely a 20th-century phenomenon and emerged as infectious causes of death fell from > 70 percent of mortality to < 5 percent. Now, cancer and vascular disease are definitively the main morbidities and causes of death in both genders.
In the longitudinal U.S. Health and Retirement Study, for all SES, women had more morbidities at later ages, despite their longer LE (Ailshire and Crimmins, 2013). Low SES was associated with poorer balance, grip strength, and walking speed, and higher blood pressure at later ages, which predicted higher mortality in the following 3 years (Ailshire and Crimmins, 2013). For ages 80 and greater, health-damaging behaviors (alcohol, smoking, low physical activity) accounted for most (68 percent) of the excess mortality with low SES (Nandi et al., 2014).
Besides gender differences in smoking, diet, exercise, and health-promoting behaviors, there may be some underlying biological factors. A growing literature also documents relevant sex differences in animal models studied in vitro, for example, vasoresponses (Zhang et al., 2012), and resistance of cardiomyocytes to hypoxia, which is greater in female-derived primary cultures (Ross and Howlett, 2012). We anticipate a biological substratum in sex differences of chronic disease incidence and progression, in which gene x environment (GxE) effects differ by SES. Invertebrate models show social influences on lifespan through pheromones and sex differences in the p53 gene tumor-suppressor (Finch and Tower, 2014).
FUTURE HUMAN AGING
What lies ahead? How future humans will experience aging may vary widely within populations because of environmental changes and because of access to expansive medical advances. There is also likely to be much greater emphasis on health as defined in the positive sense, and not just as the absence of illness, disease, and disability. We first consider the range of concepts pertaining to “health.”
“Health” as the Absence of Illness, Disease, and Disability
Human health is undergoing huge improvements consequential to increased availability of food and the control of infections, entering into a post-Darwinian era with much less selection for maximizing reproduction. These changes began 250 years ago with progressive but sporadic improvements in nutrition and sanitation, followed by medical and hygienic advances in the later 19th century, and lastly by antibiotics after 1950 (Finch
and Crimmins, 2004). In developed nations, most adults expect only the occasional acute infection, while 2 billion people still carry parasitic worms (World Health Organization, 2013).
But! The reproductive success of Homo sapiens prior to the industrial age depended on its capacity for enduring chronic infections of microbial and invertebrate parasites, as well as nutritional fluctuations. A contemporary example is the Tsimane, indigenous forager-farmers of the Amazon basin, who until recently lived under minimal hygiene and medicine. Their infant mortality of 10 percent or higher (Gurven et al., 2007) and LE of 42 years are similar to pre-industrial Sweden in 1800. Infections were rampant and white blood cell counts were 3-fold higher than the U.S. average (Vasunilashorn et al., 2010). Tsimane adults have frequent gastrointestinal and respiratory conditions that keep them bed-bound several days per month (Tanner and Rosinger, in press) and additionally carry an everage of 1.3 different species of worms and protozoans (Vasulinashorn et al., 2010). These disease burdens would be considered unhealthy and immediately treated in developed countries, as indeed they are during interviews of the Tsimane by anthropologists. Yet, Tsimane women maintained a very high fertility of an average of 9.1 live births (Gurven et al., 2007). The global growth of human populations during the Neolithic, despite the burden of infections, suggests that the Tsimane were typical in maintaining high fertility under conditions of high infection and fluctuating food availability. Thus, the expectation of life relatively free of infection through old age is a modern concept.
It is useful to compare the absence-of-disease concept of health with the historical usage of this word in the English language. Our word “health” draws from the Old English h![]() lth, with positive connotation of well-being and fitness (Oxford English Dictionary). The operationalization of “health” thus leads to the consideration of well-being, a topic with an extensive literature (Ryff and Singer, 1998, 2008; Ryff, 2014). Further, the founding charter of the World Health Organization (WHO) defined health as a “state of complete physical, mental and social well-being and not just the absence of disease or infirmity” (World Health Organization, 1948). However, most “health statistics” during the 20th century have mainly focused on illness, disease, and death (morbidity and mortality). Canada is a leading counterexample, having introduced criteria for positive health and well-being two decades ago in national health statistics (Canadian Population Health Initiative, 2009). Ongoing research is addressing biological substrates of well-being (Ryff et al., 2006; Ryff, 2014).
lth, with positive connotation of well-being and fitness (Oxford English Dictionary). The operationalization of “health” thus leads to the consideration of well-being, a topic with an extensive literature (Ryff and Singer, 1998, 2008; Ryff, 2014). Further, the founding charter of the World Health Organization (WHO) defined health as a “state of complete physical, mental and social well-being and not just the absence of disease or infirmity” (World Health Organization, 1948). However, most “health statistics” during the 20th century have mainly focused on illness, disease, and death (morbidity and mortality). Canada is a leading counterexample, having introduced criteria for positive health and well-being two decades ago in national health statistics (Canadian Population Health Initiative, 2009). Ongoing research is addressing biological substrates of well-being (Ryff et al., 2006; Ryff, 2014).
How can health be conceptualized for a post-Darwinian context wherein reproductive success is no longer a criterion for viability, and when long lives of good health—in the WHO sense of the word—are an increasingly common goal? We suggest that concepts of health at later ages will
require refinement beyond the broad WHO formulation, with particular emphasis on SES and gender, as well as pathways through multiple hierarchies experienced at different time scales. As examples, despite their longer lifespans, older U.S. women are more likely to be obese than men (Wang and Beydoun, 2007) and are 50 percent more likely to survive a heart attack (Lawlor et al., 2001). Because the prevalence of chronic conditions accelerates after age 60, almost no one reaches later ages without some preclinical condition. Among centenarians, > 75 percent have clinical-grade dementia (Perls, 2004). Yet, the majority of centenarians report happiness (Jopp and Rott, 2006). More precise taxonomies of health lie in the future. Meanwhile, definitions of health and well-being are operationalized by vague norms for each age group.
We anticipate that increasing availability of social and economic network data will result in much more nuanced taxonomies of hierarchies that will facilitate classification of communities into the morphospace outlined above, as well as representing community-level dynamics via trajectories in the morphospace. Associating position in these more refined hierarchies with health status and disease/disability condition as a function of age will enable more comprehensive analyses of hierarchy and health and disease outcomes than heretofore.
Aging Futures
Lastly we ponder the future of human lifespans. Humans are entering a post-Darwinian world with minimal natural selection for resistance to acute and chronic infections, and increasing concern about chronic noninfectious conditions. As the 21st century advances, most evidence points to further global reductions of chronic parasitic diseases and of childhood infections. While ongoing efforts seem likely to eliminate most infectious scourges of the past, it is likely that the very poor will remain at greater risk (Hotez, 2008). Family size is shrinking, with negative population growth in many countries. With the increasing proportions of elderly, noninfectious disease is the main concern associated with heart disease, cancer, and mental decline. Later ages are dominated by women whose LE is now greater than men in all countries.
Could there be unexpected consequences from the relaxation of natural selection for resistance to infections? Could the alleles that conferred resistance in early life have delayed adverse consequences to later ages? This possibility extends the “antagonistic pleiotropy hypothesis” of aging. Natural selection tends to favor youth over older ages because evolutionary pressures are greatest on the reproductive ages (Haldane, 1941; Medawar, 1952; Williams, 1957). Thus, pleiotropic gene variants that enhance reproduction in young adults can have delayed adverse consequences at later ages
when natural selection is weaker. Furthermore, with relaxation of natural selection for resistance to infections, previously maladaptive alleles could enter the gene pool with unknown effects. Unfortunately, the earliest population samples available for genetics are a century after the major declines in infectious mortality.
Additionally, environmental changes are occurring. Globally, humanity faces climate change and increasing air pollution from projected global increases in fossil fuels (Finch et al., 2014). Already, the health elite are protected by living and working in buildings with filtered air. Needs for air conditioning will follow warming, adding to increasing fossil fuel use for power, as well as personal vehicles. Access to expensive medical technology will continue to be greater for health elites. The U.N. Intergovernmental Panel on Climate Change (2014) predicts increased poverty throughout the world, even in high-income countries, from rising food prices. Thus we anticipate that the already large SES differentials in health and LE will further expand. Moreover, new Darwinian selection pressures from global environmental challenges may arise, with differential SES reproductive responses. The social network analysis sketched above can be further developed to track emergent trajectories of social support that we believe will remain a key to human health and well-being across the gradients of SES and LE.
ACKNOWLEDGMENTS
Caleb E. Finch is grateful for support by the National Institutes of Health (R21, AG-040683; P01 AG-026572 (R. Brinton, PI); P01 ES-022845 (R. McConnell, PI). Burton Singer thanks the Army Research Office Multidisciplinary Research Initiative for financial support under MURI Grant #58153-MA-MUR, Prime Award #W91 INF-11-1-0036.
REFERENCES
Ailshire, J., and Crimmins, E. (2013). Physical and biological indicators of health and functioning in U.S. oldest old. Annual Review of Gerontology and Geriatrics, 33, 193-215.
Al-Khafaji, K., Tuljapurkar, S., Carey, J.R., and Page, R.E. (2009). Hierarchical demography: A general approach with an application to honey bees. Ecology, 90, 556-566.
Apicella, C.L., Marlowe, F.W., Fowler, J.H., and Christakis, N.A. (2012). Social networks and cooperation in hunter-gatherers. Nature, 481(7382), 4978-501.
Archie, E.A., Altmann, J., and Alberts, S.C. (2012). Social status predicts wound healing in wild baboons. Proceedings of the National Academy of Sciences of the United States of America, 109, 9017-9022.
Barford, A., Dorling, D., Davey Smith, G., and Shaw, M. (2006). Life expectancy: Women now on top everywhere. British Medical Journal, 332, 808.
Benavides, J.A., Huchard, E., Pettorelli, N., King, A.J., Brown, M.E., Archer, C.E., Appleton, C.C., Raymond, M., and Cowlishaw, G. (2012). From parasite encounter to infection: Multiple-scale drivers of parasite richness in a wild social primate population. American Journal of Physical Anthropology, 147, 52-63.
Benedictow, O.J. (2004). The Black Death 1364-1353: The Complete History. Suffolk, UK: The Boydell Press.
Bengtsson, T., and van Poppel, F. (2011). Socioeconomic inequalities in death from past to present: An introduction. Explorations in Economic History, 48, 343-356.
Berkman, L.F., Glass, T., Brissette, I., and Seeman, T.E. (2000). From social integration to health: Durkheim in the new millennium. Social Science and Medicine, 51, 843-857.
Bronikowski, A.M., Altmann, J., Brockman, D.K., Cords, M., Fedigan, L.M., Pusey, A., Stoinski, T., Morris, W.F., Strier, K.B., and Alberts, S.C. (2011). Aging in the natural world: Comparative data reveal similar mortality patterns across primates. Science, 331, 1325-1328.
Canadian Population Health Initiative. (2009). Exploring Positive Mental Health: Improving the Health of Canadians, 2009. Ottawa, Canada: Canadian Institute for Health Information.
Case, A., and Paxson, C. (2011). The long reach of childhood health and circumstance: Evidence from the Whitehall II study. Economics Journal, 121, F183-F204.
Corominas-Mutra, B., Rodriguez-Caso, C., Goni, J., and Sole, R. (2011). Measuring the hierarchy of feedforward networks. Chaos: An Interdisciplinary Journal of Nonlinear Science, 21(1), 016108.
Corominas-Mutra, B., Goni, J., Sole, R.V., and Rodriguez-Caso, C. (2013). On the origins of hierarchy in complex networks. Proceedings of the National Academy of Sciences of the United States of America, 110, 13316-13321.
Crimmins, E.M., and Finch, C.E. (2006). Infection, inflammation, height, and longevity. Proceedings of the National Academy of Sciences of the United States of America, 103, 498-503.
Crimmins, E.M., Kim, J.K., and Seeman, T.E. (2009). Poverty and biological risk: The earlier “aging” of the poor. Journals of Gerontology. Series A: Biological Sciences Medical Sciences, 64, 286-292.
Fewell, J.H. (2003). Social insect networks. Science, 301, 1867-1870.
Finch, C.E. (1990). Longevity, Senescence, and the Genome. Chicago, IL: University of Chicago Press.
Finch, C.E. (2010). Evolving views of ageing and longevity from Homer to Hippocrates: Emergence of natural factors, persistence of the supernatural. Greece & Rome, 57, 355-377.
Finch, C.E., and Crimmins, E.M. (2004). Inflammatory exposure and historical changes in human life spans. Science, 305, 1736-1739.
Finch, C.E., and Crimmins, E.M. (2005). Comment on “Inflammatory exposure and historical changes in human life-spans.” Science, 308(5729), 1743.
Finch, C.E., and Kirkwood, T.B. (2000). Chance, Development, and Aging. New York: Oxford University Press.
Finch, C.E., and Tower, J.G. (2014). Sex-specific aging in flies, worms, and missing great-granddads. Cell, 156(3), 398-399.
Flannery, K. (1972). The cultural evolution of civilization. Annual Review of Ecology, Evolution, and Systematics, 3, 399-426.
Flannery, K., and Marcus, J. (2012). The Creation of Inequality. Cambridge, MA: Harvard University Press.
Flatt, T., Amdam, G.V., Kirkwood, T.B., and Omholt, S.W. (2013). Life-history evolution and the polyphenic regulation of somatic maintenance and survival. Quarterly Review of Biology, 88, 185-218.
Fogel, R. (2004). The Escape from Hunger and Premature Death 1700-2100: Europe, America and the Third World. Cambridge, UK: Cambridge University Press.
Gesquiere, L.R., Learn, N.H., Simao, M.C., Onyango, P.O., Alberts, S.C., and Altmann, J. (2011). Life at the top: Rank and stress in wild male baboons. Science, 333, 357-360.
Gurven, M., Kaplan, H., and Supa, A.Z. (2007). Mortality experience of Tsimane Amerindians of Bolivia. Regional variation and temporal trends. American Journal of Human Biology, 19, 376-398.
Haldane, J.B.S. (1941). New Paths in Genetics. London, UK: Allen and Unwin.
Harlow, M., and Laurence, R. (2002). Growing up and Growing Old in Ancient Rome: A Life Course Approach. London, UK: Routledge.
Harris, B. (2004). Public health, nutrition, and the decline of mortality: The McKeown thesis revisited. Social History of Medicine, 17, 379-407.
Hausfater, G., and Watson, D.F. (1976). Social and reproductive correlates of parasite ova emissions by baboons. Nature, 262, 688-689.
Hawkes, K., and Coxworth, J.E. (2013). Grandmothers and the evolution of human longevity: A review of findings and future directions. Evolutionary Anthropology, 22, 294-302.
Hill, K., Boesch, C., Goodall, J., Pusey, A., and Williams, J. (2001). Mortality rates among wild chimpanzees. Journal of Human Evolution, 40, 437-450.
Hollingsworth, T. (1964). The demography of the British Peerage. Population Studies, 18(2), 52-70.
Hölldobler, B., and Wilson, E.O. (2009). The Superorganism. New York: W.W. Norton & Company.
Hoppa, R.D., and Vaupel, J.W. (2002). The Rostock Manifesto for paleodemography. In R.D. Hoppa and J.W. Vaupel (Eds.), Paleodemography: Age Distributions from Skeletal Samples (pp. 1-8). Cambridge, UK: Cambridge University Press.
Hotez, P.J. (2008). Neglected infections of poverty in the United States of America. PLOS Neglected Tropical Diseases, 2, e256.
Humphreys, N.A. (1887). Class mortality statistics. Journal of the Royal Statistical Society, 50(2), 255-292.
Jones, J.H., Wilson, M.L., Murray, C., and Pusey, A. (2010). Phenotypic quality influences fertility in Gombe chimpanzees. Journal of Animal Ecology, 79, 1262-1269.
Jopp, D., and Rott, C. (2006). Adaptation in very old age: Exploring the role of resources, beliefs, and attitudes for centenarians’ happiness. Psychology and Aging, 21, 266-280.
Lawlor, D.A., Ebrahim, S., and Smith, G.D. (2001). Sex matters: Secular and geographical trends in sex differences in coronary heart disease mortality. British Medical Journal, 323, 541-515.
Lonsdorf, E.V., Travis, D., Pusey, A.E., and Goodall, J. (2006). Using retrospective health data from the Gombe chimpanzee study to inform future monitoring efforts. American Journal of Primatology, 68, 897-908.
Marmot, M.G., Shipley, M.J., and Rose, G. (1984). Inequalities in death—specific explanations of a general pattern. The Lancet, 1(8384), 1003-1006.
McKeown, T. (1976). The Modern Rise of Population. London, UK: Edward Arnold.
Medawar, P. (1952). An Unsolved Problem in Biology. London, UK: HK Lewis.
Morens, D.M., and Fauci, A.S. (2001). The 1918 influenza pandemic: Insights from the 21st century. Journal of Infectious Diseases, 195, 1018-1028.
Morens, D.M., Taubenberger, J.K., and Fauci, A.S. (2009). The persistent legacy of the 1918 influenza virus. New England Journal of Medicine, 361, 225-229.
Muehlenbein, M.P. (2006). Intestinal parasite infections and fecal steroid levels in wild chimpanzees. American Journal of Physical Anthropology, 130, 546-550.
Muehlenbein, M.P., and Watts, D.P. (2010). The costs of dominance: Testosterone, cortisol and intestinal parasites in wild male chimpanzees. Biopsychosocial Medicine, 4, 21.
Muller, M.N., and Wrangham, R.W. (2004a). Dominance, aggression and testosterone in wild chimpanzees: A test of the “challenge hypothesis.” Animal Behaviour, 67, 113-123.
Muller, M.N., and Wrangham, R.W. (2004b). Dominance, cortisol and stress in wild chimpanzees (Pan troglodytes schweinfurthii). Behavioral Ecology and Sociobiology, 55, 332-340.
Münch, D., Kreibich, C.D., and Amdam, G.V. (2013). Aging and its modulation in a long-lived worker caste of the honey bee. Journal of Experimental Biology, 216, 1638-1649.
Nandi, A., Glymour, M.M., and Subramanian, S.V. (2014). Association among socioeconomic status, health behaviors, and all-cause mortality in the United States. Epidemiology, 25, 170-177.
National Research Council. (1997). Between Zeus and the Salmon: The Biodemography of Longevity. K.W. Wachter and C.E. Finch (Eds.). Committee on Population, Commission on Behavioral and Social Sciences and Education. Washington, DC: National Academy Press.
Newton-Fisher, N.E. (2002). Relationships of male chimpanzees in the Budongo Forest, Uganda. In C. Boesch, G. Hohmann, and L.F. Marchant (Eds.), Behavioural Diversity in Chimpanzees and Bonobos (pp. 125-137). Cambridge, UK: Cambridge University Press.
Oeppen, J., and Vaupel, J.W. (2002). Demography. Broken limits to life expectancy. Science, 296, 1029-1031.
Omholt, S.W. (1988). Relationships between worker longevity and the intracolonial population dynamics of the honeybee. Journal of Theoretical Biology, 130, 275-284.
Omholt, S.W., and Amdam, G.V. (2004). Epigenetic regulation of aging in honeybee workers. Science of Aging Knowledge Environment, 26, pe28.
Perls, T. (2004). Centenarians who avoid dementia. Trends in Neurosciences, 10, 633-636.
Perrenoud, A. (1975). L’inegalite social devzant la mort a Geneve au XVIIe siècle. Population, 30, 221-243.
Pliny the Younger. (1969). Letter to Domitius Apollinaris. From Letters of the Younger Pliny (B. Radice, Trans.). New York: Penguin Classic.
Pusey, A., Williams, J., and Goodall, J. (1997). The influence of dominance on the reproductive success of female chimpanzees. Science, 277, 828-831.
Remolina, S.C., Hafez, D.M., Robinson, G.E., and Hughes, K.A. (2007). Senescence in the worker honey bee Apis mellifera. Journal of Insect Physiology, 53, 1027-1033.
Risch, N., de Leon, D., Ozelius, L., Kramer, P., Almasy, L., Singer, B., Fahn, S., Breakefield, X., and Bressman, S. (1995). Genetic analysis of idiopathic torsion dystonia in Ashkenazi Jews and their recent descent from a small founder population. Nature Genetics, 9, 152-159.
Risch, N., Tang, H., Katzenstein, H., and Ekstein, J. (2003). Geographic distribution of disease mutations in the Ashkenazi Jewish population supports genetic drift over selection. American Journal of Human Genetics, 72, 812-822.
Robinson, G.E., Page, R.E., and Arensen, N. (1994). Genotypic differences in brood rearing in honey bee colonies: Context-specific? Behavioral Ecology and Sociobiology, 34, 125-138.
Ross, J.L., and Howlett, S.E. (2012). Age and ovariectomy abolish beneficial effects of female sex on rat ventricular myocytes exposed to simulated ischemia and reperfusion. PLOS ONE, 7, e38425.
Rueppell, O., Bachelier, C., Fondrk, M.K., and Page, R.E., Jr. (2007). Regulation of life history determines lifespan of worker honey bees (Apis mellifera L.). Experimental Gerontology, 42, 1020-1032.
Runcie, D.E., Wiedmann, R.T., Archie, E.A., Altmann, J., Wray, G.A., Alberts, S.C., and Tung, J. (2013). Social environment influences the relationship between genotype and gene expression in wild baboons. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 368(1618), 20120345.
Ryff, C.D. (2014). Psychological well-being revisited: Advances in the science and practice of eudaimonia. Psychotherapy and Psychosomatics, 83(1), 10-28.
Ryff, C.D., and Singer, B.H. (1998). The contours of positive human health. Psychological Inquiry, 9(1), 1-28.
Ryff, C.D., and Singer, B.H. (2008). Know thyself and become what you are: A eudaimonic approach to psychological well-being. Journal of Happiness Studies, 9(1), 13-39.
Ryff, C.D., Love, G.D., Urry, H.L., Muller, D., Rosenkranz, M.A., Friedman, E.M., Davidson, R.J., and Singer, B. (2006). Psychological well-being and ill-being: Do they have distinct or mirrored biological correlates? Psychotherapy and Psychosomatics, 75, 85-95.
Sapolsky, R.M. (2005). The influence of social hierarchy on primate health. Science, 308, 648-652.
Scheidel, W. (2007). Demography. In W. Scheidel, I. Morris, and R. Saller (Eds.), The Cambridge Economic History of the Greco-Roman World (pp. 38-86). New York: Cambridge University Press.
Schumaker, R., and Oris, M. (2011). Long-term changes in social mortality differentials, Geneva, 1625-2004. Explorations in Economic History, 48, 357-365.
Seeley, T. (2010). Honeybee Democracy. Princeton, NJ: Princeton University Press.
Seeman, T.E., McEwen, B.S., Rowe, J.W., and Singer, B.H. (2001). Allostatic load as a marker of cumulative biological risk: MacArthur studies of successful aging. Proceedings of the National Academy of Sciences of the United States of America, 98, 4770-4775.
Silk, J.B., Beehner, J.C., Bergman, T.J., Crockford, C., Engh, A.L., Moscovice, L.R., Wittig, R.M., Seyfarth, R.M., and Cheney, D.L. (2010). Strong and consistent social bonds enhance the longevity of female baboons. Current Biology, 20, 1359-1361.
Singer, B.H., and Ryff, C.D. (1999). Hierarchies of life histories and associated health risks. Annals of the New York Academy of Sciences, 896, 96-115.
Singer B.H., and Ryff, C.D. (2001). Person-centered methods for understanding aging: The integration of numbers and narratives. In R. Binstock and L.K. George (Eds.), Handbook of Aging and the Social Sciences (pp. 44-65). San Diego, CA: Academic Press.
Singer, B.H., Ryff, C.D., Carr, D., and Magee, W. (1998). Linking life histories and mental health: A person-centered strategy. In A. Raftery (Ed.), Sociological Methodology (pp. 1-51). Washington, DC: American Sociological Association.
Sorokin, P. (1959). Social and Cultural Mobility. Glencoe, IL: The Free Press.
Spilerman, S. (2000). Wealth and stratification processes. Annual Review of Sociology, 26, 497-524.
Tanner, S., and Rosinger, A. (in press). Salud y nutrición: Perfil y cambio, Ch 9. In T. Huanca and V. Reyes-Garcia (Eds.), Cambio Local—Cambio Global: El Processo de Cambio en la Sociedad Tsimane’, Amazonia Boliviana. Barcelona, Spain: Icaria Editorial.
Thorslund, M., Wastesson, J.W., Agahi, N., Lagergren, M., and Parker, M.G. (2013). The rise and fall of women’s advantage: A comparison of national trends in life expectancy at age 65 years. European Journal of Aging, 10, 271-277.
U.N. Intergovernmental Panel on Climate Change. (2014). Impacts, Adaptation, and Vulnerability. Available: http://www.ipcc.ch/report/ar5/wg2/ [June 2014].
University of Wisconsin-Madison. (2013). Wisconsin Longitudinal Study. Available: http://www.ssc.wisc.edu/wlsresearch/ [June 2014].
Vasunilashorn, S., Crimmins, E.M., Kim, J.K., Winking, J., Gurven, M., Kaplan, H., and Finch, C.E. (2010). Blood lipids, infection, and inflammatory markers in the Tsimane of Bolivia. American Journal of Human Biology, 22, 731-740.
Wang, Y., and Beydoun, M.A. (2007). The obesity epidemic in the United States—gender, age, socioeconomic, racial/ethnic, and geographic characteristics: A systematic review and meta-regression analysis. Epidemiology Review, 29, 6-28.
Weinryb, B.D. (1972). The Jews of Poland: A Social and Economic History of the Jewish Community of Poland from 1100 to 1800. Philadelphia, PA: The Jewish Publication Society of America.
Westendorp, R.G., and Kirkwood, T.B. (1998). Human longevity at the cost of reproductive success. Nature, 396, 743-746.
White, H., Boorman, S., Breiger, R. (1976). Social structure from multiple networks. I. Block-models of roles and positions. American Journal of Sociology, 81(4), 730-780.
Williams, G.C. (1957). Pleiotropy, natural selection and the evolution of senescence. Evolution, 11(4), 398-411.
Wilson, E.O., and Holldobler, B. (1988). Dense heterarchies and mass communication as the basis of organization in ant colonies. Trends in Ecology and Evolution, 3(3), 65-68.
Woods, R. (2007). Ancient and early modern mortality: Experience and understanding. Economic History Review, 60, 373-399.
World Health Organization. (1948). World Health Organization constitution. In Basic Documents (pp. 1-18). Geneva, Switzerland: World Health Organization.
World Health Organization. (2013). Soil-Transmitted Helminth Infections (Factsheet). Geneva, Switzerland: World Health Organization.
Zhang, X., Thatcher, S.E., Rateri, D.L., Bruemmer, D., Charnigo, R., Daugherty, A., and Cassis, L.A. (2012). Transient exposure of neonatal female to testosterone abrogates the sexual dimorphism of abdominal aortic aneurysms. Circulation Research, 110, e73-85.
Zhao, H., Brooks-Gunn, J., McLanahan, S., and Singer, B. (1999). Studying the real child rather than the ideal child: Bringing the person into developmental studies. In L. Bergman and R. Cairns (Eds.), Developmental Science and the Holistic Approach (pp. 393-419). Mahwah, NJ: Lawrence Erlbaum Associates.
























