
17
Carbohydrates, Protein,, and Performance
Bonnie J.Spring,1 Regina Pingitore, and Jen Schoenfeld
INTRODUCTION
Investigators have shown great interest in diets that can optimize performance in the workplace, the athletic field, and military combat situations (Kanarek and Marks-Kaufman, 1991; Lieberman, 1989; Logue, 1986; Spring, 1986). Among the candidate food constituents that might enhance performance, carbohydrates have long been accorded a special place. For example, since 1985, carbohydrates have commandeered the largest share of the U.S. Army Research Institute of Environmental Medicine’s research effort on performance-enhancing food components, absorbing 60 percent of the research effort (see Chapter 3). This chapter discusses the effects of carbohydrates on mood, cognitive performance, satiety, and physical endurance. The information presented here suggests that behavioral effects are systematically influenced by the caloric and macronutrient compositions of the meal, especially by its ratio of carbohydrate to protein; by the time of day it is consumed; by individual differences, including the temperament and usual eating habits of the
consumer; and by the surrounding psychological context, including the influences of motivation and stress.
THE HEURISTIC IMPORTANCE OF FOLK WISDOMS
Ironically, two folk beliefs have supplied much of the impetus behind research on carbohydrates and behavior (Spring and Alexander, 1989). One folk wisdom prescribes how to acutely achieve peak performance, suggesting that ingestion of simple sugars can attain this objective. The other specifies how to attain performances that need to be elicited long after eating or sustained at a high level for a prolonged period of time. Conventional wisdom endorses the use of complex carbohydrates for that objective, suggesting that they support optimal performances that stably endure. Both folk beliefs share an assumption that fluctuations in plasma glucose levels parallel and mediate the emergence of peaks and troughs in physical and mental performance. Ordinarily, one would cursorily skim over folk wisdoms and proceed rapidly to the scientific theories that have guided research on diet and behavior. In this case, however, folk wisdoms have exerted the more powerful formative influence; therefore, this chapter describes them in some detail.
Simple Sugars
Popular wisdom holds that the energy needed to achieve a stellar performance is as accessible as the nearest candy bar (Dufty, 1975). Nearly 75 percent of Americans regard sugar as a source of quick energy, according to nationwide attitudinal surveys (Fischler, 1987). The simple carbohydrates in sweet snack foods are touted to generate a burst of stamina—the “sugar buzz”—that parallels and directly results from a steep rise in plasma glucose levels. By extrapolation from the metaphor that blood sugar energizes metabolic processes, sugar becomes the presumptive energizer of behavioral processes as well (Spring and Alexander, 1989). Empirical support for the proposition that an acute rise in plasma glucose levels translates to increased mental energy is minimal at best (Gonder-Frederick et al., 1989; Spring et al., 1987). As described later, at least within the bounds of normoglycemia, this particular proposition is probably best regarded as the reification of a metaphor.
A corollary to the folk wisdom that simple sugars offer a quick pick-me-up is the conviction that the benefits are short-lived (Dufty, 1975). Perhaps because many Americans are Puritans at heart, folk beliefs threaten that few benefits can be gleaned without a cost. In the case of simple carbohydrates and
performance, that cost is “reactive hypoglycemia”: an alleged sudden plummeting of plasma glucose levels that occurs because the initially steep rise in plasma glucose level triggers insulin overproduction (Harris, 1924). In folk wisdom, at least, the extreme glucose fluctuations that result from eating simple sugars propel the performer on a roller coaster ride from superproductivity to collapse. The fact that reactive hypoglycemia is far too rare a clinical condition (Permuth, 1976) to explain the modal performance characteristics of average individuals is problematic for this theory. The proposition that a less exaggerated fall in glucose within the normoglycemic range could have adverse functional consequences remains unsupported, although much research on carbohydrates and behavior has been guided by this assumption (Prinz et al., 1980; Virkkunen, 1986).
Complex Carbohydrates
Folk wisdom’s dietary preventive against peaks and troughs in plasma glucose levels is ingestion of complex carbohydrates. By delaying and blunting both the rise and the fall of the plasma glucose concentration, starches are assumed to ensure a sustained source of metabolic energy, allegedly yielding stabler performance. Unlike the assumption that simple sugars commonly provoke reactive hypoglycemia, folk wisdoms about complex carbohydrates have at least some empirical support. Some starches, such as bread, do have lower glycemic indices than some simple carbohydrates, such as glucose (Jenkins et al., 1981). On the other hand, the glycemic indices for other starches, like potato, are virtually the same as that for glucose (Crapo, 1984). These findings, combined with evidence that parameters like fiber content, food form, digestibility, cooking method, and eating rate critically influence the glycemic response, have challenged the generalization that complex carbohydrates reliably engender lower glycemic indices than simple sugars do (Crapo, 1984).
It also remains unclear whether, at some interval after eating, higher glucose levels translate into better performances. Although some findings support this inference (Pollitt et al., 1981), others do not (Cromer et al., 1990). Nonetheless, a considerable research effort has aimed to promote stably high levels of performance by identifying diets that prolong the tail of the glucose curve, yielding moderate, sustained glucose elevations long after eating. Tests of large bolus doses or intermittent doses of complex carbohydrates, as well as doses that incorporate fiber and even protein (Arvedson et al., 1969), have all been justified on this basis.
DEFINING THE TARGET BEHAVIORS: MOOD, PERFORMANCE, SATIETY
What behaviors promote success in the military combat environment, and what dietary interventions will best support them? Stereotyped images of war suggest that the skills needed to succeed in military combat closely resemble the physical capabilities needed to excel in sports requiring physical endurance. Indeed, even today, military training places heavy emphasis on developing physical strength, stamina, and endurance. On the other hand, the nature of military operations has changed so greatly that, especially in combat situations, many contemporary soldiers work not from the trenches but, rather, from the seats of heavily computerized vehicles. Perhaps even more than previously, today’s combat environment is as likely to tax mental and emotional wherewithal as it is to strain physical endurance. Characteristic of the combat situation are demands to generate complex cognitive or psychomotor performances under conditions of boredom, physical discomfort, prolonged sleep deprivation, or severe emotional distress. Because sensorimotor and cognitive performances are essential to all domains of military operations during war or peacetime, they are given primacy in this chapter. Because it may be necessary for long intervals to elapse between eating occasions, satiety is briefly discussed. Finally, the chapter touches peripherally upon physical endurance because that topic is addressed by other contributors to this volume.
Mood: An Underrated Outcome
Mood has been underrated and performance has been overrated as indices of dietary influences on behavior. Although it can be argued that tests of mood assess subjective, unverifiable phenomena, mood scores are highly correlated with cognitive performance, especially under extreme circumstances. Consider the findings of Bugge et al. (1979), who assessed mood and cognitive performance among Norwegian military cadets who underwent 4 days of nearly continuous sleep deprivation in addition to caloric restriction. Scores on tests of coding, logical reasoning, and letter cancellation deteriorated markedly under these conditions. Furthermore, correlation coefficients between performance and mood ranged between r=0.85 and 0.90 for positive mood and between r=-0.86 and -0.94 for negative mood. These findings suggest that under highly adverse circumstances, tests of mood and cognitive performance convey virtually the same information.
Of potentially greater importance are contexts in which mood and performance convey different information. Under ordinary circumstances, most performance tests yield only coarse indicators of organismic burden or stress.
This is because few sensorimotor or information processing tasks are so demanding of cognitive resources that they utilize full cognitive capacity (Kahneman, 1973; Wickens, 1984). Because reserve or ancillary resources are available to be allocated to whatever task is at hand, performance can remain unimpaired long after distress is evident in other functional spheres including affective state and physiological homeostasis. Indeed, the experience has been that, whereas mood reliably reflects the effects of caloric deprivation, sensorimotor and cognitive performances are insensitive to caloric restriction, except under conditions that impose an additional burden (Spring et al., 1992). Two added burdens that make caloric deprivation deficits become visible are stress or demands for continuous performance without allowing recuperative time between tasks. It is suspected that these conditions elicit deficits because they drain spare capacity and thus unmask deterioration that would otherwise not be visible.
To illustrate how insensitive performance indicators can be, consider the findings of Diaz et al. (1991), who studied the productivities of two groups of Gambian subsistence farmers during the hungry season as they worked to build a road. The farmers’ work was to shovel gravel into wheelbarrows, wheel it 1.5 km to the road, unload the gravel, and repeat the cycle for 8 h. Farmers were studied under two feeding conditions: one supplying an unlimited supply of energy-dense foods; the other supplying insufficient calories to match energy expenditures. The caloric restrictions were sufficient to cause significant weight loss. Nonetheless, work productivity was unaffected: no effects could be detected on the total number of loads transported, the number of loads per working hour, or the amount of time per load. The authors attribute these results to a motivational incentive so powerful that it overrode the effects of severe energy deficit: the men were paid $0.63 per load, a generous wage by local standards.
The results of Diaz et al. (1991) can be explained by resource models, which propose that motivation, interest, and effort actually increase functional performance capacity (Kahneman, 1973). A more telling interpretation comes, perhaps, by analogy to Sholem Aleichom’s story about the man who decided to save money by feeding his horse only once per day. The horse still pulled the cart just as before. Then, to save more money, the man chose to feed the horse only three times per week. Still, the horse pulled the cart. Finally, the man decided to feed the horse only once a week. The horse died. “What a shame,” said the man, “just when the horse finally got used to eating three times a week!” One could infer, as the man did, that no problem existed until the horse died, but this would be the wrong conclusion. The correct conclusion is that the horse’s performance was an insensitive measure of the adverse effects of caloric restriction, since the horse could still walk even when it was starving to death. To the extent that performance lags behind in reflecting adverse effects of dietary or other strictures, it is useful to have more sensitive
early warning signs of impending decline. Subjective fatigue or dysphoria may offer such a potential.
Mood state can also serve as a proxy variable for approach motivation, an attribute that can profoundly influence combat performance. A soldier’s early proactive response to potential danger can make the difference between an averted disaster and a tragic loss of life. In anecdotal accounts of friendly fire accidents presented at the conference on which this volume is based, a common denominator was that the soldier had been aware of present danger but had not acted because of fatigue, lethargy, or hesitancy. If, in that state of failure to spontaneously “go the extra mile,” the recruit had been required to perform a cognitive task, he or she could probably have done so without error. The performance task would, thus, not capture highly salient information about the soldier’s state: i.e., his or her lowered motivation to spontaneously initiate action of his or her own accord. Subjective fatigue can have profound motivational consequences in the combat environment.
CARBOHYDRATES AND FATIGUE
The authors’ interest in dietary effects on mood stemmed from an initial study that compared the behavioral responses of 184 healthy adult men and women to a sucrose-rich, high-carbohydrate, low-protein meal versus those to a high-protein meal (Spring et al., 1983). Contrary to the popular wisdom that sugar is activating, the female subjects reported fatigue, and the male subjects reported calmness after eating carbohydrates. The gender difference was interpreted as evidence of a more intense but similar mood in women, because gender differences in average body weight render the carbohydrates in the fixed-size meal a proportionately higher dose for women.
In subsequent studies, a high-carbohydrate meal, given in sufficient quantity and eaten at lunchtime, was also found to trigger fatigue in men to a greater extent than an isocaloric protein meal did (Lieberman et al., 1986a; Spring et al., 1986). Moreover, the carbohydrate-rich meal did not need to be rich in simple carbohydrates to engender fatigue; a high-starch, protein-poor lunch also triggered greater fatigue than a protein-rich lunch.
Mechanisms: Opioids? Hypoglycemia? Serotonin?
Spring and colleagues (1989) studied three mechanisms that might explain the fatiguing effects of a high-carbohydrate meal. In a repeated-measures design, healthy adult women ate a standard breakfast in the laboratory and were assessed before and after fasting through the noon hour or eating one of
three lunches that were provided in counterbalanced order. The protein lunch was 318.8 g of turkey breast plus mayonnaise, supplying 780 kcal, 105.0 g of protein, and 33.3 g of fat. The carbohydrate and balanced lunches each consisted of six sweet, cookie-type lunch bars. The carbohydrate lunch supplied 799 kcal, 105 g of carbohydrate, 0.7 g of protein, and 42.7 g of fat. The balanced lunch, offering carbohydate to protein in approximately a 3/1 ratio, supplied 774 kcal, 76.0 g of carbohydrate, 27.7 g of protein, and 40 g of fat. Self-reported fatigue and plasma glucose, serum insulin, and plasma amino acid concentrations for each condition are shown in Figures 17–1 to 17–4, respectively.
Sweet Taste
One hypothesis was that both the carbohydrate and the balanced lunches would have comparable calming effects on mood because both were sweet and would, therefore, presumably exert similar effects on endogenous opioids (Fantino et al., 1986; Getto et al., 1984; Morley et al., 1981). Inspection of Figure 17–1 shows that this hypothesis was incorrect. The carbohydrate meal
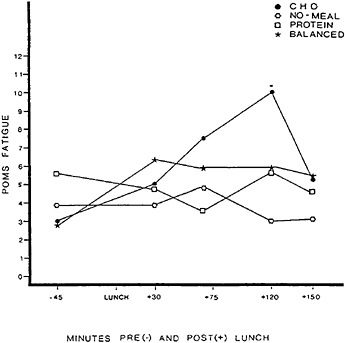
FIGURE 17–1 Fatigue reported on the Profile of Mood States (POMS) before and after eating a high-carbohydrate, low-protein lunch (CHO); a high-protein, low-carbohydrate lunch (PROTEIN); a balanced lunch containing both carbohydrate and protein (BALANCED); or fasting (NO-MEAL). SOURCE: Spring et al. (1989), used with permission.
was the only lunch that triggered a significant increase in fatigue, which occurred about 2 h after eating. In spite of their hedonic similarity to the carbohydrate cookies, the balanced cookies did not trigger fatigue.
Reactive Hypoglycemia
The next hypothesis was that if reactive hypoglycemia is responsible for carbohydrate-induced fatigue, then drowsiness should be greatest after the carbohydrate lunch and should occur in temporal proximity to an overproduction of insulin and a sharp decline in plasma glucose to hypoglycemic levels. The carbohydrate lunch did indeed trigger fatigue. As Figure 17–3 shows, however, the fatiguing effect of the carbohydrate meal could not be explained by a unique effect on insulin, because both the carbohydrate and the balanced lunches produced roughly comparable insulin responses. Furthermore, as Figure 17–2 shows, fatigue after the carbohydrate lunch could not be attributed to hypoglycemia because the plasma glucose level remained elevated at the time when fatigue occurred. The findings do not rule out the possibility that drowsiness after a carbohydrate meal could be triggered by a declining level of plasma glucose rather than by an aberrant insulin response or an absolute
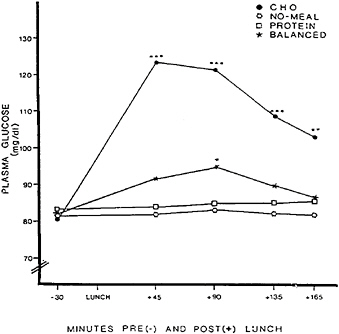
FIGURE 17–2 Plasma glucose (in milligrams per deciliter) before and after eating a high-carbohydrate, low-protein lunch (CHO); a high-protein, low-carbohydrate lunch (PROTEIN); a balanced lunch containing both carbohydrate and protein (BALANCED); or fasting (NO-MEAL).
SOURCE: Spring et al. (1989), used with permission.
hypoglycemic level of glucose. The results are, however, incompatible with a simple explanation on the basis of reactive hypoglycemia.
Plasma Amino Acids
A final hypothesis was that fatigue after eating carbohydrates results from effects on plasma amino acids, which, in turn, influence the synthesis and release of brain serotonin. Consumption of a high-carbohydrate, protein-poor meal increases the ratio of plasma tryptophan to other large neutral amino acids, which predicts increased brain tryptophan influx (Fernstrom and Wurtman, 1971). Since tryptophan is the precursor of serotonin and since the enzymes that catalyze serotonin synthesis are not fully saturated, serotonin synthesis is substrate dependent. An increase in brain tryptophan influx therefore leads to an increase in both brain serotonin synthesis and release, insofar as the latter is indexed by the cerobrospinal fluid (CSF) levels of 5-hydroxyindoleacetic acid (5-HIAA), serotonin’s major metabolite (Fernstrom and Wurtman, 1972; Wurtman et al., 1980; Yokogoshi et al., 1987). Because
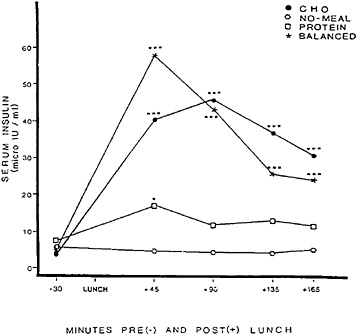
FIGURE 17–3 Serum insulin (micro-international units per milliliter) before and after eating a high-carbohydrate, low-protein lunch (CHO); a high-protein, low-carbohydrate lunch (PROTEIN); a balanced lunch containing both carbohydrate and protein (BALANCED); or fasting (NO-MEAL). SOURCE: Spring et al. (1989), used with permission.
serotonergic midbrain raphe neurons play a role in initiating the onset of sleep (Hartmann and Greenwald, 1984), drowsiness is a likely accompaniment of increased serotonin synthesis in a healthy brain. As Figure 17–4 shows, the high-carbohydrate, low-protein lunch did significantly increase the ratio of plasma tryptophan to the other large neutral amino acids that share the same transport system across the blood-brain barrier. Moreover, consistent with the hypothesis, fatigue did emerge at about the same time that the tryptophan ratio increased, although it remains unclear why fatigue dissipated even though the ratio remained elevated.
Thus far, carbohydrate-induced fatigue has emerged consistently and is most plausibly explained by the serotonin hypothesis. Neither the sweetness of lunches, nor their effects on insulin, nor their absolute effects on glucose predicted whether meals would induce fatigue. On the other hand, drowsiness occurred reliably about 2 h after a carbohydrate-rich, virtually protein-free lunch, and fatigue appeared in parallel with a rise in the tryptophan ratio.
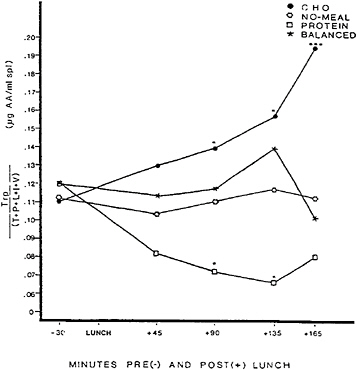
FIGURE 17–4 Ratio of plasma tryptophan (Trp) to the sum of plasma tyrosine (T), phenylalanine (P), leucine (L), isoleucine (I), and valine (V) before and after eating a high-carbohydrate, low-protein lunch (CHO); a high-protein, low-carbohydrate lunch (PROTEIN); a balanced lunch containing both carbohydrate and protein (BALANCED); or fasting (NO-MEAL). AA, amino acids. SOURCE: Spring et al. (1989), used with permission.
Breakfast Versus Lunch
Recently, the research emphasis has shifted from the midday lunch context to the study of breakfast. The results of three studies are described only briefly because they are not yet published. In a nutshell, the results suggest that there are both commonalities and differences between the manner in which carbohydrates affect mood in the morning versus the early afternoon. The core difference is that macronutrient effects are displayed across a different diurnal context. At noontime all meals exceeding about 300 cal tend to engender some fatigue. In healthy adults, both protein-rich and protein-deficient, carbohydrate-rich meals cause some drowsiness, but the latter cause greater drowsiness (cf. Rosenthal et al., 1989; Spring et al., 1983). In the morning, the diurnal pattern is reversed. Among regular breakfast eaters, a breakfast of any macronutrient composition tends to lessen fatigue in comparison with the baseline, whereas fasting leaves the person as drowsy as upon awakening. The relative activating impact of the macronutrients is, however, the same as it is at lunchtime. As breakfast, a protein-rich meal has a greater antifatiguing action than does a protein-poor, carbohydrate-rich meal.
Because of the different diurnal background, tests of carbohydrate effects on mood would be expected to produce somewhat different outcomes when the experimental meals are served as lunch rather than as breakfast. A carbohydrate test meal served as lunch should produce greater fatigue than fasting or than an isocaloric protein meal. A carbohydrate test meal served as breakfast should produce less fatigue than fasting but less activation than a protein test meal.
The Balanced Meal Threshold
To increase the plasma tryptophan ratio, a meal must be both carbohydrate-rich and protein-poor. Considerable effort has been devoted to determining just how rich in carbohydrate and how poor in protein a meal must be in order to elevate the plasma tryptophan ratio. In fasted rats, a meal containing 70–75 percent of an insulin-secreting carbohydrate increases the plasma tryptophan ratio, whereas a meal containing 25 percent carbohydrate fails to do so (Yokogoshi and Wurtman, 1986). In addition to being abundant, a carbohydrate must also be insulin-secreting (e.g., glucose, sucrose, and dextrose) if it is to elevate the tryptophan ratio. In contrast, fructose, the sugar that predominates in fruits, has a relatively weak effect on insulin. Consequently, a high-carbohydrate breakfast chiefly comprising fruit juice, even if sweetened by sucrose, fails to increase the plasma tryptophan ratio (Teff et al., 1989).
A protein-rich meal decreases the tryptophan ratio probably by contributing much larger quantities of large neutral amino acids other than tryptophan to the bloodstream. Moreover, a protein-rich meal has been found to decrease CSF tryptophan and 5-HIAA levels (Perez-Cruet et al., 1972). Proteins, however, differ in how effectively they suppress a carbohydrate-induced rise in the plasma tryptophan ratio. For example, in rats, whereas 5 percent casein or 10 percent peanut meal or gelatin fully blocks the rise triggered by a 70 percent carbohydrate meal, 10 percent lactalbumin fails to do so, and 10 percent egg white does so only partially (Yokogoshi and Wurtman, 1986).
For humans, how little protein blocks a carbohydrate-induced rise in the tryptophan ratio, creating what is, from the perspective of this ratio, a balanced meal? To answer this question, Teff and colleagues (1989) served healthy male subjects breakfast puddings containing either 0, 4, 8, or 12 percent protein. They also studied a conventional breakfast of Danish pastry and coffee, in which protein was about 10 percent the amount of carbohydrate. The ratio of plasma tryptophan to other large neutral amino acids was determined before and 2 h after each breakfast. The results are shown in Table 17–1.
Teff and colleagues (1989) found that only the pure carbohydrate breakfast elevated the plasma tryptophan ratio significantly. The 4 percent protein breakfast also elevated the ratio, although not significantly. On the other hand, breakfasts, including Danish pastry, that contained at least 8 percent protein actually lowered the tryptophan ratio slightly.
Overview of Studies on Carbohydrates and Fatigue
If increased brain tryptophan influx and serotonin synthesis is the mechanism that causes drowsiness after a high-carbohydrate meal, then carbohydrate meals that actually increase the tryptophan ratio should be the ones most likely to induce fatigue. In other words, meals that contain less than 4 percent protein should be the ones most likely to cause drowsiness. Table 17–2 presents an overview of studies that tested whether a high-carbohydrate meal induces postmeal fatigue.
Inspection of Table 17–2 indicates that carbohydrate-induced fatigue was observed most reliably in studies that used a test meal containing 0 percent protein. In contrast, fatigue was not consistently detected following carbohydrate meals that were of more balanced composition, containing more than 4 percent protein. It is also noteworthy that only one of three studies of breakfast observed carbohydrate-induced fatigue. As discussed earlier, for regular breakfast eaters, a morning meal is more likely to have activating than sedating effects.
TABLE 17–1 Effect of Dietary Treatments on Plasma Amino Acid Ratios
Military Applications
Carbohydrate-rich, protein-poor foods could intentionally be applied to engender fatigue in troops required to go to sleep at unusual hours. It is likely that this application would be most effective in the afternoon and possibly evening hours but would not be recommended for morning usage. Whereas most spontaneously selected meals would incorporate too much protein to be useful soporifics, macronutrient control would be possible in rations used to induce sleep in military personnel.
Individual Differences in Affective Response to Carbohydrates
Individual differences characterize the affective response to carbohydrates. In contrast to the fatiguing effects just described, individuals with certain
TABLE 17–2 Studies Testing Whether a Carbohydrate Meal Induces Greater Fatigue than a Protein Meal or Fasting Control
conditions experience activation and reduction of depression or anxiety after eating carbohydrates. The activating and mood-enhancing effects of carbohydrates have been observed in patients with seasonal affective disorder (Rosenthal et al., 1989), obese individuals who snack preferentially on carbohydrate-rich foods (Lieberman et al., 1986b), and females with premenstrual distress syndrome (Wurtman et al, 1989). Shared features of these clinical conditions include dysphoric mood and weight management difficulties arising from overeating of high-carbohydrate snacks.
Carbohydrates and Satiety
If extended time periods must elapse between eating occasions, carbohydrate-rich, protein-poor meals have a liability in that they are not very satiating. A widely replicated finding is that, weight for weight, protein is substantially more satiating than carbohydrate (Blundell and Hill, 1988). The greater satiating power of protein is both immediate and sustained. A protein-rich meal leaves a greater sensation of fullness at the end of a meal than does an isocaloric carbohydrate-rich meal (Spring et al., 1983). Furthermore, the satiating effect carries over until the next meal. For example, Teff et al. (1989) found that a high-protein breakfast with only half the energy value of a high-carbohydrate breakfast exerted an equipotent satiating effect on lunchtime food selections.
Carbohydrates and Mood: Conclusions
Several conclusions can be made about the effects of carbohydrates on mood.
-
A high-carbohydrate meal that is lacking in protein is followed by more fatigue than occurs after a meal higher in protein.
-
Diurnal differences characterize the effects of carbohydrates in the morning versus those in the early afternoon. After a high-carbohydrate, low-protein lunch, fatigue increases more than after a higher-protein lunch. After a high-carbohydrate, low-protein breakfast, fatigue decreases less than after a higher-protein breakfast.
-
The fatiguing effects of carbohydrate emerge most reliably when meals contain less than 4 percent protein.
-
Individual differences characterize the affective response to carbohydrate. In certain clinical conditions characterized by dysphoric mood, weight management difficulties, and heightened snacking on carbohydrate-rich foods, carbohydrate has activating, antidysphoric effects.
-
Weight for weight, carbohydrate is less satiating than protein.
CARBOHYDRATES AND COGNITIVE PERFORMANCE
As was the case for mood, cognitive performance is affected differently by carbohydrates taken at breakfast, at lunch, and as snacks. Therefore, these contexts are considered separately.
Breakfast
The next section on lunch discusses the postmeal dips in performance that are engendered by eating. At breakfast, however, the picture is different. The only group of subjects who have been found to show a postmeal dip in performance after eating breakfast are individuals who regularly skip breakfast (Richards, 1972). Some evidence indicates that military personnel who regularly eat breakfast exhibit greater physical fitness than those who do not (Trent and Conway, 1988). Since breakfast is one component of any appropriate meal plan that the military will implement to ensure health and fitness, the comments below pertain only to individuals who regularly eat breakfast.
A large body of research has examined whether skipping breakfast has adverse effects on the late-morning performance of cognitive tasks and whether meals of particular composition are differentially effective in preventing cognitive decline. Data are most extensive for young children, with some evidence supporting (Pollitt et al., 1981, 1983) and other evidence contradicting (Simeon and Grantham-McGregor, 1989) the hypothesis that, for healthy youngsters, skipping breakfast engenders late-morning impairment on cognitive tests. Findings for adolescents are more consistently negative, with most studies failing to find adverse effects of fasting on late-morning cognitive test scores (Arvedson et al., 1969; Cromer et al., 1990; Dickie and Bender, 1982a,b). Few studies have been carried out with adults, but in those studies that have been done in adults, decreased work output emerges reliably, and attentional or cognitive difficulties emerge intermittently (Daum et al., 1950; Tuttle et al., 1949, 1950).
Several factors influence whether late-morning performance impairments occur after skipping breakfast. First, the adverse cognitive effects of fasting are especially likely to occur in those who have a prior or current history of malnourishment (Simeon and Grantham-McGregor, 1989). Second, skipping breakfast is more likely to impair the performance of those who usually eat breakfast (Richards, 1971, 1972). Third, emotional and/or physical stress and continuous time pressure increase the likelihood that late-morning impairments will appear on tests of complex cognitive functions. Eating a breakfast of any composition works against the emergence of late-morning performance deficits, and when differences between meals are found, cognitive performance is better after the higher-protein breakfast (Spring et al., 1992).
Lunch
A large variety of cognitive performances, including vigilance, reaction time, sorting, and arithmetic, show a steady increase during the course of daylight hours (Blake, 1971). This trend is marred by a temporary decline in efficiency in the middle of the day. Some have argued that the midday decline, which has become known as the “postlunch dip,” is of entirely endogenous origins (Hildebrandt et al., 1974), but there is increasing evidence that the deterioration in performance is caused at least partly by eating lunch (Smith and Miles, 1986b). The slight plateauing of the body temperature curve that is evident in early to midafternoon (Blake, 1971) may signify a vulnerable period in the diurnal rhythm, especially in contrast to the strong morning upswing. A heavy lunch is one burden that can apparently make midday performance liabilities become manifest.
Figure 17–5 shows the findings of Craig et al. (1981) for the percent drop in efficiency on a visual discrimination task after the lunch interval for students who consumed a three-course lunch versus those who fasted. Whereas a significant impairment in perceptual discrimination occurred as a function of eating lunch, the ability to discriminate was unchanged among those who skipped lunch. The magnitude of the decline was approximately a 10 percent change in performance efficiency, about comparable to the degree of impairment that would occur after missing a night of sleep (Craig, 1986). Various studies show that the postlunch dip is at a maximum 1–3 h after lunch and dissipates afterward (Craig et al., 1986; Spring et al., 1986).
Moderating Variables
Several factors moderate the magnitude and even the occurrence of the postlunch dip in performance. One of the more powerful influences is the number of calories consumed at the lunchtime meal, which, in turn, exerts its effect in interaction with usual lunchtime eating habits. Figure 17–6 shows Craig and Richardson’s (1989) finding for the percent change in omission errors on a sustained-attention task as a function of the size of the subject’s usual lunch and the size of the experimental meal. The results indicate that a big three-course lunch of about 1,380 cal significantly increased errors on the attentional task. Moreover, the largest drop in performance was found for subjects who were accustomed to eating a light lunch. In contrast, a light lunch of less than 300 cal did not cause a postlunch dip in performance.
The macronutrient composition of the meal also influences the likelihood that a lunch will elicit a postlunch dip and affects the magnitude of performance deterioration that will be observed. Carbohydrate-rich, protein-poor meals elicit greater performance deterioration than do isocaloric protein-rich
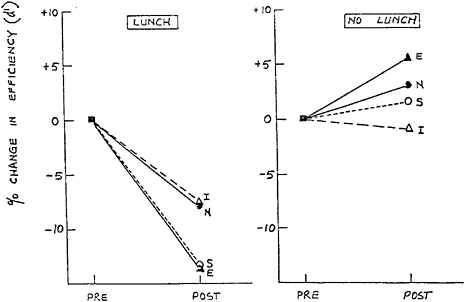
FIGURE 17–5 Afternoon drop in performance (expressed as a percentage of the morning perceptual discrimination efficiency) in lunch and no lunch groups for subjects scoring above and below the median on Eysenck Personality Inventory Introversion (I), Extraversion (E), Neuroticism (N), and Stability (S) scales. SOURCE: Craig et al. (1981), with permission.
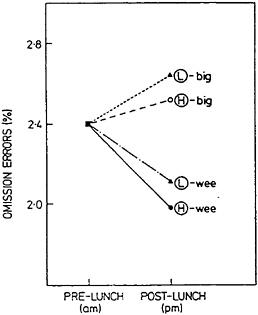
FIGURE 17–6 Postlunch percent omission errors on a sustained attention task relative to the undifferentiated prelunch covariate as a function of whether the subject usually eats a light (L) or heavy (H) lunch and whether the experimental lunch is large (big) or small (wee). SOURCE: Craig and Richardson (1989), with permission.
meals (Lieberman et al., 1986a; Simonson et al., 1948; Spring et al., 1983, 1986). A low-protein, high-carbohydrate lunch tends to induce slowing of reaction time, vigilance impairments, and decrements in sustained attention, but it does not augment susceptibility to distraction (Spring, 1984). Like their effects on mood, it is likely that the different effects of carbohydrate and protein lunches on performance will be observed for moderate-calorie meals (ranging between 300 and 1,000 kcal) and for the carbohydrate test meals that contain less than 4 percent protein. On the other hand, Smith et al. (1988) found different performance effects of larger meals that varied in their macronutrient compositions, even though all meals contained a mixture of carbohydrate and protein. Smith and colleagues reported that higher-carbohydrate meals containing either starch or sugar slowed reactions to peripheral visual stimuli, whereas higher-protein meals enhanced susceptibility to distraction.
There are also individual differences in susceptibility to the postlunch dip as a function of personality. As Figure 17–5 shows, Craig et al. (1981) found that the magnitude of the postlunch performance deterioration was greatest in stable extroverts, who can be considered to be underaroused and to have low levels of the anxiety trait. This result is consistent with findings from other studies (Smith and Miles, 1986a).
Skipping Lunch
Whether by design or happenstance, research on breakfast has focused on the relatively delayed effects of skipping the meal, whereas research on lunch has focused on the relatively acute effects of eating the meal. Given that it is sometimes necessary to skip lunch, it is worthwhile to ask whether skipping lunch results in adverse effects later in the afternoon, perhaps as a function of energy insufficiency.
Very little research has addressed skipping lunch, but two studies are available. In a laboratory setting, Kanarek and Swinney (1990) found only modest negative consquences of skipping lunch: subjects’ reading times were slowed. In a field setting, Lisper and Eriksson (1980) found the more pronounced adverse effects of skipping lunch that are shown in Figure 17–7. Their subjects drove for 8 h on four occasions, taking a rest break after 4 h. The experimenters varied the rest period—short (15 min.) or long (60 min.)—and whether the subjects were permitted to eat a hot lunch. The dependent measure, speed on a subsidiary reaction time test taken while driving, is an excellent measure of the reserve capacity available to be allocated to the driving task if needed. As the data in Figure 17–7 indicate, skipping lunch had an adverse effect on performance. By the end of the drive, subjects’ response times were slower if they had skipped lunch; i.e., they had less available spare
capacity to dedicate to driving. Lisper and Eriksson’s findings also illustrate another important phenomenon: that food does not operate in a psychological vacuum. As Figure 17–7 shows, on the day of the long pause without food, performance had deteriorated even before the lunch break. Apparently, subjects were anticipating a long, depriving wait in the middle of their 8-h drive. This finding serves as an important reminder that a meal is more than just a source of calories that reverse or prevent energy deficits. Food is also an an important psychological incentive that can enhance motivation and, hence, performance capacity.
Snacks
Carbohydrate-rich, protein-poor snacks are more likely to be eaten in the late afternoon than at any other time of day except evenings (Lieberman et al., 1989), yet no data characterize their effects on performance at that time of day. Snacks are one of the few eating vehicles likely to supply less than 5 percent protein. Carbonated beverages, graham crackers, and some cupcakes, cookies, and candies are examples of such foods. Lisper and Eriksson (1980) found late-afternoon performance deterioration that appeared to be related to energy insufficiency. To the extent that these subtle cognitive deficits are similar in
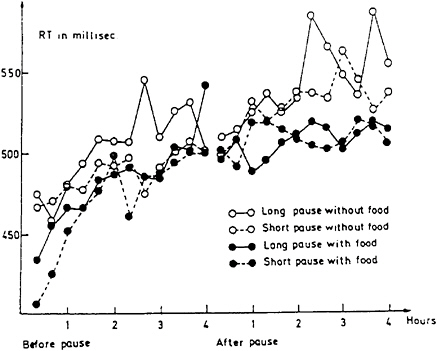
FIGURE 17–7 Changes in arithmetic means of reaction time (RT) over successive hours of driving before and after a rest break or food. SOURCE: Lisper and Eriksson (1980).
form and origin to the late-morning difficulties seen after skipping breakfast, it is plausible that late-afternoon snacking could enhance performance. Indeed, Kanarek and Swinney (1990) found support for this possibility. In their study, a calorie-rich snack improved performance on tests of sustained attention and complex cognitive functions compared with the performance after drinking a very low calorie diet soda. No differences were found between the effects of a confectionary snack versus those of a yogurt, but both snacks were balanced, containing more than 25 percent protein.
Glucose and Memory
In a series of studies of humans, Gold and colleagues report that ingesting a drink sweetened with glucose improves the memory performance of elderly subjects to a greater extent than does a saccharin-sweetened drink (Gonder-Frederick et al., 1987; Hall et al., 1989; Manning et al., 1990). They interpret their data to suggest that aging is associated with impaired brain uptake of glucose from blood, which can be attenuated by increasing circulating blood glucose levels in these subjects.
It is possible that relatively pure carbohydrate solutions have unique effects in young and elderly individuals. Nonetheless, it is striking that in the very large literature on the effects of sugars upon childrens’ cognitive performance, there are a few reports of adverse effects (Goldman et al., 1986; Rosen et al., 1988), an abundance of reports of no discernible effects (cf. reviews by Spring and Alexander [1989] and Spring et al., [1987]), and no published reports of beneficial effects.
Another interpretation of the findings of Gold and colleagues is possible. In all of that group’s published studies, the procedure has been to test patients after an overnight fast. In other words, the glucose load has served as a form of breakfast, and the saccharin load has served as a form of continued fasting. Additionally, because blood was drawn, the experimental paradigm includes the kind of stressors that can invoke cognitive impairments under morning fasting conditions. If this interpretation is correct, then the glucose load enhances memory in comparison with fasting only because it contains calories. If so, on the basis of the literature reviewed earlier in the section entitled “Breakfast,” an isocaloric high-protein load should enhance cognitive performance to a comparable or greater extent.
Conclusions about Carbohydrates and Cognitive Performance
In summary, several generalizations can characterize the effects of carbohydrates on cognitive performance at breakfast, lunch, and snacks.
-
In the morning, eating usually improves cognitive performance in comparison with fasting. The adverse effects of skipping breakfast are more likely to be observed under time pressure and stress and for regular breakfast eaters. High-protein breakfasts are more satiating than low-protein breakfasts and are somewhat more effective in sustaining late-morning performance.
-
Performance on complex tasks tends to deteriorate in the early afternoon. The postlunch dip appears to be partly due to diurnal fluctuations and partly due to caloric intake. A higher than normal caloric intake exacerbates the postlunch dip, as does protein intake of less than 4 percent. Individuals differ in their susceptibilities to the postlunch dip, and psychological influences can moderate the impact of nutritional factors on performance.
-
The research literature on sugar and childrens’ performance provides no support for the hypothesis that a sugar-rich snack improves performance. Some literature describing studies in adults suggests that snacks can improve rich performance, but the extent to which macronutrient composition influences the degree of benefit is not well understood.
CARBOHYDRATES AND PHYSICAL ENDURANCE
Thus far, in describing cognitive performance, the argument has been advanced that any benefit of carbohydrates is limited, is largely restricted to contexts in which foods break a fast or reverse caloric insufficiencies, and is less than what would be achieved by substituting or augmenting the diet with protein. The picture changes drastically when examining physical performance, however, with a high-carbohydrate diet becoming the regimen of choice.
A diet high in carbohydrates has clear-cut benefits in the case of moderate-to high-intensity, long-endurance activities. The mechanism underlying these benefits is relatively straightforward, in that there is a direct relationship between the intensity and duration of exercise and glucose utilization by the muscles. The advantages of a high-carbohydrate diet are most evident for rigorous long-endurance activities and for individuals who are in good physical condition. The benefits include postponement of fatigue, improved performance time, and improved work output. Not only is a low-fat, high-carbohydrate diet needed to bolster glycogen supplies, but there is also a need to replenish plasma glucose levels during the activity, preferably via solutions comprising glucose polymers. Wright et al. (1991), for example, obtained the best results in terms of both power output and time to exhaustion by a combination of carbohydrate feeding before and during exercise.
CONCLUSIONS AND RECOMMENDATIONS
-
The most promising indications for high-carbohydrate, low-protein foods are as soporific agents or as part of a regimen to enhance fitness for rigorous, long-endurance physical exercise.
-
Findings suggest that meals that contain a mixture of protein and carbohydrate exert more beneficial effects on cognition than do meals that are virtually protein-free. Very little is known, however, of what balance between carbohydrate and protein is actually optimal for performance.
-
Important differences exist between the behavioral effects of foods ingested at various times of day. Therefore, it is essential that any new ration or experimental food be tested in the diurnal contexts in which it will be used.
-
There are encouraging data to suggest that snacks may offer a vehicle for enhancing cognitive performance at intervals between meals. One could even speculate that intermittent snacks might offer an alternative to meals, especially for troops that are in transit. Anecdotal accounts also suggest that soldiers in combat are more willing to eat snacks than to eat full meal rations. Both because of their appeal and because of their positive effects on performance, snacks warrant additional research. Virtually nothing is known about what macronutrient composition would make an ideal snack, and there are conflicting interpretations of the potential cognitive benefits of pure carbohydrate (glucose) snacks or meals.
-
One could argue that research on diet and behavior in the military environment has overemphasized either simple cognitive or sensorimotor performances or physical performance. These domains of behavior may fail to capture other subtle and functionally very significant characteristics, such as motivation to spontaneously undertake or persist at activities, stress tolerance, or sociability. Additional work is warranted to develop sensitive measures of motivational and affective states and coping abilities to determine how these are affected by diet.
-
Eating is considerably more than a process of consuming calories and macronutrients. The motivational, sociocultural, and interpersonal aspects of eating in the combat environment warrant additional attention.
REFERENCES
Arvedson, I., G.Sterky, and K.Tjernstrom 1969 Breakfast habits of Swedish schoolchildren: Effect of protein content. J. Am. Diet Assoc. 55:257–261
Blake, M.J.F. 1971 Temperament and time of day. Pp. 109–148 in Biological Rhythms and Human Performance, W P.Colquhoun, ed. London: Academic Press.
Blundell, J.E., and A.J.Hill 1988 On the mechanism of action of dexfenfluramine: Effect on alliesthesia and appetite motivation in lean and obese subjects. Clin. Neuropharmacol. 11(Suppl. 1):S121–S134.
Bugge, J F., P.K.Opstad, and P.M.Magnus 1979 Changes in the circadian rhythm of performance and mood in healthy young men exposed to prolonged heavy physical work, sleep deprivation and caloric deficit. Aviat Space Environ. Med. 50:663–668.
Craig, A. 1986 Acute effects of meals on perceptual and cognitive functioning. Nutr. Rev. 44(Suppl.):163–171.
Craig, A., and E.Richardson 1989 Effects of experimental and habitual lunch-size on performance, arousal, hunger and mood. Int. Arch. Occup. Environ. Health 61:313–319.
Craig, A., K.Baer, and A.Diekmann 1981 The effects of lunch on sensory-perceptual functioning in man. Int. Arch. Occup. Environ. Health 49:105–114.
Crapo, P. 1984 Theory versus fact: The glycemic response to foods. Nutr. Today March 19 (April:6–11)
Cromer, B.A., K.J.Tarnowski, A.M.Stein, P.Harton, and D.Thornton 1990 The school breakfast program and cognition in adolescents. Dev. Behav. Pediatr. 11:295–300.
Daum, K., W.W.Tuttle, C.Martin, and L.Myers 1950 Effect of various types of breakfasts on physiologic response. J. Am. Diet. Assoc. 26:503–509.
Diaz, E., G.R.Goldberg, M.Taylor, J.M.Savage, D.Sellen, W.A.Coward, and A.M.Prentice 1991 Effects of dietary supplementation on work performance in Gambian laborers. Am. J. Clin. Nutr. 53:803–811.
Dickie, N.H., and A.E.Bender 1982a Breakfast and performance. Human Nutr. Appl. Nutr. 36A:46–56.
1982b Breakfast and performance in school children. Br. J. Nutr. 48:483–496.
Dufty, W. 1975 Sugar Blues. New York: Warner Books.
Fantino, M., J.Hosotte, and M.Apfelbaum 1986 Opioid antagonist, naltrexone, reduces preference for sucrose in humans. Am. J. Physiol. 251(1,II):R91–R96.
Fernstrom, J.D., and R.J.Wurtman 1971 Brain serotonin content: Increase following ingestion of carbohydrate diet. Science 174:1023–1025.
1972 Brain serotonin content: Physiological regulation by plasma neutral amino acids. Science 178:414–416.
Fischler, C. 1987 Attitudes toward sugar and sweetness in historical and social perspective. In Sweetness, J.Dobbing, ed. New York: Springer-Verlag.
Getto, C.J., D.T.Fullerton, and I.H.Carlson 1984 Plasma immunoreactive beta-endorphin response to glucose ingestion in human obesity. Appetite 5:329–335.
Goldman, J.A., R.H.Lerman, J.H.Contois, and J.N.Udall 1986 Behavioral effects of sucrose on preschool children. J. Abnormal Child Psychol. 14:565–577.
Gonder-Frederick, L., J.L.Hall, J.Vogt, D.J.Cox, J.Green, and P.E.Gold 1987 Memory enhancement in elderly humans: Effects of glucose ingestion. Physiol. Behav. 41:503–504.
Gonder-Frederick, L.A., D.J.Cox, S.A.Bobbitt, and J.W.Pennebaker 1989 Mood changes associated with blood glucose fluctuations in insulin-dependent diabetes mellitus. Health Psychol. 8:45–59.
Hall, J.L., L.A.Gonder-Frederick, W.W.Chewning, J.Silveira, and P.E.Gold 1989 Glucose enhancement of memory in young and aged humans. Neuropsychologia 27:1129–1138.
Harris, S. 1924 Hyperinsulinism and dysinsulin. J. Am. Med. Assoc. 83:729–733.
Hartmann, E., and D.Greenwald 1984 Tryptophan and human sleep: An analysis of 43 studies. Pp. 297–304 in Progress in Tryptophan and Serotonin Research, H.G.Schlossberger, W.Kochen, B.Kinzen, and H.Steinhart, eds. New York: Walter de Gruyter.
Hartmann, E., C.L.Spinweber, and J.Fernstrom 1977 Diet, amino acids and sleep. Sleep Res. 6:61.
Hildebrandt, G., W.Rohmert, and J.Rutenfranz 1974 Twelve and 24 hr rhythms in error frequency of locomotive drivers and the influence of tiredness. Int. J. Chronobiol. 2:175–180.
Jenkins, D., T.Wolever, R.Taylor, H.Barker, H.Fielden, J.Baldwin, A.Boling, H.Newman, A.Jenkins, and D.Goff 1981 Glycemic index of foods: A physiological basis for carbohydrate exchange. Am. J. Clin. Nutr 34:362–366.
Kahneman, D. 1973 Attention and Effort. Englewood Cliffs, N.J.: Prentice-Hall.
Kanarek, R., and R.Marks-Kaufman 1991 Nutrition and Behavior. New York: Van Nostrand.
Kanarek, R.B., and D.Swinney 1990 Effects of food snacks on cognitive performance in male college students. Appetite 14:15–27.
Lieberman, H.R. 1989 Cognitive effects of various food constituents. Pp. 251–272 in Handbook of the Psychophysiology of Human Eating, R.Shepard, ed. New York: John Wiley.
Lieberman, H.R., B.Spring, and G.S.Garfield 1986a The behavioral effects of food constituents: Strategies used in studies of amino acids, protein, carbohydrate and caffeine. Nutr. Rev. 44:61–69.
Lieberman, H.R., J.Wurtman, and B.Chew 1986b Changes in mood after carbohydrate consumption among obese individuals. Am. J. Clin. Nutr. 45:772–778.
Lieberman, H.R., J.J.Wurtman, and M.H.Teicher 1989 Aging, nutrient choice, activity and behavioral responses to nutrients. Ann. N.Y. Acad. Sci. 561:196–208.
Lisper, H.O., and B.Eriksson 1980 Effects of the length of a rest break and food intake on subsidiary reaction-time performance in an 8-hour driving task. J. Appl. Psychol. 65:123–126.
Logue, A.W. 1986 The Psychology of Eating and Drinking. New York: W.H.Freeman and Company.
Manning, C.A., J.L.Hall, and P.E.Gold 1990 Glucose effects on memory and other neuropsychological tests in elderly humans. Psychol. Sci. 1:307–311.
Morley, J.E., A.S.Levine, D.M.Brown, and B.S.Handwerger 1981 Evidence for in-vivo and in-vitro modulation of the opiate receptor by glucose. Soc. Neurosci. Abstr. 7:854.
Perez-Cruet, J., A.Tagliamonte, P.Tagliamonte, and G.L.Gessa 1972 Changes in brain serotonin metabolism associated with fasting and satiation in rats. Life Sci. 11:31–39.
Permuth, M.A. 1976 Postprandial hypoglycemia. Diabetic Care 25:719–733.
Pollitt, E., K.L.Leibel, and D.Greenfield 1981 Brief fasting, stress and cognition in children. Am. J. Clin. Nutr. 34:1526–1533.
Pollitt, E., N.L.Lewis, C.Garza, and R.J.Shulman 1983 Fasting and cognitive function. J. Psychiatric Res. 17:169–174.
Prinz, R.J., W.A.Roberts, and E.Hantman 1980 Dietary correlates of hyperactive behavior in children. J. Consult. Clin. Psychol. 48:760–769.
Richards, M.M.C. 1971 The effect of nutrition state on performance. Dissertation for Diploma in Nutrition, Dunn Nutritional Laboratory, University of Cambridge.
Richards, M.M.C. 1972 Studies on breakfast and mental performance. Nutrition 26:219–223.
Rosen, L.A., S.R.Booth, M.E.Bender, M.L.McGrath, S.Sorrell, and R.S.Drabman 1988 Effects of sugar (sucrose) on children’s behavior. J. Consult. Clin. Psychol. 56:583–589.
Rosenthal, N.E., M.J.Genhart, B.Cabellero, F.M.Jacobsen, R.G.Skwerer, R.D.Coursey, S. Rogers, and B.Spring 1989 Psychobiological effects of carbohydrate- and protein-rich meals in patients with seasonal affective disorder and normal controls. Biol. Psychiatry 25:1029–1040.
Simeon, D.T., and S.Grantham-McGregor 1989 Effects of missing breakfast on the cognitive functions of school children of differing nutritional status. Am. J. Clin. Nutr. 49:646–653.
Simonson, E., J.Brozek, and A.Keys 1948 Effects of meals on visual performance and fatigue. J. Appl. Psychol. 1:270–278.
Smith, A., and C.Miles 1986a Acute effects of meals, noise and nightwork. Br. J. Psychol. 77:337–387.
Smith, A.P., and C.Miles 1986b The effects of lunch on cognitive vigilance tasks. Ergonomics 29:1251–1261.
Smith, A., S.Leekam, A.Ralph, and G.McNeill 1988 The influence of meal composition on post-lunch changes in performance efficiency and mood. Appetite 10:195–203.
Spring, B. 1984 Recent research on the behavioral effects of tryptophan and carbohydrate. Nutr. Health 3:55–67.
1986 Effects of foods and nutrients on the behavior of normal individuals. Pp. 1–47 in Nutrition and the Brain, Vol. 7, R.J.Wurtman and J.Wurtman, eds. New York: Raven Press.
Spring, B., and B.L.Alexander 1989 Sugar and hyperactivity: Another look. Pp. 231–249 in Handbook of the Psychophysiology of Human Eating, R.Shepard, ed. New York: John Wiley.
Spring, B., O.Maller, J.Wurtman, L.Digman, and L.Cozolino 1983 Effects of protein and carbohydrate meals on mood and performance: Interactions with sex and age. J. Psychiatric Res. 17:155–167.
Spring, B., H.Lieberman, G.Swope, and G.S.Garfield 1986 Effects of carbohydrates on mood and behavior. Nutri. Rev. 44:51–60.
Spring, B., J.Chiodo, and D.Bowen 1987 Carbohydrates, tryptophan and behavior: A methodological review. Psychol. Bull. 102:234–256.
Spring, B., J.Chiodo, M.Harden, M.Bourgeois, J.Mason, and L.Lutherer 1989 Psychobiological effects of carbohydrates. J. Clin. Psychiatry 50 (Suppl. 5):274–333.
Spring, B., R.Pingitore, M.Bourgeois, K.Kessler, and E.Bruckner 1992 The effects and non-effects of skipping breakfast: The results of three studies. Report presented at the Annual Meeting of the American Psychological Association, Washington, D.C.
Teff, K.L., S.N.Young, L.Marchand, and M.I.Botez 1989 Acute effect of protein or carbohydrate breakfasts on human cerebrospinal fluid monoamine precursor and metabolite levels. J. Neurochem. 52:235–241.
Thayer, R.E. 1987 Energy, tiredness, and tension effects of a sugar snack versus moderate exercise. J. Personality Social Psychol. 52:119–125.
Trent, L.K., and T.L.Conway 1988 Dietary factors related to physical fitness among navy shipboard men. Am. J. Health Promotion 3:12–25.
Tuttle, W.W., M.Wilson, and K.Daum 1949 Effect of altered breakfast habits on physiologic response. J. Appl. Physiol. 1:545–559.
Tuttle, W.W., K.Daum, L.Myers, and C.Martin 1950 Effect of omitting breakfast on the physiologic response of men. J. Am. Diet. Assoc. 26:332–335.
Virkkunen, M. 1986 Reactive hypoglycemic tendency among habitually violent offenders. Nutr. Rev. 44(Suppl):94–103.
Wickens, C.D. 1984 Processing resources in attention. Pp. 63–102 in Varieties of Attention, R. Parasuraman and D.R.Davies, eds. Orlando: Academic Press.
Wright, D.A., W.M.Sherman, and A.R.Dernbach 1991 Carbohydrate feedings before, during, or in combination improve cycling endurance performance. J. Appl. Physiol. 71 1082–1088.
Wurtman, J.J., A.Brzezinski, R.J.Wurtman, and B.Laferrere 1989 Effect of nutrient intake on premenstrual depression. Am. J. Obstet. Gynecol. 161:1228–1234.
Wurtman, R.J., F.Hefti, and E.Melamed 1980 Precursor control of neurotransmitter synthesis. Pharmacol. Rev. 32:315–335.
Yokogoshi, H., and R.J.Wurtman 1986 Meal composition of plasma amino acid ratios: Effect of various proteins or carbohydrates, and of various protein concentrations. Metabolism 35:837–842.
Yokogoshi, H., T.Iwata, K.Ishida, and A.Yoshida 1987 Effect of amino acid supplementation to low protein diet on brain and plasma levels of tryptophan and brain 5-hydroxyindoles in rats. J. Nutr. 117:42–47.
DISCUSSION
RICHARD JANSEN: That was certainly an excellent review. It makes some order out of chaos. Putting this research together with your previous research, you have high carbohydrates producing a subjective feeling of fatigue. But in the context of exercise, high carbohydrate levels lengthen how long the athlete can perform.
BONNIE SPRING: To some extent, I think you are noting it is the difference between subjective and objective findings. I also think we need to acknowledge some very different uses of the term performance. I think we have created a lot of confusion for ourselves by using the generic term performance as if the same thing is being captured by the task of doing mental work at a desk and the task of running a marathon. I think that these two kinds of work are so totally different that it is hard to generalize from one to the other.
RICHARD JANSEN: Even physical performances that require prolonged endurance demands tremendous mental effort to create the motivation to keep on going and to monitor internal and external cues.
BONNIE SPRING: You raise another interesting empirical question. I have never seen a study that looked at complex decision making while somebody was running a marathon. I’ll bet it would not be too hard to conduct such a study, but I do not know anybody who has done that. I wonder if there might actually be an incompatibility between carbohydrate loading and complex cognitive functioning contrary to what you suggest. I am not sure that we can assume that athletes are really at their mentally most agile while they are running. They may feel great, but I do not know what their mental performance is. There may also be independent reasons why carbohydrate loading benefits endurance performance. For example, carbohydrates may exert pain-suppressing effects, by enhancing brain tryptophan influx.
JOHN IVY: We did a study 5 years ago and I never published the mental aspects of the study, but we had individuals go through an exhaustive workout—about 50 percent maximum oxygen consumption—which took about 4 or 4.5 h. We gave them a mental test prior to the start of the workout again at the point of exhaustion. They actually did better after they were exhausted than they did prior to the exercise.
RICHARD WURTMAN: What was the test?
JOHN IVY: It was a mathematical test. They had to do so many calculations in a set period of time. We had two groups: one in which pre-loaded with carbohydrates and one which did not, and no difference in results between the two groups was found.
EDWARD HORTON: I think what Bonnie Spring has brought up here is that you use the definition of fatigue very differently when you are talking about muscular fatigue versus mental fatigue, but I think that the issues of what happens when you are exercising and how well your brain works are very important questions to investigators.
A number of studies have looked at the effect of physical conditioning programs on mood and self-image and things like that, which brings up yet another question, which is how do carbohydrates affect one’s performance differently if one is physically conditioned than when one is not conditioned. So there are many interactions that would be very interesting to investigate.
BONNIE SPRING: There is also interesting literature on the carbohydrate loading protocols that, in the old days, involved a period of carbohydrate deprivation prior to carbohydrate loading. People following those protocols tended to get very irritable and cranky during the phase of carbohydrate deprivation.
ELDON ASKEW: How important do you think the type of snack would be in the military context of meals, skipped meals, or missed meals during the day? It might be possible to find a snack but not find a meal. Do you think we might find a performance enhancement?
BONNIE SPRING: I think you might find that a snack enhances performance if it reverses a performance impairment that was caused by caloric deprivation. When performance deficits appear as a result of caloric deprivation, for example, after skipping breakfast, those effects show up most clearly when an individual is under stress. The point has been made repeatedly that in the laboratory you do not come near the level of stress you are going to find in the field. So if performance in the field is undermined by both caloric depreivation and profound levels of stress, performance decrements should be observable and at least partially reversible by provision of calories. Snacks might also be the solution to the fact that you cannot get people in the field to eat a whole meal. Maybe you could get them to consume adequate caloric intake via a snack at every 4-h intervals.
DAVID SCHNAKENBERG: We have many fixed training scenarios, maybe operational scenarios, that will try to go to a point where you are going to go
to battle. You may have forced marches but the commanders, if they know what they are doing, will indulge a break at a certain time for personal leave or a water break because you want everyone to drink, to remain hydrated. We have not yet given doctrine to say that while you are drinking also break out and eat a small, 200-to-300 calorie food bar. We could obviously compose it to be high in carbohydrates, high fat, or mixed, but if we had some good thoughts in terms of how to design a realistic experimental situation and look at some important endpoints, we could come up with some answers as to whether it really makes a difference. We know it makes a difference if they drink and maybe change their socks, but we have not yet really convinced people to eat because it will deliver a benefit.
PATRICK DUNNE: I am a former athlete and I would suggest looking at a different athletic context. Most of your exercise physiology studies are done on single, individual athletes, so the whole issue of coupling performance to group decision making does not happen. But how about studies on team sports where you have 70 percent ![]() [maximal oxygen uptake] while playing basketball, and there is a lot of group decision making involved? Maybe individuals playing team sports are better models for military performance than the maxed-out marathoner. We must also look at the impact of the exercise on the person who has fatigue but whose catecholamine system is also turned on, which is a more complex interaction. I know it is easier to study one system at a time, but you are going to get your answers in the interactions.
[maximal oxygen uptake] while playing basketball, and there is a lot of group decision making involved? Maybe individuals playing team sports are better models for military performance than the maxed-out marathoner. We must also look at the impact of the exercise on the person who has fatigue but whose catecholamine system is also turned on, which is a more complex interaction. I know it is easier to study one system at a time, but you are going to get your answers in the interactions.
RICHARD WURTMAN: A test is now underway by the Boston Celtics. They are going to test what I guess Steve Ahlers was talking about. The evidence that when you run a marathon, the plasma choline level before the marathon and of course the original concern that it might impair neuromuscular transmission might diminish acetylcholine production in muscles, but if you are creating plasma choline, you are also affecting brain acetylcholine levels. The Boston Celtics have bezen testing it, and they allege that in the fourth quarter they do better.






























