
Workshop Overview
GLOBAL HEALTH IMPACTS OF VECTOR-BORNE DISEASES1
Pathogens transmitted among humans, animals, or plants by insects and arthropod vectors have been responsible for significant morbidity and mortality throughout recorded history. Such vector-borne diseases—including malaria, dengue, yellow fever, plague, trypanosomiasis, and leishmaniasis—together accounted for more human disease and death in the 17th through early 20th centuries than all other causes combined (Gubler, 1998). By the mid-20th century, implementation of strategies to reduce populations of the mosquitoes that spread malaria, yellow fever, and dengue effectively reduced the impact of these diseases on human health—albeit temporarily.
Over the past three decades, previously controlled vector-borne diseases have resurged or reemerged in new geographic locations, and several newly identified pathogens and vectors have triggered disease outbreaks in plants and animals, including humans. A variety of factors underlie this trend among emerging vector-borne diseases, including
- The rapid expansion of global travel and trade, enabling the geographic spread of pathogens, vectors, and animals that serve as so-called reservoirs2 of disease;
___________________
1 The planning committee’s role was limited to planning the workshop, and the workshop summary has been prepared by the workshop rapporteur as a factual summary of what occurred at the workshop. Statements, recommendations, and opinions expressed are those of individual presenters and participants, and are not necessarily endorsed or verified by the National Academies of Sciences, Engineering, and Medicine, and they should not be construed as reflecting any group consensus.
2 Defined in glossary, Appendix D.
- Recent, unprecedented, population growth associated with rampant and unplanned urbanization in the tropics, and the resulting increased juxtaposition of humans, animal reservoirs of pathogens, and vector species in geographically constrained environments;
- Societal, cultural, and behavioral practices that encourage disease transmission; and
- Decreased support for and deterioration of the public health surveillance and control infrastructure for infectious diseases in general, and specifically for vector-borne and zoonotic diseases.
Domestic and international capabilities to detect, identify, and effectively respond to vector-borne diseases are limited. Few vaccines have been developed against vector-borne pathogens. At the same time, drug resistance has increased in vector-borne pathogens while their vectors are increasingly resistant to insecticide controls. Furthermore, the ranks of scientists trained to conduct research in key fields including medical entomology, vector ecology, and tropical medicine have dwindled, threatening prospects for addressing vector-borne diseases now and in the future.
In June 2007, as these circumstances became alarmingly apparent, the Forum on Microbial Threats hosted a workshop to explore the dynamic relationships among host, pathogen(s), vector(s), and ecosystems that characterize vector-borne diseases. Revisiting this topic in September 2014, the forum examined trends and patterns in the incidence and prevalence of vector-borne diseases in an increasingly interconnected and ecologically disturbed world, as well as recent developments to meet these dynamic threats. This public workshop featured invited presentations and discussions that described the emergence and global movement of vector-borne diseases, considered research priorities for understanding their biology and ecology, and assessed global preparedness for and progress toward their prevention, control, and mitigation.
WORKSHOP CONTEXT
Disease Burden
Vector-borne diseases3 have long been associated with significant human illness and death. Over half the world’s human populations are currently at risk from vector-borne infections, which collectively account for 17 percent of the human global infectious disease burden (CDC, 2014d). In April 2014, the World Health Organization (WHO) devoted its annual World Health Day to vector-borne illnesses, issuing a global brief that profiled key diseases and their individual and collective impacts (WHO, 2014a). In her foreword to this report, WHO Director-General Margaret Chan noted that vector-borne illnesses caused more than one million deaths each year, but that “death counts, though alarming, vastly underestimate the human misery and hardship caused by these diseases, as many people who survive infection are left permanently debilitated, disfigured, maimed, or blind.” These burdens are borne most heavily by the world’s poorest people, communities, and countries.
Explosive epidemics have marked the recent resurgence of several previously controlled vector-borne diseases, including plague, dengue, and yellow fever. Less sensational—but equally destructive—infectious, vector-borne disease outbreaks in plants, domestic animals, and wildlife have disrupted ecosystems and reduced agricultural productivity. In addition to these acute impacts, persistent vector-borne diseases impose a significant burden on plant, animal, and human health and are an impediment to socioeconomic development (IOM, 2008). This is exacerbated by the chronic or long-term effects of diseases including West Nile viral fever, dengue, chikungunya, and Chagas disease, which have also been associated with chronic or long-term sequelae (Garcia et al., 2011, 2014; Montgomery et al., 2014; Murray et al., 2014; Schilte et al., 2013).
___________________
3 A disease that is transmitted to humans, plants, or animals by an insect or other arthropod (see next footnote) is called a vector-borne disease. (Plant pathologists refer to these as vector-associated diseases.) From the perspective of infectious diseases, vectors—which can be either living (biological) or nonliving (mechanical)—are the transmitters of disease-causing organisms; that is, they carry pathogens from one host to another. By common usage, vectors are considered to be invertebrate animals, usually arthropods. A broader definition of vector-borne disease recognizes that other animals can serve in the role of infectious disease vector by harboring pathogens that cause disease only in susceptible populations. These include invertebrates other than arthropods (e.g., snails, in the case of schistosomiasis), rodents (which spread a variety of viral diseases, including hantavirus pulmonary syndrome), fungi, plants, and even humans (in the case of sudden oak death), who may also serve as vectors for a variety of plant diseases (IOM, 2008).

SOURCE: As presented by Lyle Petersen on September 16, 2014.
Vectors and Pathogens
As illustrated in Figure WO-1, many vector-borne pathogens (viruses, bacteria, fungi, and parasites) are transmitted among and between their primary and incidental hosts by arthropods such as mosquitoes, ticks, biting flies, and aphids.4
These pathogens include the mosquito-borne protozoans (Plasmodium spp.) that cause malaria5 and the tick-borne parasite that causes babesiosis, Babesia microti; the newly described beetle-borne fungus, Geosmithia morbida, that causes thousand canker disease of black walnut trees6; the tick-borne bacterium
___________________
4 Arthropods (members of the phylum Arthropoda) are invertebrates with jointed limbs, segmented bodies, and exoskeletons made of chitin. They include insects, spiders, crustaceans (e.g., shrimp, lobsters), and centipedes.
5 There are many different types of plasmodium parasite, but only five types cause malaria in humans. These are
- Plasmodium falciparum—Mainly found in Africa, it is the most common type of malaria parasite and is responsible for most malaria deaths worldwide.
- Plasmodium vivax—Mainly found in Asia and South America, this parasite causes milder symptoms than P. falciparum, but it can stay in the liver for up to 3 years, which can result in relapses.
- Plasmodium ovale—Fairly uncommon and usually found in West Africa, it can remain in the liver for several years without producing symptoms.
- Plasmodium malariae—This rare species is usually found only in Africa.
- Plasmodium knowlesi—This very rare species is found in parts of South East Asia.
6 See http://www.fs.fed.us/psw/publications/seybold/psw_2010_seybold008(tisserat).pdf (accessed March 25, 2016).
that causes Lyme disease, Borrelia burgdorferi; and the mosquito-borne West Nile and dengue viruses.
The arthropod-borne viruses, or arboviruses, are the largest class of vector-borne human pathogens. More than 500 arboviruses have been described, of which about 100 are known to cause diseases that include dengue, chikungunya, and several types of encephalitis (Gray and Banerjee, 1999; Gubler, 1998; Weaver and Reisen, 2010). Arboviruses circulate among wild animals, and many can be transmitted to humans and agriculturally important domestic animals through a process known as spillover (Weaver and Reisen, 2010). Infectious disease outbreaks resulting from such spillover events include epidemics of West Nile viral fever in the United States and of Rift Valley fever in Africa and the Middle East. Arthropod vectors also transmit most identified plant viruses (Hogenhout et al., 2008), as well as several important fungal and bacterial pathogens of plants (Fletcher and Wayadande, 2002; Gergerich and Dolja, 2006; Weintraub and Beanland, 2006).
Vector–pathogen relationships are central to the epidemiologies of many important plant diseases (Gergerich and Dolja, 2006; Purcell, 1982; Weintraub and Beanland, 2006). While only certain bacterial pathogens of plants require a vector for transmission, most plant viruses are spread from infected to healthy plants via a plant-feeding arthropod, nematode, or plant-parasitic fungus. Even humans appear to serve as vectors of plant disease. Sudden oak death, an emergent pathogen that has caused widespread dieback of several tree species in West Coast forests, has been spread to new areas by hikers, mountain bikers, and equestrians (COMTF, 2013). With sudden oak death, asymptomatic plants are actually the more important vectors.
Several important bacterial pathogens are delivered directly into plants’ sugar-transporting phloem or water-transporting xylem networks by insects that feed on plant vascular fluids (Fletcher and Wayadande, 2002). These unusual pathogens and their multiple hosts provide fascinating examples of complex webs of organismal interactions. They include
- Spiroplasmas and phytoplasmas, which are tiny bacteria transmitted mainly by leafhopper insects. Pathogenic strains cause more than 700 distinct plant diseases, including corn stunt, coconut lethal yellowing, and pear decline (ARS, 2013; Fletcher and Wayadande, 2002; Weintraub and Beanland, 2006).
- Fastidious phloem-colonizing bacteria, so called because they cannot be consistently cultivated from infected hosts (which include species of herbaceous plants, trees, vegetables, fruits, grains, and ornamental plants) (Fletcher and Wayadande, 2002). Diseases caused by this group include citrus greening, which causes major losses in Asia and Africa and has been introduced recently into the United States, and cucurbit yellow vine.
- Fastidious xylem-limited bacteria, transmitted by xylem-feeding sharpshooter insects and spittle bugs. The best studied among these pathogens, Xylella fastidiosa, causes economically important damage in a wide range of plant hosts. In grapevines, it causes Pierce’s disease, a significant threat to California’s table grape and wine industries (Fletcher and Wayadande, 2002; NRC, 2004).
Viral infections of plants, such as the Citrus tristeza virus, stunt growth, lower yield, reduce fruit quality, and thereby diminish agricultural productivity (Gergerich and Dolja, 2006). Aphids transmit Barley yellow dwarf virus, the most widely distributed viral disease of cereals, among oats, wheat, maize, triticale, and rice (Miller and Rascochova, 1997). Aphids also spread plum pox, a severe disease of stone fruit trees that is easily spread from orchard to orchard (Damsteegt et al., 2007).
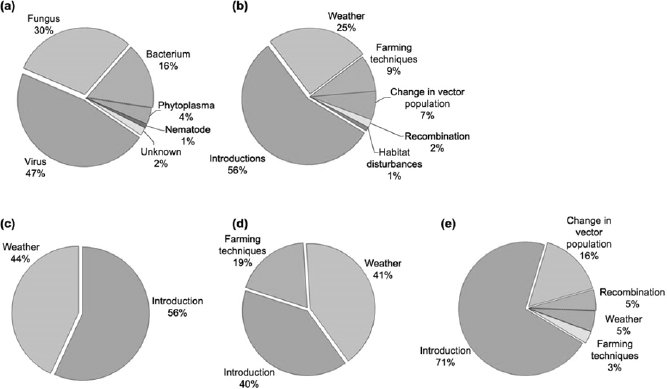
As winters become warmer in northern latitudes, more bacterial and fungal pathogens will likely survive through the winter, which may lead to more severe plant diseases, and increases in their geographic range. A shift in climate may also influence host resistance and growth, resulting in lowered resistance to fungal and viral diseases in plants (Harvell et al., 2002). Figure WO-2 illustrates major taxonomic pathogen groups causing emerging infectious disease in plants. According to Harvell et al. (2002), if climate change modifies host or pathogen geographic ranges, formerly separate species could converge, resulting in more severe disease outbreaks.
Ecology and Evolution
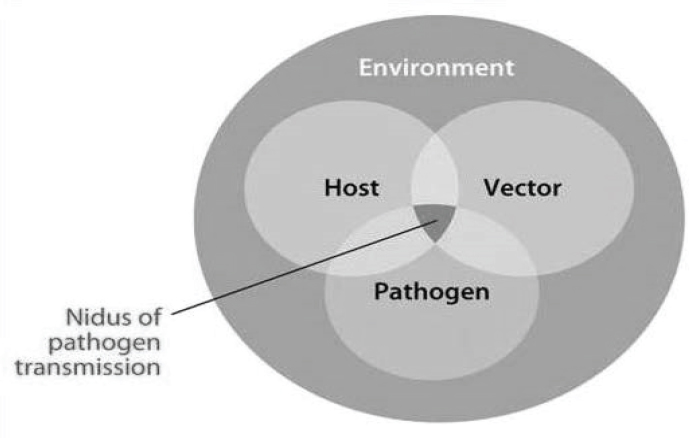
Vector-borne pathogen transmission occurs when host, vector, and pathogen interact in space and time within a permissive environment, as illustrated in Figure WO-3. Several environmental components (e.g., vegetation, climate, geology) may define the geographic area within which transmission takes place for a particular vector–host–pathogen system (Reisen, 2010).
Speaker Rodrigo Almeida (see Appendix A1), of the University of California, Berkeley, dissected the ecological complexity of vector-borne diseases into the following layers: the environment; the individual ecologies of pathogen, vector, and host; the outcome of their various interactions; and the effects of disease management. Local variation in the interplay among ecological forces shaping vector-borne diseases may produce dramatic shifts in disease transmission dynamics.
Similarly, several speakers described the effects of pathogen, vector, and host evolution on the transmission of West Nile viral fever, dengue, and chikungunya, among other vector-borne diseases.
SOURCE: Anderson et al., 2004. Reproduced with permission from Elsevier.
SOURCE: Reisen, 2010. Reproduced with permission of Annual Review of Entomology, Volume 55, © by Annual Reviews, http://www.annualreviews.org.
Epidemics and Emergence
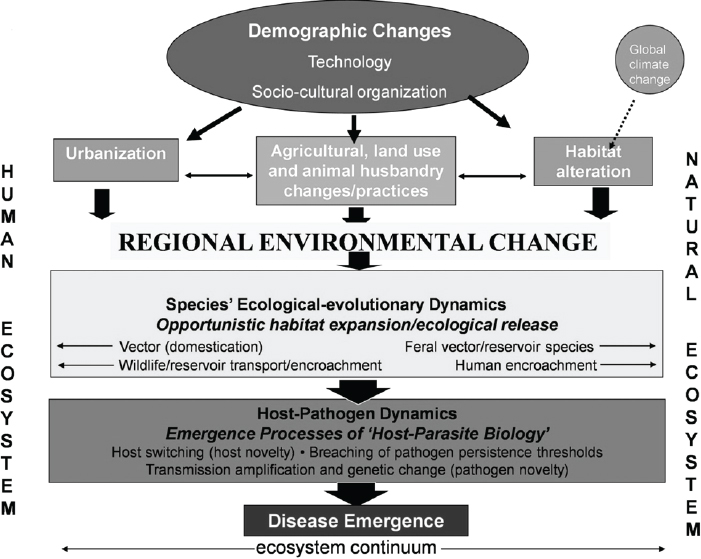
Emerging infectious diseases are caused by pathogens that (1) have increased in incidence, geographic, or host range (Funk et al., 2013); (2) have altered capabilities for pathogenesis; (3) have newly evolved; or (4) have been discovered or newly recognized (Anderson et al., 2004; Daszak et al., 2000; IOM, 1992). Recent epidemics of vector-borne disease have arisen from specific conditions occurring within the context of the large-scale drivers of infectious disease emergence listed in Box WO-1. Local surges in vector density, as well as increased vector competence—a measure of a given vector’s intrinsic capacity to be infected by a pathogen, to replicate it, and to transmit it—fuel outbreaks (see Kilpatrick and Randolph in Appendix A6). Epidemics have also arisen in naïve host populations, whose exposure to vector-borne diseases has increased with the globalization of travel and trade, and with the decline of vector control efforts.
For viruses such as the West Nile virus (WNV) and dengue virus (DENV) that have recently expanded their geographic range, increased transmission has driven selection for strains with greater epidemic potential, while increased gene flow among vector populations has been associated with higher viral transmission rates. Figure WO-4 depicts the confluence of multiple drivers of vector-borne disease emergence in humans, all of which were explored in detail in the forum’s initial workshop on vector-borne diseases (IOM, 2008).
Many of these same factors, in particular the global expansion of travel and trade, have driven the emergence of vector-borne plant diseases. Speaker Anna Whitfield, of Kansas State University, noted many similarities among vector-borne diseases of plants, animals, and humans, and in the health and research challenges
SOURCE: Adapted from Wilcox and Gubler (2005) with permission from the Japanese Society for Hygiene.
they present (see Appendix A12). She described several emerging vector-borne plant diseases that threaten U.S. agriculture and horticulture, including
- Citrus greening disease, also known as huanglongbing, is caused by the bacterium Candidatus Liberibacter asiaticus, which is transmitted by an invasive insect, the Asian citrus psyllid (Diaphorina citri). Since its first appearance in Florida in 1998, it has become a major threat to that state’s citrus crop, and it has spread across the southern continental United States to California, as well as to Hawaii and Mexico (University of California Agriculture and Natural Resources, 2013).
- An entirely new genus of viruses, Emaravirus, of which all known members are transmitted by eriophyid mites, infects a broad variety of plants including roses typically thought to be pest resistant, raspberries, pigeon-pea, and the European mountain ash (Mielke-Ehret and Muhlbach, 2012).
- The soybean vein necrosis virus, transmitted by thrips, was first reported in Arkansas and Tennessee in 2008. Soybean vein necrosis is now the most widespread viral disease of soybeans in the United States (Zhou and Tzanetakis, 2013).
Recent Developments
The vast and complex challenges identified in the forum’s 2007 workshop on vector-borne diseases continue to preoccupy researchers and policy makers (IOM, 2008). However, the field has undergone considerable change in the intervening years. In its fall 2014 workshop, the forum chose to highlight recent developments in the identification, emergence, and transmission of vector-borne diseases, as well as the public health response to vector-borne infections; advances in our understanding of the epidemiology and ecology of vector-borne diseases; and new insights on mitigating their effects. All of these topics, of course, raised further questions to be explored.
OVERVIEWS: VECTOR-BORNE DISEASE IN HUMANS, PLANTS, AND ANIMALS
The workshop opened with three presentations examining vector-borne disease systems that affect humans, plants, and animals. Speakers described dynamic interactions among pathogens, vectors, hosts, and their ecosystems, relating them to historic patterns of disease and patterns of emergence. Speakers also reviewed efforts to halt vector-borne diseases, considered possible future initiatives, and predicted possible future transmission patterns.
Emerging Human Mosquito- and Tick-Borne Diseases
Speaker Lyle Petersen from the Centers for Disease Control and Prevention (CDC) focused on mosquito- and tick-borne diseases that posed the greatest threat to the health of the U.S. population. Many of these pathogens are transmitted primarily among other animal species (reservoir hosts). For example, primates are the natural reservoir hosts of DENV (Bean et al., 2013) and chikungunya virus (CHIKV), and there is evidence that some animals, including nonprimates, such as rodents, birds, and small mammals, may also act as reservoirs for CHIKV (WHO, 2014b).
Vector-Borne Diseases of Concern in the United States
Humans are incidental hosts for most mosquito-borne viruses including WNV, as well as for tick-borne pathogens, which include Borrelia burgdorferi (the bacterial cause of Lyme disease), the recently discovered Heartland virus,7 and parasites of the genus Babesia (the agents of babesiosis). All such diseases
___________________
7 Heartland virus belongs to a family of viruses called Phleboviruses. Viruses in this family are found all over the world. Some of these viruses can cause people to get sick. Most of the phleboviruses that cause people to become ill are passed through the bite of a mosquito, tick, or sandfly. http://www.cdc.gov/ncezid/dvbd/heartland/ (accessed on October 5, 2015).
feature “incredibly complex” transmission cycles, and when herd immunity8 within the animal reservoirs becomes important in slowing transmission, it is often transient, Petersen observed. Mosquito-borne diseases that incidentally infect humans may amplify quickly in response to any of a wide spectrum of factors that are difficult to anticipate far in advance, he continued (see Petersen et al. in Appendix A9). By contrast, pathogens amplify gradually in the longer-lived tick, producing more predictable transmission patterns. Such “slow burn” epidemics tend to garner little public attention in comparison to dramatic, sporadic outbreaks of mosquito-borne illness, he pointed out.
WNV, which first emerged in the United States as a human pathogen in 1999, is a prototypic arbovirus for which people serve as incidental hosts, according to Petersen. “It had to be brought in [to this country] by man, probably by importation of an infected animal,” he said. “I think it is also important that it emerged during a heat wave,” he added, because heat has been shown to increase the transmission efficiency of WNV by increasing concentrations of WNV in vector mosquitoes and shortening the time between an infected blood meal and when they become infectious (extrinsic incubation period). Evolution has also played a significant role: the emergent strain featured a key mutation that increased transmissibility, which was further improved by a second mutation in a replacement strain that arose in 2002. In addition, he explained, “There is continued co-evolution both in birds [the reservoir host for WNV] and in the virus, where birds are becoming less susceptible to illness and death following infection, but at the same time, the viruses may be becoming more virulent by creating higher viremia in birds. So, in essence, it is an arms race between the host and the pathogen.”
To date, it is possible that more than five million WNV infections have occurred in the United States. The vast majority have been asymptomatic, Petersen stated, but a fraction of a percent of them has progressed to severe neuroinvasive disease, of which more than 17,000 cases have been reported. Many of these cases occurred during three major outbreaks, which took place in 2002, 2003, and 2012—all during heat waves, he added. As illustrated in Figure WO-5, certain geographic regions of the United States seem to be at higher ecological risk for West Nile viral disease compared to other regions of the United States. “When the virus came across the U.S., there [was] no way we could have predicted that South Dakota would become the highest-incidence state for a tropical virus,” he observed.
Some tick species serve as vectors for several different human diseases, while some pathogens can be spread by several different tick species. Petersen noted that ticks may be carriers of viruses, bacteria, and parasites that incidentally infect humans. Over the past century, land use changes, combined with
___________________
8 Herd immunity occurs when a sufficient percentage of a population is immune to a pathogen to prevent its transmission. The more efficiently a pathogen can spread between members of a “herd,” the greater the percentage that must acquire immunity to stop its transmission. Human immunity is inconsequential for pathogens that infect humans incidentally.
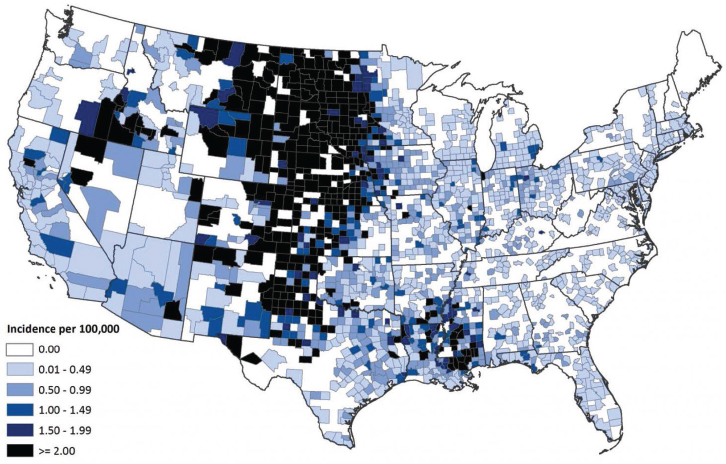
SOURCE: ArboNET, Arboviral Diseases Branch, CDC.
favorable environmental conditions, have enabled several tick species to increase in numbers and expand their geographic ranges, according to Petersen (see Figure WO-6). As a result, he observed, the incidence of essentially all tick-borne human diseases reported in the United States has increased. In addition, several novel tick-borne human pathogens have recently been identified, including relatives of known bacterial disease agents, as well as the Heartland virus, which Petersen characterized as a potential cause of hundreds to thousands of severely debilitating (and occasionally lethal) cases of illness per year (see later discussion in the section, “Changing Paradigms for Tick-Borne Diseases in the Americas”).
Humans serve as primary hosts for the mosquito-borne dengue and chikungunya viruses, both of which are on the rise in the Americas. According to the CDC, dengue is caused by any one of four related viruses transmitted by mosquitoes (CDC, 2015). Infection with any of the four Flaviviruses can cause a painful febrile illness, dengue fever, or the life-threatening dengue hemorrhagic fever. Dengue virus now infects about 400 million people each year, having resurged after DDT-based vector control efforts were halted in the 1970s, according to Petersen. Dengue’s expansion has also been abetted by the introduction of an additional vector species, Aedes albopictus, from Asia to the United States in 1985 (in a shipment of used tires). Before this introduction 30 years ago, dengue’s geographic “footprint” was limited to a more range-restricted vector, Aedes
aegypti, in the Americas. Figure WO-7 illustrates the dramatic increase of the incidence of dengue in the Americas, which accelerated after reaching an apparent turning point around 2000. “The ecological factors all sort of aligned. The creation of megacities in the tropical world and all of the problems that trended with them have suddenly caused this incidence of dengue to go up and up and up,” he observed.
Today, in tropical locations such as Puerto Rico, over 90 percent of residents have already been infected by DENV, Petersen stated. Can dengue fever—once a common illness in the southern United States—reemerge in this country? There is certainly reason to worry that it might, he observed. Ae. aegypti is endemic in the South and has expanded its geographic range to new areas, such as California, while Ae. albopictus has spread throughout the East. At the same time, increasing numbers of dengue-infected travelers are entering the United States. Yet, since 2009, only 10 dengue outbreaks (8 in Texas and 2 in Florida) have occurred in the United States, and each involved limited numbers of cases within restricted areas. “The U.S.–Mexico border is like the Berlin Wall of dengue,” he quipped. “You
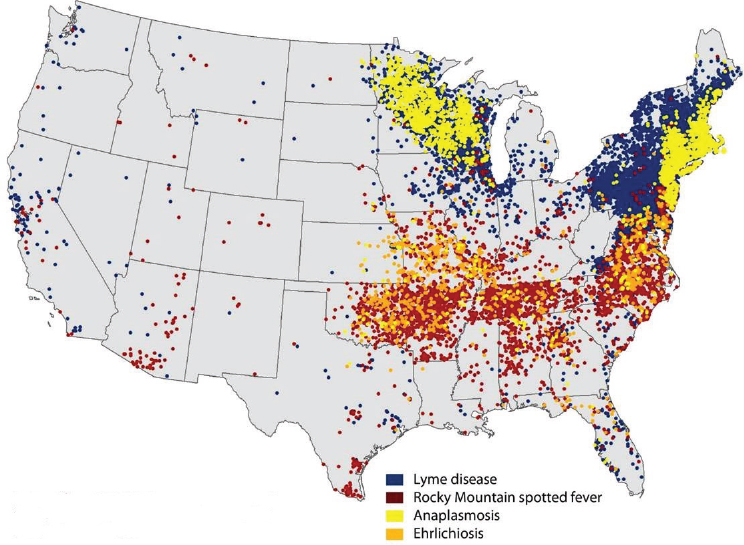
NOTE: In 2012, no cases of tick-borne illness were reported from Hawaii. Alaska reported 10 travel-related cases of Lyme disease.
SOURCE: CDC.
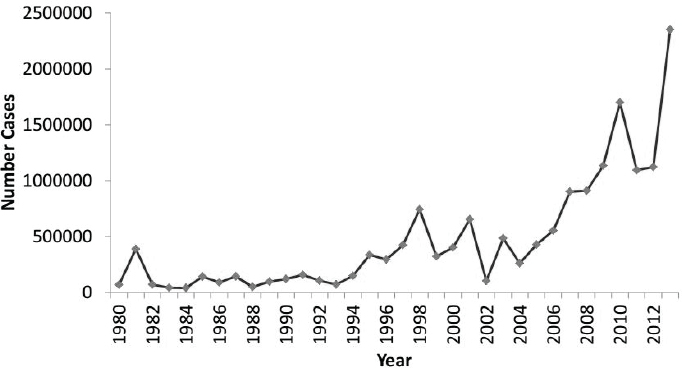
SOURCES: As presented by Lyle Petersen on September 16, 2014. Data from Pan American Health Organization.
find these huge outbreaks on the Mexican side of the border, just right across the Rio Grande River.” His group’s investigation of this paradox revealed vastly different human behaviors and environments in adjacent towns on either side of the border, and suggested that the lack of air conditioning and more crowded living conditions in Matamoros, Mexico, resulted in much higher rates of dengue transmission in comparison to Brownsville, Texas (Ramos et al., 2008). For now, it appears, lifestyle and living conditions help to protect the United States from dengue becoming endemic.
CHIKV is an Alphavirus that, like DENV, is transmitted between humans by both Ae. aegypti (its traditional urban vector) and Ae. albopictus, Petersen stated. Yet, unlike dengue, chikungunya infection is usually symptomatic, causing fever, debilitating joint pain, and often a rash. The virus emerged in the Americas in late 2013, on the island of St. Martin, and quickly spread across the Caribbean, as illustrated in Figure WO-8.
In May 2014, the Caribbean Public Health Authority declared that chikungunya had reached epidemic status (Carribean 360, 2014). By September 2014, more than 700,000 cases had been reported to the Pan American Health Organization (PAHO), with 113 deaths. As with dengue, these included only a few isolated cases of locally acquired chikungunya in the contiguous United States, all of them in Florida.
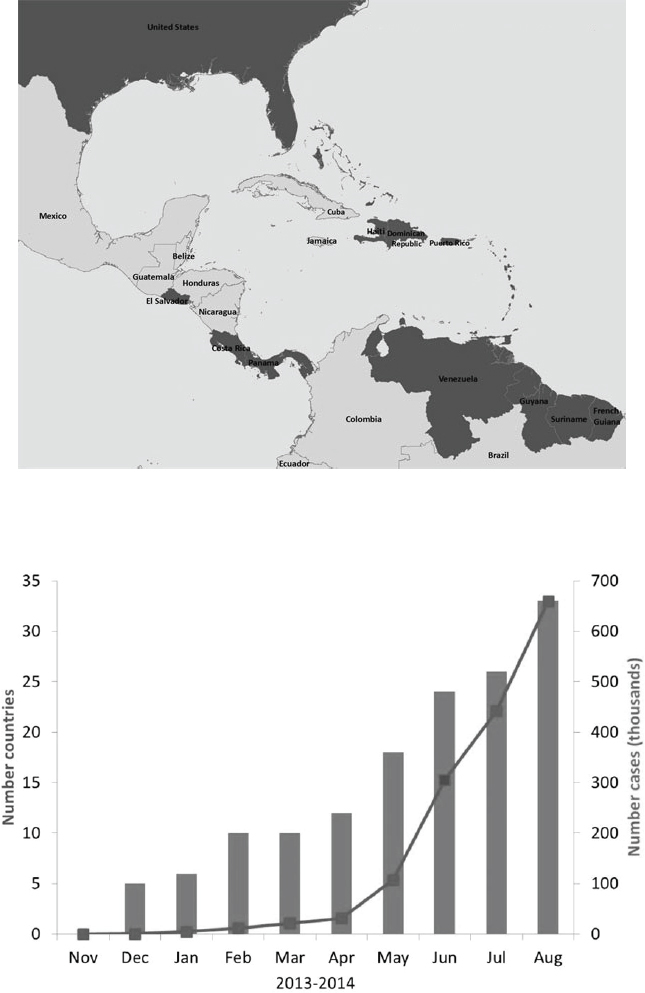
SOURCES: Top, CDC; bottom, as presented by Lyle Petersen on September 16, 2014. Data from Pan American Health Organization.
U.S. Outlook and Preparedness
Petersen offered the following general predictions of vector-borne disease activity in the United States and its territories:
- Continued focal and regional outbreaks of West Nile viral disease;
- Significant public health effects of dengue (ongoing) and chikungunya (until herd immunity is established) in U.S. territories;
- Increased incidence and distribution of tick-borne diseases;
- Discovery of additional novel tick-borne pathogens, some of public health significance; and
- Importation and emergence of nonendemic pathogens, such as the Zika virus.9
Are we prepared to meet these challenges? “In some instances, I think the glass is reasonably full,” Petersen concluded. The ArboNET surveillance system, developed to track WNV, is the only system in the world that simultaneously collects human, animal, and vector data, he said. It can and has—in about a dozen cases—been adapted to monitor additional emergent and endemic arboviruses. Advanced molecular detection systems have proved extremely valuable in detecting imported and novel vector-borne pathogens. Another important legacy of the response to WNV is greatly improved communication capacity among physicians, public health agencies, and medical centers regarding actual or potential vector-borne disease outbreaks, he added. On the other hand, he warned that the existing system for tick-borne disease surveillance is becoming overwhelmed by the rising numbers of cases. More broadly, he observed, ecological parameters of pathogen transmission remain largely unknown, which limits the usefulness of disease models—as does the fact that many communities are not prepared to respond to vector-borne disease threats owing to inadequate surveillance and/or mosquito control capacity.
For prevention and treatment, “The glass is only half full,” Petersen noted. While screening has—at great expense—nearly eliminated the risk of acquiring blood-borne WNV or Trypanosoma cruzi (the protozoan agent of Chagas disease), the U.S. blood transfusion system cannot currently detect the pathogens that cause dengue, babesiosis, chikungunya, ehrlichiosis, or anaplasmosis, or the next novel or imported vector-borne pathogen, Petersen pointed out (see “Blood Donation Screening for Vector-Borne Diseases”). Effective treatment regimens are available for the vector-borne bacterial diseases, but these conditions often go unrecognized, undiagnosed, or improperly treated, he observed. No such
___________________
9 Zika virus is a Flavivirus related to yellow fever, dengue, West Nile, and Japanese encephalitis viruses. In 2007, it caused an outbreak of relatively mild disease characterized by rash, arthralgia, and conjunctivitis on Yap Island in the southwestern Pacific Ocean. This was the first time that Zika virus was detected outside of Africa and Asia (Hayes, 2009).
therapeutics exist for viruses, and while promising vaccines are in development against DENV and WNV—and also against Borrelia burgdorferi, the bacterial agent of Lyme disease—it is unclear when and if these will become commercially available, in part, because they may lack a robust commercial domestic market (see “Outlook for West Nile and Chikungunya Vaccines”).
Much the same can be said about promising pesticides in development for vector control, Petersen stated. Of particular concern, he noted that no effective, scalable, vector control method exists for Ixodes scapuarlis or Ae. aegypti, each of which represents a major threat to public health throughout the Americas.
Lessons from a Model Plant Disease
As previously alluded to, after describing the ecological “layers” within which vector-borne plant diseases occur and the multiple factors that influence their transmission dynamics (see Figure WO-1), Almeida used the example of the vector-borne bacterium Xylella fastidiosa to illustrate these concepts (see Appendix A1). Typically a benign colonist of more than 300 species of plant species, X. fastidiosa is transmitted by insects that feed on the liquids transported within the xylem of host plants.10 The bacterium can, under certain circumstances, grow so profusely that it blocks the upward flow of fluids in the plant, resulting in scorched leaves and shriveled fruit. This condition affects several important woody crop plants and trees, most notably grapevines in the United States (called Pierce’s disease of grapevines). Pierce’s disease escalated from a low-level problem into a major threat to California’s viticultural industries following the arrival of an alternate vector—the glassy-winged sharpshooter (Homalodisca vitripennis)—to California in the late 1980s (Fletcher and Wayadande, 2002). Since then, the geographic and host plant range—and the economic, political, and social significance—of X. fastidiosa have expanded in the United States and internationally.
The emergence of Pierce’s disease has paralleled significant changes in the study of vector-borne plant diseases, as well as in their dynamics, Almeida pointed out. Around the time that the glassy-winged sharpshooter invaded California, plant pathologists relied on studies of pathogen host range, epidemiological surveys, and the outcomes of vector control measures attempted in the field to inform mitigation efforts. These ecologically based methods were sufficient to manage diseases within a limited geographic area and time frame—a sensible approach, prior to the global transmission of economically important diseases, he said. Similarly, research priorities were short to medium term and were directed toward managing disease, not toward understanding patterns of transmission or factors of emergence.
___________________
10 Xylem is the conductive tissue in vascular plants through which water and nutrients flow upward from the roots.
“The present is a little more complicated,” Almeida continued. Today, significant vector-borne plant disease threats frequently involve pathogens and/or vectors introduced to new ecosystems via international travel and trade, and sequence-based identification methods allow researchers to trace the origins of outbreaks. By these means, researchers determined that pathogenic subspecies of X. fastidiosa occur with a distinct range within the Americas, and specific subspecies now threaten olives and other important crops in Europe, as well as grapes and the Asian pear in Taiwan.
Before it was identified in southern Italy for the first time in October 2013, X. fastidiosa was not believed to be able to cause disease in olives, Almeida noted. Given the olives’ economic and cultural significance in this region (as both a source of oil and a tourism attraction), as well as the potential for pathogen spillover to grapes and citrus, it is perhaps no wonder that the European agricultural community is extremely concerned about it. Plant scientists are often not trained to handle such situations where the disease has large economic, trade, and social consequences at the international sphere; in this case it also included the generation of conspiracy theories suggesting that Almeida himself had spread the disease to Italy, as an agent of developers or agribusiness, or that X. fastidiosa itself does not exist. “It is really an interesting problem that goes way beyond a plant disease and how to manage it.”
How do plant diseases actually move to new ecosystems? In many cases it occurs on ships, according to Almeida, as contrasted to their human counterparts, which move readily by air travel. Current data strongly suggest that the epidemic in Italy originated from ornamental coffee plants imported from Costa Rica. Pest and diseases also travel with “suitcase plant material,” such as the anecdotal case of a grapevine cutting of Israeli origin smuggled into California by a grower (which is supported by molecular data), along with what has become a major insect pest, he noted. Vector introductions provide a second important path to plant disease outbreaks, if they increase transmission of an existing pathogen. This occurred in California when the glassy-winged sharpshooter quickly achieved large populations, which permitted more frequent encounters with X. fastidiosa, more successful infections of various plant species, and, ultimately, higher incidence of disease, he explained. New associations between the novel vector and X. fastidiosa, coupled with the vector’s ability to transmit additional pathogen strains among a broader range of host plants, may also have contributed to the recent emergence of Pierce’s disease and other X. fastidiosa diseases in California, he suggested.
In addition, complex insect–pathogen–plant interactions must be understood at the molecular level in order to address vector-borne plant disease threats. Almeida observed that plant pathogen transmission hinges on complex interactions among surface proteins and receptors in pathogens and vectors. Having identified some of such moieties in X. fastidiosa, Almeida and coworkers are attempting to inhibit bacterial attachment to insect tissues, which might be achieved by
genetically modifying host plants to produce molecules that block key interactions. This approach has recently been shown to work for an insect-transmitted plant virus (Whitfield and Rotenberg, 2015, reprinted in Appendix A12). Host plants could also be genetically manipulated to produce molecules that kill specific insect vectors that feed on them, he added.
Another active area of current research examines how, over the course of evolution, pathogens manipulate vector behavior directly, or through their effects on host plants, so as to increase transmission efficiency. Many such interactions have been identified, employing a broad range of mechanisms and strategies, Almeida reported.
Vector-Borne Disease in Animals
“We are just barely getting a grasp on patterns of vector-borne diseases in animals,” observed speaker William Karesh, of the EcoHealth Alliance, as he introduced this topic. However, he continued, the importance of vector-borne animal diseases is increasingly apparent. Vector-borne animal pathogens are included in priority pathogen categories by the National Institute of Allergy and Infectious Diseases, in the catalog of significant trade-related animal diseases by the World Organisation for Animal Health (OIE), and among novel pathogens listed by the United States Agency for International Development’s PREDICT project, he reported. Nearly one-third of all known viruses that infect mammals are vector borne, and a recent analysis of 86 emerging zoonotic viruses determined that, among those transmitted from wild animals to humans, 40 percent were vector borne, including all viruses for which wild birds served as reservoir species.
Recent studies of transmission patterns reveal potential strategies for addressing vector-borne animal diseases, Karesh noted (see Appendix A5). For example, researchers have shown that while vector-borne mammalian viruses tend to have a broad host range, they are generally transmitted among these hosts by a single vector. In some cases where that is true, he suggested, controlling vector populations and their ability to spread pathogens to humans or animals offers relatively simple routes to reducing disease transmission, as compared with pathogens that follow multiple transmission routes between animals and people, and are therefore more prone to spillover. The identification of land use change and international travel and trade as primary drivers of emergence of vector-borne animal diseases should guide disease surveillance and prevention efforts, he added.
Disease Patterns in Emerging Pathogens
Karesh described the state of knowledge regarding patterns of disease for the notable emerging vector-borne animal pathogens in the following subsections.
Schmallenberg virus emerged suddenly in sheep in Germany in 2011—related viruses had previously been identified only in Africa, the Middle East, and Asia—and from there spread across Europe much as the West Nile virus moved across the United States upon its emergence in 1999, Karesh recalled. The biting midge (Culicoides species), considered the major vector of Schmallenberg virus, is “extremely efficient in transmitting the virus to sheep,” he said. However, he added, “Sheep are not so efficient in transmitting the virus back into midges.” Nevertheless, the virus is highly contagious, with each infected animal producing as many as seven infections (European Food Safety Authority, 2014). These infections—which last a few days and cause fever, reduced milk yield, diarrhea, and abortion—have primarily been reported among ruminants (none have been reported in humans).
Schmallenberg virus infection is not listed as a reportable disease by the OIE, Karesh noted. “Currently, the disease does not meet the criteria for OIE listing,” he explained. “If an animal is positive it means they are protected for life. If they are negative it means they are free of the disease.” On the other hand, he continued, the disease can be disastrous for individual farmers, who operate on small economic margins. Vector control might seem a reasonable way to reduce disease transmission, but it would not be an easy route to take, given our limited understanding of the ecology of the widespread midge, he observed. Vaccines are available but have not been widely used, he remarked, perhaps because the transient infection is not perceived as sufficiently burdensome to warrant prevention.
WNV, as Petersen noted, infects a wide range of bird species. Elucidating the resulting patterns of disease has been difficult, according to Karesh, because “there are so many variables at play.” While climate and weather likely influence disease incidence, many additional factors vary across the geographic range of the virus, he noted (Crowder et al., 2013). The number of avian WNV cases reported to the CDC has varied widely from year to year, he stated (Lindsey et al., 2014). Some of this variation may reflect the 60 percent decline in dead bird surveillance that occurred between 2004 and 2012, and which occurred simultaneously with a significant decrease of pathogen surveillance in trapped mosquitoes, Karesh noted (Hadler et al., 2014). Thus, “It is very hard to say whether we are having a changing pattern when we are changing the tools we are using to monitor patterns . . . [and] we are investing less in surveillance,” he concluded.
Tick-borne pathogens threaten animal and human health worldwide. Ten percent of tick species carry such pathogens, Karesh reported (Jongejan and Uilenberg, 2004). The resulting diseases affect 80 percent of the world’s cattle, at a cost of up to $19 billion per year, which is borne disproportionately by resource-limited countries in the tropics and subtropics (Minjauw and McLeod, 2003).
Once again, vector control is not a likely solution to this problem, Karesh observed. Effective vector control to address tick-borne disease in livestock
would need to extend to wild animals that are part of transmission cycles, making it prohibitively expensive, particularly for people earning less than 1USD per day, which is common in India and Africa (Minjauw and McLeod, 2003). In such circumstances, it can cost more to control disease in cattle than it does to raise cows, he pointed out.
Multidisciplinary Research on Rift Valley Fever Virus
“We can’t really lump the vector-borne diseases together and say they are all headed in the same direction,” Karesh concluded (Kilpatrick and Randolph, 2012, reprinted in Appendix A6). Pathogen introductions, ecological shifts, and changes in host immunity all affect patterns of disease, he noted. Thus, rather than tackle vector-borne diseases as a whole, he proposed that researchers undertake multidisciplinary, long-term, broad-based studies of individual vector-borne diseases.
The EcoHealth Alliance and a large group of collaborating agencies are currently attempting such a study of Rift Valley fever11 in South Africa. There, according to Karesh, rainfall patterns are predictable as much as 3 months in advance. This would in theory provide adequate warning to vaccinate animals against the spread of Rift Valley fever virus (RVFV). But, according to Karesh, partly because of sociological reasons that may be difficult to counteract, this has not happened. Therefore, it may be equally important to develop a targeted, effective approach to disease control that will enlist the support of people and governments. A better strategy against Rift Valley fever in South Africa might take advantage of herd immunity, which also appears to influence outbreak patterns there, he observed.
To investigate this possibility in detail, Karesh and coworkers have embarked on a plan to monitor changes in immunity to RVFV in individual animals, flocks, herds, mixed-species populations, and mixed populations of wildlife and domestic animals within a 40,000 km2 area to identify factors that influence immunity at a population scale. The project, which began around the time of the workshop, is expected to last 5 years, he said. It will track antibody levels in local domestic animals (including cattle, goats, and sheep), free-ranging wildlife (including several antelope species) and those on game ranches, mosquitoes, and people. Those measurements will be integrated with data on vegetation and weather, in order to accomplish the following series of objectives:
___________________
11 Rift Valley fever is a viral zoonosis that primarily affects animals but also has the capacity to infect humans. Infection can cause severe disease in both animals and humans. The disease also results in significant economic losses due to death and abortion among RVF-infected livestock. The virus was first identified in 1931 after an epidemic struck sheep on a farm in the Rift Valley of Kenya. Since then, outbreaks have been reported in sub-Saharan and North Africa. In 1997–1998, a major outbreak occurred in Kenya, Somalia, and Tanzania, and in September 2000, cases were confirmed in Saudi Arabia and Yemen, marking the first reported occurrence of the disease outside the African continent and raising concerns that it could extend to other parts of Asia and Europe. Source: http://www.who.int/mediacentre/factsheets/fs207/en (accessed March 25, 2016).
- To compare how immunity to RVFV changes over time in vaccinated and unvaccinated sheep and antelope;
- To determine herd immunity in wildlife and domestic animals;
- To investigate the ecology of RVFV and its mosquito vector, with reference to soil types, vegetation, and climate; and
- To evaluate human behavioral practices and measure immunity among people working on farms within the study area and detect new infections.
By collecting data at the individual, population, and meta-population levels, among both domesticated and wild animals, the researchers expect to discover useful distinctions in herd immunity to RVFV among different populations. Karesh noted, for example, that herd immunity in cattle kept by pastoral farmers—which tend to live longer than those raised in commercial feedlot systems—is likely to be more persistent. Likewise, he said, long-lived wildlife such as buffalo might also have high immunity to RVFV. Such information should allow these investigators to better anticipate when particular animal populations are susceptible to outbreaks of disease that could, in turn, spill over into humans—knowledge that would support more efficient vaccination programs or other disease control measures, he concluded. Implementation will require social engagement which, he said, means “making sure [the program] . . . is cost-effective, convincing people that [it] is worth doing, and really understanding where they are coming from so we can come up with some solutions that make sense.”
Common Ground
In her introduction to this workshop session, moderator Mary Wilson of the Harvard School of Public Health (now at the University of California, San Francisco), encouraged participants to recognize commonalities among the pathogen–vector–host–environment systems described by the three speakers, and to consider research and policy issues that lie at these points of intersection. In the course of their presentations, the speakers raised several such ideas that were further explored in discussion immediately afterward, and throughout the workshop.
Need for Consistent, Comprehensive Surveillance
Both Petersen and Karesh noted that initially robust support for WNV surveillance has declined with disease incidence following the 2002–2003 outbreaks. Limited resources were available to respond to another peak outbreak in 2012. While it is now possible to make accurate local predictions of WNV outbreaks in time to prevent their occurrence, Petersen said, communities are not investing in local surveillance, nor are they willing to implement vector control measures in advance of human cases of the disease.
“By the time an effective response is mounted at a local level, the outbreak is often well on its way and possibly on the downhill slope,” Petersen stated. “That is exactly what we saw in the big outbreak in Dallas. By the time a widespread response was mounted, three-quarters of the cases had already occurred. We were able to show quite nicely that it did stop the outbreak, but it was done too late. So there is really a problem with the intensity of surveillance and getting people to actually respond effectively in a timely way.” Speaker James Hadler, of Yale University, further explored this dilemma (see subsequent section, “Loss of Arbovirus Disease Surveillance Capacity in the United States”).
Surveillance for most other vector-borne pathogens—including novel, emerging ones—is less thorough than for WNV. Vector-borne plant diseases pose an especially difficult problem for surveillance, according to several participants. Until a plant disease becomes epidemic, it is perceived as a problem only to the farmers whose crops are infected, Almeida noted. “For a plant disease to come to the radar you need thousands if not hundreds of thousands of plants to be sick,” he said.
On the other hand, forum member David Rizzo of the University of California, Davis, noted that many plant pathogens are emerging as the result of ecosystem disturbance, much as are animal pathogens. “I can think of a half a dozen in the United States right now with the potential to spill over into agriculture,” he stated, such as laurel wilt, which now threatens avocado crops. “By the time you see millions of dead plants, then it is too late to really do something,” he warned. Unfortunately, he added, little such surveillance is occurring, especially in natural ecosystems.
Gaps in Training and Their Consequences
All three speakers and several discussants expressed concerns regarding the limited opportunities for the education and training of the next generation of vector-borne disease researchers, coupled with the decline of certain key disciplines. Training of scientists working with vector-borne diseases in plant pathology fails to emphasize either quantitative work or field ecology, Almeida noted. Petersen similarly criticized the public health community for its neglect of research and training in ecology—as well as medical entomology—and warned that insufficient funding and career opportunities for scientists in these disciplines would diminish future capacity to address vector-borne disease threats. In a later presentation, Christopher Paddock, of the CDC, added cartography to the list of endangered core disciplines for vector-borne disease research. When mosquito vectors of emerging pathogens cannot readily be identified by species owing to a lack of skilled medical entomologists, Karesh observed, “it is going to be an ugly world.”
Today’s investigators also lack the ability to connect the study of vector-borne diseases—and emerging diseases in general—with their social, political,
and economical consequences. “I think there will be a push for incorporating social sciences into what we do,” Almeida predicted, and offered as an example the work of a postdoctoral researcher in his laboratory (Dr. Adam Zeilinger), shown in Figure WO-9.
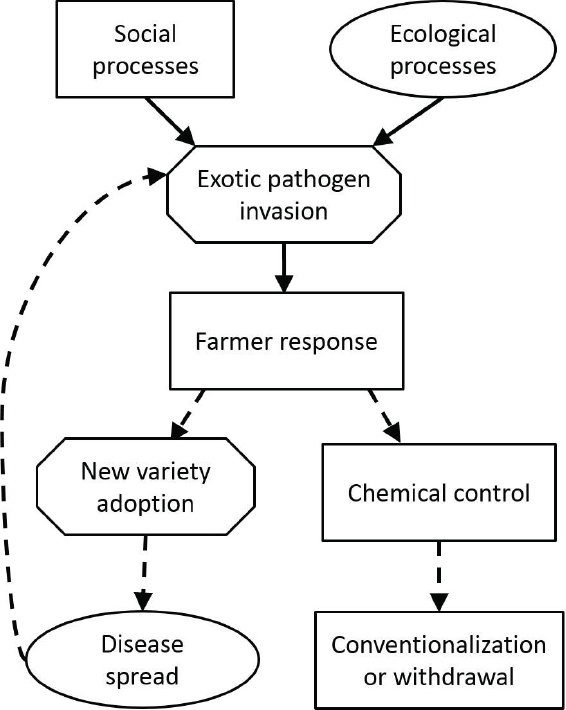
SOURCE: Figure created by Dr. Adam Zeilinger, University of California, Berkeley. Reproduced with permission.
Advantages of the One Health Approach
The narrow training of investigators both reflects and drives a specialized, technology-centric approach to the study of vector-borne diseases. Speakers and participants alike noted that as the field has become increasingly focused on molecular-level interactions between pathogen, host, and vector, it has lost sight of the ecological contexts of these interactions, as well the many benefits of interdisciplinarity.
In plant pathology, Almeida observed, basic questions are being overlooked. For example, in the case of citrus greening—a disease that threatens to decimate Florida’s citrus industry—some of the most fundamental experiments have yet to be performed. If “we still don’t know how long it takes for a plant to express symptoms once it is infected by an insect,” inferences from epidemiological models will suffer from lack of information, according to Almeida.
Moreover, solutions to specific disease problems are being studied in isolation and in ignorance of the drivers of disease emergence, which might productively be addressed interdisciplinarily. “Is plant pathology, as well as other disciplines, willing to diversify and view agricultural systems in a more holistic manner?” Almeida wondered. “That is not clear,” because there is limited research of that kind currently going on—nor is it at all clear, he observed, whether or how funding agencies would support such an approach to transdisciplinary research.
Almeida, along with several other workshop participants, decried a funding bias toward research in molecular biology in general, and specifically in support of the use of transgenics to address vector-borne disease. In addition, lack of funding has made programs highly competitive. “USDA funding rates for fundamental science [i.e., single Principal Investigator grants] are currently less than 10 percent,” Almeida observed. He also noted, however, that the National Science Foundation recently revived the dwindling science of systematics through a targeted training grant program. Petersen described a similar effort by the CDC to boost medical entomology following the emergence of WNV in the early 2000s that has since been discontinued.
Increasing recognition of the value of the One Health paradigm—defined as “the collaborative effort of multiple disciplines—working locally, nationally, and globally—to attain optimal health for people, animals, and the environment” (AVMA, 2008)—offers hope that these trends may be reversed. Forum member Kevin Russell of the Armed Forces Health Surveillance Center pointed out that the Global Health Security Agenda emphasized a multisectorial approach to global health security, which depends in part upon the economic ramifications of plant, animal, and human diseases. Karesh noted that the Department of Defense is funding the described long-term, broad-based RVFV project he discussed in his presentation, which aligns with Global Health Security Agenda mandates for a One Health approach to disease control encompassing humans, wildlife, and livestock. “I think there are some cost-savings and efficiencies when we start to
pull together . . . thinking on animal and human and plant vector-borne diseases,” Karesh predicted. “We have to really start interacting more at that level and using that to leverage reducing budgets.”
Ecological Complexity
Underpinning the concept of One Health and influencing patterns of disease is an understanding of the essential role of the environment to address vector-borne diseases, both individually and collectively. Petersen observed, for example, that WNV outbreaks cannot be predicted beyond the local scale because “the ecology is incredibly complicated. It varies from place to place.” Some common denominators, such as heat waves, are generally predictive of outbreaks, he continued, but ultimately, “If you start narrowing down the ecology, Phoenix is not like Chicago.” Even within parts of Phoenix—which, in the middle of the desert, is an unlikely but raging hot spot for WNV—there is variation in transmission of the virus, he added. “The ecology of the whole United States is amazingly complicated and varied,” Petersen observed. This point was illustrated again and again over the course of the workshop, through the lens of various vector-borne diseases in natural, agricultural, urban, and suburban contexts.
CHANGING DISEASE PATTERNS
Four speakers illustrated the dynamic nature of vector-borne diseases in presentations describing the evolution and epidemiology of dengue and chikungunya, shifting patterns of insect-borne parasitic infections closely associated with poverty, leishmaniasis and Chagas disease, and the recent range expansion of multiple tick-borne diseases in the United States.
Arbovirus Evolution in Humans and Mosquitoes
Speaker Rebecca Rico-Hesse, of Baylor College of Medicine, used the examples of DENV and CHIKV to illustrate how evolution influences dynamic relationships among pathogens, vectors, and hosts (see Appendix A10). Consisting of little more than a strand of RNA that encodes 8 to 10 proteins, these highly mutable and adaptable arboviruses represent “the smallest, most simple organisms that we know of that have changed history of humankind,” she pointed out. DENV and CHIKV are transmitted to humans by the mosquito vectors Ae. aegypti and Ae. albopictus, which have quite distinct habitats and biting habits.
Global Spread of a More Virulent Dengue Virus
Most human dengue infections produce the flulike illness known as dengue fever, but some cases progress to the life-threatening dengue hemorrhagic fever
(DHF), with massive internal bleeding. Any of the four serotypes of DENV can cause dengue fever, and it has been known for decades that the risk of DHF increases if a person is serially infected by two different viral serotypes (Rico-Hesse, 2009). The majority of dengue epidemics to date, and most cases of DHF, have been linked to serotype 2 (DENV-2), which has been isolated in Asia, Africa, and the Americas, Rico-Hesse reported (Cologna et al., 2005). A single genotype within DENV-2, once limited to Southeast Asia, has been detected in the majority of isolates from patients with DHF—including, in recent years, patients in the Americas and West Africa, as well as in Southeast Asia. “This genotype, the one that is more severe and more virulent, has displaced all of the other ones,” she concluded. A similar displacement of less virulent genotypes worldwide by a more virulent virus has occurred within the DENV-3 serotype as well, she added.
How did the Southeast Asian genotype of DENV-2 outcompete native genotypes? Comparative infection experiments in cultured human dendritic cells—the cell type targeted by DENV—reveal that while the American genotype infects a larger number of cells, the Southeast Asian virus replicates much more efficiently, generating a larger number of viruses per cell infected, according to Rico-Hesse. Experiments in mosquitoes showed that both genotypes were equally capable of binding the insect’s midgut, but that the Southeast Asian virus was more prevalent in the salivary glands, and therefore more available for transmission to humans, she added. Both features contribute to the 60-fold-higher transmission efficiency of the Southeast Asian genotype, which in turn explains its ability to displace the American genotype, she concluded.
To investigate the source of heightened virulence in the Southeast Asian genotype of DENV-2, Rico-Hesse and colleagues created “humanized” mice, in which up to 80 percent of their white blood cells are of human origin (Brehm et al., 2013). “What we have is a mouse that gets infected, gets viremic and gets a rash, gets thrombocytopenic, and does all of the things just like humans do with dengue fever,” she explained. By infecting these mice, via mosquito bite, with selected viruses, the researchers were able to determine that the Southeast Asian genotype remained longer in the bloodstream and achieved higher titers than other genotypes. Subsequent theoretical studies of nucleotide folding among genotypes of DENV-2 suggest that subtle structural differences may determine virulence, she said; they plan to test these ideas in experiments with chimeric viruses.
The researchers also discovered that infecting via mosquito, rather than by inoculation, significantly increased DENV viremia in humanized mice, Rico-Hesse reported (Cox et al., 2012). This, she said, likely resulted from immune deficiencies that limit the mice’s ability to respond to infection as a human would. Interestingly, the humanized mice also made antibodies to mosquito saliva alone, and these, too, were very long lived. “We can’t say that mosquito saliva is not important,” she insisted. “We have to start including this in any studies
of pathogenesis, control of vaccination strategies. The mosquito saliva is doing things very unexpectedly in the human immune system.”
Chikungunya Expansion and Adaptation
Urban epidemics of chikungunya are characterized by rapid spread and high infection rates, leading in most cases to symptoms that resemble those of dengue: acute fever and debilitating joint pain (Nasci, 2014). These typically resolve within a week; however, joint pain and fatigue may persist for 2 years or longer in some individuals. In contrast to DENV, CHIKV infects endothelial cells and fibroblasts, but how it interacts with the human immune system and the mechanisms by which pathogenesis manifests remain a mystery, according to Rico-Hesse. She and coworkers plan to investigate these processes—as well as dengue pathogenesis—with a recently developed mouse model that better mimics the human immune system.
CHIKV was first identified in Tanzania in 1952, and subsequently found throughout Africa and Asia, where it caused periodic small outbreaks (Nasci, 2014). In June 2004, a chikungunya epidemic on Lamu Island, Kenya, spread to other islands in the Indian Ocean. This epidemic produced nearly half a million cases of chikungunya. A later epidemic resulted in more than 1.5 million cases in India, which then continued on through Southeast Asia to islands in the Pacific Ocean. Since then, CHIKV has been recognized as an important emerging vector-borne pathogen. The first locally transmitted cases of chikungunya in the Western Hemisphere were reported in October 2013, on the island of St. Martin; others have since been reported on several more islands in the Caribbean.
The recent range expansion of CHIKV resulted from separate advances by two of the three known viral genotypes, Rico-Hesse explained (Thiberville et al., 2013). The East Central South African genotype of CHIKV, which caused the first wave of epidemics, has not yet reached the Americas, but the Asian genotype has, she said. Recent evidence shows that the Asian genotype can be transmitted with equal efficiency by both mosquito vectors, one of which—Aedes albopictus—is well adapted to temperate climates (Vega-Rua et al., 2014). She concluded by observing that, “We [now] have a chikungunya virus that can be spread in Aedes albopictus—which, by the way, is everywhere in Houston.”
Dengue and Chikungunya in the Americas
Building on Petersen’s update on dengue and chikungunya emergence, Harold Margolis, of the CDC, described how rapid increases in their incidence in the Americas has prompted changes in diagnostic methods and protocols for all febrile disease syndromes.
Dengue
Since around 2000, as Petersen noted, dengue case reports have risen rapidly in the Americas. Margolis pointed out, however, that when considered on a country-by-country basis, this trend has been far from uniform. Rather, he observed, case rates have increased sharply in countries most affected by urbanization and migration. “Part of this increase is how we are recognizing the disease and what we are measuring and what we are diagnosing,” he suggested.
The symptoms of dengue fever can resolve within a week, or during the same period it can progress to severe hemorrhagic disease or death, Margolis noted. Many of dengue’s symptoms resemble those of several other febrile diseases, such as leptospirosis. In Puerto Rico, for instance, the diagnostic testing of patients meeting WHO criteria for suspect dengue typically finds that only about half of them are actually infected with the virus, he reported. On the other hand, researchers in Thailand determined that among a group of nearly 400 schoolchildren who tested positive for DENV infection, only half exhibited symptoms that met WHO criteria for clinical diagnosis (Sabchareon et al., 2012). Clearly, he concluded, “The only way you know if somebody has dengue is diagnostic testing.”
Margolis reported that major changes in dengue diagnostics in recent years are improving this situation. Once a slow and complicated process requiring both acute and convalescent samples for immunoglobins (IgM) testing, diagnosis by specific DENV subtype or by IgM can now be performed quickly on a single acute-phase sample through rRT-PCR, he explained. It is now possible to detect about 90 percent of cases that will seroconvert through molecular diagnostic testing. In the United States, where most molecular diagnostic tests like these are performed commercially, routine testing for DENV occurs only in public health laboratories, he said, although guidelines promoting DENV testing are under development by the CDC and the Association of Public Health Laboratories. Meanwhile, PAHO has established a network of dengue diagnostic laboratories, where the disease is endemic, throughout tropical and subtropical South and Central America and the Caribbean (PAHO, 2014a).
Because most of the United States is nonendemic for dengue, the majority of current cases involve returning travelers, Margolis noted. Dengue is the leading cause of acute febrile illness in travelers returning from the Americas, the Caribbean, and Asia (Freedman et al., 2006). Between 2000 and 2007, the number of such cases requiring hospitalization tripled in the United States (Streit et al., 2011). With thousands of travelers returning from dengue-endemic areas, coupled with the presence of Ae. aegypti in Florida, Texas, and Arizona, it is not surprising that a few episodes of limited local transmission have recently occurred in the United States, he remarked. The potential for more widespread local transmission, particularly involving Ae. albopictus, remains to be determined (Eisen and Moore, 2013).
Chikungunya
Like dengue fever, chikungunya is an acute febrile illness that can be reliably diagnosed only through molecular diagnostic testing, preferably by polymerase chain reaction (PCR),12 according to Margolis. Only supportive treatment (typically with nonsteroidal anti-inflammatory drugs) is available for chikungunya, and it is important to rule out dengue before proceeding, given the risk of hemorrhage, he pointed out.
The rapid geographic expansion and rise in chikungunya cases in the Americas, since its emergence in 2013, will continue, Margolis predicted, echoing Petersen’s earlier observations. Locally acquired cases have been confirmed in many countries throughout the Americas (CDC, 2014c), with the first such case in the United States reported in Florida in July 2014 (CDC, 2014b). This development was anticipated, and is expected to be repeated, given the large numbers of U.S. travelers returning from locations where major outbreaks have occurred (including Puerto Rico, as Margolis noted), coupled with the presence of both competent mosquito vector species—Ae. albopictus and Ae. aegypti—in this country (CDC, 2014a; Fischer et al., 2014; Khan et al., 2014).
At the time of the workshop, about 1,400 cases of chikungunya per week were being reported in Puerto Rico, mainly in the metropolitan San Juan area, Margolis stated.
The introduced virus has been traced to the Dominican Republic, he said, and many of these early cases have arisen in the city’s Dominican community. For such a “virgin soil” epidemic, it is difficult to predict how many epidemic cycles will occur before herd immunity is established. CHIKV’s rapid expansion clearly demonstrates that its mosquito vector, Ae. aegypti, is not controlled, he observed.
Dynamics of Leishmaniasis and Chagas Disease
Two vector-borne parasitic diseases, leishmaniasis and Chagas disease, are strongly associated with poor living conditions that expose people to the insect vectors that carry them. “These are diseases of poverty,” observed speaker James Maguire, of the Harvard Medical School. “In some senses the vector is poor. I think the poor vectors are picking on poor people.” This is also true of African sleeping sickness, which, like leishmaniasis and Chagas disease, is caused by a member of a group of flagellated protozoa known as kinetoplastids.13 Members of this group parasitize a broad range of animals and plants (Wiser, 2013).
___________________
12 PCR is a laboratory technique used to amplify DNA sequences. The method involves using short DNA sequences called primers to select the portion of the genome to be amplified. The temperature of the sample is repeatedly raised and lowered to help a DNA replication enzyme copy the target DNA sequence. The technique can produce a billion copies of the target sequence in just a few hours (http://www.genome.gov/Glossary/?id=159).
13 The major distinguishing feature of this group is a subcellular structure known as the kinetoplast, a distinct region of the mitochondria (Wiser, 2013).
Leishmaniasis
The more than 20 species of Leishmania capable of causing leishmaniasis can be vectored by nearly 100 species of sand flies, according to Maguire. Cases have been reported on every continent except Antarctica and Australia, and about 1.5 million new infections occur each year. All except two species of Leishmania that infect humans are zoonotic, and most species tend to cause subclinical disease. Clinical leishmaniasis presents in three main forms: cutaneous, mucosal, and visceral. All three are treatable to some degree, he said (Antinori et al., 2012).
While Leishmania have existed for at least 80 million years, they have only coexisted with humans for several millennia—an association that has produced tremendous diversity, he continued. “This is still a very dynamic set of organisms,” he observed. “They are emerging. They are reemerging. They are expanding their geographic range. This is a parasite that is definitely on the move.”
Maguire presented the following examples to illustrate the spectrum of drivers that influence leishmaniasis’ transmission patterns and geographic range:
- Cutaneous leishmaniasis, present in the Americas prior to human arrival, has recently exhibited increased incidence and broadening geographic range in response to human incursions into the forest (e.g., chicle harvesting) and the expansion of human settlement to formerly forested areas. Its complex transmission patterns involve multiple parasite and sand fly species, reservoir hosts, and varied ecology.
- Cutaneous leishmaniasis caused by a parasite that was once apparently sylvatic has become adapted exclusively to the domestic environment in a Brazilian community, with dogs and humans serving as its sole hosts.
- The range of locally transmitted cutaneous leishmaniasis in Oklahoma and Texas has moved northeast with increasing temperature, as predicted by models (Clarke et al., 2013). Similar predictions of expanded range with increasing temperature have also been borne out for visceral leishmaniasis in Europe.
- Most cases of cutaneous leishmaniasis in the United States have occurred among travelers, including significant numbers of military personnel. As a result, that sector has greatly advanced the prevention, diagnosis, and treatment of leishmaniasis, Maguire said.
- An apparently non-vector-borne outbreak of visceral leishmaniasis in a kennel of fox hounds in New York state spread to 18 other states as infected dogs traveled to participate in hunts. No human cases resulted, despite the presence in the area of a competent vector species.
- Visceral leishmaniasis in Brazil, long a rural disease, shifted around 1980 to a primarily urban or peri-urban disease, coincident with drought-induced mass migration from rural to urban areas. Epidemics occurred in several major cities, and incidence and geographic range increased, unabated by reactive spraying and campaigns to cull infected dogs (current
-
strategies under investigation include insecticide-impregnated collars for dogs, and a canine vaccine).
- At the same time, as HIV has moved into more rural regions of Brazil, co-infection with visceral leishmaniasis—which tends to produce severe disease—is on the increase. Moreover, co-infected individuals were found to be highly infectious to sand flies.
- In Europe, co-infection with HIV and leishmaniasis has occurred most frequently via shared needles, rather than insect bite.
- A recent large outbreak of leishmaniasis in Madrid was probably related to the development of a park within the city, which altered the ecology of hares, which served as a reservoir for the parasite (Carrillo et al., 2013).
- An explosive and lethal outbreak of visceral leishmaniasis occurred in East Africa during the early years of a civil war in southern Sudan, after refugees migrated through a region with high concentrations of sand flies.
- Eighty percent of visceral leishmaniasis cases occur in Bangladesh, India, Nepal, and South Asia, where humans are the sole hosts. Nearly extinguished by indoor insecticide spraying in the 1960s, leishmaniasis resurged in this region after malaria eradication efforts were abandoned.
Chagas Disease
Several species of blood-sucking triatomine bugs native to the American continents (but not the Caribbean) transmit Trypanosoma cruzi, which is known to infect over 100 species of mammals, Maguire stated. The first human hosts encountered and displaced the bug when clearing forests several thousand years ago. Some species adapted to the domestic environment and are today responsible for most human infections, he explained. About eight million people are permanently infected with this parasite that, decades after infection, can provoke life-threatening heart or gastrointestinal disease. Available treatments are “not satisfactory” and “toxic,” and it is uncertain whether they prevent the development of heart disease, he noted (Rassi et al., 2010).
Maguire described several settings in South and Central America where he and coworkers had investigated transmission of Chagas disease since the 1960s. Each illustrated one or more factors that supported disease transmission. In one community, only about half of the population was infected—those who could not afford a house with plaster walls and a tile roof to prevent colonization by bugs. Other outbreaks coincided with the introduction of a new vector species as roads were built, and as religious pilgrims visited the area. Researchers accidentally introduced a domesticated vector prevalent in one region into El Salvador. The introduced vector spread through Central America along the Pan American highway and beyond, becoming more important in this new territory than the native vector for Chagas disease. Maguire recalled that when poor migrants from rural
areas to cities supported themselves by selling their blood, a major outbreak of transfusion-associated cases of Chagas occurred.
In the early 1990s, the six southernmost countries in South America—which accounted for the majority of cases in the hemisphere—collaborated in an effort to knock out parasite transmission by the major vector in the Southern Cone region, Triatoma infestans, and to end blood-borne transmission of Chagas disease. “It cost $30 million to $50 million dollars a year versus billions of dollars of economic losses from the disease,” Maguire reported, and “it was incredibly successful.” As a result, Uruguay and Chile are now free of parasite transmission, Brazil is free of transmission by T. infestans, and the other countries have only low transmission rates. Additional initiatives in the Andean and Central American regions used similar tactics to target different vectors of Chagas disease. Together, these efforts have lowered the prevalence of infection from as high as 18 million in the 1980s to its current level of about 8 million.
A number of obstacles stand in the way of eliminating Chagas disease altogether in these regions, Maguire noted:
- the re-infestation of houses with sylvatic T. infestans or other sylvatic triatomine bugs that can adapt to domestic environments;
- passive transport of alternative vectors from other regions;
- the development of insecticide resistance by vector species; and
- ongoing migration from rural to urban areas.
In the Amazon region, which is particularly rich in reservoir and vector species, outbreaks of acute Chagas disease have been traced to the ingestion of juice contaminated with vector feces, he noted.
Maguire observed that in the United States, there are as many people infected with T. cruzi—about 300,000—as in 8 of the other 20 countries where Chagas disease is endemic (Montgomery et al., 2014). An estimated 30,000 cases of chronic Chagas-related heart disease and hundreds of cases of congenital disease remain undiagnosed, he added. Nearly half the states in the United States are inhabited by several species of the triatomine bug vector and mammals—including dogs—that are heavily infected with T. cruzi. Both acute and asymptomatic disease have been reported, mostly among immigrants from South and Central America, but there have also been 23 locally transmitted cases, 5 transfusion-associated cases, and a single case of congenital disease, he reported.
A similar situation exists in Europe and Japan, where such nonvectorial routes of transmission have outstripped vector-borne Chagas disease, Maguire observed. This is increasingly true in South and Central America as well. To control transmission through this diversity of routes will require infected people to be identified, served by the health care system, and treated with effective drugs, he concluded.
Changing Paradigms for Tick-Borne Diseases in the Americas
Ticks rank second only to mosquitoes as arthropod vectors of medical importance, according to speaker Christopher Paddock of the CDC. In the United States, tick species in four genera—Amblyomma, Ixodes, Dermacentor, and Rhipicephalus—transmit the majority of human pathogens. Each of these has three different feeding stages and wide host ranges, so their potential to transmit zoonotic disease is “really tremendous,” he said—and it has been increasingly realized in this country over the past century, during which nearly every recognized tick-borne disease emerged (see Paddock et al. in Appendix A8). “It has been a century of discovery and change,” he observed. “There have been lots of newly recognized diseases. There have also been dramatic shifts in the incidence and distribution of certain historically recognized disease like Rocky Mountain spotted fever . . . [as well as] paradigm changes in terms of vector and ecology associations with these pathogens.” In particular, he noted, human activity has profoundly affected tick-borne infections, which continue to shift in scope and magnitude.
As an example of this trend, Paddock described recent changes in the epidemiology of Rocky Mountain spotted fever (RMSF), recognized as the first tickborne disease of humans in this country more than 100 years ago. Until a decade ago, only two vectors were believed to transmit the bacterial pathogen, Rickettsia rickettsii, the Rocky Mountain wood tick in the East, and the American dog tick—Dermacentor variabilis—in the West. Neither species inhabits Arizona, yet in 2003 a boy there died of RMSF, and soon afterward, 14 members of his small mountain community were found to be infected. Upon investigation, the brown dog tick—which typically does not bite humans but is present throughout the Americas and on other continents as well—was identified as the vector in these cases. The local population of those ticks had exploded, driving them to expand their host range.
This situation has been duplicated in other Arizona communities such that Paddock described it at this meeting as the “new normal” of RMSF epidemiology in the western United States. In 2011, for example, 77 cases of RMSF were reported in Arizona, including 6 deaths. Tragically, he added, children less than 10 years of age comprise the majority of RMSF cases vectored by the brown dog tick, because young children typically share dogs’ habitats more than do other age groups. More severe outbreaks involving this vector have occurred in Northern Mexico, he added, resulting in significant mortality. “This may become a border health issue,” he warned. Increasing numbers of cases are being reported from cities like Calexico, California, and Nogales, Arizona, involving people who traveled to endemic regions within Mexico and then returned to the United States, where their symptoms caused them to be hospitalized.
Over the last decade, additional rickettsial diseases have been identified as distinct from RMSF, Paddock added. Cases of milder R. parkeri rickettsiosis
(inferred to exist in 1948 by the eponymous R. R. Parker) have been confirmed in several states since 2004 (Cragun et al., 2010; Paddock et al., 2008). In California, discovery of the first cases of 364D rickettsiosis has led to the suggestion that this pathogen is behind most cases of “RMSF” in that state (Johnston et al., 2013). Another relatively mild—and distinct—rickettsiosis was recently identified in Brazil where RMSF is also present (Angerami et al., 2009). All of these rickettsioses respond to treatment with doxycycline, Paddock noted, but it is nevertheless important to distinguish one from another. “If you are going to accurately describe the clinical features and the epidemiology of these diseases you really have to know . . . what is causing them,” he insisted.
An analogous situation exists with Borrelia miyamotoi, a tick-borne pathogen closely related to the Lyme disease bacterium, Borrelia burgdorferi, Paddock continued. Patients infected with B. miyamotoi can have antibodies that cross-react with B. burgdorferi antigens. B. miyamotoi was first associated with human disease in 2011, and has been detected in ticks in areas where Lyme disease is endemic in both the northeastern and western United States. The seroprevalence of B. miyamotoi infection among residents of New England, for example, has been determined to be as high as 4 percent. Its clinical spectrum remains to be determined, but the few cases that have been evaluated range from fever to very severe meningoencephalitis, he said. Meanwhile, novel clinical manifestations of Lyme disease have been identified, including sudden cardiac death among patients in their 20s and 30s (CDC, 2013).
The Heartland virus, previously described by Petersen as an emerging tick-borne disease of newly recognized importance, was first isolated in 2009 from two patients in Missouri who were initially suspected to have ehrlichiosis (McMullan et al., 2012). All confirmed infections with this virus that have occurred were in men older than 50 years of age, in whom it is a life-threatening infection, Paddock stated. Cases have been sporadically identified throughout the range of its vector, the Lone Star tick, he noted, so there are probably many more unrecognized infections. “It is going to be wherever this tick exists in the United States, which is also expanding in its range,” he observed.
Explosive population growth among white-tailed deer has surely driven the expansion and emergence of tick-borne disease in the United States, Paddock remarked (Paddock and Yabsley, 2007). As many as 30 million deer now inhabit this country, as compared with an estimated 300,000 animals at the beginning of the 20th century. Deer are keystone hosts for the two most important vector species of tick-borne pathogens in the United States, namely black-legged ticks (Ixodes scapularis), known to transmit at least seven different human diseases, including Lyme, and Lone Star ticks (Amblyomma americanum), which carry Heartland virus and two species of Ehrlichia known to cause human disease.
THE PUBLIC HEALTH RESPONSE
Four speakers described the considerable challenges encountered by public health organizations in the Americas and Europe as they attempt to identify and adapt to changing patterns of vector-borne diseases.
Vector-Borne Disease Surveillance and Response in Latin America and the Caribbean
Many countries in the Americas are endemic to both vector-borne and neglected tropical diseases, which present similar public health challenges, according to speaker Luis Gerardo Castellanos of PAHO. PAHO, founded with the First General International Sanitary Convention of the American Republics in 1902, is the world’s oldest continually functioning international public health agency. Today PAHO represents 35 countries that make up nearly one-third of the world’s land mass, and 14 percent of its population. The organization also includes associate members, observer states, and participating states representing territories in the region. PAHO’s initial mission of controlling epidemic diseases has broadened to include noncommunicable disease control, health education, and environmental improvements designed to help all people, especially those in need, he explained.
Yellow fever was the first vector-borne disease to be battled in the Americas, Castellanos noted. PAHO coordinated 11 countries in an attempt to eliminate yellow fever and malaria from the Panama Canal Zone, following identification of their common vector, Ae. aegypti (see section on “History and Current Challenges of Dengue Vector Control” for a detailed account of this effort and its aftermath). Currently malaria is endemic in 21 countries represented by PAHO, and more than 430,000 cases and 82 deaths were reported in 2013—a 64 percent reduction in cases since 2000, he reported. Fourteen member states are free of local malaria transmission today, he added, and according the WHO, seven additional countries may soon qualify as malaria free. PAHO has supported this progress by preparing strategic plans of action that, once approved by a country’s minister of health, become binding and are documented by annual progress reports, he explained.
All four serotypes of DENV, also spread by Ae. aegypti, are present and causing disease throughout Latin America and the Caribbean, according to Castellanos. In 2003, PAHO undertook an integrated management strategy14 to control the spread of dengue which, in turn, informed the WHO’s Global Strategy for Dengue Prevention and Control, launched in 2012. That the Americas currently report more dengue cases, but lower case fatality, than any other global region he
___________________
14 See http://www.paho.org/hq/index.php?option=com_content&view=article&id=4501&Itemid=41038&lang=en (accessed March 25, 2016).
attributed to “a very robust surveillance system across all countries” (see Castellanos in Appendix A4).
The emergence of chikungunya in the Americas in late 2013 was anticipated by PAHO which, in 2010, began developing preparedness plans for the Caribbean region in collaboration with partners including the CDC and the Institut Pasteur, Castellanos said. Although these plans were in place by 2012, as of November 2014, more than 900,000 suspected and nearly 16,000 confirmed locally transmitted cases of chikungunya have been reported in the Americas (PAHO, 2014b). PAHO and its partners have established a network of referral laboratories located in Argentina, Brazil, Cuba, French Guyana, and the United States to support the entire region in responding to this challenge.
In the meantime, PAHO continues to pursue elimination of several infectious diseases throughout the Americas, including onchocerciasis (river blindness) in Ecuador, which is soon to be declared free of the disease by WHO, according to Castellanos; Mexico and Guatemala are expected to gain that designation by 2016. He also noted the following vector-borne diseases expected to achieve elimination: trachoma in Mexico; lymphatic filariasis in Brazil; schistosomiasis in Suriname, the Dominican Republic, and St. Lucia; malaria in Argentina and Paraguay; and Chagas disease within several cities, provinces, or departments of Argentina, Columbia, Mexico, and Peru. Despite these gains, he observed, “Vector-borne diseases will continue to be a dynamic public health threat to countries in the Americas.” Governments and international stakeholders must therefore commit themselves to preventing the further spread of these diseases, he concluded.
Vector-Borne Disease Emergence and Spread in Europe
The European Union is a hot spot for infectious disease emergence, and it is highly interconnected with other hot spots internationally, according to speaker Jan Semenza of the European Centre for Disease Prevention and Control (ECDC). Based on a foresight study conducted by ECDC in 2008, Europe is at risk of vector-borne disease threats owing to anticipated changes in drivers of infectious diseases by 2020. These predictions were validated in 2013 with an analysis of the infectious disease threats (and their contributing drivers) that occurred over the preceding 5 years that were identified by epidemic intelligence at ECDC. Semenza described these studies and their results in more detail (see Appendix A11) and elaborated on the ECDC’s strategy to tackle these threats from vector-borne disease emergence.
For their prospective study of infectious disease threats to Europe through 2020, the ECDC first assembled expert panels to identify major drivers of infectious disease in the region, Semenza reported. These were determined to fall into one of three broad categories: globalization and environmental change; social and demographic change; and public health systems (Suk and Semenza, 2011). Based
on the results of an extensive literature review structured around these disease drivers, the researchers created scenarios to anticipate their influence on eight likely, significant near-term infectious disease threats to Europe as illustrated in Figure WO-10. Among them, introductions of vector-borne diseases or shifts in their transmission were predicted to be driven primarily by trends subsumed under the category of globalization, migration, and environmental change, as well as by social inequality (Suk and Semenza, 2011).
To validate the conclusions of this “thought experiment,” as Semenza described it, the investigators conducted a retrospective analysis of ECDC epidemic intelligence data on infectious disease threats to European member states of the ECDC between July 2008 and December 2013. Among the 116 qualifying public health events they analyzed, nearly one-quarter involved vector-borne diseases, Semenza reported. Sixty-one percent of these vector-borne disease threats were attributed to global and environmental change, he added.
Given the disproportionate contribution of global and environmental change drivers to vector-borne disease threats in Europe, the ECDC turned to infectious disease experts for advice in crafting strategies to meet these challenges. Many of these experts considered climate change to be of particular concern as a driver of selected vector-borne disease such as Lyme disease, West Nile viral fever, tickborne encephalitis, and leishmaniasis, Semenza stated (Semenza et al., 2012). The arrival and dispersal of tropical pathogens commonly associated with warmer temperatures is a potential threat to the safety of the blood supply. A ranking of emerging infectious diseases that can be a threat to substances of human origin (such as blood cells, tissues, or organs) in the European Union was compiled, based on an assessment of experts in the field (Semenza and Domanovic, 2013).
The majority of these infectious disease experts also expressed concern that their national disease surveillance and health systems were unprepared to deal with the effects of climate change on infectious disease dynamics, Semenza noted (Semenza et al., 2012). To prioritize surveillance improvements in light of these public health challenges, ECDC evaluated both notifiable and non-notifiable infectious diseases in terms of the strength of their link with climate change and the potential severity of their consequences to society, he said (Lindgren et al., 2012). Top-ranked diseases, in need of more surveillance activities, included Lyme disease, dengue fever, tick-borne encephalitis (TBE), Rift Valley fever, chikungunya, and leishmaniasis, he reported.
To address gaps in surveillance and preparedness, ECDC has built the European Environment Epidemiology (E3) Network. The E3 Network is built to monitor environmental precursors of epidemic events, including vector-borne disease outbreaks, to facilitate a more effective public health response, Semenza said. “We have compiled and processed a large number of environmental data that are now available for epidemiologic analysis such as prediction modeling,” he explained. These data and prediction models are hosted at the E3 Geoportal (Semenza et al., 2013). The following examples illustrate how data from the
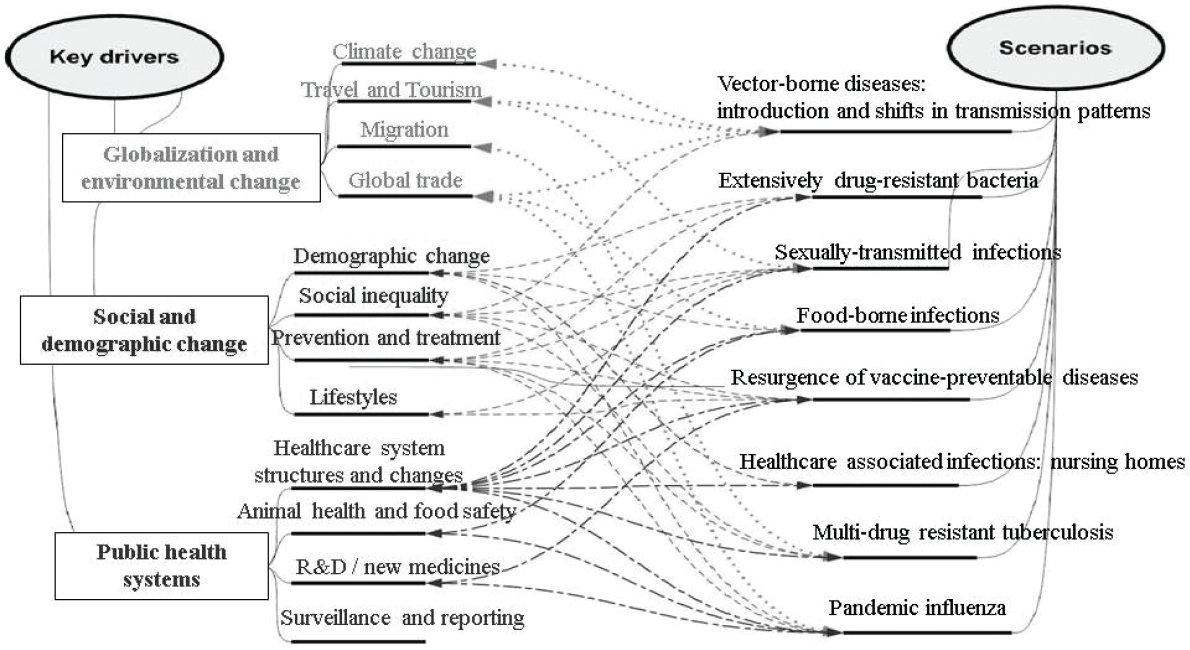
NOTE: R&D = research and development.
SOURCE: Suk and Semenza, 2011. Reproduced with permission from the Sheridan Press, on behalf of the American Public Health Association.
E3 Geoportal have been used to inform effective public health action to address vector-borne diseases:
- Following the reintroduction and autochthonous transmission of malaria to Greece in 2009–2012, environmental data from the E3 Geoportal were used to develop a multivariate model to identify areas at risk for transmission based on environmental and climatic conditions. These insights were used to enable targeted pesticide spraying, dedicated surveillance, and public outreach. Disease transmission was subsequently interrupted, ending the outbreak in 2013 (Sudre et al., 2013).
- The risk for transmission of TBE in certain areas of southern Sweden was characterized with environmental data from the E3 Geoportal. By delineating these areas environmentally suitable for transmission, residents and tourists can be alerted to the risks and targeted with vaccination campaigns; the same approach could potentially be expanded to other areas at risk for TBE in northern Europe.
- The relationship between temperature deviations from the mean and WNV infections were assessed during the WNV outbreaks in Europe in 2010. Environmental and meteorological data (July temperature) from the E3 Geoportal were used to develop a predictive model of WNV that can now be used to predict outbreak areas in the future (Paz et al., 2013; Tran et al., 2014).
- The risk for dengue importation into Europe was modeled for 2010 with air passenger volume from dengue active areas internationally. These analyses can be used in the future to predict the airports most at risk and the timing of potential onward transmission in the destination country (Semenza et al., 2014).
- Similarly, these data were also used to identify spatiotemporal risk parameters for chikungunya importation into Europe from the Americas.
In addition to these applications of the E3 Network, the ECDC has compiled a handbook with practical climate change adaptation measures for infectious diseases: Climate Change and Communicable Diseases in the EU Member States15 (Ebi et al., 2013). “By monitoring the environmental precursors of disease we hope to be able to help forecasting and predicting these patterns of disease emergence in order to enhance preparedness and reduce human and economic costs, particularly in resource strapped regions in Europe,” Semenza concluded.
___________________
15 See http://www.ecdc.europa.eu/en/publications/Publications/1003_TED_handbook_climatechange.pdf (accessed March 25, 2016).
Loss of Arbovirus Disease Surveillance Capacity in the United States
Before discussing the findings of a recent assessment of national capacity for arbovirus surveillance, speaker James Hadler of Yale University described the structure of public health surveillance in the United States. His review emphasized that surveillance for diseases of public health importance is a state function. Even in the case of nationally notifiable conditions, states collect and transmit information to the CDC as they see fit. He also noted that 80 percent of all surveillance by state health departments is federally funded (mainly through the CDC) and that, as a condition of funding, the CDC can require standardized surveillance methods and reporting.
Hadler noted that arboviruses such as WNV are of particular interest to public health—and therefore candidates for surveillance—for several reasons. Some cause severe morbidity and death; they are associated with large, rapidly developing outbreaks with the potential to overwhelm the health care system; they can be transmitted through the blood supply (or via organ transplant) as well as by insects; and both the infections and outbreaks they cause are potentially preventable if we know which arboviruses are present and the level of threat associated with them.
Before the emergence of WNV in the United States in 1999, no federal funding supported state or local surveillance for arboviral infections, which was limited largely to the voluntary reporting of human and animal cases of several types of encephalitis—and in several states was nonexistent, Hadler stated. That year federal funding for WNV surveillance was distributed from the CDC to the affected states through a cooperative agreement program known as Epidemiology and Laboratory Capacity (ELC). By 2004, WNV had reached every state except Washington, Alaska, and Hawaii, and ELC funding and guidance were extended to all 50 states and six major cities or counties. Also, by 2004, the ArboNET electronic national reporting system (previously discussed by Petersen) was collecting information from every state on avian mortality and surveillance of sentinel birds, horses, mosquitoes, and human infections. A 2005 Council of State and Territorial Epidemiologists (CSTE) survey of state and selected local health departments found that federal funding had enabled the development of broad-based, multisectorial WNV surveillance capacity in all states and recommended that states be permitted to expand the use of ELC funding to more broadly address vector-borne disease surveillance (CDC, 2006).
This recommendation was implemented, Hadler said, but as neuroinvasive cases declined rapidly after peaking in 2003, annual ELC funding for WNV surveillance gradually shrunk from $24 million in 2004 to $9 million in 2012, as shown in Figure WO-11. In 2012, however, neuroinvasive West Nile viral disease cases spiked to levels not seen in nearly a decade. “It was clear that WNV still had the potential to cause large-scale outbreaks, measurable not just locally, but nationwide,” he observed. In response to this development, and also to the threat of other emerging arboviruses such as DENV and CHIKV, CSTE—in partnership
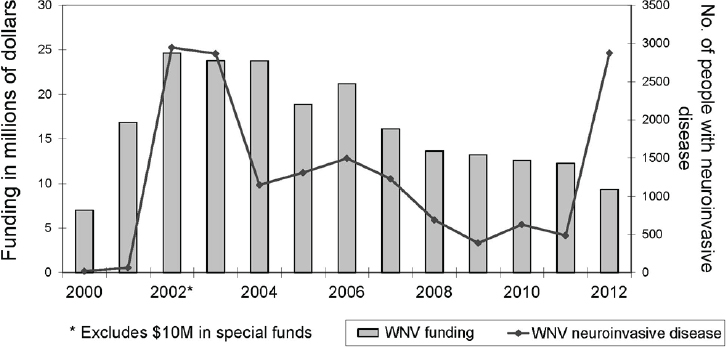
NOTE: WNV = West Nile virus.
SOURCE: Hadler et al., 2014 (CDC).
with state health departments and additional public health organizations—set out to examine how arbovirus surveillance capacity had changed through years of lean ELC funding. Their follow-up assessment found that since 2004, while nearly all states continued to conduct at least passive human disease surveillance, 22 percent of jurisdictions had stopped conducting active human surveillance, 13 percent had stopped mosquito surveillance, 70 percent had reduced mosquito trapping and testing, and 64 percent had eliminated avian mortality surveillance (Hadler et al., 2014).
Hadler reported that state surveillance capacity has decreased substantially since 2004, particularly with respect to conducting active human surveillance (including offering testing), mosquito surveillance, and laboratory resources. “From our perspective this has really reached a tipping point in some states,” he said. Due in part to decreases in ELC funding, he observed, “The ability to rapidly detect emerging and outbreak level threats and thus initiate prevention activities has clearly been compromised.” The CSTE report recommends that the CDC examine national arboviral surveillance capacity and secure additional support as needed to ensure mosquito surveillance occurs in all metropolitan areas with historically high West Nile viral disease burden (Hadler et al., 2014).
Hadler characterized current national capacity to conduct surveillance for vector-borne pathogens other than WNV as “very patchy,” partly because of limited laboratory capacity at the state and local levels. CSTE estimates that a 50 to 60 percent increase in full-time employees will be required to enable state
and local health departments to meet their criteria for “full capacity” arbovirus surveillance (Hadler et al., 2014). According to Hadler, public health jurisdictions at “full capacity” for arbovirus surveillance meet the following three criteria:
- They have the ability to complete a standard case report form on every suspected/confirmed case and report it to ArboNET.
- They have the ability to test by IgM for all relevant arboviruses on any cerebrospinal fluid (CSF)/serum specimen submitted on a suspected case of arboviral disease.
- They have a surveillance system that includes mosquito surveillance to routinely monitor both larval and adult arboviral activity in all parts of the jurisdiction in which there is the potential for human outbreaks of arboviral disease based on past experience.
In subsequent discussions, workshop participants considered how to address the issue of inadequate surveillance for mosquito-borne diseases in particular. Both Petersen and Margolis noted that while mosquito surveillance proved a good early indicator of outbreaks of West Nile fever, that is not the case for dengue, nor might it be for chikungunya. Petersen noted that surveillance for mosquito-borne diseases in the continental United States is limited largely to Culex mosquitoes, which are very different from the Aedes species.
Moreover, Petersen said, mosquito-based surveillance is useful for preventing or controlling outbreaks only if it generates a speedy response. “Really, we want to aim to build local capacity for mosquito-based surveillance. They are the ones making the decisions. They need to make them quickly.” To this end, forum member Roger Breeze of Lawrence Livermore National Laboratory recommended that health departments take advantage of technological advances, such as multiplex PCR, to expedite pathogen analysis. “We are still stuck in a very 1990s paradigm, and bemoaning the fact that we don’t have lots of people doing 1990s technology that you can do with a machine,” he observed. He also noted that the Department of Defense is attempting to develop “a completely autonomous system to analyze what [pathogens are] . . . flying around in the mosquito and report to you.”
Blood Donation Screening for Vector-Borne Diseases
Beginning with syphilis testing in 1938, and increasingly since the emergence of HIV in the early 1980s, blood donations in the United States have been screened for a growing number of pathogens, according to speaker Susan Stramer, of the American Red Cross. Many vector-borne disease agents have been shown to be, or suspected to be capable of, transfusion transmission, which is important because of the large, explosive nature of outbreaks caused by these agents. Few interventions for such agents are available, and treatments can be
costly with the development of new therapeutics likely to be slow, she observed. Thus, vector-borne diseases figure prominently among those infectious diseases deemed threats to the U.S. blood supply,16 as determined by the AABB (formerly the American Association of Blood Banks). To estimate the magnitude of such a threat, she explained, researchers attempt to answer a series of key questions about the pathogen and the disease it causes, including
- Does the disease have an asymptomatic blood-borne phase, enabling donors who feel and appear well to transmit it?
- Does the pathogen survive through blood component preparation, distribution, and storage?
- How severe is the disease? What is the outcome in those who are immunosuppressed?
- Is the disease treatable?
- Is the pathogen present in the donor population? If so, is it increasing or decreasing?
- Does the public fear this pathogen (whether or not that fear is justified)?
- What intervention(s) would effectively protect the blood supply from this pathogen? (Dodd, 2012)
Such questions have been incorporated into models such as one Stramer described called the European Upfront Risk Analysis Tool,17 which estimates transmission risk in blood. In her presentation to the workshop, she described recent and current attempts to assess and address the transfusion-associated transmission risk posed by several emerging vector-borne pathogens. Each of these pathogens, she noted, has been addressed on a case-by-case basis. Collectively, they illustrate the need to create decision-making processes for protecting the blood supply from the wide range of vector-borne pathogens.
Lessons from WNV
The response to WNV serves as a model of success in recognizing and preventing transfusion-associated transmission of a vector-borne disease, Stramer said. Although most WNV infections are asymptomatic, interventions introduced less than 1 year after the first transfusion-associated cases were identified greatly
___________________
16 The August 2009 issue of Transfusion included a supplement on emerging infectious disease (Stramer et al.) agents and their potential threat to transfusion safety. Members of AABB’s Transfusion Transmitted Diseases Committee identified 68 infectious agents and described them in detail, providing background information about each agent, along with a variety of assessments such as the clinical features of the agent and those characteristics specifically related to transfusion transmission. New fact sheets on emerging threats and updates to previously published fact sheets are also available. See http://www.aabb.org/tm/eid/Pages/default.aspx for details (accessed March 25, 2016).
reduced exposure to the virus, she stated. Continued refinement of testing procedures has further decreased “breakthrough” transmission. To date, more than 3,700 WNV-positive donations have been removed from the blood supply, she reported.
WNV taught stewards of the U.S. blood supply several important lessons, Stramer noted. It was their first experience in dealing with a transfusion-transmissible infection that was an acute, rather than chronic infection, like HIV, or hepatitis B or C viruses. While recognizing the potential of nucleic acid testing to provide rapid results, they also discovered that testing pooled blood samples—which can save both time and cost—may be insufficiently sensitive to low levels of virus, she said.
The DENV Conundrum
Like WNV, DENV frequently produces asymptomatic infections, Stramer observed. There is as yet no Food and Drug Administration (FDA)-licensed screening test for DENV. Investigational testing under way in Puerto Rico since 2010, however, has produced comparable results to established WNV protocols, she reported. In retrospective tests of more than 15,000 blood donations acquired at the peak of the Puerto Rican dengue epidemic, about 1 in 500 samples tested positive for DENV (Stramer et al., 2012).
On the other hand, only three clusters of transfusion-associated DENV transmission have been reported, in Hong Kong, Puerto Rico, and Singapore (updated data, however, indicate seven clusters). This does not appear to be an artifact of inadequate surveillance, Stramer said. She noted, however, that it is often difficult to distinguish mosquito-borne from blood-borne cases of dengue in developing countries, where even hospitalized patients may be significantly exposed to mosquito bites. It is also possible that either immunosuppression or the simultaneous receipt of antibodies to DENV in transfused blood may reduce apparent transfusion-associated infections, she added. Lastly, recognition of dengue symptoms in severely ill patients may be difficult against the background of underlying disease in the recipient. Alternatively, as Rico-Hesse suggested, amplifying factors in mosquito saliva may significantly increase the effectiveness of vector-borne transmission.
Responses to CHIKV
Like DENV, blood donated within the United States is not presently being routinely screened for CHIKV RNA, Stramer reported. However, unlike DENV infection, which CHIKV resembles in terms of the progress of viremia and antibody development, approximately three in four cases of chikungunya infection are symptomatic, she noted. This would tend to reduce the number of infected donors, and also theoretically make it possible to intercept donations from people
who report postdonation symptoms within a few days. To date, there have been no documented CHIKV transmissions associated with blood transfusions, but this may also be caused by the limitations noted above for DENV, she reported.
Blood screening and other preventive measures have been taken in response to epidemic chikungunya elsewhere in the world, most notably after an explosive outbreak on the Indian Ocean island of Réunion between 2005 and 2007, in which more than 40 percent of the island’s inhabitants became infected with CHIKV. This viral strain—which acquired a mutation that increased its ability to replicate in Ae. Albopictus—was eventually introduced to northern Italy, and from there spread through Europe. Upon recognition of this introduction, blood collection was halted in at-risk areas of northern Italy, and in France, donors who had recently traveled to Réunion were deferred and nucleic acid testing for the virus was instituted, she said. The French also used this crisis to test a process known as platelet pathogen inactivation, which employs a broad-spectrum agent to prevent blood-borne transmission, which they found to be both safe and effective (Rasongles et al., 2009). The local collection of platelets, with an intervention, was required because of the short shelf life of platelets. Similar processes have since been shown to inactivate other pathogens, including DENV, in plasma and other blood products (Musso et al., 2014). Subsequent to this report (December 2014), the FDA cleared the process for use in the United States.
CHIKV was detected in blood samples from Caribbean donors within a few months of its emergence there (Gallian et al., 2014). Concerns about the blood supply in Puerto Rico, which has become endemic for CHIKV, and in the United States at large, have been discussed and are summarized below (Katz, 2014):
- Do nothing and watch, as we did before the emergence of WNV in summer 2002, responding if and when transfusion-transmission risk is demonstrated.
- Enhance our ability to identify the approximately 80 percent of donors who would be expected to have symptoms, by effectively eliciting callbacks by donors who get sick after a donation, so that we can recall their products.
- Understand donor travel and temporal donation patterns following travel, allowing us to model the effects of a short-term deferral for travel to affected areas. While operationally challenging, this may mitigate many acute tropical virus “sins.” (Stramer added, “We can temporarily stop collections in areas where we see focal outbreaks”; however, this is not sustainable and is costly.)
- Engage our test builders to have “on-the-shelf” nucleic acid assays to detect CHIKV using available test platforms. (Stramer characterized this option as cost prohibitive.)
An option not listed above could use pathogen inactivation systems as used by the French in Réunion to inactivate CHIKV as well as other emerging arboviruses.
In August 2014, under administrative order by the Puerto Rican Ministry of Health, blood donation centers began asking potential donors whether they or anyone in their neighborhood had experienced either symptoms or a diagnosis of CHIKV and/or DENV, and they told donors repeatedly to report postdonation symptoms within 3 days (and in the case of platelet or plasma donors, to confirm their symptom-free status if contacted, or their donation would be discarded). This order was subsequently modified to allow the use of pathogen-reduction systems available as licensed or through treatment-use studies.
Like CHIKV, no transmission of the recently emerged Zika virus has been reported to date, even though it is very closely related to DENV, according to Stramer; however, similar interventions have been taken to prevent its transfusion-associated transmission. In Oceana, she observed, “They have multiple outbreaks ongoing simultaneously: there is Zika, dengue viruses -1, -2, and -3, and chikungunya [virus], so the three can occur quite successfully together.” Because of these risks, several research blood donation screening interventions and pathogen reduction have been introduced in remote settings where importing blood components is not feasible, she explained.
Low Threat for Chagas
Few cases of blood-borne Chagas disease have occurred in the United States, Stramer reported, and blood-borne transmissions have only been documented by platelets, due to the fragility of the parasite (platelets are stored at room temperature and agitated to promote oxygen availability, likely enhancing survival of the parasite over their 5-day shelf life). Those transmission-associated cases that have occurred involved long-infected donors who came from endemic areas, she said. An extensive incidence study that followed over 4 million donors and greater than 6 million person-years of observation over the course of 4 years did not find any cases of incident infection, thus supporting a policy of selective testing involving testing each donor only once. The risk of missing a new case of infection was estimated 0.61 per million. Meanwhile, among more than 24 million donations screened between 2007 and 2014, the American Red Cross found about 1 in 36,000 positive donors, she stated.
Documenting and Preventing Transfusion-Associated Babesiosis
Of several tick-borne pathogens of concern to the blood supply, parasites of the genus Babesia—which infect red blood cells—are the most important, according to Stramer. General mortality for babesiosis, a malaria-like illness, ranges from 6 to 9 percent, but it is much higher for transfused recipients with underlying comorbidities and for other typically vulnerable patients, she observed.
There are hundreds of apparent transfusion-associated cases of babesiosis primarily confined to the northeastern United States and upper Midwest, she said, but fewer than 170 cases have been well documented. There is no FDA-licensed screening test for Babesia microti, the agent responsible for nearly all transfusion transmissions. Current interventions are limited to questions asked of donors, such as “Have you had babesiosis?” Potential improvements should not include questioning patients about their history of tick bites, since most donors do not know if they have been bitten, and if they have, which includes up to 9 percent of donors in endemic areas, such donors likely removed attached ticks during the grace period prior to B. microti infection. The only currently realistic intervention is to test all donations in endemic U.S. states for B. microti using both antibody and DNA tests. Investigational testing, including a retrospective study of donated blood, supports the use of these tests as an intervention against further blood-borne transmission of B. microti (Moritz et al., 2014).
ASSESSING AND ADDRESSING DRIVERS OF VECTOR-BORNE DISEASES
Three workshop speakers described diverse research efforts to investigate a spectrum of factors that potentially influence the transmission dynamics of vector-borne diseases, and to elucidate their mechanisms of action.
Weather, Agriculture, Climate, and Outbreak Patterns
Recent weather extremes have influenced agricultural production and created conditions conducive to outbreaks of certain vector-borne diseases, according to speaker Ken Linthicum of the U.S. Department of Agriculture (Anyamba et al., 2014; see adaptation, Linthicum et al., in Appendix A7). He described how he and coworkers investigated a series of extreme weather events between 2010 and 2012 that strongly affected agricultural production in major growing regions of Australia, East Africa, Russia, Southern Africa, and the continental United States—and where, sometimes simultaneously, outbreaks of vector-borne diseases (including dengue, Rift Valley fever, and West Nile virus disease) occurred. Using satellite data that track both vegetation density and land surface temperature, along with data on rainfall during the growing season, the researchers mapped anomalous conditions in these areas in detail, and compared these locations with places where major vector-borne disease outbreaks occurred during this period.
Linthicum and coworkers observed, for example, that when Texas experienced a 100-year drought in 2012, the overall vegetation index declined by 66 percent, production of cotton—a major crop—was cut in half, and a record-setting outbreak of West Nile virus disease erupted, he said. Drought in East Africa resulted in a loss of sorghum production at the same time as a large dengue
outbreak. Meanwhile, in areas where there was increased rainfall during this period, as occurred in southern Africa and southeast Australia, corn and cotton production increased coincident with outbreaks of Rift Valley fever in southern Africa, and Murray Valley encephalitis18 outbreaks took place in Australia (Anyamba et al., 2014).
In addition to these acute, short-term impacts of weather anomalies, shifts in climate affect vector-borne disease patterns over the long term, Linthicum stated. There is a close link between the climate fluctuation phenomenon known as the El Niño/Southern Oscillation (ENSO)—as illustrated in Figure WO-12—and global rainfall anomalies.
Global patterns of floods and droughts influence the emergence, propagation, and survival of mosquito vectors and ultimately the transmission of mosquito-borne pathogens associated with diseases that include Rift Valley fever, dengue and dengue hemorrhagic fever (DHF), and chikungunya, he explained (Anyamba et al., 2012; see adaptation, Linthicum et al., in Appendix A7). The result, he observed, is “episodic patterns of disease outbreaks that are in tune with climate variability.” For example, he noted:
- Hot and dry periods that occur during El Niño events in Southeast Asia have preceded significant peaks in DHF cases.
- Chikungunya outbreaks occurring between 2004 and 2010 were in some locations associated with extremely hot temperatures and/or drought, but in others with extremely wet conditions (Anyamba et al., 2012).
- In the Horn of Africa, recent outbreak clusters of chikungunya (2004−2006) were associated with severe drought, and Rift Valley fever (2006−2009) with heavy rainfall (Anyamba et al., 2012).
- Plasmodium vivax malaria reemerged in the Republic of Korea (post-eradication in the late 1970s) in 1993 during an extremely hot and dry period, and gained in incidence during subsequent periods of similar conditions (Linthicum et al., 2014).
- Global sea surface temperatures and rainfall patterns during the spring, summer, and fall 2014, and winter-spring of 2015, suggested that an El Niño event was imminent (see Figure WO-13). Figure WO-14 illustrates predicted regions of elevated risk for outbreaks of several vector-borne diseases if such an event occurred in 2014–2015.
___________________
18 Murray Valley encephalitis (MVEV) is caused by a mosquito-borne virus that is found across Australia, Papua New Guinea, and Irian Jaya. MVEV is endemic to northern Australia and causes occasional outbreaks across southeastern Australia. 2011 saw a dramatic increase in MVEV activity in endemic regions and the reemergence of MVEV in southeastern Australia. This followed significant regional flooding and increased numbers of the main mosquito vector, Culex annulirostris, and was evident from the widespread seroconversion of sentinel chickens, fatalities among horses, and several cases in humans, resulting in at least three deaths. The last major outbreak in Australia was in 1974, during which 58 cases were identified and the mortality rate was about 20 percent (Knox et al., 2012).
SOURCE: Anyamba et al., 2012. Available from PLoS Neglected Tropical Diseases under Creative Commons license.
While extremes of temperature and precipitation have significant implications for the emergence and spread of vector-borne diseases, the magnitude of ENSO influence on some of these extremes cannot currently be predicted, Linthicum cautioned. Disease transmission dynamics in different environments and populations may vary widely, he observed, reflecting a broad spectrum of influences on vector species, vector population sizes, and vectorial capacity. Nevertheless, he continued, “There’s obviously a need to invest in early ground surveillance during periods of unusual weather conditions”—including rapid field diagnostics for vector identification and virus isolation.
SOURCES: NOAA, 2015; Reynolds et al., 2002.
Globalization, Land Use, Global Warming, and the Invasion of West Nile Virus
Continuing the discussion on WNV, A. Marm Kilpatrick of the University of California, Santa Cruz, described its emergence in the United States as a case study in the intersection of multiple disease drivers, including novel pathogen introductions, land use, climate change, and the evolution of pathogen, host, and vector, as illustrated in Figure WO-15 (Kilpatrick, 2011) (see also Appendix A6). While West Nile viral disease patterns are often characterized as complex, he noted, “I think it’s actually our job as scientists to take that complexity and distill it down to those factors that matter the most.” Hence in his presentation, he posed—and to a large extent, answered—a series of questions intended to accomplish this goal.
How might WNV have arrived in North America? There are five main vehicles for such zoonotic vector-borne pathogens, Kilpatrick stated: infected humans, wind-transported mosquitoes, human-transported mosquitoes (e.g., on planes or boats), human-transported nonhuman hosts (e.g., poultry), and migratory hosts (e.g., birds). Using mathematical models of these scenarios, Kilpatrick and coworkers projected that WNV was most likely to be introduced to Hawaii (Kilpatrick et al., 2004) and the Galapagos (Kilpatrick et al., 2006) by mosquitoes
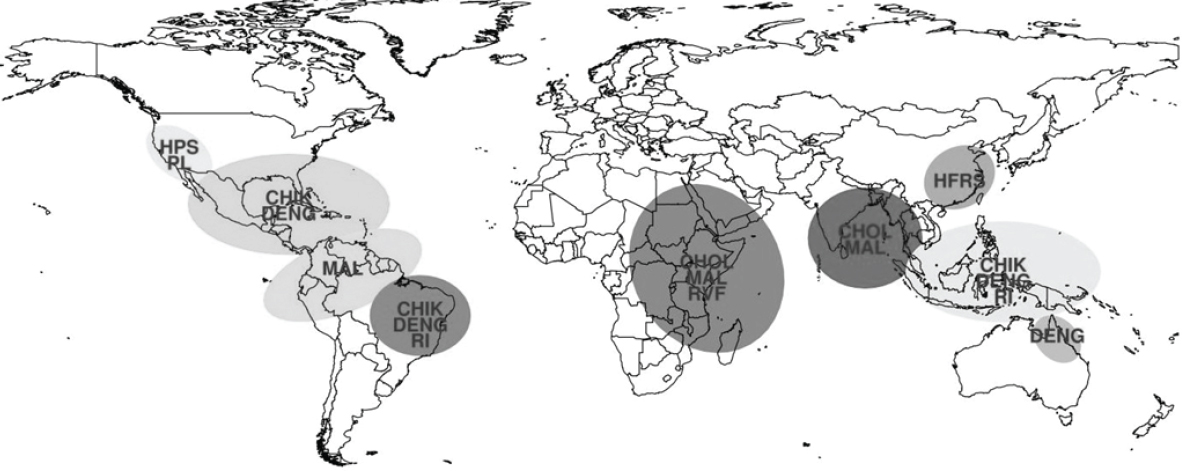
NOTE: CHIK = chikungunya; CHOL = cholera; DENG = dengue fever; HFRS = hemorrhagic fever with renal syndrome; HPS = hantavirus pulmonary syndrome; MAL = malaria; PL = plague; RI = respiratory illness; RVF = Rift Valley fever.
SOURCE: Chretien et al., 2015. Available from PLoS Current Outbreaks under Creative Commons license.

SOURCE: Kilpatrick, 2011, © AAAS.
that arrived on airplanes, but that migratory birds would provide an easier entry for WNV into the Caribbean (Douglas et al., 2007), he explained.
Key WNV Vectors and Hosts
What are the important vectors and hosts for WNV in the United States? Taking a quantitative approach, Kilpatrick and coworkers determined that in New York—where the epidemic began—only two of the more than 170 species of North American mosquitoes—Culex pipiens and Culex restuans—dominated WNV transmission among both birds and humans (Kilpatrick et al., 2005). These
two species were similarly dominant vectors of WNV in the Washington, D.C., area (Kilpatrick unpublished data, 2004−2012); in Colorado, C. pipiens and Culex tarsalis are the primary vectors of WNV (Kilpatrick and Pape, 2013). Such information is crucial to targeted vector control, he observed.
Kilpatrick also noted several reasons for identifying the primary hosts of an introduced vector-borne disease: in order to direct wildlife vaccination or cull host species, should those measures be adopted; to predict hot spots for epidemic disease; and to map temporal-spatial variation in transmission. In the case of WNV, that means identifying which among hundreds of North American bird species are likeliest to transmit the virus. Their quantitative approach led them to the American robin, which proved both highly infectious to WNV, and highly preferred by mosquitoes, he reported (Hamer et al., 2011; Kilpatrick et al., 2006). Thus, he concluded, “It turns out that really there are relatively few mosquito [species] involved in any given place in West Nile Virus transmission, and even relatively few bird [species] involved as well, and I think that’s actually quite good news.” Kilpatrick quickly noted that he and coworkers found that WNV has evolved since its introduction to be transmitted more efficiently by mosquitoes (Kilpatrick et al., 2008), and also to more efficiently infect birds (Duggal et al., 2014).
Urbanization and WNV Transmission
Briefly summarizing a large body of research on factors influencing spatial variation in West Nile transmission, Kilpatrick noted abundant evidence that transmission intensity is higher in urban areas; however, he added, the mechanisms driving that pattern remain to be determined. His own investigations suggest that one reason is an increase of larval habitat for mosquitoes in urban areas, which increases the density of vectors that transmit WNV. Another reason is that vector species in the forests differ from those in urban settings, and the urban vector, C. pipiens, feeds on an especially infective host, the American robin, he stated.
This research also led Kilpatrick to conclude that spatial variation in WNV transmission must be understood on a smaller scale. In the Washington, DC, area, for example, he and coworkers found that mosquito traps placed as close as 100 meters will vary widely both in the number of mosquitoes they capture, and the percentage of those mosquitoes infected with WNV. Based on these observations he concluded, “The proper scale for analyzing transmission is probably in the tens of meters.” Castellanos of the PAHO came to a similar conclusion based on PAHO’s surveys of malaria transmission in Latin America. “We have made the analysis down to a house unit, and we have houses producing malaria repetitively and houses not producing malaria,” he reported. On the other hand, Kilpatrick noted, there is significant variation at higher scales as well—encompassing different types of land uses, such as open fields versus parking lots. It may actually
be possible to predict patterns in WNV transmission at the small scale based on an understanding of both spatial and temporal drivers, he observed.
Environmental Influences on Malaria Transmission
Speaker Matt Thomas of Pennsylvania State University expanded upon the discussion of small-scale variations in vector-borne disease transmission by considering the effects of temperature. Far more immediate than the anticipated effects of climate change, daily temperature ranges and extremes in temperature influence pathogen transmission, he stated, and these effects are tempered by other environmental factors—claims he illustrated with examples from his work on malaria.
Diurnal Temperature Range
Malaria, Thomas observed, is the most important and longest studied of all vector-borne diseases, yet there remain “massive gaps in our knowledge.” Taking what he termed a mechanistic approach, he began his remarks by explaining the components of vectorial capacity, a measure of the transmission potential of a vector population (a specific mosquito species, in the case of malaria). The vectorial capacity equation incorporates variables representing the density of vector species in a given area, their rate of biting and of feeding, their longevity as compared with the developmental period of the pathogen (the malarial parasite), and the degree of vector competence: in this case, how effectively the mosquito picks up the parasite, harbors it and supports its development, and transmits it to the host (humans or experimental animals).
Temperature has long been considered a key driver of malaria transmission because the ecology, physiology, and behavior of the mosquito vector—an ectoderm—are strongly influenced by variations in temperature, Thomas said. All of the previously noted contributors to malarial vector capacity are strongly and differently affected by temperature in nonlinear ways, he reported (Mordecai et al., 2013). Despite this fact, experiments examining the effects of temperature on various aspects of vector capacity have been conducted over a range of temperatures, but at constant temperature within each experimental cohort. For example, he said, researchers measuring the effect of temperature on mosquito development rate might measure that trait in separate mosquito populations in incubators set constantly at 20°C, 25°C, 30°C, and 35°C, to represent the possible range of mean monthly temperatures in the mosquito’s natural environment. However, recent studies comparing malaria transmission within traditional mud huts and modern brick and tin homes in Tanzania found significant differences that were ascribable not to the mean temperature (which was largely equivalent between the two dwelling types), but to the diurnal temperature range, which could be much broader in the tin-roofed huts (von Seidlein et al., 2012).
As a result of this finding, Thomas and coworkers have been studying the effects of diurnal variation on vector competence in the Asian malaria vector Anopheles stephensi. Using incubators programmed to run at a constant temperature, or at that same temperature as its mean, but with variable diurnal highs and lows around it, the researchers monitored the development rate and survivorship of mosquito larvae (Paaijmans et al., 2013). The results, shown in Figure WO-16, reveal that while diurnal temperature variation did not have a significant effect on survivorship under optimum mean temperature conditions, similar variation under high average temperature conditions slows development, he reported—and “the bigger the daily temperature variation the worse things get,” he said. “This is rather an important result,” he added, because these effects would not be discern-able in an experiment that did not feature variations in temperature. Moreover, he added, “You get the reverse effect at the cold end: temperature variation matters there too, but actually it makes things better.” For example, he noted, at a constant temperature of 18°C, few larvae survive, but many more do if the temperature varies diurnally around that mean. Thus, he concluded, “You can’t define the upper or lower limit for this mosquito’s survivorship simply based on
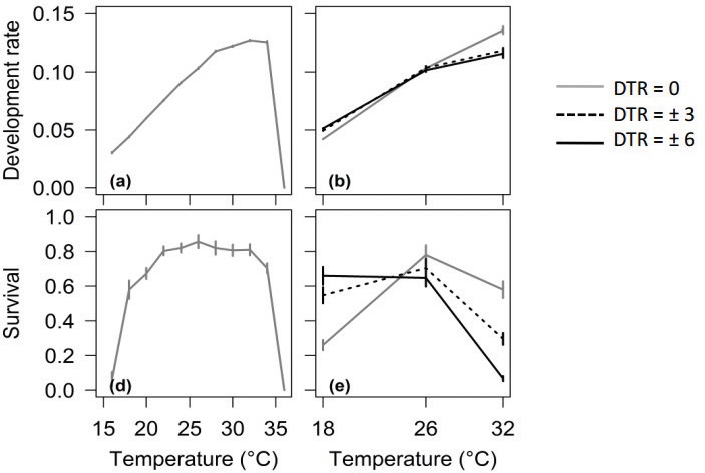
NOTE: DTR = daily temperature range.
SOURCE: © Paaijmans et al., 2013. Published in Global Change Biology, John Wiley & Sons Ltd.
mean temperatures. [Yet] all the models, all the studies that we do, nearly all of them use mean temperatures.” Those include recent, apparently promising attempts to stably infect mosquitoes with the endosymbiotic bacterium, Wolbachia, and thereby interrupt transmission of the malarial parasite to humans (Murdock et al., 2014).
Thomas also shared results of recent experiments examining additional effects of temperature on vector competence in A. stephensi, as well as in A. gambiae, the most important African mosquito vector for P. falciparum, the most important human malarial parasite. Given a meal of infected blood, they asked, how did temperature affect the proportion of mosquitos that became infectious? At the standard temperature for raising mosquito vectors of malaria, 27°C, about half of the mosquitoes of both species became infected, he reported; if the mean temperature increases, fewer mosquitoes become infected. Diurnal temperature range exaggerates this effect, and at high temperatures and broad range, inhibits infection altogether, they found. Thomas and coworkers found a similar pattern of temperature effects on the proportion of A. stephensi that actually harbored sporozoites in their salivary glands following an infected blood meal—the mosquitoes that, Thomas noted, “are the ones that can actually transmit the disease . . . the ones that are going to kill you.” However, in this case, A. gambiae responded far less strongly to both mean temperature and to diurnal temperature variation.
It would be very informative to collect microenvironmental data in many different locations actually inhabited by mosquitoes, rather than in insectaries or other artificial settings, Thomas suggested—for example, within houses. “What’s the temperature in the house? We don’t actually know,” he acknowledged. “We need to go out and do the leg work.”
Larval Nutrition
The quality of habitat available to mosquito larvae can vary greatly within a short time (e.g., whether it is a wet or dry year) and also within a small geographic space, as the result of land use changes that may affect larval nutrition. Little work has been conducted to measure the effects of such variations in larval habitat quality on the ability of adult mosquitoes to transmit malaria, according to Thomas—a question he and colleagues are now attempting to address.
In their recently completed study of A. stephensi infected with P. falciparum malaria, larvae were raised in either “high-food” or “low-food” environments that differed threefold in the quantity of available food. Adult mosquitoes emerging from the “high-food” environment were significantly more likely than the “low-food” adults to be infected with the parasite, particularly at the sporozoite stage, Thomas reported. “That’s just the vector competence component of the vectorial capacity equation,” he noted. The investigators also measured the impact of food availability on additional variables including vector density, biting rate, survival, and how quickly the malaria parasite developed within them. When compared on
the basis of complete vectorial capacity, they found a 45-fold difference in transmission potential between the two groups, favoring the high-food mosquitoes.
Could these findings be applied to vector control? Perhaps, Thomas remarked, in the sense that habitat manipulation strategies—already a keystone of integrated vector management—should target larval habitats. However, he added, the main purpose of these nutritional studies is to better understand how changes in the quality of larval habitats influence patterns of seasonality in malaria transmission.
Small Changes with Large Effects?
In contrast to climate change, with its event horizon measured in decades, the environmental changes Thomas described can happen overnight, he observed—and their impacts on the risk for vector-borne disease transmission can be dramatic. Very few of the many modeling and empirical studies that have been conducted in an attempt to predict future patterns of disease, or to explain current variations in transmission risk, take such short-term effects into account, he observed.
Reflecting on earlier speculation by Kilpatrick that many small-scale effects on transmission either cancel each other out or amount to little more than noise as compared with a very few important influences in a given disease system, Thomas observed that without measuring such effects, we cannot be certain of their magnitude. Moreover, he later noted, “If we’re going to understand local transmission and understand better the consequences of change in a local context, then we really need to think about understanding the local ecology and the sympatric pairings between those vectors and the parasites.”
Perspectives on Disease Drivers
Global Change and Transmission Risk
In the discussion that followed these presentations, and also in comments raised in earlier sessions, several participants expressed concern that disproportionate attention was being paid to the potential influence of climate change on transmission risk for vector-borne diseases, and particularly for West Nile viral disease, where evidence for climate and temperature effects appears particularly thin. “Are we really focusing on the right type of global change by doing so much work on issues around climate change, when really the impact of WNV in the United States may vary with temperature, or it may not?” asked forum member Peter Daszak, of EcoHealth Alliance. Instead, he suggested, it seems that trade routes, travel, globalization, and land use change issues may be the most potent drivers of vector-borne disease transmission over the next two to three decades.
Kilpatrick agreed, asserting that the total number of West Nile viral disease cases driven by changes in either temperature or precipitation is orders of magnitude lower than those driven by changes in land use—and that the same could be said about many vector-borne diseases, because land use changes increase humans’ exposure to biting vectors and, thereby, to disease. Climate does have an impact on transmission, he explained, and climate change may increase transmission in certain circumstances, such as at the geographical limits of vector distribution (Siraj et al., 2014). Mercedes Pascal’s group has been looking for climate links to malaria and other pathogens for a while, and the message that comes relatively clearly from both that work as well as the larger body of work is that climate at the distributional edges of a pathogen or a vector can have a huge role in changing the geographic distribution of a disease, but in the middle of a pathogen’s range it appears that other variables are much more important (Gething et al., 2010; Rogers and Randolph, 2000; Siraj et al., 2014).
So, for example, Pascal’s article was suggesting that at the upper altitudinal limits of malaria, climate could drive it up and down for Lyme disease. There has been some nice work by Nick Ogden and candidates showing that in fact the vector is kind of moving north more in warmer years than other years, and there are a number of cases like that (Ogden et al., 2008).
Many environmental parameters are changing faster than the climate, Linthicum acknowledged, “But I think we have to also keep in mind the long-term impacts of climate change,” he advised. Thomas described his work with climate modelers to attempt to anticipate how transmission pattern effects of diurnal temperature range might change with predicted shifts in mean temperature. While there seems to be a narrow range of responses to a mean change in temperature, it is unclear whether that change will occur equally across the temperature spectrum, he observed. “Perhaps we’ve done a pretty good job with the climate models,” he suggested. Far less is understood about how environmental changes predicted by climate models would affect pathogens, vectors, hosts, and their interactions, Thomas said. Solid empirical data are needed to characterize these relationships.
From Models to Mitigation
Impressed with the variety and depth of models of vector-borne disease transmission risk, and with Linthicum’s Rift Valley fever model in particular, forum member Julie Pavlin of the Armed Forces Health Surveillance Center asked to what extent these models were being translated into actions that benefited public health. The responses she received were mixed. While Linthicum expressed frustration at initially having tried and failed in part to initiate timely responses from public health and agriculture officials to mitigate predicted Rift Valley fever activity that then occurred, they later were able to achieve better communication and ultimately better response. Kilpatrick said that some of his
group’s work had been used by many local health departments to guide vector control activities.
Thomas noted that implementing studies of malaria transmission risk is very challenging. “Early warning systems might be useful in terms of allowing for some level of preparedness,” he explained, but studies of transmission drivers might more productively be used to analyze trends in malaria cases in order to determine the effects of specific preventive measures, such as bed net use on transmission rates, separate from environmental variables, including seasonal precipitation (Aregawi et al., 2014).
What does one do when there is disagreement among multiple models of transmission risk for a vector-borne disease? Both Kilpatrick and Thomas recommended that all models be tested with rigorous local studies. “I personally won’t be confident in the mechanisms we think are driving . . . [variations in vector-borne disease transmission] until we have both broad scale correlational patterns and local-scale studies that support the actual mechanisms,” Kilpatrick asserted. For example, he noted, links between remotely sensed climate and human cases of malaria are well established, but the mechanisms that connect those phenomena have not been defined. “If you can actually open up that black box and show that . . . when we have a higher temperature that does lead to an earlier transmission season, higher mosquito abundance, higher mosquito infection rates, and then more human cases, then I’ll start to believe,” he said. “Short of that I think we’re just waving our hands, and we can get it completely wrong.”
On the other hand, Linthicum noted, such high-resolution information may not be necessary to have a significant impact on public health. In Africa, for example, where Rift Valley fever occurs over very large geographic areas affecting many hundreds of thousands of animals, “There’s no point in becoming very specific; what you need to do is to warn people when those risks are going to be elevated,” he argued, “and then there are a number of things that could be done on a large scale to really mitigate that [threat].”
“One could spend forever doing elegant research with exquisite temperature fluctuations in the lab,” Thomas imagined, “but ultimately we need to get that out in the real world, we need to have it . . . inform practice.” In the real world, there will be broad patterns and significant variation and context dependence, he observed. Only by examining what actually happens, by analyzing case histories, can we discern the most important drivers of transmission risk in a given situation. He therefore advocated in favor of focal studies of disease transmission at sentinel sites, with the goal of trying to identify and understand the drivers involved and their interactions, and to gauge the effectiveness of possible interventions. “The best way of progressing is to learn by doing,” Thomas insisted.
NOVEL APPROACHES AND INTERVENTION STRATEGIES
In the final workshop session, speakers reviewed past efforts to address vector-borne diseases and described a range of strategies and methods to tackle key obstacles in their prevention, diagnosis, and treatment.
History and Current Challenges of Dengue Vector Control (Or, Why Did Gorgas Succeed? And Why Have We Failed?)
Vector control has an important role in addressing vector-borne diseases, observed speaker Paul Reiter of the Institut Pasteur. In recounting the history of this approach, he began with Carlos Finlay’s hypothesis that yellow fever is transmitted by Aedes aegypti. This was experimentally confirmed on the arrival of Walter Reed. With this knowledge, William Gorgas19 had spectacular success in eliminating yellow fever from Havana, Cuba, and later during the construction of the Panama Canal. Fred Soper, director of the Pan American Sanitary Bureau (later PAHO) between 1947 and 1959, implemented Gorgas’s approach throughout Latin America (Johns Hopkins Bloomberg School of Public Health, 1991)—a contribution to public health that Reiter deemed “quite exceptional.”
Fast-forward to the mid-1970s in Singapore. From the late 1960s until the mid-1980s, dengue—also transmitted by Ae. aegypti and once a major cause of illness in that country—had been drastically reduced by vector source reduction but began to rise thereafter until it became a major public health problem once again. As consultant to the Singapore government, Reiter hypothesized that suppression of the disease had been so successful that half the population was now nonimmune; i.e., the herd immunity had been greatly reduced in the host populations so that, even in low numbers, mosquitoes were now more efficient in transmission. “We made certain recommendations, and things looked like they were getting better,” he recalled—until a massive epidemic struck in 2005. Again Reiter was consulted, along with Duane Gubler (“sort of the emperor of dengue epidemiology worldwide”), and after their suggestions were implemented, case numbers declined—“until 2013, when, despite major control efforts, dengue suddenly took off again,” he reported.
Singapore spends some $60 billion a year on Ae. aegypti control yet incidence continues to rise. For decades, government sources state that Havana (about the same size as Singapore) was the only country in the New World that was free of dengue but despite official statistics it is well known that dengue is rampant there. In truth, there is nowhere on this Earth where dengue is under control. . . . If they can’t do it in Singapore, no one is going to be able to do it with the weapons that we have at present.
___________________
19 Later Sir William Gorgas. Although an American citizen, he was knighted by King George IV for his achievements and given a funeral in St. Paul’s Cathedral upon his death.
Why not? Reiter outlined his response in the form of this poem, entitled “Ode to Ae. aegypti Control”:
Those golden days . . . .
The perfect way . . . .
Let us-(s)pray
The mess today!
Source Reduction
The “golden days” began with Gorgas, whose military pursuit of Ae. aegypti through “source reduction”—the elimination of breeding sites—purged Havana of yellow fever within 5 months, Reiter stated. Mosquito habitats were eliminated by various means: water storage vessels and wash basins were covered, water in horse troughs was exchanged regularly, gutters were made to drain properly, and so on. The results were certainly impressive but it is important to note this was an entirely different era from today: breeding sites were much less common; there were few motor vehicles, so used tires, a classic mosquito breeding site, were absent; cities were much smaller; and there were no plastics or other disposable items that could serve as water collection breeding sites. Moreover, fear of yellow fever increased acceptance of the intrusive measures used to enforce control, he added.
When Gorgas was subsequently assigned to combat yellow fever and malaria during construction of the Panama Canal (mosquito-borne disease was an important reason why the French canal project had failed), he first attempted to rely on the insecticide pyrethrum to do the job, Reiter said. Three attempts failed before he returned to source reduction, with which he finally achieved success.
Insecticides
Insecticide treatment would not become “the perfect way” to combat mosquito-borne diseases until after World War II, when DDT became available. Soper employed it to eradicate Ae. aegypti from 22 countries in less than 10 years, or so it has been claimed, Reiter noted. Certainly, dengue and yellow fever transmission was brought to a halt.
With the banning of DDT, beginning in the 1960s, a new era of insecticide-based vector control began. While DDT had been applied directly to infested containers and their immediate (50 cm) perimeter, post-DDT insecticides were broadcast as aerosols—“let us spray”—by hand-held foggers, road vehicles, or aircraft, Reiter stated. Ae. Aegypti, however, is an indoor mosquito; this, among other behavioral traits, may explain why spraying has not effectively reduced the diseases they carry (Reiter, 2007). Nevertheless, he pointed out, “There’s a noisy machine with a nasty smell with a big loud noise and the flashing lights.
So that really persuades people that they’re being looked after.” But, he added, “the bottom line is that many countries are trying to control dengue, and they fail . . . if you look at any of the public health data it’s absurd to say that we are actually controlling it.”
Now What?
Our cities are huge, human populations are dense and mobile, public health funding is scarce—“It’s a perfect paradise for the mosquito,” Reiter lamented. It is often difficult to access areas that should be treated to reduce populations of Ae. aegypti, insecticide resistance is a problem, and public participation in cleanup campaigns is inadequate to achieve source reduction, he noted. How does one face “the mess today?”
There are a lot of things we don’t know about the biology and ecology of Ae. aegypti, Reiter observed, and that missing information may provide routes to effective vector control. For example, he said, “We don’t know how many [water] containers we have to reduce in order to stop transmission, how many mosquitoes, the comparative economics of the different approaches to control, or their sustainability.” According to his group’s mathematical models, a 90 percent reduction in the numbers of mosquitos would still produce very little on the overall transmission rate for dengue.
The same model demonstrated that only a few mosquitoes could effectively transmit dengue in a human population that has low herd immunity (see Figure WO-17). This, he concluded, reveals why Gorgas and Soper succeeded where we are now failing: their effective reduction of the vector occurred in human populations with high herd immunity (Reiter, 2014).
If that is indeed the key to successful dengue control, we need to better understand how to build and exploit herd immunity, Reiter insisted. “I don’t believe that vector control on its own is going to be the answer, even though I’m a medical entomologist. But I do believe that augmentation of the herd immunity by vaccination, in combination with vector control, may prove more effective than either approach on its own.”
Reiter was emphatic that new and novel approaches to vector control are urgently needed. These may include a return to focal insecticide application; the use of Wolbachia to reduce mosquito infectivity, as mentioned by Thomas; and the use of juvenile hormone mimic—a compound that disrupts mosquito metamorphosis, and which can be distributed by female mosquitoes among multiple breeding sites. In his opinion, however, the method that shows the most promise is based on transgenics: males of a strain of Ae. aegypti that carry a dominant lethal gene are released to mate with “wild” females but the gene ensures that the resulting progeny cannot survive to adulthood. Studies in a number of countries have demonstrated remarkable results.
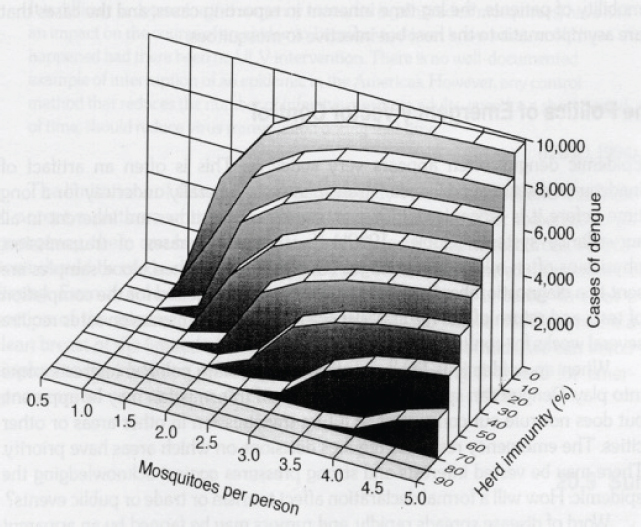
SOURCE: Reiter, 1992. Reproduced with permission from CAB International.
The Public Health Imperative for Improved Ae. aegypti Control
According to Barry Beaty from Colorado State University, “it has been humankind’s great misfortune to share time and space with Aedes aegypti (see Figure WO-18). Following domestication of this mosquito in Africa, it traveled with humans throughout the tropical and subtropical world (Powell and Tabachnick, 2013). Beaty describes Ae. aegypti as the Norway rat of mosquitoes. Because of its intimate association with humans in homes, schools, and work places and its extreme preference for feeding on humans, it is an excellent vector of yellow fever, dengue, and chikungunya viruses, and this mosquito has caused and continues to cause inestimable morbidity and mortality in humans (García-Rejón et al., 2008, 2011). The same biological and behavioral attributes that allowed domesticated Ae. aegypti to colonize the tropical world via sailing ships have served the vector well in the modern era.
Aedes aegypti is uniquely adapted to the modern urban environment, exploiting new breeding sites such as septic systems and storm drains, and it has proven intractable to sustainable control in large urban areas. Classically used control methods such as environmental source reduction and space spraying have not
sustainably stemmed the pandemics of dengue and chikungunya. Indeed, Ae. aegypti is essentially now hyperabundant throughout the tropical world, and notably in areas where it had previously been eliminated or greatly reduced in abundance (Gubler, 2011). Large urban areas, with their sizeable Ae. aegypti populations, are now very receptive to the introduction, spread, and trafficking of arboviruses (see Beaty et al. in Appendix A3). The introduction of chikungunya virus in 2013, and its explosive spread throughout Latin America (Nasci, 2014) in a very short time, is testimony to the epidemic potential of Ae. aegypti–transmitted pathogens. Unfortunately, our ability to intervene in such epidemics is likely to worsen.
The alarming and rapid emergence of pyrethroid resistance in Ae. aegypti threatens the efficacy of many of the chemical control efforts for this important vector. The emergence of knockdown resistance (kdr) in Ae. aegypti in Mexico (Garcia et al., 2009) has been mirrored in Ae. aegypti throughout much of the tropical and subtropical world. Similarly kdr has exploded in Anopheles gambiae threatening the efficacy of long-lasting insecticide-treated nets for malaria control. Pyrethroids are also the insecticide of choice for control of vectors of other globally important diseases such as Chagas, lymphatic filariasis, and leishmaniasis, and they are widely used by public health agencies for control of other insect vectors, such as for those that transmit West Nile virus, and other insect vectors of globally important pathogens. The evolution of kdr has been associated with dramatic increases in metabolic resistance in important vector species. Evidence

SOURCES: James Gathany/CDC, 2006.
is accumulating that pyrethroid resistance is becoming operationally significant, which could lead to the loss of this key class of insecticides in the armamentarium for vector control.
According to Beaty, this would be a public health catastrophe on the order of emerging antibiotic resistance in bacteria and parasites. For the foreseeable future, chemical insecticides will remain critically important for controlling vector populations. Development of new, environmentally sensitive insecticides with new modes of action from existing insecticides is a public health imperative. One novel approach in this regard has been the development of the Innovative Vector Control Consortium (IVCC). The IVCC partners with industry to develop new insecticides with different modes of action than pyrethroids, which will permit rotational or mosaic applications of insecticides to minimize development of resistance and to provide improved stewardship of existing and new insecticides. New insecticides as well as new innovative strategies for vector control are needed, including insecticide resistance blocking strategies (Devine et al., 2009), Wolbachia interruption of DENV transmission by Ae. aegypti mosquitoes (Moreira et al., 2009), genetic strategies to reduce vector populations (Black et al., 2011), and other innovative approaches to be vigorously pursued to prevent and control Ae. aegypti transmitted diseases. The burden of these diseases is too great to bear, and it is clearly time to declare “war” on Ae. aegypti and to sustainably control this enemy of humankind.
Strategies for Malaria Eradication
As recently as 1998, it seemed quite unrealistic that malaria could be eradicated, according to speaker Alan Magill of The Bill & Melinda Gates Foundation. That year, malaria killed two million people and infected half a billion. Resistance had developed to key therapeutics, and insecticide-treated bed nets—a preventive measure recently proven in clinical trials—were not yet widely distributed. Epidemic HIV in East Africa led to co-infection, increasing the burden of both diseases.
By 2013, however, the picture had changed drastically (White et al., 2014). As compared with 1998, acute malaria cases had declined by about 60 percent, and mortality by nearly 70 percent, Magill reported. Massive increases in donor funding spurred the implementation of new preventive measures—long-lasting insecticide-treated bed nets and indoor residual insecticide spraying—along with rapid diagnostic tests and targeted interventions for vulnerable populations. This turnaround demonstrates how adequate resources in support of effective tools can produce significant gains in public health, he concluded—and it offers hope that the next step, the eradication of malaria, can be achieved by both applying current interventions continuously and at high coverage, and by applying current interventions with new strategies. “There’s really no backing off on this,” he insisted.
But Magill also noted daunting challenges to continued progress against malaria, including emerging resistance to drugs and insecticides, and maintaining current levels of support of funding required for malaria eradication. The disease has been controlled many times in many locations, but resurgence predictably has occurred because efforts were not sustained (Cohen et al., 2012). Therefore, he argued, global eradication is the only permanent solution for malaria. Moreover, he pointed to evidence that complete eradication has been stable in 46 out of 50 countries where it has been achieved (Smith et al., 2013). “The trick here is getting to zero,” he emphasized. “What often happens is you get to very low levels, you have many residual pockets of transmission, and if you don’t actually finish the job then resurgence is pretty much inevitable.” Taking that final step from control to eradication is both difficult and expensive, he added, “but if you really do get parasites out of people . . . then maintaining that state is a little easier.”
Interrupting Transmission
In 2007, Melinda Gates described the foundation’s commitment to eradicating malaria as a quest for equity. “It’s about a recognition that those areas of the world that suffer from malaria really can’t get ahead,” Magill explained. The debate as to whether poverty causes malaria or vice versa isn’t useful, he added, since eradicating malaria will surely advance some of the world’s poorest people.
Malaria has three possible futures, Magill observed: resurgence, control (sustaining and slowly improving progress against the disease to date), or accelerating toward eradication. To achieve the latter outcome, the Gates Foundation has defined a strategy called Accelerate to Zero, intended to focus current and future tools in an intensive effort to interrupt malaria transmission. The cornerstones of this strategy are the detection of the human parasite reservoir, the elimination of that reservoir, and the effective prevention of transmission, he explained. “If you can’t cure people and prevent transmission concurrently, then both of those approaches will ultimately fail,” he insisted.
Because the malaria parasite biomass resides almost entirely in humans, it must be diagnosed and treated in infected people in order to be reduced, Magill stated. Thus the Gates Foundation supports current efforts to test, treat, and track malaria infections. Every person who presents with a fever in a malaria-endemic country should have a reliable diagnostic test, and if proven to be infected,20
___________________
20 In the discussion that followed this session, forum member Lonnie King of The Ohio State University asked what would be done for those patients who tested negative for malaria. Magill responded that unfortunately, there are not good point-of-care diagnostics for other common febrile diseases, such as Q fever and leptospirosis; however, an initiative is under way to address nonmalarial febrile diseases collectively, with antibiotics for suspected or confirmed bacterial infections, and supportive care in other cases. Moreover, he said, even people who test positive for malaria may actually be suffering from another infection. Forum member Gerald Keusch, of Boston University, noted that a rapid diagnostic test capable of differentiating between malaria and pneumonia in children was shown to reduce the use of antimalarial drugs by about two-thirds, with excellent survival rates.
get treatment with best available therapy, he said—and then, their case should be tracked along with others in a surveillance system. However, not all malarial infections are symptomatic, he noted; “This is the classic iceberg . . . there is a vast reservoir of infected people out there in the community who are happily carrying their parasites and their gametocytes, and they are going on in this uninterrupted circle of transmission with their mosquito vectors, and we’re doing absolutely nothing about that today” (Lindblade et al., 2013). To address this problem, the Gates Foundation advocates the complete cure of asymptomatic fathers and mothers as a way to save their children’s lives. This will demand new rapid diagnostic tests that can identify asymptomatic people in communities, as well as mass “screen and treat” campaigns, he said.
These efforts will be most effective if they are targeted toward communities that serve as sources for widespread malaria transmission, Magill continued. The Gates Foundation supports efforts to map malaria transmission patterns, analyses to determine the most strategic areas to focus treatment efforts, and the creation of databases for use by ministries of health in affected countries, he reported.
The goal of malaria eradication is the interruption of transmission, Magill emphasized. But today, while many people with malaria get treated and recover, they may continue to carry viable P. falciparum gametocytes for the next 4 to 8 weeks, and therefore continue to transmit the disease. “There’s never been a single attempt to actually interrupt transmission by targeting the gametocyte,” he asserted. “What we need is what we call complete cure, which is complete parasitologic cure. We need a drug regimen that will not just make you better . . . but we also need them to get rid of the parasites that transmit.” Thus, he said, the Gates Foundation is working with partners to develop drugs that will kill malaria gametocytes. Two lead candidates “have extremely significant and very promising transmission-blocking and gametocidal effects,” he reported; ultimately, it is hoped that they can be delivered as a single pill, along with drugs that cure clinical disease—and that this would be achieved for both P. falciparum and P. vivax malaria.
Situational Solutions
In 1937, malariologist Lewis Hackett observed,
Everything about malaria is so molded and altered by local conditions that it becomes a thousand different diseases and epidemiological puzzles. Like chess, it is played with a few pieces, but is capable of an infinite variety of situations.
Recognizing the enduring truth of this depiction, the Gates Foundation supports a variety of means to tackling malaria, and to applying them as targeted, locally adapted solutions, Magill explained. For example, despite the deployment of insecticide-treated bed nets and indoor residual spraying of insecticides, significant residual malaria transmission occurs as mosquito populations adapt both
genetically and behaviorally, he reported (Killeen, 2014). Vector control could, in theory, reduce these problems, but rigorous field trials are rarely conducted on these measures, he noted (Vontas et al., 2014). To meet this need, the Gates Foundation initiated and supports the UK-based IVCC,21 a group of experts who partner with agrichemical companies worldwide to develop novel insecticides to address a broad range of vector-borne diseases.
The pairing of insecticide-treated bed nets with artemisinin-based combination treatment for malaria has proven extremely effective in reducing cases, as shown in a long-standing epidemiological study in Senegal (Trape et al., 2014). However, resistance to artemisinin has emerged in Southeast Asia (Ashley et al., 2014), while in Haiti, where the primary vector for malaria is the outdoor-dwelling mosquito Anopheles albimanus, bed nets are not a useful preventive measure. There, in partnership with PAHO, The Bill & Melinda Gates Foundation is working to eradicate the disease through the identification of transmission hot spots, with a combination of focal indoor residual insecticide spraying and drug treatment to eliminate the human parasite reservoir, Magill said.
The Gates Foundation also supports vaccine development, but with the emphasis on preventing infection to interrupt malaria transmission, rather than targeting disease prevention, Magill stated. An antidisease focus for vaccination could actually enable the continuation of asymptomatic parasitemia, he asserted.
In conclusion, Magill emphasized that eradicating malaria will require new concepts, tools, and strategies, and an end to a “one size fits all” approach to addressing this complex and varied disease. “The next decade will be a period of intense experimentation and learning, leading to a rapidly evolving policy environment for new tools and technologies,” he predicted.
Transgenic Insects for Vector Control
Effective control strategies for arboviral diseases can target their insect vectors, observed speaker Luke Alphey of the United Kingdom’s Pirbright Institute. He described work under way to create and deploy genetically modified Ae. aegypti mosquitoes, designed to reduce dengue transmission by this species, which also transmits the chikungunya and Zika viruses (see Alphey et al. in Appendix A2).
The number of people becoming infected with dengue each year is now approaching that for malaria, Alphey noted, although far fewer people die of dengue. There are no drugs that specifically treat it, nor are bed nets effective deterrents, since the mosquitoes that carry the virus do not bite at night. Thus dengue control has focused on reducing mosquito populations—with varying degrees of success, as previously described by Reiter.
___________________
21 See www.ivcc.com (accessed March 25, 2016).
As an alternative to vector control through source reduction or insecticide treatments, genetic strategies involve changing the mosquito genome so as to spread a repressible developmental defect that would interfere with reproduction or a modification that would interfere with disease transmission, Alphey explained (Alphey et al., 2010). This is not a new idea, he noted: irradiated sterile insects have been used for more than 50 years, on very large scales, to control several agricultural pests. Since then, several additional genetic control strategies have been developed that could be used to combat dengue or other mosquito-borne diseases (see Figure WO-19).
There are two possible goals of genetic control strategies, Alphey explained: to reduce the size of a vector population or to change it in a way that reduces disease transmission—that is, by somehow reducing vector competence throughout the population. The genetic changes introduced to accomplish these goals may either be self-limiting—they will eventually be washed out by lethality or by natural selection—or they may be self-sustaining, becoming established in the target vector population and possibly spreading to other populations, he added. Genetic control strategies share several key features, Alphey stated, including the fact that they protect every person within the area where they are deployed. By way of contrast, human-directed disease control programs often disproportionately favor the wealthy, powerful, and educated. All genetic control strategies exploit insect mate-seeking behavior to disperse the control agent, and also to ensure species-specific effects. Thus, he reassured the audience, “We are not talking about eliminating all mosquitoes.”

NOTE: HEG = homing endonuclease gene; RIDL = release of insects carrying dominant lethal.
SOURCE: As presented by Luke Alphey on September 17, 2014.
RIDL Technology
Alphey’s presentation focused primarily on a genetic control strategy known as “release of insects carrying dominant lethal” (RIDL), which features genetically engineered male mosquitoes carrying repressible dominant lethal transgenes that are released to mate with wild females, producing doomed progeny (Alphey, 2014b). For example, he described one transgene with a promoter that is expressed only in female flight muscle tissue and which, when passed to the female offspring of genetically engineered male mosquitoes, compromises the females’ ability to fly (Fu et al., 2010). “Of course flightless mosquitoes can’t survive in the wild,” he observed. “They can’t move away from the breeding site, they can’t find a host, they can’t avoid predators. Actually they can’t mate, even in the laboratory.” Male offspring, on the other hand, can fly and mate, and thereby spread the lethal gene through the population.
Alphey reported that another type of RIDL system—in which the repressible transgene kills all of the progeny from the engineered father and wild mother (Phuc et al., 2007)—has proven effective in suppressing Ae. aegypti populations throughout a series of phased tests, culminating in a successful field release in the Cayman Islands, which achieved an 82 percent reduction in the target mosquito population (Harris et al., 2012). The researchers have also conducted field trials with RIDL mosquitoes in Malaysia, Brazil, and Panama, he added. In Brazil, Alphey observed, target mosquito populations in different settings were reduced by 94 to 99 percent.
To gain public support for these potentially controversial experiments, the researchers used informal presentations in a variety of settings—door-to-door visits, television, radio, and print media—to explain their work and to demonstrate its safety. “Dengue control is widely recognized as desirable, and it’s also recognized that current methods aren’t adequate and new methods are needed,” Alphey said, explaining the high levels of public approval these projects received. At the time of the workshop, a proposal for a field trial in the Florida Keys was under consideration, he added. While there has been some objection to this trial by environmental activists, an independent pollster found that 61 percent of residents questioned supported the use of genetically engineered mosquito technology, as compared with 18 percent who did not (Florida Keys Mosquito Control District, 2013). Moreover, 81 percent of respondents considered genetically engineered mosquito technology safe, as compared with 73 percent who considered the use of chemicals and insecticides to be safe.
Appropriate Application
While not a “magic bullet,” RIDL technology is appropriate for certain disease vectors in certain settings, Alphey said. For example the two mosquito species Ae. aegypti and Ae. albopictus (for which RIDL technology has also been developed)—which spread DENV, CHIKV, and Zika, among other pathogens—could be
specifically targeted with the technology. Ae. aegypti eggs are dry and easily stored, he added. “They could just be shipped out in packets for people to rear at the bottom of their yard.” Likewise, Kilpatrick’s narrowing of the mosquito species most likely to transmit WNV suggests that RIDL could also be brought to bear on that disease.
Because Ae. aegypti and Ae. albopictus are alien invasive species in the Americas, “If you could eliminate them, then that might be seen as ecologically desirable rather than undesirable thing,” Alphey observed. Indeed, he added, when Ae. aegypti was eliminated from about 20 South American countries during the DDT era, no adverse ecological effects were reported. On the other hand, a native vector species could serve an important ecological role; in that case, he advocated using methods that make vectors less able to transmit pathogens but without reducing their populations. “There are different options, and you look at this on a case-by-case basis,” he concluded.
Exploiting Virus–Vector Interactions
Arthropod-borne plant pathogens—and plant viruses in particular—present a major threat to global food security, according to speaker Anna Whitfield of Kansas State University. The viruses she studies, tomato spotted wilt virus (TSWV) and maize mosaic virus (MMV), are related to viruses that infect animals, and their transmission cycles resemble those of other vector-borne animal and human viruses such as CHIKV, she noted. TSWV and MMV are acquired by their insect vectors when they feed on infected plants. The viruses then infect the insect’s gut, and eventually move to its salivary glands, where they replicate, and from which they are transmitted to naïve host plants during feeding (Blanc et al., 2014).
Each step in this transmission cycle can potentially be disrupted, Whitfield said; thus her group and other researchers are exploring many opportunities to control plant disease (see Whitfield and Rotenberg in Appendix A12). “We don’t view these strategies as a silver bullet,” she explained. Managing plant diseases generally requires an integrated approach; therefore, she characterized the methods she described as “just another tool in the toolbox for plant production.”
Viral Acquisition
TSWV, a Bunyavirus, is related to RVFV and other members of the genus Hantavirus, Whitfield noted. TSWV is globally distributed, has an exceptionally wide host range of more than 1,000 plant species, and annually contributes to more than $1 billion in losses of crops that include tomatoes, peanuts, and peppers. There is no single effective control strategy for TSWV, as it easily overcomes genetic resistance bred into crop plants, and its thrips vector develops resistance to pesticides. By pursuing the interruption of TSWV transmission by molecular means, she and coworkers are trying to provide another tool to add to their current integrated pest management system.
Whitfield described a glycoprotein known as GN that projects from the membrane surface of TSWV and mediates viral attachment to the thrips vector’s midgut tissue. The researchers made GN in soluble form, fed it to thrips, and found that it not only specifically bound their midgut tissue, but that it inhibited TSWV transmission—presumably by blocking its binding site, she explained. They then made transgenic plants that expressed the GN protein, infected them with TSWV, and let thrips feed on them; compared with insects fed on equivalent nontransgenic plants, the thrips fed on GN plants had significantly lower TSWV titers and rates of viral transmission, she reported (Montero-Astúa et al., 2014). The transgenic GN plants sustain an initial infection with TSWV, she said, but because they are expressing the viral attachment protein, they block subsequent transmission of the virus. “We think that these could be a promising tool for control of TSWV spread from secondary infection,” she concluded.
A similar strategy has been demonstrated in another vector-borne plant virus system, Whitfield noted. Bonning and coworkers (2014) produced transgenic plants that expressed a soluble luteoviral coat protein fused to a spider toxin. Aphids fed on these plants internalized the chimeric protein as if it were a virus, delivering the toxin to its body cavity and killing the insect. This approach provides much-needed options for controlling aphids, a major transmitter of plant viruses—and it could be applied to other virus–insect systems, employing a range of potential toxins (Whitfield et al., 2014).
Viral Dissemination and Transmission
Whitfield’s laboratory is also exploring vector proteins that interact with TSWV and MMV, enabling these viruses to traverse multiple barriers and reach the vector’s salivary glands, where they replicate and from which they are disseminated. The researchers have identified a suite of proteins consistent among members of the Rhabdovirus genus like MMV and among members of the Tospovirus genus like TSWV that appear to interact with or respond to the insect vectors of several types of plant viruses, she reported. “These are the type of proteins that we would like to follow up on by targeting and disrupting vector acquisition and transmission,” she stated.
RNAi for Vector Control
Viral-vector interactions could be further exploited as a means of vector control through the use of RNA interference (RNAi),22 Whitfield observed (Kupferschmidt, 2013). Research in several different insect species has
___________________
22 The term RNA interference was coined to describe a cellular mechanism that uses the gene’s own DNA sequence to turn it off, a process that researchers call silencing. In a wide variety of organisms, including animals, plants, and fungi, RNAi is triggered by double-stranded RNA (dsRNA). http://www.umassmed.edu/rti/biology/how-rnai-works (accessed August 12, 2016).
demonstrated the effectiveness of this method for silencing expression of the crucial enzyme, vacuolar ATPase (v-ATPase) (Yao et al., 2013). When Whitfield and coworkers delivered double-stranded RNA fragments either orally or by microinjection to nymphs of the insect vector of MMV, the corn planthopper Peregrinus maidis, it experienced higher rates of mortality, as well as reduced egg production (apparently due to abnormal development of female reproductive organs). Similar decreases in survival and fecundity were achieved with thrip vectors when treated with dsRNA of v-ATPase (Badillo-Vargas et al., 2015). Studies are currently under way to treat plants, including crops threatened by citrus greening disease, with double-stranded RNA to control its insect vector, the Asian citrus psyllid (Diaphorina citri).
RNAi techniques have also been developed to control arthropod vectors of animal viruses, Whitfield noted. For example, Kang and coworkers (2014) showed that silencing v-ATPase and another gene (inhibitor of inosine-5′monophosphate dehydrogenase) in Ae. aegypti not only reduced mosquito survival and egg production, but it also suppressed host factors for DENV-2, thereby decreasing infectivity. The successful results with RNAi to silence the v-ATPase genes of plant and animal disease insect vectors highlight not only the similarities in basic vector biology but also that of emerging vector control strategies. These commonalities suggest that increased communication and collaboration between investigators working with plant and animal vectors could be beneficial for the control of vector-borne diseases.
Outlook for West Nile and Chikungunya Vaccines
Vaccines represent the most cost-effective means of controlling many infectious diseases, and they offer significant prevention against zoonotic vector-borne diseases for which humans serve as a dead-end host, observed speaker Thomas Monath of Hookipa BioTech AG and PaxVax, Inc. Repeated vaccination of humans can prevent them from contracting such diseases, which he said cannot be eradicated due to the size or persistent infection of animal reservoirs.
Figure WO-20 provides an overview of the state of vaccine development for vector-borne diseases. “Yellow fever vaccine is probably the closest to a silver bullet that we have,” Monath said. “The Nobel Prize was awarded for it, and it really has driven that disease nearly to extinction, though it still remains a threat.” He highlighted vaccines against WNV and Japanese encephalitis virus as good examples of prevention against diseases that pose an ongoing threat to human hosts. By contrast, he noted, vaccines against vector-borne pathogens that can be transmitted by humans, such as dengue and chikungunya, could potentially contribute to the eradication of those diseases.
Focusing on vaccines to prevent West Nile viral disease and chikungunya, Monath suggested that lessons learned in the process of developing the WNV vaccine could inform the development of vaccines against other emerging

NOTE: TBE = Tick-borne encephalitis; VEE = Venezuelan equine encephalitis.
SOURCE: As presented by Thomas Monath on September 17, 2014.
pathogens—most notably CHIKV, with which WNV shares several common threads, as shown in Box WO-2.
A Vaccine Against WNV?
The first vaccine initiative against WNV started within months of its emergence in New York in 1999, Monath recalled. “The first vaccine for horses was approved in 2001,” he said. “By that time there were at least six companies engaged in vaccine development for humans,” he added—all of which have since halted their efforts. Today, only the National Institutes of Health (NIH) continues to pursue a vaccine against WNV—and without industry involvement, he expressed doubt that such a candidate would reach advanced stages of development.
Technical obstacles to developing a WNV or CHIKV vaccine are minimal, Monath stated. “These are fairly straightforward targets.” The real problems, he continued, are economic: uncertain market size, high development costs, a challenging regulatory pathway, and the risk that the disease will be controlled by natural immunity before the vaccine is developed (a possibility for any emerging infection, he pointed out, and for chikungunya in particular).
Given these hurdles, who is developing vaccines for emerging diseases? “It’s mostly biotech companies,” Monath said. “They’re cash constrained,” he noted, and to pursue a vaccine, they must invest a significant amount [of capital] without guarantee of return—which tends to discourage investors. Due to this reality, small companies with candidate vaccines have partnered with large ones to defray the expenses associated with late-stage development. Large pharmaceutical companies, Monath observed, have limited interest in vaccines. “They were, as you saw, engaged in West Nile at the beginning, but they’ve put a tiny fraction of their treasure to work on these kinds of targets, and they’re basically watching and waiting to see what happens,” he noted.
Monath was part of the effort to develop a WNV vaccine at his former employer, Acambis, now owned by Sanofi Pasteur. Their product, ChimeriVax-WN, is a live, attenuated, virus vaccine based on the yellow fever vaccine, which they produced within months of the 1999 outbreak (Dayan et al., 2013). A single inoculation provided long-lasting immunity in nearly everyone who received it, he reported. It was shown to be safe, even in elderly people, and it couldn’t be transmitted by mosquitoes (and thus would not stimulate antivector immunity). “It really looked like an ideal vaccine,” he observed.
In 2001, Acambis began advanced preclinical studies in nonhuman primates and demonstrated the vaccine’s effectiveness in horses, he said. The horse vaccine was licensed in 2002 to a large company, Intervet. With Phase 1 clinical trials completed, ChimeriVax-WN was manufactured at final scale by 2004—but by then, the epidemic had peaked, and soon afterward, market surveys revealed
a public wary of a live viral vaccine for a disease they and their physicians perceived as low risk. Acambis also learned that primary care physicians, who would be primarily responsible for administering the vaccine, would likely not be mandated to provide it. Nevertheless, in 2007, Sanofi licensed ChimeriVax-WN.
The next year, Sanofi bought Acambis. Development of ChimeriVax-WN stalled during the next 2 years of declining disease incidence and unfavorable market and regulatory conditions, Monath recalled. In 2010, Sanofi ceased further development of the vaccine. Other companies made the same decision, he noted. By the time of the next WNV outbreak in 2012, few vaccines were in development. “Arbovirologists understood that this was going to come back and we would have a problem, and it would be nice to have a vaccine as part of the armamentarium against a reemerging disease,” he observed, “but it’s very hard to rekindle or restart a program that’s become dormant in a company.”
“I think we can conclude that if past is prologue, we’ll have more episodes like 2012,” Monath continued. Nevertheless, taking into account a variety of regulatory issues and the cost of a Phase 3 trial, Sanofi concluded that it still did not make financial sense to develop the vaccine. Given that decision, should the U.S. government fund the development of a WNV vaccine stockpile in order to respond to future outbreaks? Even a stockpile restricted to immunizing the elderly, who are most susceptible to neuroinvasive disease, would cost about $156,000 per case averted, Monath warned.
Moreover, he wondered, “How are you going to use a vaccine like this in an emergency? These epidemics tend to evolve quite quickly . . . [and] in the United States we really don’t have any experience with mass immunization campaigns, especially those involving adults.” On the other hand, a growing burden of long-term symptoms associated with WNV could tip the scales to justify further vaccine development, Monath observed (Garcia et al., 2014; Maxmen, 2012; Murray et al., 2014).
What About Chikungunya?
Fifteen years after the emergence of WNV in the United States, the threat of chikungunya is déjà vu, Monath observed (Morens and Fauci, 2014). As was illustrated in Box WO-2, the two diseases are quite similar in terms of epidemiology, clinical impact, and apparent long-term sequelae (Weaver et al., 2012). However, there are some important differences in the challenges these viruses present for vaccine development, he added. For example, humans are a dead-end host for WNV, but they participate in the transmission of chikungunya. “I think that could actually help us develop a vaccine, speed up the clinical development in part, and be a driver for an intervention,” he observed. The rarity of asymptomatic CHIKV infections as compared with WNV could also expedite vaccine development, he added.
Much has occurred with the emergence of WNV in the United States: the emergence of CHIKV in the Americas has spurred several companies and government institutes to pursue development of a vaccine using a broad range of approaches, as shown in Figure WO-21. NIH researchers have produced a CHIKV-like particle that so far has proven safe and immunogenic, Monath reported (Akahata et al., 2010; Chang et al., 2014). “This is a real potential product, and I think there are a number of companies interested in licensing it,” he said.
As with WNV, the obstacles to developing a CHIKV vaccine are not technical, but economic, Monath said. Once again, key issues involve target population, regulatory pathway, timing, and funding sources. Chikungunya’s dramatic emergence has raised the profile of a potential vaccine, as does the likelihood that CHIKV will persist for decades in a large human population with abundant mosquito vectors, through which many travelers pass, he noted. There is also the potentially significant burden of chronic, long-term disease. Whereas the WNV vaccine was relatively feasible but had low market interest, a CHIKV vaccine, though potentially more difficult to develop, could have a larger potential market both in the United States and worldwide if chikungunya continues to spread, Monath predicted.
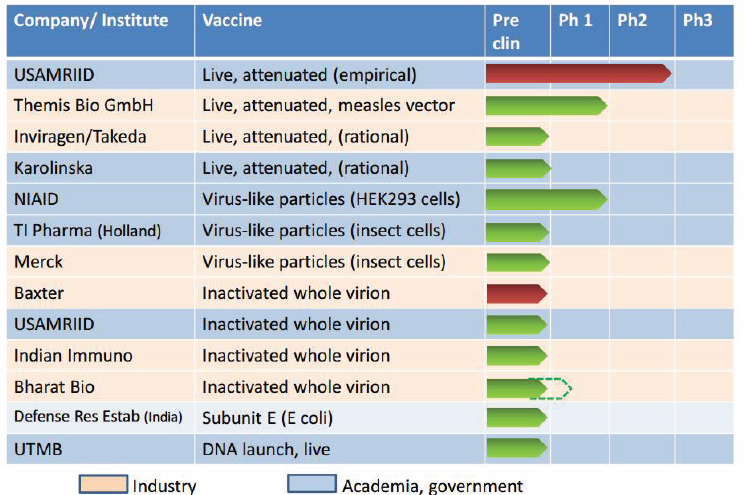
SOURCE: As presented by Thomas Monath on September 17, 2014.
Protecting Humans with Animal Vaccines
Monath briefly discussed prospects for immunizing animal species that are reservoirs for emerging human pathogens, a strategy that is being investigated to prevent several vector-borne diseases, including Rift Valley fever (in livestock), Venezuelan equine encephalitis, and leishmaniasis (in dogs). Some innovative approaches to animal vaccines include oral bait vaccines for Lyme disease, as well as “generic” immunization against ticks, according to Monath. Given Kilpatrick’s indictment of robins as a key reservoir for West Nile virus, he noted, “An intriguing idea now obviously has to be a recombinant worm that robins eat,” he observed.
Monath remarked that the work of Ken Linthicum (see Appendix A7) and others demonstrates that if you can predict Rift Valley fever activity and immunize the hosts that contribute to amplification of the virus, you can prevent human disease and prevent direct animal-to-human transmission, which may provide a model for other vaccine-preventable diseases. He noted that the approval process for animal vaccines is less onerous than for human vaccines, and therefore potentially a more attractive investment for pharmaceutical companies.
Both WNV and CHIKV vaccines present obstacles to industry that could be reduced through push/pull incentives, Monath suggested, as well as direct government funding for advanced development. The accumulation of a government stockpile, which could begin when the vaccine was still under emergency use authorization or not yet licensed, would also boost development. Ultimately, he concluded, “We need a faster, easier, less expensive way to get these kinds of vaccines through the regulatory process, reduce the cost of development, and improve the return on the investment for industry.”
REFERENCES
Akahata, W., Z. Y. Yang, H. Andersen, S. Sun, H. A. Holdaway, W. P. Kong, M. G. Lewis, S. Higgs, M. G. Rossmann, S. Rao, and G. J. Nabel. 2010. A virus-like particle vaccine for epidemic chikungunya virus protects nonhuman primates against infection. Nature Medicine 16(3):334-338. doi: 10.1038/nm.2105.
Almeida, R. 2007. Vector-borne plant diseases: Factors driving the emergence and spread of pathogens. Presentation at the Forum on Microbial Threats workshop entitled “Vector-borne diseases: Understanding the environmental, human health, and ecological connections,” June 19–20, Ft. Collins, CO.
Almeida, R. 2014. The past, present, and future of vector-borne plant diseases. Presentation at IOM workshop on vector-borne diseases, Washington, DC.
Alphey, L. 2014a. Development and evaluation of transgenic insects for use in the control of vector-borne disease. Presentation at IOM workshop on vector-borne disease, Washington, DC.
Alphey, L. 2014b. Genetic control of mosquitoes. Annual Review of Entomology 59:205-224. doi: 10.1146/annurev-ento-011613-162002.
Alphey, L., M. Benedict, R. Bellini, G. G. Clark, D. A. Dame, M. W. Service, and S. L. Dobson. 2010. Sterile-insect methods for control of mosquito-borne diseases: An analysis. Vector Borne Zoonotic Diseases 10(3):295-311. doi: 10.1089/vbz.2009.0014.
Anderson, P. K., A. A. Cunningham, N. G. Patel, F. J. Morales, P. R. Epstein, and P. Daszak. 2004. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends in Ecology and Evolution 19(10):535-544. doi: 10.1016/j.tree.2004.07.021.
Angerami, R. N., A. M. da Silva, E. M. Nascimento, S. Colombo, M. Y. Wada, F. C. dos Santos, D. M. Mancini, R. C. de Oliveira, G. Katz, E. C. Martins, and L. J. da Silva. 2009. Brazilian spotted fever: Two faces of a same disease? A comparative study of clinical aspects between an old and a new endemic area in Brazil. Clinical Microbiology and Infection 15(Suppl 2):207-208. doi: 10.1111/j.1469-0691.2008.02160.x.
Antinori, S., L. Schifanella, and M. Corbellino. 2012. Leishmaniasis: New insights from an old and neglected disease. European Journal of Clinical Microbiology and Infectious Disease 31(2):109-118. doi: 10.1007/s10096-011-1276-0.
Anyamba, A., K. J. Linthicum, J. L. Small, K. M. Collins, C. J. Tucker, E. W. Pak, S. C. Britch, J. R. Eastman, J. E. Pinzon, and K. L. Russell. 2012. Climate teleconnections and recent patterns of human and animal disease outbreaks. PLoS Neglected Tropical Diseases 6(1):e1465. doi: 10.1371/journal.pntd.0001465.
Anyamba, A., J. L. Small, S. C. Britch, C. J. Tucker, E. W. Pak, C. A. Reynolds, J. Crutchfield, and K. J. Linthicum. 2014. Recent weather extremes and impacts on agricultural production and vector-borne disease outbreak patterns. PLoS ONE 9(3):e92538. doi: 10.1371/journal. pone.0092538.
Aregawi, M., M. Lynch, W. Bekele, H. Kebede, D. Jima, H. S. Taffese, M. A. Yenehun, A. Lilay, R. Williams, M. Thomson, F. Nafo-Traore, K. Admasu, T. A. Gebreyesus, and M. Coosemans. 2014. Time series analysis of trends in malaria cases and deaths at hospitals and the effect of antimalarial interventions, 2001-2011, Ethiopia. PLoS ONE 9(11):e106359. doi: 10.1371/journal.pone.0106359.
ARS (Agricultural Research Service, USDA). 2013. Phytoplasma Resource Center. http://plantpathology.ba.ars.usda.gov/phytoplasma.html (accessed July 26, 2013).
Ashley, E. A., M. Dhorda, R. M. Fairhurst, C. Amaratunga, P. Lim, S. Suon, S. Sreng, J. M. Anderson, S. Mao, B. Sam, C. Sopha, C. M. Chuor, C. Nguon, S. Sovannaroth, S. Pukrittayakamee, P. Jittamala, K. Chotivanich, K. Chutasmit, C. Suchatsoonthorn, R. Runcharoen, T. T. Hien, N. T. Thuy-Nhien, N. V. Thanh, N. H. Phu, Y. Htut, K. T. Han, K. H. Aye, O. A. Mokuolu, R. R. Olaosebikan, O. O. Folaranmi, M. Mayxay, M. Khanthavong, B. Hongvanthong, P. N. Newton, M. A. Onyamboko, C. I. Fanello, A. K. Tshefu, N. Mishra, N. Valecha, A. P. Phyo, F. Nosten, P. Yi, R. Tripura, S. Borrmann, M. Bashraheil, J. Peshu, M. A. Faiz, A. Ghose, M. A. Hossain, R. Samad, M. R. Rahman, M. M. Hasan, A. Islam, O. Miotto, R. Amato, B. MacInnis, J. Stalker, D. P. Kwiatkowski, Z. Bozdech, A. Jeeyapant, P. Y. Cheah, T. Sakulthaew, J. Chalk, B. Intharabut, K. Silamut, S. J. Lee, B. Vihokhern, C. Kunasol, M. Imwong, J. Tarning, W. J. Taylor, S. Yeung, C. J. Woodrow, J. A. Flegg, D. Das, J. Smith, M. Venkatesan, C. V. Plowe, K. Stepniewska, P. J. Guerin, A. M. Dondorp, N. P. Day, N. J. White, and Tracking Resistance to Artemisinin Collaboration. 2014. Spread of artemisinin resistance in Plasmodium falciparum malaria. New England Journal of Medicine 371(5):411-423. doi: 10.1056/NEJMoa1314981.
AVMA (American Veterinary Medical Association). 2008. One Health: A new professional imperative. Schaumburg, IL: American Veterinary Medical Association.
Badillo-Vargas, I., D. Rotenberg, B. A. Schneweis, and A. E. Whitfield. 2015. RNA interference tools for Frankliniella occidentalis. Journal of Insect Physiology 76:36-46.
Bean, A. G. D., M. L. Baker, C. R. Stewart, C. Cowled, C. Deffrasnes, L.-F. Wang, and J. W. Lowenthal. 2013. Studying immunity to zoonotic diseases in the natural host—Keeping it real. Nature Reviews Immunology 13:851-861.
Black, W. C., 4th, L. Alphey, and A. A. James. 2011. Why RIDL is not SIT. Trends in Parasitology 27(8):362-370.
Blanc, S., M. Drucker, and M. Uzest. 2014. Localizing viruses in their insect vectors. Annual Review of Phytopathology 52:403-425. doi: 10.1146/annurev-phyto-102313-045920.
Bonning, B. C., N. Pal, S. Liu, Z. Wang, S. Sivakumar, P. M. Dixon, G. F. King, and W. A. Miller. 2014. Toxin delivery by the coat protein of an aphid-vectored plant virus provides plant resistance to aphids. Nature Biotechnology 32(1):102-105.
Brehm, M. A., N. Jouvet, D. L. Greiner, and L. D. Shultz. 2013. Humanized mice for the study of infectious diseases. Current Opinion in Immunology 25(4):428-435. doi: 10.1016/j.coi.2013.05.012.
Carribean 360. 2014. Chikungunya declared an epidemic in the Caribbean. http://www.caribbean360.com/news/chikungunya-declared-an-epidemic-in-the-caribbean (accessed December 12, 2014).
Carrillo, E., J. Moreno, and I. Cruz. 2013. What is responsible for a large and unusual outbreak of leishmaniasis in Madrid? Trends in Parasitology 29(12):579-580. doi: 10.1016/j.pt.2013.10.007.
CDC (Centers for Disease Control and Prevention). 2006. Assessing capacity for surveillance, prevention, and control of West Nile virus infection—United States, 1999 and 2004. Morbidity and Mortality Weekly Report 55(6):150-153.
CDC. 2013. Three sudden cardiac deaths associated with Lyme carditis—United States, November 2012-July 2013. Morbidity and Mortality Weekly Report 62(49):993-996.
CDC. 2014a. Chikungunya nowcast for the Americas. http://www.cdc.gov/chikungunya/modeling (accessed November 24, 2014).
CDC. 2014b. First chikungunya case acquired in the United States reported in Florida. http://www.cdc.gov/media/releases/2014/p0717-chikungunya.html (accessed November 24, 2014).
CDC. 2014c. Geographic distribution: Where has chikungunya virus been found? http://www.cdc.gov/chikungunya/geo/index.html (accessed November 24, 2014).
CDC. 2014d. World Health Day—Vector-borne diseases. http://www.cdc.gov/features/worldhealth day2014 (accessed October 20, 2014).
CDC. 2015. Dengue. http://www.cdc.gov/Dengue (accessed October 5, 2015).
Chang, L. J., K. A. Dowd, F. H. Mendoza, J. G. Saunders, S. Sitar, S. H. Plummer, G. Yamshchikov, U. N. Sarwar, Z. Hu, M. E. Enama, R. T. Bailer, R. A. Koup, R. M. Schwartz, W. Akahata, G. J. Nabel, J. R. Mascola, T. C. Pierson, B. S. Graham, J. E. Ledgerwood, and VRC 311 Study Team. 2014. Safety and tolerability of chikungunya virus-like particle vaccine in healthy adults: A phase 1 dose-escalation trial. Lancet 384(9959):2046-2052. doi: 10.1016/S0140-6736(14)61185-5.
Chretien, J.-P., A. Anyamba, J. L. Small, S. C. Britch, J. L. Sanchez, A. C. Halbach, C. J. Tucker, and K. J. Linthicum. 2015. Global climate anomalies and potential infectious disease risks: 2014-2015. PLoS Currents Outbreaks Jan 26. Edition 1. doi: 10.1371/currents.outbreaks. 95fbc4a8fb4695e049baabfc2fc8289f.
Clarke, C. F., K. K. Bradley, J. H. Wright, and J. Glowicz. 2013. Case report: Emergence of autochthonous cutaneous leishmaniasis in northeastern Texas and southeastern Oklahoma. American Journal of Tropical Medicine and Hygiene 88(1):157-161. doi: 10.4269/ajtmh.2012.11-0717.
Cohen, J. M., D. L. Smith, C. Cotter, A. Ward, G. Yamey, O. J. Sabot, and B. Moonen. 2012. Malaria resurgence: A systematic review and assessment of its causes. Malaria Journal 11:122. doi: 10.1186/1475-2875-11-122.
Cologna, R., P. M. Armstrong, and R. Rico-Hesse. 2005. Selection for virulent dengue viruses occurs in humans and mosquitoes. Journal of Virology 79(2):853-859. doi: 10.1128/JVI.79.2.853-859.2005.
COMTF (California Oak Mortality Task Force). 2013. Sudden oak death overview. http://www.suddenoakdeath.org/about-sudden-oak-death/history-background (accessed August 11, 2015).
Cox, J., J. Mota, S. Sukupolvi-Petty, M. S. Diamond, and R. Rico-Hesse. 2012. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. Journal of Virology 86(14):7637-7649. doi: 10.1128/JVI.00534-12.
Cragun, W. C., B. L. Bartlett, M. W. Ellis, A. Z. Hoover, S. K. Tyring, N. Mendoza, T. J. Vento, W. L. Nicholson, M. E. Eremeeva, J. P. Olano, R. P. Rapini, and C. D. Paddock. 2010. The expanding spectrum of eschar-associated rickettsioses in the United States. Archives of Dermatology 146(6):641-648. doi: 10.1001/archdermatol.2010.48.
Crowder, D. W., E. A. Dykstra, J. M. Brauner, A. Duffy, C. Reed, E. Martin, W. Peterson, Y. Carriere, P. Dutilleul, and J. P. Owen. 2013. West Nile virus prevalence across landscapes is mediated by local effects of agriculture on vector and host communities. PLoS ONE 8(1):e55006. doi: 10.1371/journal.pone.0055006.
Damsteegt, V. D., R. Scorza, A. L. Stone, W. L. Schneider, K. K. Webb, M. A. Demuth, and F. E. Gildow. 2007. Prunus host range of plum pox virus (PPV) in the United States by aphid and graft inoculation. Plant Disease 91:18-23.
Daszak, P., A. A. Cunningham, and A. D. Hyatt. 2000. Emerging infectious diseases of wildlife—Threats to biodiversity and human health. Science 287(5452):443-449.
Dayan, G. H., K. Pugachev, J. Bevilacqua, J. Lang, and T. P. Monath. 2013. Preclinical and clinical development of a YFV 17 D-based chimeric vaccine against West Nile virus. Viruses 5(12):3048-3070. doi: 10.3390/v5123048.
Devine, G. J., E. Z. Perea, G. F. Killeen, J. D. Stancil, S. J. Clark, and A. C. Morrison. 2009. Using adult mosquitoes to transfer insecticides to Aedes aegypti larval habitats. Proceedings of the National Academy of Sciences of the United States of America 106(28):11530-11534.
Dodd, R. Y. 2012. Emerging infections and transfusion safety. In Practical transfusion medicine, 4th ed., edited by M. F. Murphy, D. H. Pamphilon, and N. M. Heddle. Chichester, UK: John Wiley & Sons. Pp. 161-167.
Douglas, K. O., A. M. Kilpatrick, P. N. Levett, and M. C. Lavoie. 2007. A quantitative risk assessment of West Nile virus introduction into Barbados. West Indian Medical Journal 56(5):394-397.
Duggal, N. K., A. Bosco-Lauth, R. A. Bowen, S. S. Wheeler, W. K. Reisen, T. A. Felix, B. R. Mann, H. Romo, D. M. Swetnam, A. D. T. Barrett, and A. C. Brault. 2014. Evidence for co-evolution of West Nile virus and house sparrows in North America. PLoS Neglected Tropical Diseases 8.
Ebi, K. L., E. Lindgren, J. E. Suk, and J. C. Semenza. 2013. Adaptation to the infectious disease impacts of climate change. Climatic Change 118:355-365.
Eisen, L., and C. G. Moore. 2013. Aedes (Stegomyia) aegypti in the continental United States: A vector at the cool margin of its geographic range. Journal of Medical Entomology 50(3):467-478.
European Food Safety Authority. 2014. Schmallenberg virus: State of art. EFSA Journal 12(5).
Fischer, M., J. E. Staples, National Center for Emerging Arboviral Diseases Branch, and CDC Zoonotic Infectious Diseases. 2014. Notes from the field: Chikungunya virus spreads in the Americas−Caribbean and South America, 2013-2014. Morbidity and Mortality Weekly Report 63(22):500-501.
Fletcher, J., and A. Wayadande. 2002. Fastidious vascular-colonizing bacteria. http://www.apsnet.org/edcenter/intropp/PathogenGroups/Pages/Fastidious.aspx (accessed December 12, 2014).
Florida Keys Mosquito Control District. 2014. http://keysmosquito.org/wp-content/uploads/2013/02/All-Keys-Survey-Results-2013.pdf (accessed June 2, 2015).
Freedman, D. O., L. H. Weld, P. E. Kozarsky, T. Fisk, R. Robins, F. von Sonnenburg, J. S. Keystone, P. Pandey, M. S. Cetron, and Network GeoSentinel Surveillance. 2006. Spectrum of disease and relation to place of exposure among ill returned travelers. New England Journal of Medicine 354(2):119-130. doi: 10.1056/NEJMoa051331.
Fu, G., R. S. Lees, D. Nimmo, D. Aw, L. Jin, P. Gray, T. U. Berendonk, H. White-Cooper, S. Scaife, H. Kim Phuc, O. Marinotti, N. Jasinskiene, A.A. James, and L. Alphey. 2010. Female-specific flightless phenotype for mosquito control. Proceedings of the National Academy of Sciences of the United States of America 107:4550-4554.
Funk, S., T. L. Bogich, K. E. Jones, A. M. Kilpatrick, and P. Daszak. 2013. Quantifying trends in disease impact to produce a consistent and reproducible definition of an emerging infectious disease. PLoS ONE 8:e69951.
Gallian, P., X. de Lamballerie, N. Salez, G. Piorkowski, P. Richard, L. Paturel, R. Djoudi, I. Leparc-Goffart, P. Tiberghien, J. Chiaroni, and R. N. Charrel. 2014. Prospective detection of chikungunya virus in blood donors, Caribbean 2014. Blood 123(23):3679-3681. doi: 10.1182/blood-2014-03-564880.
Garcia, G. P, A. E. Flores, I. Fernández-Salas, K. Saavedra-Rodríguez, G. Reyes-Solis, S. Lozano-Fuentes, J. Guillermo Bond, M. Casas-Martínez, J. M. Ramsey, J. García-Rejón, M. Domínguez-Galera, H. Ranson, J. Hemingway, L. Eisen, and W. C. Black 4th. 2009. Recent rapid rise of a permethrin knock down resistance allele in Aedes aegypti in México. PLoS Neglected Tropical Diseases 3(10):e531.
Garcia, G., N. Gonzalez, A. B. Perez, B. Sierra, E. Aguirre, D. Rizo, A. Izquierdo, L. Sanchez, D. Diaz, M. Lezcay, B. Pacheco, K. Hirayama, and M. G. Guzman. 2011. Long-term persistence of clinical symptoms in dengue-infected persons and its association with immunological disorders. International Journal of Infectious Diseases 15(1):e38-e43. doi: 10.1016/j.ijid.2010.09.008.
Garcia, M. N., A. M. Hause, C. M. Walker, J. S. Orange, R. Hasbun, and K. O. Murray. 2014. Evaluation of prolonged fatigue post-West Nile virus infection and association of fatigue with elevated antiviral and proinflammatory cytokines. Viral Immunology 27(7):327-333. doi: 10.1089/vim.2014.0035.
García-Rejón, J., M. A. Loroño-Pino, J. A. Farfan-Ale, L. Flores-Flores, E. Del Pilar Rosado-Paredes, N. Rivero-Cardenas, R. Najera-Vazquez, S. Gomez-Carro, V. Lira-Zumbardo, P. Gonzalez-Martinez, S. Lozano-Fuentes, D. Elizondo-Quiroga, B. J. Beaty, and L. Eisen. 2008. Dengue virus-infected Aedes aegypti in the home environment. American Journal of Tropical Medicine and Hygiene 79(6):940-950.
García-Rejón, J. E., M. A. Loroño-Pino, J. A. Farfán-Ale, L. F. Flores-Flores, M. P. López-Uribe, R. Najera-Vazquez Mdel, G. Nuñez-Ayala, B. J. Beaty, and L. Eisen. 2011. Mosquito infestation and dengue virus infection in Aedes aegypti females in schools in Merida, Mexico. American Journal of Tropical Medicine and Hygiene 84(3):489-496. doi: 10.4269/ajtmh.2011.10-0654.
Gergerich, R. C., and V. V. Dolja. 2006. Introduction to plant viruses, the invisible foe. http://www.apsnet.org/edcenter/intropp/PathogenGroups/Pages/PlantViruses.asp (accessed December 12, 2014).
Gething, P. W., D. L. Smith, A. P. Patil, A. J. Tatem, R. W. Snow, and S. I. Hay. 2010. Climate change and the global malaria recession. Nature 465:342-346.
Gray, S. M., and N. Banerjee. 1999. Mechanisms of arthropod transmission of plant and animal viruses. Microbiol and Molecular Biology Reviews 63(1):128-148.
Gubler, D. J. 1998. Resurgent vector-borne diseases as a global health problem. Emerging Infectious Diseases 4(3):442-450. doi: 10.3201/eid0403.980326.
Gubler, D. J. 2011. Dengue, urbanization and globalization: The unholy trinity of the 21st century. Tropical Medicine and Health 39:3-11. doi:10.2149/tmh.2011-S05.
Hackett. L. W. 1937. Malaria in Europe, an ecological study. London: Oxford University Press.
Hadler, J. L., D. Patel, K. Bradley, J. M. Hughes, C. Blackmore, P. Etkind, L. Kan, J. Getchell, J. Blumenstock, J. Engel, and Control Centers for Disease and Prevention. 2014. National capacity for surveillance, prevention, and control of West Nile virus and other arbovirus infections−United States, 2004 and 2012. Morbidity and Mortality Weekly Report 63(13):281-284.
Hamer, G. L., L. F. Chaves, T. K. Anderson, U. D. Kitron, J. D. Brawn, M. O. Ruiz, S. R. Loss, E. D. Walker, and T. L. Goldberg. 2011. Fine-scale variation in vector host use and force of infection drive localized patterns of West Nile virus transmission. PLoS ONE 6:e23767.
Harris, A. F., A. R. McKemey, D. Nimmo, Z. Curtis, I. Black, S. A. Morgan, M. N. Oviedo, R. Lacroix, N. Naish, N. I. Morrison, A. Collado, J. Stevenson, S. Scaife, T. Dafa’alla, G. Fu, C. Phillips, A. Miles, N. Raduan, N. Kelly, C. Beech, C. A. Donnelly, W. D. Petrie, and L. Alphey. 2012. Successful suppression of a field mosquito population by sustained release of engineered male mosquitoes. Nature Biotechnology 30(9):828-830. doi: 10.1038/nbt.2350.
Harvell, C. D., C. E. Mitchell, J. R. Ward, S. Altizer, A. P. Dobson, R. S. Ostfeld, and M. D. Samuel. 2002. Climate warming and disease risks for terrestrial and marine biota. Science 296(5576):2158-2162.
Hayes, E. B. 2009. Zika virus outside Africa. Emerging Infectious Diseases 15(9):1347-1350. doi: 10.3201/eid1509.090442.
Hogenhout, S. A., E.-D. Ammar, A. E. Whitfield, and M. G. Redinbaugh. 2008. Insect vector interactions with persistently transmitted viruses. Annual Review of Phytopathology 46:327-359. doi: 10.1146/annurev.phyto.022508.092135.
IOM (Insitute of Medicine). 1992. Emerging infections: Microbial threats to health in the United States. Washington, DC: National Academy Press.
IOM. 2008. Vector-borne diseases: Understanding the environment, human health, and ecological connections. Washington, DC: The National Academies Press.
Johns Hopkins Bloomberg School of Public Health. 1991. Fred Soper, DrPH, MPH. http://www.jhsph.edu/about/history/heroes-of-public-health/fred-soper.html (accessed December 5, 2014).
Johnston, S. H., C. A. Glaser, K. Padgett, D. A. Wadford, A. Espinosa, N. Espinosa, M. E. Eremeeva, K. Tait, B. Hobson, S. Shtivelman, C. Hsieh, and S. L. Messenger. 2013. Rickettsia spp. 364D causing a cluster of eschar-associated illness, California. Pediatric Infectious Disease Journal 32(9):1036-1039. doi: 10.1097/INF.0b013e318296b24b.
Jongejan, F., and G. Uilenberg. 2004. The global importance of ticks. Parasitology 129(Suppl):S3-S14.
Kang, S., A. R. Shields, N. Jupatanakul, and G. Dimopoulos. 2014. Suppressing dengue-2 infection by chemical inhibition of Aedes aegypti host factors. PLoS Neglected Tropical Diseases 8(8): e3084.
Katz, L. 2014. Not much happening in the real world, so i’ll update you on mine. America’s Blood Centers. http://www.americasblood.org/blog/not-much-happening-in-the-real-world,-so-i’llupdate-you-on-mine.aspx (accessed November 30, 2014).
Khan, K., I. Bogoch, J. S. Brownstein, J. Miniota, A. Nicolucci, W. Hu, E. O. Nsoesie, M. Cetron, M. I. Creatore, M. German, and A. Wilder-Smith. 2014. Assessing the origin of and potential for international spread of chikungunya virus from the Caribbean. PLoS Currents 6. doi: 10.1371/currents.outbreaks.2134a0a7bf37fd8d388181539fea2da5.
Killeen, G. F. 2014. Characterizing, controlling and eliminating residual malaria transmission. Malaria Journal 13:330.
Kilpatrick, A. M. 2011. Globalization, land use, and the invasion of West Nile virus. Science 334(6054):323-327. doi: 10.1126/science.1201010.
Kilpatrick, A. M. 2014. Globalization, land use, global warming, and the invasion of West Nile virus. Presentation at IOM workshop on vector-borne diseases, Washington, DC.
Kilpatrick, A. M., and W. J. Pape. 2013. Predicting human West Nile virus infections with mosquito surveillance data. American Journal of Epidemiology 178(5):829-835. doi: 10.1093/aje/kwt046.
Kilpatrick, A. M., and S. E. Randolph. 2012. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 380(9857):1946-1955. doi: 10.1016/S0140-6736(12)61151-9.
Kilpatrick, A. M., Y. Gluzberg, J. Burgett, and P. Daszak. 2004. A quantitative risk assessment of the pathways by which West Nile virus could reach Hawaii. Ecohealth 1(2):205-209.
Kilpatrick, A. M., L. D. Kramer, S. R. Campbell, E. O. Alleyne, A. P. Dobson, and P. Daszak. 2005. West Nile virus risk assessment and the bridge vector paradigm. Emerging Infectious Diseases 11(3):425-429. doi: 10.3201/eid1103.040364.
Kilpatrick, A. M., P. Daszak, M. J. Jones, P. P. Marra, and L. D. Kramer. 2006. Host heterogeneity dominates West Nile virus transmission. Proceedings of the Royal Society B: Biological Sciences 273:2327-2333.
Kilpatrick, A. M., M. A. Meola, R. M. Moudy, and L. D. Kramer. 2008. Temperature, viral genetics, and the transmission of West Nile virus by Culex pipiens mosquitoes. PLoS Pathogens 4(6):e1000092. doi: 10.1371/journal.ppat.1000092.
Knox, J., R. U. Cowan, J. S. Doyle, M. K. Ligtermoet, J. S. Archer, J. N. Burrow, S. Y. Tong, B. J. Currie, J. S. Mackenzie, D. W. Smith, M. Catton, R. J. Moran, C. A. Aboltins, and J. S. Richards. 2012. Murray Valley encephalitis: A review of clinical features, diagnosis and treatment. Medical Journal of Australia 196(5):322-326.
Kupferschmidt, K. 2013. A lethal dose of RNA. Science 341(6147):732-733. doi: 10.1126/science.341.6147.732.
Lindblade, K. A., L. Steinhardt, A. Samuels, S. P. Kachur, and L. Slutsker. 2013. The silent threat: Asymptomatic parasitemia and malaria transmission. Expert Review of Anti-Infective Therapy 11(6):623-639. doi: 10.1586/eri.13.45.
Lindgren, E., Y. Andersson, J. E. Suk, B. Sudre, and J. C. Semenza. 2012. Public health. Monitoring EU emerging infectious disease risk due to climate change. Science 336(6080):418-419. doi: 10.1126/science.1215735.
Lindsey, N. P., J. A. Lehman, J. E. Staples, M. Fischer, National Center for Emerging Division of Vector-Borne Diseases, and CDC Zoonotic Infectious Diseases. 2014. West Nile virus and other arboviral diseases—United States, 2013. Morbidity and Mortality Weekly Report 63(24):521-526.
Linthicum, K. J., A. Anyamba, B. Killenbeck, W. J. Lee, H. C. Lee, T. A. Klein, H. C. Kim, J. A. Pavlin, S. C. Britch, J. Small, C. J. Tucker, and J. C. Gaydos. 2014. Association of temperature and historical dynamics of malaria in the Republic of Korea, including reemergence in 1993. Military Medicine 179(7):806-814. doi: 10.7205/MILMED-D-13-00545.
Maxmen, A. 2012. The hidden threat of West Nile virus. Nature 489(7416):349-350. doi: 10.1038/489349a.
McMullan, L. K., S. M. Folk, A. J. Kelly, A. MacNeil, C. S. Goldsmith, M. G. Metcalfe, B. C. Batten, C. G. Albarino, S. R. Zaki, P. E. Rollin, W. L. Nicholson, and S. T. Nichol. 2012. A new phlebovirus associated with severe febrile illness in Missouri. New England Journal of Medicine 367(9):834-841. doi: 10.1056/NEJMoa1203378.
Mielke-Ehret, N., and H. P. Muhlbach. 2012. Emaravirus: A novel genus of multipartite, negative strand RNA plant viruses. Viruses 4(9):1515-1536. doi: 10.3390/v4091515.
Miller, W. A., and L. Rasochova. 1997. Barley yellow dwarf viruses. Annual Review of Phytopathology 35:167-190.
Minjauw, B., and A. McLeod. 2003. Tick-borne diseases and poverty. The impact of ticks and tickborne diseases on the livelihood of small-scale and marginal livestock owners in India and eastern and southern Africa. University of Edinburgh, UK: DFID Animal Health Programme, Centre for Tropical Veterinary Medicine.
Monath, T. P. 2014. Dengue, Japanese encephalitis, West Nile, chikungunya, and yellow fever: Challenges for the development and use of vaccines. Presentation at IOM workshop on vector-borne diseases, Washington, DC.
Montero-Astúa, M., D. Rotenberg, A. Leach-Kieffaber, B. A. Schneweis, S. Park, J. K. Park, T. L. German, and A. E. Whitfield. 2014. Disruption of vector transmission by a plant-expressed viral glycoprotein. Molecular Plant-Microbe Interactions Journal 27(3):296-304. doi: 10.1094/MPMI-09-13-0287-FI.
Montgomery, S. P., M. C. Starr, P. T. Cantey, M. S. Edwards, and S. K. Meymandi. 2014. Neglected parasitic infections in the United States: Chagas disease. American Journal of Tropical Medicine and Hygiene 90(5):814-818. doi: 10.4269/ajtmh.13-0726.
Mordecai, E. A., K. P. Paaijmans, L. R. Johnson, C. Balzer, T. Ben-Horin, E. de Moor, A. McNally, S. Pawar, S. J. Ryan, T. C. Smith, and K. D. Lafferty. 2013. Optimal temperature for malaria transmission is dramatically lower than previously predicted. Ecology Letters 16(1):22-30. doi: 10.1111/ele.12015.
Moreira, L. A., I. Iturbe-Ormaetxe, J. A. Jeffery, G. Lu, A. T. Pyke, L. M. Hedges, B. C. Rocha, S. Hall-Mendelin, A. Day, M. Riegler, L. E. Hugo, K. N. Johnson, B. H. Kay, E. A. McGraw, A. F. van den Hurk, P. A. Ryan, and S. L. O’Neill. 2009. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, chikungunya, and plasmodium. Cell 139(7):1268-1278. doi: 10.1016/j.cell.2009.11.042.
Morens, D. M., and A. S. Fauci. 2014. Chikungunya at the door—déjà vu all over again? New England Journal of Medicine 371(10):885-887. doi: 10.1056/NEJMp1408509.
Moritz, E. D., C. S. Winton, S. T. Johnson, D. E. Krysztof, R. L. Townsend, G. A. Foster, P. Devine, P. Molloy, E. Brissette, V. P. Berardi, and S. L. Stramer. 2014. Investigational screening for Babesia microti in a large repository of blood donor samples from nonendemic and endemic areas of the United States. Transfusion 54(9):2226-2236. doi: 10.1111/trf.12693.
Murdock, C. C., S. Blanford, G. L. Hughes, J. L. Rasgon, and M. B. Thomas. 2014. Temperature alters Plasmodium blocking by Wolbachia. Scientific Reports 4:3932. doi: 10.1038/srep03932.
Murray, K. O., M. N. Garcia, M. H. Rahbar, D. Martinez, S. A. Khuwaja, R. R. Arafat, and S. Rossmann. 2014. Survival analysis, long-term outcomes, and percentage of recovery up to 8 years post-infection among the Houston West Nile virus cohort. PLoS ONE 9 (7):e102953. doi: 10.1371/journal.pone.0102953.
Musso, D., V. Richard, J. Broult, and V. M. Cao-Lormeau. 2014. Inactivation of dengue virus in plasma with amotosalen and ultraviolet A illumination. Transfusion 54(11):2924-2930. doi: 10.1111/trf.12713.
Nasci, R. S. 2014. Movement of chikungunya virus into the Western Hemisphere. Emerging Infectious Diseases 8:1394-1395. doi: 10.3201/eid2008.140333.
The National Oceanic and Atmospheric Administration’s (NOAA) Climate Prediction Center (CPC) 2014, El Niño conditions advisory. http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/enso_advisory/index.shtml (accessed March 25, 2016).
NOAA (National Oceanic and Atmospheric Administration). 2015. Global temperature record. http://ncdc.noaa.gov/cag/timeseries/global/globe/land_ocean/ytd/12/1880-2014 (accessed March 25, 2016).
NRC (National Research Council). 2004. California agricultural research priorities: Pierce’s disease. Washington, DC: The National Academies Press.
Ogden, N. H., L. St-Onge, I. K. Barker, S. Brazeau, M. Bigras-Poulin, D. F. Charron, C. M. Francis, A. Heagy, L. Robbin Lindsay, A. Maarouf, P. Michel, F. Milord, C. J. O’Callaghan, L. Trudel, and R. A. Thompson. 2008. Risk maps for range expansion of the Lyme disease vector, Ixodes scapularis, in Canada now and with climate change. International Journal of Health Geographics 7:24.
Paaijmans, K. P., R. L. Heinig, R. A. Seliga, J. I. Blanford, S. Blanford, C. C. Murdock, and M. B. Thomas. 2013. Temperature variation makes ectotherms more sensitive to climate change. Global Change Biology 19(8):2373-2380. doi: 10.1111/gcb.12240.
Paddock, C. D., and M. J. Yabsley. 2007. Ecological havoc, the rise of white-tailed deer, and the emergence of Amblyomma americanum-associated zoonoses in the United States. Current Topics in Microbiology and Immunology 315:289-324.
Paddock, C. D., R. W. Finley, C. S. Wright, H. N. Robinson, B. J. Schrodt, C. C. Lane, O. Ekenna, M. A. Blass, C. L. Tamminga, C. A. Ohl, S. L. McLellan, J. Goddard, R. C. Holman, J. J. Openshaw, J. W. Sumner, S. R. Zaki, and M. E. Eremeeva. 2008. Rickettsia parkeri rickettsiosis and its clinical distinction from Rocky Mountain spotted fever. Clinical Infectious Diseases 47(9):1188-1196. doi: 10.1086/592254.
PAHO (Pan American Health Organization). 2014a. Dengue Laboratory Network of the Americas (RELDA). http://www.paho.org/hq/index.php?option=com_content&view=article&id=4497&Itemid=39306&lang=en (accessed December 13, 2014).
PAHO. 2014b. Number of reported cases of chikungunya fever in the Americas, by country or territory, epidemiological week 47 (November 21, 2014). Washington, DC: Pan American Health Organization.
Paz, S., D. Malkinson, M. S. Green, G. Tsioni, A. Papa, K. Danis, A. Sirbu, C. Ceianu, K. Katalin, E. Ferenczi, H. Zeller, and J. C. Semenza. 2013. Permissive summer temperatures of the 2010 European West Nile fever upsurge. PLoS ONE 8(2):e56398. doi: 10.1371/journal.pone.0056398.
Petersen, L.R. 2014. Emerging vector-borne diseases in the United States: What’s next and are we prepared? Presentation at the IOM workshop on vector-borne diseases, Washington, DC.
Phuc, H. K., M. H. Andreasen, R. S. Burton, C. Vass, M. J. Epton, G. Pape, G, Fu, K. C. Condon, S. Scaife, C. A. Donnelly, P. G. Coleman, H. White-Cooper, and L. Alphey. 2007. Late-acting dominant lethal genetic systems and mosquito control. BMC Biology 5:11.
Powell, J. R., and W. J. Tabachnick. 2013. History of domestication and spread of Aedes aegypti—a review. Memórias do Instituto Oswaldo Cruz 108(Suppl 1):11-17. doi: 10.1590/0074-0276130395.
Purcell, A. H. 1982. Insect vector relationships with procaryotic plant pathogens. Annual Review of Phytopathology 20(1):397-417.
Ramos, M. M., H. Mohammed, E. Zielinski-Gutierrez, M. H. Hayden, J. L. Lopez, M. Fournier, A. R. Trujillo, R. Burton, J. M. Brunkard, L. Anaya-Lopez, A. A. Banicki, P. K. Morales, B. Smith, J. L. Munoz, S. H. Waterman, and Group Dengue Serosurvey Working. 2008. Epidemic dengue and dengue hemorrhagic fever at the Texas-Mexico border: Results of a household-based seroepidemiologic survey, December 2005. American Journal of Tropical Medicine and Hygiene 78(3):364-369.
Rasongles, P., M. F. Angelini-Tibert, P. Simon, C. Currie, H. Isola, D. Kientz, M. Slaedts, M. Jacquet, D. Sundin, L. Lin, L. Corash, and J. P. Cazenave. 2009. Transfusion of platelet components prepared with photochemical pathogen inactivation treatment during a chikungunya virus epidemic in Ile de La Reunion. Transfusion 49(6):1083-1091. doi: 10.1111/j.1537-2995.2009.02111.x.
Rassi, A., Jr., A. Rassi, and J. A. Marin-Neto. 2010. Chagas disease. Lancet 375(9723):1388-1402. doi: 10.1016/S0140-6736(10)60061-X.
Reisen, W. K. 2010. Landscape epidemiology of vector-borne diseases. Annual Review of Entomology 55:461-483. doi: 10.1146/annurev-ento-112408-085419.
Reiter, P. 2007. Oviposition, dispersal, and survival in Aedes aegypti: Implications for the efficacy of control strategies. Vector Borne Zoonotic Diseases 7(2):261-273. doi: 10.1089/vbz.2006.0630.
Reiter, P. 2014. Surveillance and control of urban dengue vectors. In Dengue and dengue hemorrhagic fever, 2nd ed., edited by D. J. Gubler, E. E. Ooi, S. Vasudevan, and J. Farrar. London, United Kingdom: CAB International. Pp. 481-518.
Reynolds, R. W., N. A. Rayner, T. M. Smith, D. C. Stokes, and W. Wanqiu. 2002. An improved in situ and satellite SST analysis for climate. Journal of Climate 15(13):1609-1625.
Rico-Hesse, R. 2009. Dengue virus markers of virulence and pathogenicity. Future Virology 4(6):581. doi: 10.2217/fvl.09.51.
Rogers, D. J., and S. E. Randolph. 2000. The global spread of malaria in a future, warmer world. Science 289:1763-1766.
Sabchareon, A., C. Sirivichayakul, K. Limkittikul, P. Chanthavanich, S. Suvannadabba, V. Jiwariyavej, W. Dulyachai, K. Pengsaa, H. S. Margolis, and G. W. Letson. 2012. Dengue infection in children in Ratchaburi, Thailand: A cohort study. I. Epidemiology of symptomatic acute dengue infection in children, 2006-2009. PLoS Neglected Tropical Diseases 6(7):e1732. doi: 10.1371/journal.pntd.0001732.
Schilte, C., F. Staikowsky, T. Couderc, Y. Madec, F. Carpentier, S. Kassab, M. L. Albert, M. Lecuit, and A. Michault. 2013. Chikungunya virus-associated long-term arthralgia: A 36-month prospective longitudinal study. PLoS Neglected Tropical Diseases 7(3):e2137. doi: 10.1371/journal. pntd.0002137.
Semenza, J. C., and D. Domanovic. 2013. Blood supply under threat. Nature Climate Change 3(5):432-435.
Semenza, J. C., J. E. Suk, V. Estevez, K. L. Ebi, and E. Lindgren. 2012. Mapping climate change vulnerabilities to infectious diseases in Europe. Environmental Health Perspectives 120(3):385-392. doi: 10.1289/ehp.1103805.
Semenza, J. C., B. Sudre, T. Oni, J. E. Suk, and J. Giesecke. 2013. Linking environmental drivers to infectious diseases: The European environment and epidemiology network. PLoS Neglected Tropical Diseases 7(7):e2323. doi: 10.1371/journal.pntd.0002323.
Semenza, J. C., B. Sudre, J. Miniota, M. Rossi, W. Hu, D. Kossowsky, J. E. Suk, W. Van Bortel, and K. Khan. 2014. International dispersal of dengue through air travel: Importation risk for Europe. PLoS Neglected Tropical Diseases 8(12):e3278. doi: 10.1371/journal.pntd.0003278.
Siraj, A. S., M. Santos-Vega, M. J. Bouma, D. Yadeta, D. Ruiz Carrascal, and M. Pascual. 2014. Altitudinal changes in malaria incidence in highlands of Ethiopia and Colombia. Science 343(6175):1154-1158.
Smith, D. L., J. M. Cohen, C. Chiyaka, G. Johnston, P. W. Gething, R. Gosling, C. O. Buckee, R. Laxminarayan, S. I. Hay, and A. J. Tatem. 2013. A sticky situation: The unexpected stability of malaria elimination. Philosophical Transactions of the Royal Society of London B: Biological Sciences 368(1623):20120145. doi: 10.1098/rstb.2012.0145.
Stramer, S. L., J. M. Linnen, J. M. Carrick, G. A. Foster, D. E. Krysztof, S. Zou, R. Y. Dodd, L. M. Tirado-Marrero, E. Hunsperger, G. A. Santiago, J. L. Munoz-Jordan, and K. M. Tomashek. 2012. Dengue viremia in blood donors identified by RNA and detection of dengue transfusion transmission during the 2007 dengue outbreak in Puerto Rico. Transfusion 52(8):1657-1666. doi: 10.1111/j.1537-2995.2012.03566.x.
Streit, J. A., M. Yang, J. E. Cavanaugh, and P. M. Polgreen. 2011. Upward trend in dengue incidence among hospitalized patients, United States. Emerging Infectious Diseases 17(5):914-916. doi: 10.3201/eid1705.101023.
Sudre, B., M. Rossi, W. Van Bortel, K. Danis, A. Baka, N. Vakalis, and J. C. Semenza. 2013. Mapping environmental suitability for malaria transmission, Greece. Emerging Infectious Diseases 19(5):784-786. doi: 10.3201/eid1905.120811.
Suk, J. E., and J. C. Semenza. 2011. Future infectious disease threats to Europe. America Journal of Public Health 101(11):2068-2079. doi: 10.2105/AJPH.2011.300181.
Thiberville, S. D., N. Moyen, L. Dupuis-Maguiraga, A. Nougairede, E. A. Gould, P. Roques, and X. de Lamballerie. 2013. Chikungunya fever: Epidemiology, clinical syndrome, pathogenesis and therapy. Antiviral Research 99(3):345-370. doi: 10.1016/j.antiviral.2013.06.009.
Tran, A., B. Sudre, S. Paz, M. Rossi, A. Desbrosse, V. Chevalier, and J. C. Semenza. 2014. Environmental predictors of West Nile fever risk in Europe. International Journal of Health Geographics 13:26. doi: 10.1186/1476-072X-13-26.
Trape, J. F., A. Tall, C. Sokhna, A. B. Ly, N. Diagne, O. Ndiath, C. Mazenot, V. Richard, A. Badiane, F. Dieye-Ba, J. Faye, G. Ndiaye, F. Diene Sarr, C. Roucher, C. Bouganali, H. Bassene, A. Toure-Balde, C. Roussilhon, R. Perraut, A. Spiegel, J. L. Sarthou, L. P. da Silva, O. Mercereau-Puijalon, P. Druilhe, and C. Rogier. 2014. The rise and fall of malaria in a West African rural community, Dielmo, Senegal, from 1990 to 2012: A 22-year longitudinal study. Lancet Infectious Diseases 14(6):476-488. doi: 10.1016/S1473-3099(14)70712-1.
University of California Agriculture and Natural Resources. 2013. Asian citrus psyllid and huanglongbing disease. http://www.ipm.ucdavis.edu/PMG/PESTNOTES/pn74155.html (accessed December 12, 2014).
Vega-Rua, A., K. Zouache, R. Girod, A. B. Failloux, and R. Lourenco-de-Oliveira. 2014. High level of vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor in the spread of chikungunya virus. Journal of Virology 88(11):6294-6306. doi: 10.1128/JVI.00370-14.
von Seidlein, L., K. Ikonomidis, R. Bruun, M. Jawara, M. Pinder, B. G. Knols, and J. B. Knudsen. 2012. Airflow attenuation and bed net utilization: Observations from Africa and Asia. Malaria Journal 11:200. doi: 10.1186/1475-2875-11-200.
Vontas, J., S. Moore, I. Kleinschmidt, H. Ranson, S. Lindsay, C. Lengeler, N. Hamon, T. McLean, and J. Hemingway. 2014. Framework for rapid assessment and adoption of new vector control tools. Trends in Parasitology 30(4):191-204. doi: 10.1016/j.pt.2014.02.005.
Weaver, S. C., and W. K. Reisen. 2010. Present and future arboviral threats. Antiviral Research 85(2):328-345. doi: 10.1016/j.antiviral.2009.10.008.
Weaver, S. C., J. E. Osorio, J. A. Livengood, R. Chen, and D. T. Stinchcomb. 2012. Chikungunya virus and prospects for a vaccine. Expert Review of Vaccines 11(9):1087-1101. doi: 10.1586/erv.12.84.
Weintraub, P. G., and L. Beanland. 2006. Insect vectors of phytoplasmas. Annual Review of Entomology 51:91-111. doi: 10.1146/annurev.ento.51.110104.151039.
White, N. J., S. Pukrittayakamee, T. T. Hien, M. A. Faiz, O. A. Mokuolu, and A. M. Dondorp. 2014. Malaria. Lancet 383(9918):723-35. doi: 10.1016/S0140-6736(13)60024-0.
Whitfield, A. E., and D. Rotenberg. 2015. Disruption of insect transmission of plant viruses. Current Opinion in Insect Science 8:79-87.
Whitfield, A. E., D. Rotenberg, and T. L. German. 2014. Plant pest destruction goes viral. Nature Biotechnology 32(1):65-66. doi: 10.1038/nbt.2787.
WHO (World Health Organization). 2014a. A global brief on vector-borne diseases. Geneva, Switzerland: World Health Organization.
WHO. 2014b. Chikungunya Fact Sheet No. 327. http://www.who.int/mediacentre/factsheets/fs327/en/ (accessed December 29, 2014).
Wilcox, B. A., and D. J. Gubler. 2005. Disease ecology and the global emergence of zoonotic pathogens. Environmental Health and Preventive Medicine 10(5)263-272.
Wiser, M. F. 2013. Kinetoplastids. http://www.tulane.edu/~wiser/protozoology/notes/kinet.html (accessed November 25, 2014).
Yao, J., D. Rotenberg, A. Afsharifar, K. Barandoc-Alviar, and A. E. Whitfield. 2013. Development of RNAi methods for Peregrinus maidis, the corn planthopper. PLoS ONE 8(8):e70243. doi: 10.1371/journal.pone.0070243.
Zhou, J., and I. E. Tzanetakis. 2013. Epidemiology of soybean vein necrosis-associated virus. Phytopathology 103(9):966-971. doi: 10.1094/PHYTO-12-12-0322-R.


























































































