
21
The Role of Carnitine in Enhancing Physical Performance
Peggy R.Borum1
INTRODUCTION
Human muscle contains a high concentration of carnitine. The first well-documented function of carnitine was found to be facilitation of the transport of long-chain fatty acids into the matrix of the mitochondrion, which is the site for beta-oxidation. Fatty acids are an important fuel for muscle metabolism. During endurance exercise, the oxidation of fatty acids spares the use of muscle glycogen and delays the onset of fatigue. Muscle tissue cannot synthesize carnitine and thus is dependent on the transport of carnitine from the bloodstream. Blood carnitine is derived from endogenous biosynthesis in the liver and kidney and from exogenous sources in the diet.
There is controversy as to whether or not administration of carnitine improves exercise performance. The difference in the intensity of exercise, the training or conditioning of the subjects, the quantity of carnitine administered, the route of administration, and the timing of administration relative to the exercise have led to different experimental results (Vecchiet et al., 1990).
FUNCTIONS OF CARNITINE
Carnitine Stimulates Fatty Acid Oxidation
The impermeability of all membranes to carbon chains activated to the coenzyme A (CoA) level, as diagramed in Figure 21–1 is well recognized. The carnitine transport system consists of carnitine and three proteins that result in the transport of the carbon chain moiety of acyl-CoA across a membrane and reesterified to another CoA molecule residing on the opposite side of the membrane (Borum, 1983). As diagramed in Figure 21–2, at the completion of the action of the carnitine transport system, the carbon chain is located on the opposite side of the membrane and is esterified to a different CoA molecule.
Long-chain fatty acids activated to the CoA level were the first acylcarnitines to be documented as being transported across a membrane, as diagramed in Figure 21–3. However, the acyl-CoA may be of any chain length and may be the metabolite derived from fatty acids, carbohydrates, or amino acids. The importance of carnitine in the oxidation of medium-chain fatty acids has been discussed recently (Borum, 1992).
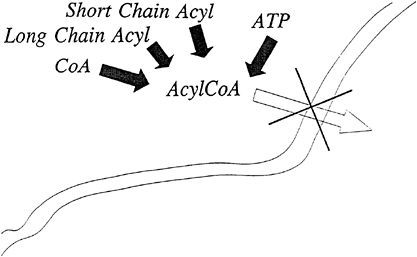
FIGURE 21–1 Acyl-coenzyme A (AcylCoA) moieties cannot penetrate membranes.
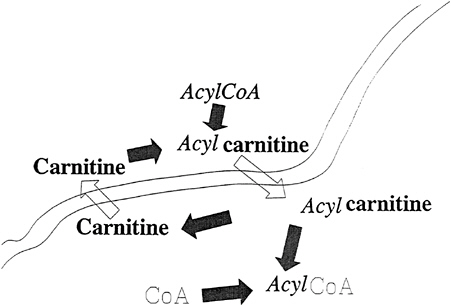
FIGURE 21–2 The carnitine transport system results in the acyl moiety of the acyl-coenzyme A (acylCoA) being transported to the opposite side of the membrane and esterified to a CoA moiety on the opposite side of the membrane.
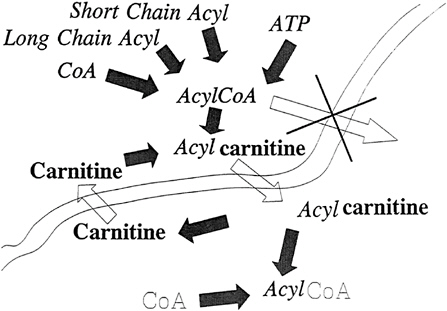
FIGURE 21–3 The carnitine transport systems function in the transport of both short-chain and long-chain acyl-coenzyme A (acylCoA) across membranes.
Carnitine Transports Acyl-CoA Across Membranes
It is clear that in beta-oxidation of long-chain fatty acids, carnitine is essential for the transport of long-chain acyl-CoA as long-chain acylcarnitine across the inner membrane of the mitochondrion. Carnitine is also essential in the transport of short-chain acyl-CoA as short-chain acylcarnitine across several membranes. During some physiological conditions, the goal of the transport process is to provide a needed substrate to the subcellular organelle where it is required. During other physiological conditions, the goal of the transport is to remove from a subcellular organelle a metabolite that is accumulating to toxic levels. In the latter condition, carnitine is functioning in a detoxification process. During exercise, carnitine is critical in providing needed substrates and in removing potentially toxic compounds.
Carnitine Prevents Accumulation of Lactate
During intense exercise, acetyl-CoA is produced at a faster rate than it can be used in the Krebs cycle. The rate-limiting factor in the Krebs cycle is frequently the availability of oxygen. During intense exercise, oxygen is limiting and the muscle produces lactate. The advantages of lactate production are that it permits the continuation of glycolysis with production of nicotinamide adenine dinucleotide, does not deplete limiting concentrations of CoA, and can be removed from the cell. The disadvantages of lactate production are that it can cause acidosis and it is a low-energy compound. An alternative pathway is for pyruvate to form acetyl-CoA, which in turn forms acetylcarnitine, which does not deplete limiting concentrations of CoA and which can be removed from the cell.
Carnitine Stimulates Utilization of Carbohydrate and Amino Acids
Since a common metabolite in the oxidation of fatty acids, amino acids, and glucose is acetyl-CoA, the role of carnitine in the transport of acetyl-CoA as acetylcarnitine across membranes places carnitine in the role of facilitating the oxidation of all three fuels used by the body.
Acylcarnitine Is a Storehouse of High Energy That Is Transportable
An important characteristic of acylcarnitine is that it is a high-energy compound, and thus, the acyl-CoA can be reformed without the use of an ATP molecule (Borum, 1986). Figure 21–4 illustrates the fact that the transport of acylcarnitine across a membrane delivers both the carbon chain and high energy (equivalent to an ATP molecule) to the new site. Thus, another advantage of the formation of acetylcarnitine rather than lactate by muscle is that high metabolic energy is preserved.
Acylcarnitine Permits Administration of High Energy
In addition to the supplementation of free carnitine, it is also possible to use acetylcarnitine and propionylcarnitine as supplements. Supplementation with the latter two compounds provides the carnitine moiety, the carbon chain moiety, and high metabolic energy. Additional research is needed to evaluate the potential benefit of supplementing individuals with a compound that potentially provides high metabolic energy.
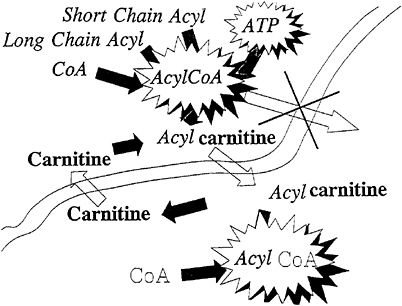
FIGURE 21–4 The carnitine transport system maintains the high energy level of the acylcoenzyme A (acylCoA), allowing the acyl-CoA to be esterified on the opposite side of the membrane without the use of an additional ATP molecule. The overall effect of the action of the carnitine transport system is the delivery of both the acyl group and high energy to the new site on the opposite side of the membrane.
ASSAY METHODS FOR CARNITINE AND ACYLCARNITINE
The most widely used assay for carnitine and acylcarnitine is the radioenzymatic assay (Borum, 1990). The assay uses radiolabeled acetyl-coenzyme A (acetyl-CoA) and an unknown amount of carnitine as substrates for the enzyme carnitine acetyltransferase and separates the radiolabeled acetylcarnitine product from the unreacted radiolabeled substrate. When alkaline hydrolysis is used to remove all acyl groups, the assay provides an excellent means of measuring total carnitine. The assay cannot be used to identify specific acylcarnitines. Although it is frequently used in combination with acid precipitation techniques to estimate long-chain acylcarnitines and short-chain acylcarnitines, investigators have found that caution must be used unless the measurement of the different fractions of acylcarnitine and free carnitine are documented to equal the total amount of carnitine measured. Specific acylcarnitines can be identified by gas chromatography-mass spectrometry techniques. The published methods require research-grade instruments and procedures that are not practical for measuring acylcarnitine levels in a large number of samples. However, ongoing work in several laboratories should permit measurement of specific acylcarnitines with bench-top instruments. These analytical techniques should greatly enhance investigators’ ability to address many of the issues discussed below.
POTENTIAL USE OF CARNITINE TO ENHANCE PHYSICAL PERFORMANCE
Intensity of Exercise Affects Carnitine Metabolism
High- and low-intensity types of exercise require two qualitatively distinct states of skeletal muscle metabolism. The transition between these two metabolic states for different exercise intensities occurs at a work load approximated by the lactate threshold, which is defined as the exercise intensity at which elevated plasma lactate concentrations are first observed (Hiatt et al., 1989).
Muscle acetyl-CoA and acetylcarnitine contents increase dramatically at the onset of intense exercise. When the duration of exercise is prolonged, acetyl-CoA and acetylcarnitine contents decrease in different proportions but are maintained above resting levels at exhaustion when muscle glycogen levels are depleted. As fatigue approaches, the carbohydrate-derived acetyl-CoA content decreases at a rate more rapid than the decrease in acetylcarnitine (Spriet et al., 1992b).
Muscle carnitine metabolism does not change in a graded fashion with exercise, but only when exercise is of sufficient intensity to qualitatively alter muscle substrate metabolism. Six normal male subjects exercised on a bicycle ergometer on two separate occasions with a constant work load. Low-intensity exercise was performed for 60 min at a work load equal to 50 percent of the lactate threshold, and high-intensity exercise was performed for 30 min at a work load between the lactate threshold and the maximal work capacity for the individual (Hiatt et al, 1989).
The data in Table 21–1 show that low-intensity exercise for 60 min has no detectable effect on muscle carnitine composition, but high-intensity exercise for 10 min decreases the free carnitine concentration and dramatically increases the short-chain acylcarnitine concentration of muscle. There were only minor changes in these muscle parameters between 10 and 30 min of intense exercise. After 60 min of recovery from high-intensity exercise, the total muscle carnitine content returned to baseline values. The short-chain acylcarnitine content remained elevated and the free carnitine content was only 46 percent of the content in the resting state, reflecting a continued redistribution of the carnitine pool. The finding of a persistent change in muscle carnitine metabolism into recovery is consistent with the observation that in humans, metabolic rate and substrate utilization remain altered for
TABLE 21–1 Changes in Muscle Acylcarnitine Levels During Exercise (percentage of value at rest)
|
Carnitine Fraction |
60 min of Low-Intensity Exercise |
10 min of High-Intensity Exercise |
|
Free |
100 |
34 |
|
Short chain |
100 |
550 |
|
Long chain |
100 |
100 |
|
Total |
100 |
81 |
|
SOURCE: Adapted from Hiatt et al. (1989). |
||
several hours after exercise and may have implications for the performance of repetitive bouts of high-intensity exercise (Hiatt et al., 1989).
The effect of exercise on plasma carnitine metabolism differed from that observed in muscle. With low-intensity exercise there were no changes in the profile of plasma carnitine with the exception of a small increase in plasma long-chain acylcarnitine. The plasma free carnitine level was increased at 15 min of high-intensity exercise but was unchanged from the values at rest
during the remainder of the exercise and the recovery period. The plasma short-chain acylcarnitine concentration increased from that at rest during exercise and remained elevated 10 min into recovery before returning to baseline levels at 30 min of recovery. Compared with the large changes in muscle after 10 min of exercise, the ratio of short-chain acylcarnitine to total acid-soluble carnitine in plasma increased modestly and only after 30 min of exercise. In contrast to muscle, the ratio of short-chain acylcarnitine to total acid-soluble carnitine in plasma had normalized by 30 min of recovery (Hiatt et al., 1989).
Muscle Training Affects Nutrient Utilization During Exercise
Training of muscle may result in muscle adaptations for the metabolism of fatty acids during prolonged exercise. Six trained and six untrained human males exercised for 3 h at 60 percent of their maximal dynamic knee extension capacity. Arterial plasma free fatty acid concentrations increased over time in both groups. Fractional uptake of free fatty acids across the thigh remained unchanged over time in the trained group (15 percent) but decreased in the untrained group. Thus, the total free fatty acid oxidation was higher in the muscles of the trained group. Glucose uptake increased in both groups over time and was significantly higher in the untrained group during the last hour of exercise. Although the contribution of extracellular substrates to thigh oxidative metabolism increased during the third hour of exercise in both groups of subjects, only in the thighs of the trained group was there an increased utilization of the plasma free fatty acids (Turcotte et al., 1992).
Carnitine Affects Pyruvate Metabolism
Pyruvate oxidation was measured in intact mitochondria isolated from fresh human skeletal muscle obtained from the excision of pectoralis minor in the course of a mastectomy for breast cancer; the muscle was free of macroscopic neoplastic infiltration, however. The data listed in Table 21–2 show that pyruvate oxidation increased significantly in the presence of L-carnitine and that inhibitors of the transport of either pyruvate or carnitine across the intact membrane greatly decreased the oxidation of pyruvate (Uziel et al., 1988).
Pyruvate dehydrogenase complex activity (PDHC) was measured in mitochondria made permeable to cofactors by exposure to hypotonic medium. PDHC activity increased in the presence of L-carnitine as shown in Table 21–2. As expected, because of the use of leaky mitochondria, the addition of an
TABLE 21–2 Pyruvate Metabolism (Percentage of Control Value)
|
Addition |
Pyruvate Oxidation in Intact Mitochondria |
Pyruvate Dehydrogenase Complex Activity |
|
1 mM L-Carnitine |
176 |
182 |
|
Inhibitor of pyruvate transport |
6 |
109 |
|
Inhibitor of pyruvate transport +1 mM L-carnitine |
5 |
ND |
|
Inhibitor of carnitine transport |
88 |
100 |
|
Inhibitor of carnitine transport |
95 |
164 |
|
1mM D-Carnitine |
ND |
100 |
|
1 mM Deoxycarnitine |
ND |
103 |
|
1 mM Choline |
ND |
100 |
|
NOTE: ND, not determined. SOURCE: Adapted from Uziel et al. (1988). |
||
inhibitor of either pyruvate transport or carnitine transport across membranes had no effect on activity. D-Carnitine, deoxycarnitine, and choline, which are structural analogs of L-carnitine, had no effect on activity. When L-carnitine was omitted, only 0.2 percent of the pyruvate oxidized was transformed to acetylcarnitine. When 1 mM carnitine was added, PDHC activity nearly doubled and 82 percent of the pyruvate oxidized was transformed to acetylcarnitine. PDHC stimulation occurred only at pyruvate concentrations greater than 0.25 mM, suggesting that carnitine and carnitine acetyltransferase can buffer the acetyl-CoA excess in human mitochondria when pyruvate is oxidized at high rates (Uziel et al., 1988).
Ten moderately trained male subjects aged 20–30 years performed a maximal exercise test on a bicycle ergometer, beginning with a light warm-up load followed by progressive incremental increases of 50 W every 3 min. The tests were stopped when one of the following was reached: (1) achievement of theoretical maximal heart rate, (2) muscular exhaustion, or (3) onset of severe dyspnea. Three days later the test was repeated after a randomized loading of either placebo or L-carnitine (2 g) (Siliprandi et al., 1990). Ninety minutes after carnitine administration and with the subjects at rest, plasma free carnitine levels increased, and short-chain carnitine levels did not change. In both the placebo and carnitine trials, exercise decreased plasma free carnitine
levels and increased plasma short-chain carnitine levels. Plasma long-chain carnitine levels did not change in any instance (Siliprandi et al., 1990).
Maximal exercise increased plasma lactate levels 15-fold and pyruvate levels 3.5-fold in the placebo trial. In each subject the increases in both lactate and pyruvate levels with maximal exercise were significantly lower after carnitine administration, in spite of the significantly greater amount of work carried out by 7 of the 10 subjects. In the carnitine trial the increase in lactate levels was lower throughout the trial, and the return to baseline levels during the 12-min recovery period was the same in both the placebo and carnitine trials. There was an inverse correlation of lactate and acetylcarnitine in plasma (Siliprandi et al., 1990).
Both free carnitine levels and short-chain carnitine levels increased in urine after carnitine administration (especially free carnitine). Exercise did not induce appreciable changes. When individual acylcarnitine levels were estimated by isotope-exchange high-pressure liquid chromatography in the placebo trial, exercise induced a significant decrease in acetylcarnitine levels and a concomitant increase in the amount of a four-carbon (C4) compound (the authors suggested that it was isobutyrylcarnitine). After carnitine administration but with no exercise, there was a decrease in the amount of the C4 compound. Exercise after carnitine loading showed an increase in acetylcarnitine levels and an almost disappearance of the C4 compound. It is important to note that all these data are percentages of the total short-chain acylcarnitine and that the absolute amount of the short-chain acylcarnitine was much greater in the carnitine trial than in the placebo trial (Siliprandi et al., 1990).
The authors interpreted the increase in plasma and urinary short-chain acylcarnitine levels with carnitine administration as reflecting a washout of short-chain acyls that accumulate in tissues as the CoA ester during exercise. They suggested that in the placebo trial, pyruvate dehydrogenase was inhibited, which caused an energy crisis, and that branched-chain amino acids were used for energy, which gave rise to the C4 compound. They suggest that, with carnitine supplementation, the branched-chain amino acids were not needed because pyruvate dehydrogenase was not so inhibited (Siliprandi et al., 1990). The authors did not address the issue of why there is a greater need for oxidation of branched-chain amino acids at rest with no carnitine supplementation than at maximal exercise with carnitine supplementation. The identification of the C4 compound as isobutyrylcarnitine should only be considered tentative and should be further investigated.
The possible effect of supplying free fatty acids during intense exercise on carnitine metabolism and the use of carbohydrate versus fat have been investigated. Rats were randomly assigned to a no-fat or a high-fat treatment and were perfused (single pass) by using one of three conditions: (1) 10 min at rest, (2) 10 min at rest and 1 min of stimulation, and (3) 10 min at rest and
5 min of stimulation. The right hind limb was used to obtain control samples of the soleus (89 percent slow oxidative fibers and 11 percent fast oxidative-glycolytic fibers) and red gastrocnemius from the deep portion of the gastrocnemius medial head (59 percent fast oxidative-glycolytic fibers, 35 percent slow oxidative fibers; and 9 percent fast glycolytic fibers). The perfusion media contained no fat in the fat-free group and approximately 1 mM oleate in the high-fat group (Spriet et al., 1992a). It should be noted that the fatty acid profile of the perfusion medium was nonphysiological.
Oxygen uptake was similar in the fat-free and high-fat treatments at rest and during stimulation. Muscle acetylcarnitine plotted versus acetyl-CoA levels for both treatments and all conditions gave a positive linear relationship (r= 0.82 for red gastrocnemius and (r=0.70 for soleus) (Spriet et al., 1992a).
Provision of high levels of free fatty acids while at rest increased acetyl-CoA and acetylcarnitine contents, despite an unchanged oxygen uptake, suggesting that little regulation of free fatty acid metabolism exists in skeletal muscle while at rest. Altered free fatty acid provision did not affect the increases in acetyl-CoA and acetylcarnitine during intense stimulation, suggesting that carbohydrate-derived acetyl-CoA dominates during exercise of this intensity (Spriet et al., 1992a).
Carnitine Supplementation May Increase Use of Fatty Acids During Exercise
Ten highly conditioned subjects performed a control test of 45 min of cycling at 66 percent of maximal oxygen uptake ![]() ; this was followed by 60 min of recovery in a sitting position. Each subject repeated this trial after 28 days of placebo and L-Carnitine treatment of 2 g/day (double blinded crossover design). There were no differences between the control test and the placebo test throughout the exercise and recovery periods (Gorostiaga et al., 1989).
; this was followed by 60 min of recovery in a sitting position. Each subject repeated this trial after 28 days of placebo and L-Carnitine treatment of 2 g/day (double blinded crossover design). There were no differences between the control test and the placebo test throughout the exercise and recovery periods (Gorostiaga et al., 1989).
The respiratory quotient was lower (P>0.05) in carnitine-treated subjects than in placebo or control subjects during exercise. The oxygen uptake, heart rate, blood glycerol concentrations, and resting plasma fatty acid concentrations listed in Table 21–3 showed a trend toward higher values in the carnitine-treated period but was not significant at the 0.05 level (Gorostiaga et al., 1989). These observations suggest increased lipid utilization by muscle during exercise with carnitine treatment, but the data are far from dramatic.
TABLE 21–3 Exercise After Placebo or Carnitine Supplementation in the Same Subjects
Carnitine Supplementation May Preserve Available Coenzyme A Pool
It can be calculated that during perfusion of the rat hind limb at rest, 97.7 percent (soleus) to 99.1 percent (red gastrocnemius) of the extra acetyl-CoA produced by adding free fatty acid to the perfusion medium was buffered by the formation of acetylcarnitine. Total acetylcarnitine accumulations during perfusion at rest were 5- to 2.5-fold greater than the resting CoA contents of red gastrocnemius and soleus, respectively. If there was no buffering of acetylcarnitine, the entire mitochondrial CoA store would be acetylated in 9 s in the red gastrocnemius and in 20 s in the soleus. After peak accumulation of acetylcarnitine during muscle stimulation, approximately 20 and 41 percent of the total carnitine pool would be acetylated in the soleus and red gastrocnemius, respectively (Spriet et al., 1992a).
In an experiment involving skiing uphill and downhill in difficult terrain at race pace for more than 13 h, the authors (Decombaz et al., 1992) calculated the estimated fuel cost of the exercise to be 38 MJ or 9,100 kcal. The authors assumed the following: 353 g of carbohydrate intake, 850 g of glycogen from muscles (25 kg, 3 percent glycogen), 100 g of liver glycogen, and 10 percent of energy from protein. Therefore 350 g of fat must have been oxidized. If 90
percent of the fat (1,100 nmol of free fatty acid) was oxidized in the muscle, the pool of muscle carnitine must have been acylated 13-fold during the race (1,100 mmol of free fatty acid×25 kg of muscle×3.33 mmol of total carnitine/kg of muscle) with no net loss of carnitine (Decombaz et al., 1992).
Eight subjects exercised for 3–4 min on a bicycle ergometer at work loads corresponding to 30, 60, and 90 percent of their ![]() , The graded exercise in that investigation was very important, and investigations that use different percent
, The graded exercise in that investigation was very important, and investigations that use different percent ![]() may not be comparable. Muscle biopsy specimens were taken at rest, at the end of the work period, and after 10 min of recovery. During the incremental exercise test, there was an increase in muscle lactate levels, an increase in the levels of the active form of the pyruvate dehydrogenase complex, and an increase in both acetyl-CoA and acetylcarnitine levels, with a corresponding decrease in CoA and free carnitine levels. At work loads of 60 and 90 percent of
may not be comparable. Muscle biopsy specimens were taken at rest, at the end of the work period, and after 10 min of recovery. During the incremental exercise test, there was an increase in muscle lactate levels, an increase in the levels of the active form of the pyruvate dehydrogenase complex, and an increase in both acetyl-CoA and acetylcarnitine levels, with a corresponding decrease in CoA and free carnitine levels. At work loads of 60 and 90 percent of ![]() , the accumulation of lactate showed that the rate of pyruvate formation from glucose was higher than the catalytic rate of the pyruvate dehydrogenase complex. The accumulation of acetyl-CoA and acetylcarnitine showed that the rate of condensation of acetyl groups with oxaloacetate was less than their rate of formation (Constantin-Teodosiu et al., 1991).
, the accumulation of lactate showed that the rate of pyruvate formation from glucose was higher than the catalytic rate of the pyruvate dehydrogenase complex. The accumulation of acetyl-CoA and acetylcarnitine showed that the rate of condensation of acetyl groups with oxaloacetate was less than their rate of formation (Constantin-Teodosiu et al., 1991).
The authors calculated that if the pyruvate dehydrogenase complex is fully active during exercise, the following calculations would apply to muscle (Constantin-Teodosiu et al., 1991):
-
Maximum rate of pyruvate degradation during exercise=20–25 µmol/s/kg of muscle,
-
CoA concentration at rest=9 µmol/kg of muscle,
-
L-carnitine concentration at rest=3,600 µmol/kg of muscle, and
-
the entire store of CoA in muscle could theoretically be acetylated within less than 1 s.
The high concentration of carnitine and the high levels of activity of carnitine acetyltransferase and carnitine acetylcarnitine translocase in muscle tissues enable carnitine to act as a buffer for excess acetyl-CoA production opposing the depletion of CoA. Depletion of CoA would result in the inhibition of the pyruvate dehydrogenase complex and the inhibition of oxoglutarate dehydrogenase in the citric acid cycle. It should be remembered that since acetylcarnitine is a high-energy compound, the acetyl units stored as acetylcarnitine during heavy exercise would be available if the exercise intensity decreased. Also, the acetyl unit would not require metabolic energy to re-form acetyl-CoA.
Carnitine Supplementation May Increase Work Output
Ten healthy males (moderately trained, but the authors [Vecchiet et al., 1990] did not give) ![]() ) performed three tests to maximal exercise intensity separated by 72 h. The intensity of the exercise was increased by 50 W every 3 min. The first test was performed without any treatment. The other two tests began 90 min after administration of a single oral dose of carnitine (2 g) or placebo in random order. Tests were terminated after muscular exhaustion or upon reaching the theoretical maximal heart rate (Vecchiet et al., 1990).
) performed three tests to maximal exercise intensity separated by 72 h. The intensity of the exercise was increased by 50 W every 3 min. The first test was performed without any treatment. The other two tests began 90 min after administration of a single oral dose of carnitine (2 g) or placebo in random order. Tests were terminated after muscular exhaustion or upon reaching the theoretical maximal heart rate (Vecchiet et al., 1990).
As exercise intensity increased, both ![]() and the blood lactate level increased. After carnitine treatment, all subjects tolerated similar exercise intensities but with a significant reduction of
and the blood lactate level increased. After carnitine treatment, all subjects tolerated similar exercise intensities but with a significant reduction of ![]() and the blood lactate level. When the subjects were required to exercise to their maximal capacity, 9 of the 10 subjects were able to do substantially more work in the session in which they received carnitine. Both the work output and the
and the blood lactate level. When the subjects were required to exercise to their maximal capacity, 9 of the 10 subjects were able to do substantially more work in the session in which they received carnitine. Both the work output and the ![]() were significantly increased by carnitine pretreatment (Vecchiet et al., 1990). Other investigators have not found L-carnitine supplementation to modify either the physiological parameters or the circulating metabolites (Oyono-Enguelle et al., 1988).
were significantly increased by carnitine pretreatment (Vecchiet et al., 1990). Other investigators have not found L-carnitine supplementation to modify either the physiological parameters or the circulating metabolites (Oyono-Enguelle et al., 1988).
Mixed-type latissimus dorsi of the dog was used to show the effect of carnitine on an in situ fatigue test. L-Carnitine appeared to improve the force of this muscle by 34 percent while the muscle was stimulated in situ. Ten mongrel dogs were used on different days with at least a 1-week interval. The musculus latissimus dorsi was trained by continuous stimulation with a frequency that was increased every 2 weeks until a frequency of 60 contractions per min was reached. An 8-min test was performed by using a pacemaker as the control test. The test was repeated after a rest period of 30 min, during which time the drug to be investigated was infused. The authors (Dubelaar et al., 1991) did not address the issue of whether the experimental design would distinguish between an effect of carnitine on exercise and an effect on the recovery period between the two exercise sessions. Drugs (saline; 0.15 mmol of L-carnitine, D-carnitine, or choline per kg of body weight) were infused at a rate of 0.15 mmol/min, and only one drug was tested in each experiment (Dubelaar et al., 1991).
When two control tests were repeated at a 30-min interval during which saline was infused, no significant change in force occurred. A third test after another 30-min rest period showed the same pattern. Administration of L-carnitine resulted in a significant increase in force throughout the second test, resulting in a significantly higher amount of work carried out by the muscle. The effect of L-carnitine on the contractile force was less pronounced in animals in which the latissimus dorsi was conditioned for 12 weeks. During
this training period, the percentage of type I muscle fibers increased from 23 ±3 to 69±16 percent (Dubelaar et al., 1991).
After intravenous administration of carnitine, total plasma carnitine levels rose from 23±0.6 to 322±61 nmol/ml, but the concentration in the muscle remained the same. The structural analog choline did not improve the contraction force of the muscle. D-Carnitine decreased the contraction force significantly during the last minutes of the exercise. In the presence of high insulin levels, which is known to inhibit fatty acid oxidation, the insulin itself had no effect on contraction force, but the stimulation of the force by carnitine was no longer seen in the presence of insulin and glucose (Dubelaar et al., 1991).
Exercise May Alter Metabolic Compartmentation of Carnitine and Acylcarnitine
Eighteen cross-country skiers took part in a race in the Alps that lasted an average of 13 h and 26 min. The skiers’ heart rates indicated performance at 72 percent of ![]() . The reduced oxygen pressure at the high altitude added a hypoxic stress. Carnitine intake 2 weeks before the race was 50±4 mg/kg. The total carnitine concentration of resting muscle (vastus lateralis) was measured twice, with a 2-year interval for eight of the subjects. Muscle carnitine concentrations did not change with time and did not correlate with carnitine intake, but they showed consistent interindividual differences (Decombaz et al., 1992).
. The reduced oxygen pressure at the high altitude added a hypoxic stress. Carnitine intake 2 weeks before the race was 50±4 mg/kg. The total carnitine concentration of resting muscle (vastus lateralis) was measured twice, with a 2-year interval for eight of the subjects. Muscle carnitine concentrations did not change with time and did not correlate with carnitine intake, but they showed consistent interindividual differences (Decombaz et al., 1992).
After exercise, the total muscle carnitine concentration was unaltered, but the free carnitine concentration decreased 20 percent and the short-chain carnitine concentration increased 108 percent. The correlation between the drop of one and the rise of the other was weak (r=0.23). This suggests that, in addition to acylation in situ, there may be significant interorgan exchange in the form of an efflux of free carnitine from the muscle and an influx of acetylcarnitine from the liver (where it may be raised after exercise) to the muscle. A significant finding is the stability of total carnitine levels in muscle after long and demanding periods of exercise. Therefore, the persistently elevated urinary carnitine excretion with exercise would not be at the expense of muscle carnitine. The ratio of short-chain acylcarnitine to total soluble carnitine rose from 19 to 37 percent. There was no correlation between the total carnitine concentration in muscle at rest and finishing time. There was a lack of correlation between muscle carnitine and maximal aerobic power or duration of training. This lack of correlation indicates that endurance conditioning has little effect on skeletal muscle carnitine concentrations in humans (Decombaz et al., 1992).
Seven moderately trained male subjects ages, 19–31 years; ![]() , 3.3 to 4.3 liters/min participated in a study of prolonged (120 min) moderate-intensity exercise. The first submaximal bicycle ergometer exercise session lasted 120 min at approximately 50 percent of the individual
, 3.3 to 4.3 liters/min participated in a study of prolonged (120 min) moderate-intensity exercise. The first submaximal bicycle ergometer exercise session lasted 120 min at approximately 50 percent of the individual ![]() . After 1–2 months, a second exercise session immediately preceded by 5 days of oral L-carnitine (inner salt, 1 g 5 times per day) and 1 g of L-carnitine on the morning of the test was conducted. It should be noted that the dose was much higher than the amount of carnitine that would be expected to be obtained in the diet (Soop et al., 1988).
. After 1–2 months, a second exercise session immediately preceded by 5 days of oral L-carnitine (inner salt, 1 g 5 times per day) and 1 g of L-carnitine on the morning of the test was conducted. It should be noted that the dose was much higher than the amount of carnitine that would be expected to be obtained in the diet (Soop et al., 1988).
Blood samples were collected from the femoral arterial catheter and one of the femoral venous catheters at rest, after 40 and 120 min of exercise, and at 40 min postexercise. A continuous intravenous infusion of [14C]oleic acids bound to human albumin (1.6 µCi/min) was given during the 30 min preexercise and during the last 20 min of exercise (Soop et al., 1988). The experimental design appeared to assume that the metabolism of oleic acid is indicative of the metabolism of all fatty acids. The experiment should be repeated with a labeled physiological mixture of fatty acids.
Carnitine supplementation did not influence the relation between arterial oleic acid concentration and leg uptake. During exercise there was a progressive increase in total and acylated plasma carnitine concentrations and a decrease in the concentrations of plasma free carnitine. At 40 min postexercise, there was a decline in total and acylated carnitine levels in comparison with those at 120 min, but the levels at both times were still higher than those preexercise. The ratio of acylated carnitine to free carnitine increased from preexercise values during the exercise and remained elevated at 40 min postexercise (Soop et al., 1988).
The following data concerning the release and uptake of carnitine by muscle was collected during the exercise session with no carnitine supplementation:
Free carnitine:
-
Rest—no evidence of net uptake or release;
-
40 min of exercise—release almost significant;
-
120 min of exercise—significant release;
-
40 min postexercise—no net exchange.
Acylated carnitine
-
Rest—no net exchange;
-
40 min of exercise—no net exchange;
-
120 min of exercise—tendency toward uptake; and
-
40 min postexercise—significant release.
The exercise-induced changes in plasma carnitine concentrations after carnitine supplementation followed the same pattern that was observed before carnitine supplementation, except that the free carnitine concentration rose in response to exercise (Soop et al., 1988).
Urinary carnitine excretion did not differ before or during exercise with no carnitine supplementation. More than 10 times as much carnitine was excreted during exercise with carnitine supplementation, and a larger percentage was found to be free carnitine (Soop et al., 1988).
As a result of the exercise-induced release of free carnitine and a simultaneous tendency toward uptake of acylated carnitine, no net exchange of total carnitine was observed during exercise. The release of free carnitine from the muscle occurred concomitantly with a decrease in the plasma free carnitine level. The level of plasma acylated carnitine rose, in spite of no evidence of a release by the leg muscle. Thus, the observed fall in plasma free carnitine levels and the rise in acylated carnitine levels in response to exercise is difficult to explain solely in terms of alterations in muscle carnitine exchange. Results suggest that during exercise, free carnitine is released from muscle and acylated at a site other than contracting muscle. The most likely site for carnitine acylation is the liver. The carnitine released from muscle during exercise is either redistributed to tissues other than muscle or accumulated in plasma, since urinary carnitine excretion was not affected by exercise (Soop et al., 1988).
Since several studies have shown no effect of carnitine supplementation on the total muscle carnitine concentration during exercise but have shown a beneficial effect of the carnitine supplementation on exercise performance, some authors speculate that there is an “extramyocytal” effect of carnitine. “The question arises whether there is a compartment in muscle other than the striated muscle cells, from which carnitine can be lost during muscle performance. The question could be rephrased, Do endothelial and/or vascular smooth muscle cells lose carnitine during muscle stimulation?” (Dubelaar et al., 1991:E192). These authors have previously shown that in cultured endothelial cells, the bulk of ATP may come from carnitine-dependent fatty acid oxidation. It is possible that the cells of the vascular wall lose carnitine because of the relative ischemia during muscle stimulation. The lesser effect of carnitine in the trained muscle is in line with this hypothesis, since the number of capillaries is known to increase by training, leading to better oxygenation and possibly preventing functional decompensation of the relatively vulnerable vascular cells (Dubelaar et al., 1991).
DIETARY INTAKE VERSUS PHARMACOLOGICAL ADMINISTRATION
Unpublished data show that most Americans consume between 50 and 100 mg of carnitine per day in their diets. Some individuals consume up to 300 mg of dietary carnitine per day. Most of the studies evaluating the effect of carnitine supplementation on exercise performance have used quantities that are severalfold higher than the typical dietary carnitine intakes. It is important to remember that the data available on the intestinal absorption and metabolic compartmentation of 100 mg of dietary carnitine may not be applicable to supplementation with pharmacological doses of carnitine. Carnitine supplementation in investigations concerning exercise have used free carnitine. Acetylcarnitine and propionylcarnitine have been used in Europe in clinical trials involving a variety of patients. The uptake of these acylcarnitine supplements by some organs may be greater than the uptake of free carnitine. Future studies should consider their use as supplements.
CONCLUSIONS AND RECOMMENDATIONS
It is clear that carnitine has an important metabolic role in the exercising muscle. However, the specific functions of carnitine and acylcarnitine and the effect of supplementation remain to be elucidated. The design of future investigations should carefully address the following issues:
-
Assay methodology—The radioenzymatic assay is appropriate for total carnitine measurements, but it should be used with caution to measure fractions of acylcarnitines. Bench-top gas chromatography-mass spectrometry methodologies should be further developed to measure specific acylcarnitines in a large number of samples. Caution should be used in the use of a carnitine concentration value as an indicator of the carnitine status in another cell type. Plasma, red blood cells, white blood cells, urine, and muscle should all be evaluated in each investigation.
-
Chemical form of supplementation—Published investigations concerning exercise have all used free carnitine as the chemical form that is used in supplementation. Acetylcarnitine and propionylcarnitine should be evaluated for their use in supplementation.
-
Timing of supplementation—The time relationship of the dose of carnitine supplementation to the time of exercise has varied from investigation to investigation and may be the cause of some of the variability of the results. The time relationship of dose and exercise needs to be carefully evaluated.
-
Dose of supplementation—In studies with carnitine supplementation, higher doses than are found in most diets have been used. The optimal dose has not be defined.
-
Physical training of subjects—Physical training alters muscle metabolism, including the metabolism of carnitine. All investigations should use subjects who have the same level of training as the individuals for whom the data will be utilized.
-
Intensity of exercise—The intensity of exercise alters muscle metabolism, including the metabolism of carnitine. All investigations should use the intensity of exercise for which the data will be utilized.
-
Animal studies—Some experimental questions require techniques that are too invasive to be performed in humans and thus require animals. Although rodents have been used for most experiments in the past, they are not the best animal model because their muscle metabolism is significantly different from that of humans. Swine would be a better choice.
-
Practical requirements—Carnitine is sufficiently stable to withstand long-term storage. It does have a bitter aftertaste, but it can be masked with citrus flavor. Carnitine has been found to be safe under most circumstances. However, more is not necessarily better, and investigations of patients with different pathophysiologies have indicated that better results can sometimes be obtained with lower doses.
REFERENCES
Borum, P.R. 1983 Carnitine. Ann. Rev. Nutr. 3:233–259.
1986 Carnitine function. Pp. 16–27 in Clinical Aspects of Human Carnitine Deficiency, P.R. Borum, ed. New York: Pergamon Press.
1990 Carnitine: Determination of total carnitine using a radioenzymatic assay. J. Nutr. Biochem. 1:111–114.
1992 Medium chain triglycerides in formula for preterm neonates: Implications for hepatic and extrahepatic metabolism. J. Pediatr. 120:S139–S145.
Constantin-Teodosiu, D., J.I.Carlin, G.Cederblad, R.C.Harris, and E.Hultman 1991 Acetyl group accumulation and pyruvate dehydrogenase activity in human muscle during incremental exercise. Acta Physiol. Scand. 143:367–372.
Decombaz, J., B.Gmuender, G.Sierro, and P.Cerretelli 1992 Muscle carnitine after strenuous endurance exercise. J. Appl. Physiol. 72:423–427.
Dubelaar, M.L., C.M.H.B.Lucas, and W.C.Hülsmann 1991 Acute effect of L-carnitine on skeletal muscle force tests in dogs. Am. J. Physiol. Endocrinol. Metab. 260:E189–E193.
Gorostiaga, E.M., C.A.Maurer, and J.P.Eclache 1989 Decrease in respiratory quotient during exercise following L-carnitine supplementation. Int. J. Sports Med. 10:169–174.
Hiatt, W.R., J.G.Regensteiner, E.E.Wolfel, L.Ruff, and E.P.Brass 1989 Carnitine and acylcarnitine metabolism during exercise in humans. Dependence on skeletal muscle metabolic state. J. Clin. Invest. 84:1167–1173.
Oyono-Enguelle, S., H.Freund, C.Ott, M.Gartner, A.Heitz, J.Marbach, F.Maccari, A.Frey, H.Bigot, and A.C.Bach 1988 Prolonged submaximal exercise and L-carnitine in humans. Eur. J. Appl. Physiol. 58:53–61.
Siliprandi, N., F.Di Lisa, G.Pieralisi, P.Ripari, F.Maccari, R.Menabo, M.A.Giamberardino, and L.Vecchiet 1990 Metabolic changes induced by maximal exercise in human subjects following L-carnitine administration. Biochim. Biophys. Acta 1034:17–21.
Soop, M., O.Bjorkman, G.Cederblad, L.Hagenfeldt, and J.Wahren 1988 Influence of carnitine supplementation on muscle substrate and carnitine metabolism during exercise. J. Appl. Physiol. 64:2394–2399.
Spriet, L.L., D.J.Dyck, G.Cederblad, and E.Hultman 1992a Effects of fat availability on acetyl-CoA and acetylcarnitine metabolism in rat skeletal muscle. Am. J. Physiol. Cell Physiol. 263:C653–C659.
Spriet, L.L., D.A.MacLean, D.J.Dyck, E.Hultman, G.Cederblad, and T.E.Graham 1992b Caffeine ingestion and muscle metabolism during prolonged exercise in humans. Am. J. Physiol. Endocrinol. Metab. 262:E891–E898.
Turcotte, L.P., E.A.Richter, and B.Kiens 1992 Increased plasma FFA uptake and oxidation during prolonged exercise in trained vs. untrained humans. Am. J. Physiol. Endocrinol. Metab. 262:E791–E799.
Uziel, G., B.Garavaglia, and S.Di Donato 1988 Carnitine stimulation of pyruvate dehydrogenase complex (PDHC) in isolated human skeletal muscle mitochondria. Muscle Nerve 11:720–724.
Vecchiet, L., F.Di Lisa, G.Pieralisi, P.Ripari, R.Menabò, M.A.Giamberardino, and N.Siliprandi 1990 Influence of L-carnitine administration on maximal physical exercise. Eur. J. Appl. Physiol. 61:486–490.
DISCUSSION
GILERT LEVEILLE: Two questions. One, when we use acetyl or propionyl carnitine as an oral supplement, do we know if it is absorbed in that form?
PEGGY BORUM: That is being looked at. There are actually clinical trials that are ongoing with acetyl and propionyl, but I do not think that the data are in yet. There is some indication that what is happening is that the acetylcarnitine that is in the gastrointestinal tract is not the same molecule that gets into the bloodstream. Acetate may be removed from acetylcarnitine in one location of the cell and acetylcarnitine resynthesized in another location using a different acetate molecule.




















