
Engineered Proteins for Visualizing and Treating Cancer
JENNIFER R. COCHRAN
Stanford University
Cancer is complex and its diagnosis and treatment can more effectively be tackled by teams of scientists, engineers, and clinicians whose expertise spans bench-to-bedside approaches.
An emerging core philosophy applies understanding of molecular mechanisms underlying disease pathophysiology as design criteria toward the development of safer and more efficacious tumor targeting agents (Kariolis et al. 2013). Armed with this knowledge, academic and industrial researchers are using a variety of approaches to create tailor-made proteins for application in cancer imaging and therapy. These efforts leverage enabling tools and technologies, including methods for (1) protein design and engineering, (2) biochemical and biophysical analyses, and (3) preclinical evaluation in animal models.
Important deliverables of this work include insight into ligand-mediated cell surface receptor interactions that drive disease, and the development of new protein-based drugs and imaging agents for translation to the clinic.
BACKGROUND
As the field of protein engineering evolved during the 1980s, modified proteins soon joined recombinant versions of natural proteins as a major class of new therapeutics. The ability to customize the biochemical and biophysical properties of proteins to augment their clinical potential has presented many exciting new opportunities for the pharmaceutical industry.
The market value of such biopharmaceuticals is currently more than $140 billion, exceeding the GDP of three-quarters of the economies in the World Bank rankings (Walsh 2014). Monoclonal antibodies used to treat cancer, rheumatoid
arthritis, and cardiovascular and other diseases account for a large share of these efforts (Drewe and Powell 2002). In 2014 the US and European markets included close to 50 monoclonal antibody drugs, a $75 billion market (Ecker et al. 2015). In 2015, the top three revenue-generating cancer drugs were monoclonal antibodies: rituximab (Rituxan®), bevacizumab (Avastin®), and trastuzumab (Herceptin®), all produced by Genentech/Roche. The size of this market underscores both the clinical and economic importance of protein therapeutics in translational medicine.
CURRENT CHALLENGES
Challenges for cancer therapeutics include the need for more selective localization to tumors versus healthy tissue, and improved tissue penetration and delivery to brain tumors, which are protected by the restrictive blood-brain barrier. Other therapeutic challenges are tumor heterogeneity that makes cancers difficult to treat, acquired drug resistance that cannot be overcome because of dose limiting drug toxicity, and lack of effective drugs to treat cancer once it has spread.
Limitations of monoclonal antibodies in addressing these and other challenges have motivated the development of alternative tumor targeting proteins with different molecular sizes and biophysical attributes, conferring altered pharmacological properties (Weidle et al. 2013). In the following sections I describe some examples of engineered protein therapeutics developed by our research team that have opportunities to affect cancer in new and impactful ways.
AN ULTRA-HIGH AFFINITY ENGINEERED PROTEIN THERAPEUTIC FOR TREATING METASTATIC DISEASE
Despite advances over the past few decades in the development of targeted therapeutics, there is a lack of effective drugs to treat cancers once they have spread (called metastasis), and 90 percent of patients succumb to metastatic disease. We teamed up with cancer biologist Amato Giaccia (Stanford Radiation Oncology) to address this challenge.
In a number of human cancers, aberrant signaling through the Axl receptor tyrosine kinase has been demonstrated to drive metastasis (Li et al. 2009), confer therapeutic resistance (Hong et al. 2013), and promote disease progression (Vajkoczy et al. 2006). Additionally, Axl overexpression has been observed in multiple solid and hematological malignancies (Linger et al. 2008), with expression levels often correlating with disease stage and poor clinical prognosis (Gustafsson et al. 2009; Hong et al. 2013; Rankin et al. 2010). Ambiguity surrounding the fundamental characteristics of Axl’s interaction with its ligand, growth arrest-specific 6 (Gas6), including its affinity and the mechanism of receptor activation, have hindered the development of effective Axl antagonists.
We used rational and combinatorial approaches to engineer an Axl “decoy receptor” that binds to the Gas6 ligand with ultra-high affinity and inhibits its
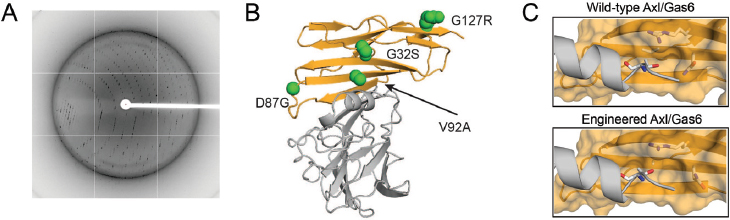
function (Kariolis et al. 2014). Upon fusion to an antibody fragment crystallizable (Fc) domain, the engineered Axl decoy receptor binds Gas6 with an affinity of ~400 femtomolar, placing it among the tightest protein-protein interactions found in nature. Crystallographic analysis of the ligand/receptor interaction, carried out in collaboration with Irimpan Mathews (SLAC National Accelerator Laboratory), showed that mutations in Axl induced structural alterations that resulted in increased Gas6/Axl binding (Figure 1).
The engineered Axl decoy receptor effectively sequestered Gas6, allowing complete abrogation of Axl signaling. Moreover, Gas6 binding affinity was critical and correlative with the ability of decoy receptors to effectively inhibit metastasis and disease progression. The engineered Axl decoy receptor inhibited up to 90 percent of metastatic nodules in two murine models of ovarian cancer compared to wild-type Axl (~50 percent inhibition), with virtually no toxic side effects (Kariolis et al. 2014).
INSPIRATION FROM NATURE TO DEVELOP A NOVEL CLASS OF TUMOR TARGETING AGENTS
A major obstacle to the development of therapeutics that target the brain is the presence of the blood-brain barrier, which prevents foreign particles and
molecules from entering the central nervous system. We recently demonstrated the promise of using engineered peptides, known as knottins, to target brain tumors for applications such as image-guided resection and targeted drug delivery (Ackerman et al. 2014a; Kintzing and Cochran 2016).
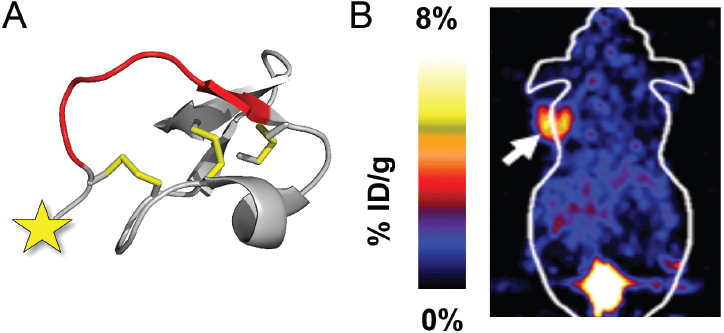
Knottins are unique peptides (30–50 amino acids) containing a disulfide-bonded core that confers outstanding proteolytic resistance and thermal stability (Kolmar 2009). They are found in a wide variety of plants, animals, insects, and fungi, and carry out diverse functions such as ion channel inhibition, enzyme inactivation, and antimicrobial activity (Zhu et al. 2003).
We used molecular engineering approaches to redirect a knottin found in squash seeds that normally functions as an enzyme inhibitor, to create an engineered knottin that binds tumor-associated receptors with high affinity (Kimura et al. 2009a). In collaboration with Zheng Cheng and Sanjiv Sam Gambhir (Stanford Radiology) we established these engineered peptides as a new class of molecular imaging agents for cancer (Kimura et al. 2009b) (Figure 2). We then showed that intravenous injection of an engineered knottin, conjugated to a near-infrared fluorescent dye molecule, targeted and illuminated intracranial brain tumors in animal models of medulloblastoma (collaborations with Matthew Scott, Stanford Developmental Biology, and Samuel Cheshier and Gerald Grant, Stanford Neurosurgery) (Ackerman et al. 2014b; Moore et al. 2013).
Disulfide-rich peptides, including knottins, have generated great interest as potential drug candidates as they offer the pharmacological benefits of small molecule drugs along with the target-binding affinity and specificity of protein biologics. We postulated that if we could use an engineered knottin peptide to visualize tumors, then we could also use it as a vehicle to deliver drugs to tumors, with a goal of minimizing toxic side effects of systemic chemotherapy.
In one study, carried out in collaboration with the Stanford ChEM-H Medicinal Chemistry Knowledge Center, the engineered knottin peptide was conjugated to the nucleoside analogue gemcitabine, using a variety of linker strategies, and an optimal candidate was shown to inhibit proliferation of breast, ovarian, pancreatic, and brain tumor cells in vitro (Cox et al. 2016). Notably, this peptide-drug conjugate was shown to kill cells via receptor-mediated internalization, and thus exhibited increased potency against pancreatic cells that acquired some resistance to treatment with gemcitabine alone.
In a second study, carried out in collaboration with Sutro Biopharma, Inc., the engineered peptide was fused to an antibody Fc domain and conjugated to the tubulin inhibitor monomethyl-auristatin-F. This knottin-Fc-drug conjugate was capable of inducing regression and prolonged survival in a flank glioblastoma model (Currier et al. 2016), highlighting promise for further clinical development.
CONCLUSIONS
Research and development efforts over the past few decades have culminated in a growing number of FDA-approved protein therapeutics that enable targeted treatment of cancer. In parallel, continued efforts to develop safer and more effective cancer therapeutics are being fueled by expanding knowledge of mechanisms underlying disease pathophysiology and the ability to customize proteins using a variety of engineering methods.
The case studies presented above provide examples of how our research team is using protein design and engineering to generate next-generation cancer therapeutics. Protein engineers are also using these powerful technologies to create molecular toolkits for answering a wide range of research questions in basic science, biotechnology, and biomedicine.
REFERENCES
Ackerman SE, Currier NV, Bergen JM, Cochran JR. 2014a. Cystine-knot peptides: Emerging tools for cancer imaging and therapy. Expert Review of Proteomics 11:561–572.
Ackerman SE, Wilson CM, Kahn SA, Kintzing JR, Jindal DA, Cheshier SH, Grant GA, Cochran JR. 2014b. A bioengineered peptide that localizes to and illuminates medulloblastoma: A new tool with potential for fluorescence-guided surgical resection. Cureus 6(9):e207.
Cox N, Kintzing JR, Smith M, Grant GA, Cochran JR. 2016. Integrin-targeting knottin peptide-drug conjugates are potent inhibitors of tumor cell proliferation. Angewandte Chemie International Edition 55(34):9894–9897.
Currier NV, Ackerman SE, Kintzing JR, Chen R, Filsinger Interrante M, Steiner A, Sato AK, Cochran JR. 2016. Targeted drug delivery with an integrin-binding knottin-Fc-MMAF conjugate produced by cell-free protein synthesis. Molecular Cancer Therapeutics 15(6):1291–1300.
Drewe E, Powell RJ. 2002. Clinically useful monoclonal antibodies in treatment. Journal of Clinical Pathology 55(2):81–85.
Ecker DM, Jones SD, Levine HL. 2015. The therapeutic monoclonal antibody market. MAbs 7(1):9–14.
Gustafsson A, Martuszewska D, Johansson M, Ekman C, Hafizi S, Ljungberg B, Dahlback B. 2009. Differential expression of Axl and Gas6 in renal cell carcinoma reflecting tumor advancement and survival. Clinical Cancer Research 15:4742–4749.
Hong J, Peng D, Chen Z, Sehdev V, Belkhiri A. 2013. Abl regulation by Axl promotes cisplatin resistance in esophageal cancer. Cancer Research 73:331–340.
Kariolis MS, Kapur S, Cochran JR. 2013. Beyond antibodies: Using biological principles to guide the development of next-generation protein therapeutics. Current Opinion in Biotechnology 24:1072–1077.
Kariolis MS, Miao YR, Jones DS 2nd, Kapur S, Mathews II, Giaccia AJ, Cochran JR. 2014. An engineered Axl “decoy receptor” effectively silences the Gas6-Axl signaling axis. Nature Chemical Biology 10:977–983.
Kimura RH, Levin AM, Cochran FV, Cochran JR. 2009a. Engineered cystine knot peptides that bind alphavbeta3, alphavbeta5, and alpha5beta1 integrins with low-nanomolar affinity. Proteins 77(2):359–369.
Kimura RH, Cheng Z, Gambhir SS, Cochran JR. 2009b. Engineered knottin peptides: A new class of agents for imaging integrin expression in living subjects. Cancer Research 69:2435–2442.
Kintzing JR, Cochran JR. 2016. Engineered knottin peptides as diagnostics, therapeutics, and drug delivery vehicles. Current Opinion in Chemical Biology 34:143–150.
Kolmar H. 2009. Biological diversity and therapeutic potential of natural and engineered cystine knot miniproteins. Current Opinion in Pharmacology 9(5):608–614.
Li Y, Ye X, Tan C, Hongo JA, Zha J, Liu J, Kallop D, Ludlam MJC, Pei L. 2009. Axl as a potential therapeutic target in cancer: Role of Axl in tumor growth, metastasis and angiogenesis. Oncogene 28:3442–3455.
Linger RM, Keating AK, Earp HS, Graham DK. 2008. Tam receptor tyrosine kinases: Biologic functions, signaling, and potential therapeutic targeting in human cancer. Advances in Cancer Research 100:35–83.
Moore SJ, Hayden Gephart MG, Bergen JM, Su YS, Rayburn H, Scott MP, Cochran JR. 2013. Engineered knottin peptide enables noninvasive optical imaging of intracranial medulloblastoma. Proceedings of the National Academy of Sciences 110(36):14598–14603.
Rankin EB, Fuh KC, Taylor TE, Krieg AJ, Musser M, Yuan J, Wei K, Kuo CJ, Longacre TA, Giaccia AJ. 2010. Axl is an essential factor and therapeutic target for metastatic ovarian cancer. Cancer Research 70(19):7570–7579.
Vajkoczy P, Knyazev P, Kunkel A, Capelle HH, Behrndt S, von Tengg-Kobligk H, Kiessling F, Eichelsbacher U, Essig M, Read TA, Erber R, Ullrich A. 2006. Dominant-negative inhibition of the Axl receptor tyrosine kinase suppresses brain tumor cell growth and invasion and prolongs survival. Proceedings of the National Academy of Sciences 103:5799–5804.
Walsh G. 2014. Biopharmaceutical benchmarks 2014. Nature Biotechnology 32:992–1000.
Weidle UH, Auer J, Brinkmann U, Georges G, Tiefenthaler G. 2013. The emerging role of new protein scaffold-based agents for treatment of cancer. Cancer Genomics and Proteomics 13:155–168.
Zhu S, Darbon H, Dyason K, Verdonck F, Tytgat J. 2003. Evolutionary origin of inhibitor cystine knot peptides. FASEB Journal 17:1765–1767.






