
Implications of Vegetation Change
Expansive changes in tundra and boreal vegetation can have significant implications for physical and ecological processes, as well as impacts on communities. Vegetation change that alters land surface darkness influences albedo and surface energy balance. Greening and browning associated with shifts in plant species composition can alter plant-soil-microbial feedbacks with long-term effects on ecosystem structure and function, while losses of tundra vegetation can reduce the insulating capacity of soils, leading to increased permafrost thaw and subsequent changes in carbon cycling and hydrological processes (Blok et al., 2010; Myers-Smith and Hik, 2013). If changes in plant species occur as a result of greening or browning, it may affect wildlife habitat and food availability, which in turn can affect community hunting practices. Other community effects of greening and browning can include access to new, local fuel sources and influence availability of jobs like firefighting.
Implications can extend from the local to global scale, including feedbacks to global climate from altered albedo and increased greenhouse gas release from thawing permafrost. Also, as discussed in the Global Context section earlier in this proceedings, northern latitudes can affect measured global atmospheric CO2 concentration, indicating the critical importance of these ecosystems in helping to moderate climate change and suggesting that greening or browning over large tracts of this region could have impacts felt globally.
This section provides greater detail about a few of these implications and demonstrates a range of potential effects.
ALBEDO AND ENERGY FEEDBACKS
Land surface temperature results from the Earth’s surface energy balance, which is the equilibrium between radiative, ground, and turbulent heat flux, explained Thomas O’Halloran, Clemson University. Radiation values (both shortwave and longwave) relate to albedo and atmospheric and surface temperatures, while heat fluxes (sensible, soil, and latent) manifest as air and ground temperature and humidity change. Among these properties, albedo is the most important in the context of climate because changes to it create a radiative forcing at the top of the atmosphere with a climate sensitivity comparable to CO2 (Davin and de Noblet-Ducoudré, 2010). O’Halloran and colleagues have developed a new method using radiative kernels to estimate how surface albedo change translates into net energy change at the top of the atmosphere. This method offers a simplified, more accessible approach when compared to the complex radiative transfer models and general circulation models that are commonly used.
The primary biological controls on the surface energy balance are vegetation structure and physiology, O’Halloran stated. Quantitative frameworks have been developed that demonstrate that equilibrium temperature change is a direct function of changes in ecosystem properties (e.g., Devaraju et al., 2018; Lee et al., 2011), suggesting that patterns of greening and browning may affect surface temperature directly. Indirectly, changes in sensible and latent heat fluxes associated with vegetation change can occur, with implications for cloud formation, incoming radiation, and air temperature. Collectively, these findings indicate there are local and global effects of vegetation change that drive feedbacks and perhaps teleconnections, which can be more fully evaluated in coupled Earth System Models. Such effects could be key in predicting the spatiotemporal propagation of greening and browning.
For the circumpolar region, there are also some relatively unique relationships between surface energy balance and vegetation (Pearson et al., 2013). Albedo is strongly influenced by the type of vegetation present and its interaction with snow, according to O’Halloran. Extensive snow cover in the tundra, where few tall shrubs stick through the snow surface, and in deciduous forest where no leaves are present, results in a highly reflective ground surface that produces a high albedo extreme (Loranty et al., 2014; Shuman et al., 2011) particularly during the spring, when the snow is still present and the sun is also shining across the region. In contrast, areas with tall shrub cover and evergreen boreal forest, where vegetation is present above the snow surface, are more absorptive and produce a low albedo extreme. The only factor that mitigates the climate effects of albedo extremes is low insolation, driven by the sun’s angle and day length at high latitudes. Another energy balance term that may be uniquely important to high latitudes is ground heat flux; this has largely been ignored by surface energy balance studies in other regions, said O’Halloran, but is critical for evaluating permafrost thaw in this context.
BIOPHYSICAL EFFECTS
Adrianna Foster, Northern Arizona University, discussed how climate change and disturbance can change boreal forest tree species composition, productivity, and structure, with implications for a wide range of biophysical characteristics. Studies have demonstrated that individual tree species vary in productivity (e.g., Beck et al., 2011), have differing plant-soil interactions (Johnstone et al., 2010; see Figure 4), and respond differently to climate variables such as precipitation and temperature (e.g., Cahoon et al., 2018). At the same time, there are strong interactions among vegetation, climate, and disturbances that occur across multiple temporal and spatial scales that can be difficult to predict and yield highly variable results when modeled. Individual [tree]-based, high resolution models can aid in simulating observed changes and interactions and may be crucial for accurate prediction of future ecosystem states, Foster said. To address the known uncertainties in linking climate, disturbance, and vegetation factors, Foster and colleagues have produced results using the individual-based ecological model, University of Virginia Forest Model Enhanced (UVAFME), which were presented at the workshop.
Foster described how UVAFME simulates tree-level response to forces such as precipitation, temperature, and soil characteristics, and provides information on forest structure, composition, and productivity change following fire disturbance with and without climate change included. Results of this model can be scaled from plot to landscape-scale to generate expected forest biomass, tree size structure, and species composition on an area basis and over time. Model output shows that fire disturbance can shift forests toward smaller tree stems, reduced aboveground biomass, and increased deciduous tree species cover relative to pre-fire conditions. This can inform trends in greening and browning by demonstrating changes in vegetation cover following disturbance, where systems could shift from browning following an event to greening during recovery. Additionally, the extent of greening differs among tree species, especially when considering evergreen versus deciduous stands. Simulated fire concurrent with climate change led to greater deciduous forest dominance and yielded reductions in the soil organic layer and deepening permafrost thaw that did not recover over time, which has potential implications for long-term carbon storage and greenhouse gas release from soils. Shifts in vegetation can also directly feedback to climate via albedo, as discussed by O’Halloran, with increasing deciduous forest cover increasing forest albedo and causing a relative cooling effect on the landscape in interior Alaska. However, UVAFME simulations suggest an expansion of conifer forest northward, which may reduce albedo at the range edge. Other notable biophysical characteristics Foster mentioned that can change as a result of disturbance and tree species shifts include fluxes of CO2 and other greenhouse gases, biogenic volatile organic compounds, evapotranspiration, soil moisture, and nutrient cycling.
ECOLOGICAL EFFECTS
Michelle Mack, Northern Arizona University, explored the ecological linkages and feedbacks among vegetation composition, ecosystem properties, and disturbance at high latitudes. Understanding these interactions can help to interpret trends in greening and browning and how they may change over time as ecosystems first experience disturbance and then recover to reach stable states that may differ from pre-disturbance conditions. Ecosystems exert strong controls over the abundance and distribution of plant species, which can affect the degree of greenness the landscape exhibits. At the same time, plant functional traits can create ecosystem-level feedbacks that modulate energy (e.g., albedo) and water flux and alter community interactions and soil resources (nutrient and carbon accumulation and cycling, rooting depth, mycobionts, etc.). These plant traits often covary and exhibit associations across species (syndromes) that either are caused by or emerge from phylogeny/historical constraints or evolutionary tradeoffs. A classic example is plant-soil-microbial feedbacks where plants in low-resource environments tend to exhibit relatively low growth rates and produce long-lived, poor quality litter that decomposes and releases nutrients slowly, which feeds back to stabilize or reinforce low nutrient availability.
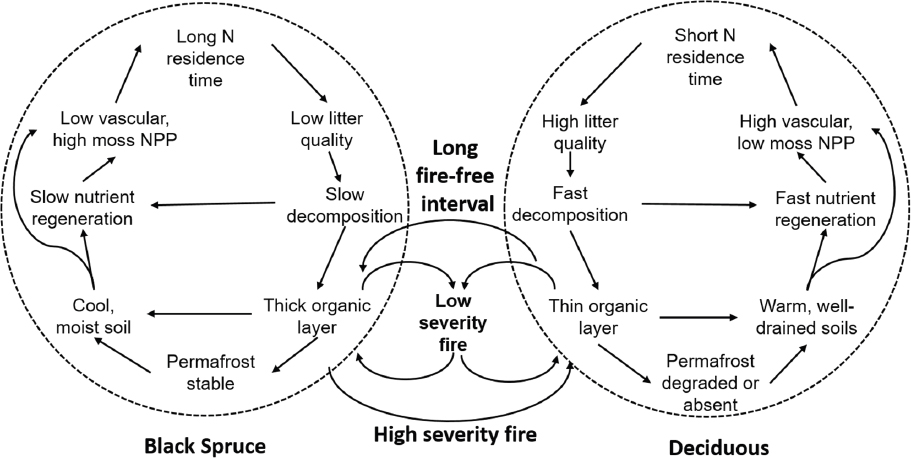
Addition or removal of key species can determine the balance between negative or stabilizing feedbacks that tend to reinforce the existing ecosystem and keep it in the same state (which may not show a trend toward greening or browning) and positive or destabilizing feedbacks that might push an ecosystem toward a new state (which might lead to an observable greening or browning trend). Disturbances can affect this stability by opening up space for recruitment of new individuals, genotypes, and species that allow for community reorganization. As demonstrated by many workshop participants, the disturbance may also drive a browning trend initially. In Alaskan boreal forest, increased fire frequency, extent, and severity are resulting in destabilizing feedbacks that change vegetation composition and associated ecosystem characteristics (see Figure 4). Historically, this region has been dominated by black spruce (which exhibits the low resource plant-soil-microbial feedbacks explained in the previous paragraph) and
relatively low-intensity fires with limited burning of the soil organic layer which, combined with large seed releases from semi-serotinous cones, would allow for spruce replacement or establishment of a mixed spruce and deciduous stand post-fire. During severe fires however, more of the soil organic layer is lost, which opens space for deciduous tree species to establish, propagating their different plant functional traits through the ecosystem and causing a shift to a new state that stabilizes with plant-soil-microbial feedbacks of a relatively higher-resource environment. Mack noted that a major implication of this shift to deciduous dominance is an observed increase in net ecosystem carbon storage in wood resulting from higher nutrient-use efficiency and the transfer of nitrogen from the soils to the wood.
WILDLIFE
Elie Gurarie, University of Maryland, provided an overview of the complex linkages among migratory caribou, vegetation change, and other factors. Caribou are the most important herbivores in the north and are distributed across ecotypes around the circumpolar region. They serve as an important prey species and are vital to subsistence and the cultural economy of indigenous communities. Migratory caribou travel in large herds located on tundra and bare ground, and many populations have exhibited dramatic declines in recent years. In the spring, migration timing is influenced by the condition of mothers, while in summer, insect harassment (i.e., mosquitoes) is an important driver of caribou movement, Gurarie said.
Many questions remain about the links between caribou and vegetation productivity. Observed latitudinal shifts in caribou migration do not correlate well with NDVI, Gurarie said. Using resource selection functions, Gurarie and colleagues have evaluated the probability of use of various habitats by caribou and have found high preference for diverse tussocks and shrub tundra, although a time series to explore how changes in vegetation may affect these relationships has not been conducted. Other work shows a relatively weak relationship between caribou movement and enhanced vegetation index (EVI; similar to NDVI) for tundra and grassland vegetation (Rickbeil et al., 2018), with no evidence of shrubs being selected for as forage. Biological mechanisms that may affect caribou-vegetation interactions include anti-herbivory defenses by some shrub species, making them less palatable and more likely to establish in tundra (Bryant et al., 2014), and compensation of shrubs to varying degrees of browsing that may change growth allocation (Champagne et al., 2012). Gurarie noted other controversial studies that suggest a phenological mismatch between caribou calving times and food availability (Post and Forchhammer, 2008), as well as work indicating that greening promotes declines in caribou populations (Fauchald et al., 2017).
Additional focus on understanding how vegetation change and patterns of greening and browning affect animals and how animals then influence that vegetation is needed, Gurarie said. Research would benefit from new land cover maps and projections of future land cover change. Additionally, integration of mechanisms identified on the ground and large-scale observations would improve understanding of the complex linkages among vegetation, caribou, and other factors affecting caribou populations. Placing cameras on caribou could also be a way to capture vegetation patterns and provide ground-based validation for other data sources.
SOCIO-ECONOMICS
Jennifer Schmidt, University of Alaska Anchorage, shared results from interviews she has conducted in several Alaskan communities designed to identify how people use, observe, and value the land. She noted that human observations, when appropriate terminology is used, are a potentially powerful
untapped tool to help assess vegetation changes. Schmidt suggested the working hypothesis that, as you increase complexity in asking about environmental observations, you tend to get more diverse responses. For instance, when community residents are asked about the size of willow shrubs (an important food source for moose), there is general consensus that size has increased. If residents are asked more generically about shrubs though, responses are more mixed with about half stating there is no change and the rest noting either they are increasing in size/extent or decreasing.
Schmidt discussed both the benefits and negative impacts of vegetation change for communities. For example, increased shrub cover may provide wind and snow barriers and riverbank stabilization. For energy production, when woody species increase in cover, the wood may be used as a much cheaper fuel source than fossil fuels, and examples of this now occurring in interior Alaska were mentioned. The long-term sustainability of a shift toward greater fuel wood reliance as the wood around communities is harvested remains unknown. Shrubs could also affect the efficiency of bifacial solar panels if they influence albedo because these panels capture light reflected off the snow surface. For hunters, greater vegetation cover may bring animals like moose closer to communities, reducing distance traveled to hunt, although it becomes harder to see moose, bears, and other wildlife in shrubs, which could pose risks. Increased shrub cover along riverbanks is unfavorable because it hinders visibility for hunters using the river for transport. In open tundra, shrubs can hide game like ptarmigan and may also create deep snow pockets in which snow machines can become stuck when traveling across the landscape in winter months. Finally, a greater extent of woody plants can increase fire occurrence, creating greater risks to communities while at the same time providing firefighting jobs and possibly salvageable fuel wood post-fire.
Schmidt also noted that the extent to which human activities influence shrub expansion is unclear, which could be further explored in the context of drivers of northern latitude greening and browning.
This page intentionally left blank.






