
6
NEW TECHNOLOGIES
The discussion that follows highlights several aspects of new technologies. It is not intended to be inclusive; rather, it illustrates some promising lines of investigation. To illustrate the potential difficulty of introducing a marker into clinical practice, we describe the use of prostate-specific antigen (PSA) in some detail. We also consider fully the importance of alpha2u-globulin, a controversial topic involving the extrapolation of animal data to human conditions.
MAKERS OF CELL INJURY, REGENERATION, AND HYPERTROPHY
To a considerable extent, the identification of markers has been facilitated by an understanding of the molecular biology of the cell and the mechanisms that are responsible for both orderly growth and neoplastic transformation.
The Cell Cycle
Nongrowing, quiescent cells have low rates of DNA synthesis and are said to be in the G0 phase of the cell cycle. These cells (Figure 6-1) can be stimulated to grow by the addition of one of several growth factors. After the binding of a growth factor to its receptor, growth-promoting signals are generated that commit the cell to a new phase of the cell cycle, G1, where biosynthetic activities resume. During the transition to the G1 phase, the cells express many genes that enable them to progress through this phase. Prominent among them are the so-called immediate early genes. The S phase starts when DNA synthesis starts and ends when the DNA content of the nucleus has doubled. Thereafter, the cells move into the G2 phase of the cycle, where new protein and RNA synthesis occurs and the cells increase in size. The M phase begins with nuclear division and ends with cytoplasmic division (Safirstein, 1994). Various points in the cycle are influenced by environmental stress and can
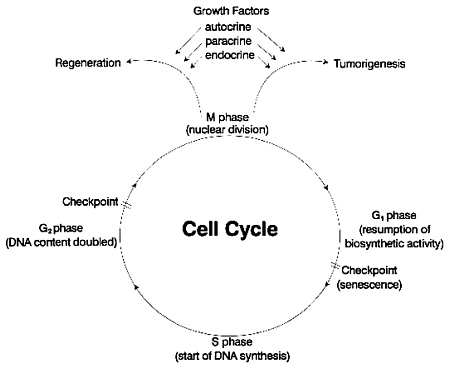
Figure 6-1 The cell cycle.
be targets for future biologic-marker research. The control of growth and differentiation involves sophisticated modes of cellular communication. Figure 6-2 illustrates actual and potential modes of growth control, as might be found in the prostate. Endocrine control is exemplified by the effects of androgen, which stimulates the production of growth factors in both epithelial and stromal cells. Fibroblasts secrete factors that bind to epithelial-cell receptors.
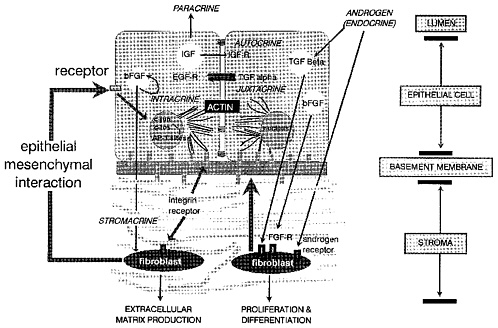
This represents a special case of paracrine growth control, "stromacrine" control. External signals, with specific growth peptide and cell-surface receptors producing tissue specificity, link with common intracellular signal pathways, which ultimately induce the growth genes c-fos and c-jun and transcription factors. Fibroblasts are responsible for controlling proliferation and differentiation, as well as for synthesizing extracellular matrix. Fibroblasts can also control epithelial-cell growth and differentiation through their effect on the basement membrane. The proteoglycan composition of basement membrane reflects whether it is growth-promoting or differentiation-promoting. If the membrane and matrix connect with the cell, differentiation generally results; thus, cell-adhesion molecules—such as integrins, fibronectin, and proteoglycans—that bind to cell receptors or matrix proteins are important inducers of differentiation. Juxtacrine growth in the epithelium occurs when a membrane-bound form of the growth peptide binds to a receptor on an adjacent cell. Juxtacrine growth control in the epithelium tends to favor differentiation. Autocrine control occurs when cells secrete growth peptides that induce their own growth by binding to their own receptors. Intracrine growth, which has so far been observed in model systems, occurs when the growth factors are prevented from being exported by mutations that inhibit cleavage of the pre-pro forms of the peptides.
Growth Factors
Renal growth is initiated by mechanisms that include changes in renal growth factors. Several of the growth factors have been found in kidney tissue itself or shown to act on renal cells in culture. The growth factors isolated from kidney tissue include epidermal growth factor (EGF), insulin-like growth factors (IGF), transforming growth factors (TGF), and platelet-derived growth factor (PDGF). Other substances, such as angiotensin II, have also been shown to have substantial growth-regulating properties.
Epidermal Growth Factor
EGF is a potent mitogen capable of stimulating the proliferation of cultured renal tubular cells (Goodyer et al., 1988; Norman et al., 1987). It is produced in the kidney (Olsen et al., 1984; Salido et al., 1986, 1989; Scott et al., 1985) with a precursor molecule, prepro-EGF (Bell et al., 1986; Fisher et al., 1989; Rall et al., 1985). Binding sites for EGF have been found in various portions of the renal tubules and the glomeruli. After injury to distal tubular cells, there might be an autocrine pathway which regulates synthesis, promotes receptor binding, and so regulates repair. Stimulation of proximal tubule replication most likely involves EGF released from other cells and the use of a paracrine pathway. EGF also stimulates the production of
IGF-I and acts directly to enhance IGF-I gene expression (Rogers et al., 1991).
In the case of renal ischemic injury, prepro-EGF mRNA synthesis or processing changes (Safirstein et al., 1989). There is a prolonged reduction of EGF excretion, a reduction in prepro-EGF mRNA, an increase in EGF-receptor density (Safirstein et al., 1990), and an increase in the binding of EGF in the postischemic kidneys. Indeed, exogenous EGF administered to rats after renal ischemic injury enhances renal tubular cell regeneration and accelerates the recovery of renal function (Humes et al., 1989; Tsau et al., 1989).
The synthesis or processing of prepro-EGF mRNA is also altered by exposure to nephrotoxic agents, such as cisplatin (Safirstein et al., 1989). In addition, an increase of EGF receptors has been seen during the renal hyperplasia that follows folic acid administration (Behrens et al., 1989) and ischemia (Saferstein et al., 1989).
Insulin-Like Growth Factors
The insulin gene family is made up of IGF-I, IGF-II, proinsulin, and relaxin. The IGF-I gene is on chromosome 12, and the insulin and IGF-II genes are on the short arm of chromosome 11. The insulin-like growth factors are bound to carrier proteins. IGF-I and IGF-II interact with a distinct cell-surface receptor. IGF-I is synthesized in most organs, where it is involved in the regulation of various metabolic and growth processes (Hammerman, 1989). IGF-I can be found in the cortical medullary collecting duct and the parts of the thin loop of Henle that are in the medulla (Andersson et al., 1988; D'Ercole et al., 1984). IGF-I mRNA has been isolated directly from the kidney; this implies that the IGF-I recovered from the kidney is synthesized there (Bortz et al., 1988). IGF-I receptors are present on glomerular mesangial cells and on proximal tubular cells (Hammerman and Rogers, 1987). Interaction with IGF-I can result in the stimulation of protein kinase activity.
In rats, IGF-I has been implicated in the compensatory hypertrophy of the kidney that follows unilateral nephrectomy, in the hypertrophy that accompanies the feeding of high-protein diets, and, more important, in regenerating tubular cells after ischemic injury. In compensatory hypertrophy, IGF-I gene expression is stimulated, and the renal concentration of IGF-I is substantially increased (El Nahas et al., 1989; Fagin and Melmed, 1987; Polychronakos et al., 1985; Stiles et al., 1985). When high-protein diets are fed, serum IGF-I increases (Hirschberg and Kopple, 1991; Isley et al., 1983; Maiter et al., 1988; Prewitt et al., 1982); this is circumstantial evidence of a causal role of IGF-I in hypertrophy. IGF-I immunoreactivity is transiently expressed by regenerating cells in the postischemic rat kidney. IGF-I immunoreactivity appears as early as 3 days after the injury. With increas-
ing differentiation of the regenerating cells, IGF-I immunoreactivity vanishes; at 14 days, it is not detectable (Andersson and Jennische, 1988). The regenerative cells express IGF-I peptide and IGF-I mRNA in a transient manner that is thought to correlate with cell differentiation better than with cell division (Matejka and Jennische, 1992).
Transforming Growth Factor Beta
Transforming growth factor beta (TGF-B) constitutes a family of ubiquitous growth peptides that are intimately involved in extracellular matrix formation, cellular proliferation and differentiation, wound-healing, cartilage and bone formation, and possibly oncogenesis. Evidence is growing that TGF-B operates in an autocrine or paracrine fashion to bring about a multiplicity of cellular actions.
In the kidney, TGF-B has been implicated in the repair processes that follow renal ischemic injury, (Humes and Daniel, 1990); it might be an important mediator in the genesis of renal fibrosis after the administration of anti-glomerular-basement-membrane IgG (Phan et al., 1990); it has been implicated in the pathogenesis of glomerulonephritis (Okuda et al., 1990) and it might contribute to the development of progressive kidney fibrosis (Yamamoto et al., 1994).
There is evidence that TGF-alpha expression is greater in prostatic epithelial cells of patients with benign prostatic hypertrophy (BPH) than in epithelial cells of normal persons (Harper et al., 1993). Extracts of human prostate with benign hypertrophy contain fibroblasts that have been shown to secrete TGF-B (Story et al., 1993). TGF-B has more effect on stromal growth than on epithelial growth (Sherwood et al., 1992), inasmuch as TGF-B concentrations are higher in stroma than in epithelium (Truong et al., 1993). The TGF family might have an important role in the pathology of BPH. For example, bFGF is known to stimulate fibroblasts and can increase expression of intracellular matrix proteins in other cell lines. aFGF is known to negate the inhibitory effects of TGFnl in prostatic cell lines and is also androgen-responsive.
Another member of the FGF family, Int-2, has a sequence very similar to that of bFGF and in transgenic mice has been found to be associated with BPH. The BPH in those mice was dissimilar to human BPH in that the mice did not have accompanying stromal and fibromuscular hyperplasia; so it was difficult to study these model systems. The prostates from the transgenic mice did not grow when transplanted into nude mice; this indicates that alterations in hormonal and growth-factor concentrations might play a role in the pathogenesis of BPH. However, in transgenic mice, growth factors do not elicit histologic changes in the prostate.
Angiotensin II
Several lines of evidence indicate that
angiotensin II (All) acts as a regulator of cell growth. For example, it has been shown that angiotensin-converting enzyme inhibitors attenuate renal hypertrophy (Anderson et al., 1986b). In cultures of rabbit proximal tubular cells, All by itself has no direct effect on protein synthesis; but in the presence of EGF (Norman et al., 1987) or PDGF (Norman et al., 1987; Wolf and Neilson, 1991), it can enhance tubular epithelial proliferation. Although All does not induce mitogenesis, it appears to be capable of promoting cellular hypertrophy and early oncogene expression (Rozengurt and Heppel, 1975). With EGF and PDGF, All can induce the expression of various cellular oncogenes (Wolf and Neilson, 1990)—an effect that can be blocked with a competitive antagonist of All. Indeed, All has many of the features of growth factors and appears to be a growth regulator in the kidney. All binds to specific cell-surface receptors on tubular cells and activates many of the intracellular signaling pathways associated with cell growth (Norman, 1991). For example, the All-induced hypertrophic response might be related to its ability to stimulate Na+ /H+ pump activity in proximal tubular cells.
Early-Response Genes
In cellular proliferation and hypertrophy, an orderly pattern of gene expression (Cowley et al., 1989) leads to the induction of mRNA and various protein products. The c-fos and c-myc genes are examples of so-called ''immediate early'' genes and encode DNA-binding proteins localized to the nucleus. The initial activity of the c-fos protein appears to be pivotal to the transcriptional activity of the cell and is necessary for the transition from the stable resting state of the cell cycle to the gap between the mitotic and S phases where DNA replication occurs. The protein product encoded by the c-myc gene serves a regulatory function in controlling DNA synthesis by binding DNA polymerase II and is necessary for the next transition, to the synthetic phase of the cell cycle. Later, other gene products appear with similar kinetic characteristics.
The pattern of expression of these gene products after stimuli that provoke either renal hypertrophy or hyperplasia has been examined. In studies of the compensatory renal hypertrophy induced by uninephrectomy (Beer et al., 1987; Norman et al., 1988), modest increases in c-myc, c-H-ras, and c-K-ras expression have been reported. Others have been able to detect both c-fos and c-myc transcripts within 15 minutes after uninephrectomy (Sawczuk et al., 1990) and Egr-1 expression within 30 minutes (Ouellette et al., 1990). In rats with acute unilateral ureteral obstruction, within minutes there is a transient induction of c-myc transcripts and a somewhat more prolonged expression of c-fos. Although both decline by 24 hours, there is evidence of reinduction at 48 hours (Sawczuk et al., 1989). After nephrotoxic injury by a large parenteral dose of folic acid, expression of c-fos, c-myc, c-K1-ras, and c-Ha-ras is markedly increased (Asselin and Mar-
cus, 1989; Cowley et al., 1989). After renal ischemic injury by bilateral renal arterial occlusion of 50 minutes (Safirstein, 1990), c-fos mRNA increases, reaches a peak at I hour, and declines rapidly to control values by 4 hours. A similar pattern has been observed for Egr-1 mRNA. During the repair phase of ischemic acute renal failure, c-myc expression in the renal cortex increases by a factor of 10 (Humes and Daniel, 1990).
Nucleic Acid Synthesis and Hypertrophy
Although attention has been given to the cell cycle and DNA synthesis, other mechanisms are operative when cells hypertrophy. The hallmark of compensatory growth is a marked increase in cellular RNA content (Hayslett, 1979). For example, after unilateral nephrectomy in the rat, RNA and protein synthesis in the remaining kidney increases within 12–24 hours (Bucher and Malt, 1971) and persists for weeks as renal mass increases (Halliburton and Thomson, 1965). The increase in RNA synthesis is a useful index of the extent of hypertrophy. Less impressive alterations in DNA synthesis have been demonstrated within 6 hours after uninephrectomy (Toback and Lowenstein, 1974). This hyperplastic response reaches a peak at 2 days (Threfall et al., 1967) but is not of the same magnitude as the hypertrophic response. At two weeks, in contrast, total RNA content rises by 40% and total DNA content by only 25%; this shows that the predominant factor in compensatory growth is hypertrophy (Threfall et al., 1967).
During compensatory growth, the processing of nucleoplasmic RNA is accelerated (Willems et al., 1969), and more ribosomal RNA (rRNA) is produced (Ash and Cuppage, 1970; Halliburton and Thomson, 1965; Kurnick and Lindsay, 1968; Toback and Lowenstein, 1974). In fact, because rRNA constitutes more than 85% of total RNA in the kidney, the characteristic growth-induced increase in the renal RNA-to-DNA ratio results from an increase in ribosome number. The principal site of all these activities is the renal cortex. Indeed, there is direct evidence that the accretion of RNA occurs in the tubular cells of the cortex (Vancura et al., 1970).
Sodium-Pump Activity and Cell Growth
On the basis of studies performed in cell cultures, it has come to be recognized that a rapid increase in sodium entry into the cell stimulates Na+ /K+ pump activity (Rozengurt and Heppel, 1975; Smith and Rozengurt, 1978) and increases intracellular K+ (Tupper et al., 1977; Wolf and Neilson, 1990). The increases are associated with a commencement of DNA synthesis (Lopez-Rivas et al., 1982). Initially, an influx of Na+ exceeds the rate at which Na+ is extruded across the basolateral mem-
brane by Na+ /K+ pump activity with a reduction of intracellular pH (Schuldiner and Rozengurt, 1982). In proximal tubular cell cultures, the increase in Na+ /H+ pump activity associated with the addition of hypertonic NaCl results in increases in cell volume and protein content without stimulation of DNA synthesis (Rozengurt, 1981); this effect suggests that cell hypertrophy is coupled with Na+ influx (Fine et al., 1985). With a reduction in renal mass, growth stimulation and increase in Na+ /H+ pump activity are closely related (Fine, 1986).
It also appears that Na+ influx is necessary for the initiation of DNA synthesis stimulated by peptide growth factors. For example, mitogenic factors stimulate Na+ influx (Rozengurt et al., 1981; Smith and Rozengurt, 1978) and H+ efflux: (Schuldiner and Rozengurt, 1982). The addition of NaCl to cultures of kidney epithelial cells results in an increase in the number of cells initiating DNA synthesis in response to EGF and insulin (Toback, 1980). Furthermore, a decrease in the rate of influx of Na' achieved by a reduction of Na+ in the culture medium prevents the development of the mitogenic response to EGF and insulin (Burns and Rozengurt, 1984). In contrast, an increase in cellular Na+ accumulation can be attenuated in the presence of a purified growth-inhibiting protein. The reversal by the addition of NaCl to the medium suggests that variation in the Na+ flux during the onset of kidney epithelial cell growth can be related to the action of a specific cell protein that inhibits DNA synthesis (Toback, 1980).
Polyamines
The polyarnines spermidine and spermine and their precursor diamine putrescine, omnipresent components of all living cells, are aliphatic polycations with three, four, and two positive charges, respectively, at physiologic pH. In the cell, they are bound to macromolecular anionic sites in nucleic acids, ribosomes, and membranes. Polyamines are believed to be essential for cellular growth, proliferation, and differentiation, although their physiologic function at the molecular level is still not well understood (Manteuffel-Cymborowska, 1993).
CANCER OF THE BLADDER
Cell Culture
In vitro models of toxic and pathologic processes of the urinary tract, besides the kidneys, have been recently developed. Because most of them have focused on growth, development, and neoplastic transformation, they have used cell cultures derived from either bladder or ureter epithelium, rather than freshly isolated cells, as an in vitro system. Reznikoff and colleagues (1983) cultured normal human uroepithelial cells from tissue explants that comprised ureteral transitional epithelial
cells. Although fetal bovine serum (7% by volume) was included in the culture medium, hormone and growth factor supplements were also included to optimize growth and differentiation. Culture of uroepithelial cells from experimental animals actually lagged behind culture of cells from humans. Johnson et al. (1985) developed a successful primary culture and later a serial cultivation process for bladder epithelial cells from normal rats.
Acute cytotoxicity of bladder carcinogens—such as biphenyls, nitrofurans, and 3-methylcholanthrene—has been examined in cultured normal human uroepithelial cells (Reznikoff et al., 1986). Reduction in cell number during culture was used as the index of toxicity. The prevalence of urinary carcinogenesis has stimulated interest in uroepithelial-cell culture as a model for the study of growth regulation and neoplastic transformation, and most published studies in which these cell cultures have been used focused on processes related to tumorigenesis and growth regulation. The studies have included investigation of effects of polyamine synthesis inhibition on growth (Messing et al., 1988), of effects of bacterial endotoxicants on cell survival and growth (Wille et al., 1992), of the role of activation of the EJ/ras oncogene in neoplastic transformation (Pratt et al., 1992), of chromosomal deletions induced by bladder carcinogens in SV40-immortalized human uroepithelial cells (Meisner et al., 1988; Reznikoff et al., 1988; Wu et al., 1991), and of second-messenger mechanisms and the carcinogenic process in SV40-immortalized human uroepithelial cells (Jacob et al, 1991).
Quantitative Fluorescence Image Analysis
Quantitative fluorescence image analysis (QFIA) is an exciting recent technique for marker research (West, 1970; West et al., 1987). It is based on the recognition of a single cancer cell in a mass of normal cells through the quantitation of a fluorescent signal to obtain a measurement of the DNA, protein, or other macromolecular content of individual cells. Aneuploidy (the presence of more or fewer than the normal diploid number of chromosomes) is apparently the result of genetic instability and is a hallmark of carcinogenesis (McGowan et al., 1988). The use of QFIA permits more precise quantitation of DNA content in individual cells. Unlike flow cytometry, QFIA selects cells on the basis of composition or structure; unlike immunohistochemistry, it is capable of true quantitation (Parry and Hemstreet, 1988; Rao et al., 1993).
That cancer cells often have aneuploidy has been known for decades, and the technique of flow cytometry was developed originally to determine ploidy. The difficulty with using overall ploidy as a marker is that normal cells yield some measurement variability in the amount of DNA, and usually some cells are in the process of dividing and hence will have higher DNA content than normal. Consequently, it is very difficult to detect abnormal individual
cells in the vast sea of normal cells (Parry and Hemstreet, 1988).
Image analysis has been used to detect cancer cells for over a decade. The main advantage of the approach is that it provides objective, reproducible results. Image analysis is well suited to so-called rare-event analysis, the detection of objects that make up 1% or less of the cell sample.
In its initial application for bladder-cancer detection, QFIA urinary cytology successfully combined the attributes of classical cytopathology with those of DNA cytometry and thereby increased sensitivity to low-grade lesions (Bass et al., 1987; Bonner et al., 1993; Hemstreet et al., 1990; Hemstreet et al., 1983). The semiautomated QFIA uses both quantitative and qualitative analyses of urinary-tract specimens and is a new, highly sensitive alternative for early cancer detection. QFIA technology has an advantage over flow cytometry in requiring fewer cells for valuations and in being able to analyze individual cells for a one-cell diagnosis of cancer rather than requiring patterns of cells. Yet the two techniques can be complementary, and together they have greater sensitivity than classical cytopathologic methods in detecting cellular transformation.
QFIA technology has also been used in several large-scale occupational studies of people at risk for bladder cancer and has shown a specificity of 93–96% (Hemstreet et al., 1988). It is being increasingly recognized as a valuable tool for screening asymptomatic people to identify biologic risks associated with working in environments that contain toxic and possibly carcinogenic substances (Bi et al., 1993). QFIA has been applied extensively in both clinical detection of bladder cancer and several trials in worker cohorts exposed to bladder carcinogens (Bi et al., 1993; Hemstreet et al., 1988).
In addition, QFIA has been used successfully to monitor for the efficacy of treatment with chemopreventive retinoids in an animal model (Hemstreet et al., 1992; Hurst et al., 1991). The numbers and DNA content of bladder-wash cells obtained from rats treated with carcinogens and chemopreventive retinoids was found to be an accurate predictor of risk of developing bladder cancers (Hemstreet et al., 1992; Hurst et al., 1991).
There are newer directions of QFIA research to learn whether commonly seen atypical cells are the result of an inflammatory or irritative process or are truly progressing toward malignancy. Fluorescence probes for cytoskeletal oncogenes and growth factor receptors in addition to DNA and RNA are being incorporated (Rao et al., 1990, 1991). The ability to image multiple probes on a single cell greatly increases its clinical usefulness. The new capabilities are being added to the experimental armamentarium for determining and possibly reducing cancer risk or reversing malignant cell transformation.
The ability to study macromolecular changes in single living cells with fluorescent probes offers a tool that exceeds the capabilities of the electron microscope and the scanning electron micro-
scope. The combination of QFIA technology with the use of monoclonal antibodies and DNA hybridization promises imaginative applications for early cancer screening and diagnosis.
Basic research with QFIA will continue to investigate the quantitation of cell-surface antigens, oncogenes, and cytoskeletal components (Jones et al., 1990; Rao et al., 1993). The findings can be correlated with nuclear DNA content and classical histopathology. The objectives will be to develop methods for understanding the fundamental oncogenic process at the single-cell level, to define biochemical markers, to identify preneoplastic conditions, and to differentiate tumors with metastatic potential (Rao et al., 1990, 1993).
Given the recognition that aneuploidy is a marker of genetic instability and is only indirectly related to changes in gene expression or deletion, QFIA was extended from DNA to protein markers, which might be more directly related to genetic changes than is aneuploidy. In addition, other studies have shown that using QFIA with Gactin (a precursor molecule of the cytoskeleton) yields an early marker of carcinogenesis that can guide chemoprevention studies and, in conjunction with tumor markers of aneuploidy and the presence of cells that react with antibodies against tumor-specific antigens, can be helpful in the diagnosis and management of bladder cancer in worker cohorts exposed to bladder carcinogens (Rao et al., 1990; Hemstreet al., 1992).
More important, those techniques might help to define individuals at risk for developing cancer and to identify and monitor for the effects of chemopreventive agents.
DIFFERENTIAL-DISPLAY POLYMERASE CHAIN REACTION
Differential-display polymerase chain reaction (DD-PCR), as first presented by Liang and Pardee (1992) and improved by Pardee (Liang et al., 1993) and Bauer and colleagues (1993) has facilitated the investigation of altered gene expression in a wide range of applications. It might well become the method of choice for the elucidation of the molecular mechanisms underlying the cellular response to xenobiotics. The responses that occur on xenobiotic challenge involve an increase ("up-regulation") or a decrease ("down-regulation") in the synthesis of structure-and differentiation-related proteins. Metabolism can be changed, various growth and signal peptides and their receptors can be altered, and the growth kinetics of the cell can be affected.
Individual sections of base pairs on DNA are necessary to encode a particular protein. Identification of the expression of these genes can use conventional techniques, such as Northern and Southern blots, but requires knowledge of the nature of the expressed genes and the availability of previously characterized or identified probes. Most mammalian cells express around 15,000 genes. Testing for each individually is a daunting task. In the cell nucleus, polymerase enzymes copy a sequence of
DNA by transcription to messenger RNA (mRNA). In the cell cytoplasm, proteins are ultimately synthesized on the basis of the information encoded in the mRNA. In theory, electrophoresis of the total RNA can separate the various mRNAs. In practice, some of the genes are expressed at very low levels, and their resolution is poor.
The general strategy of DD-PCR is to amplify partial DNA sequences copied from subsets of mRNA (cDNA) by reverse transcriptase enzymes and PCR (Bauer et al., 1993; Liang and Pardee, 1992; Liang et al. 1993). It is necessary somehow to divide all the cDNA sequences so that the ones of interest can be identified. To achieve this partitioning, a series of different primers is used. These primers initiate the reading of the DNA sequence. At the 3'end, which contains the poly(A) tail characteristic of mRNA, 12 different 3'primers are used; each one displays a different population of mRNAs. Further division can be obtained by the use of different upstream primers. A set of arbitrary primers has been shown to yield optimal results (Bauer et al., 1993; Liang et al., 1993). These delimit PCR at a specific length for each of the mRNAs. Each pair of 3' and upstream primers reproducibly anneals to the same sequences in each successive PCR cycle and selects a discrete set of mRNAs. A radioactive or fluorescent label is incorporated into the mixture to label the cDNAs and make them detectable. Differentially expressed genes are evident from differences in intensity of corresponding bands in the two gels. The sensitivity of the technique can be adjusted to any selected level of difference, thereby allowing those genes that are differentially expressed at the highest level to be detected first, then the genes at smaller levels. For example, genes with a difference in expression of a factor of 10 or higher could be identified first, then those with a difference of a factor of 5-10, and so forth. The cDNAs can be isolated, further amplified, directly cloned, or used as probes to isolate longer sequences.
DD-PCR has been widely applied to many fundamental problems in cancer research and other fields in which identification of changes in gene expression is important. The technique is likely to have a large impact on understanding of the underlying mechanisms of toxicity to the genitourinary tract and on the development of markers with which to investigate risk in human populations.
CANCER OF THE PROSTATE
Prostate-Specific Antigen
Carcinoma of the prostate is the second most common cause of cancer deaths in men. Prostate-specific antigen (PSA), a glycoprotein that is produced by the epithelial cells of the prostate and acts as a protease to liquefy the seminal coagulum (Wang et al., 1979), has been used in screening for prostatic cancer, for its staging, and as a marker of response to treatment. Although its utility has been demonstrated, controversy has developed about its proper
use. It is appropriate to consider the questions about PSA in a discussion of new technologies and the development of other markers, which are likely to be associated with similar concerns when applied to clinical decision-making.
Major issues to be addressed are the effectiveness of current screening programs to detect occult prostatic cancer and the effectiveness of surgical intervention compared with conservative management. Only an estimated 13% of men will benefit from current prostatic-cancer detection programs. Studies have compared prostatic cancers found in radical cystoprostatectomy specimens (latent form) with prostatic-cancers detected in current prostatic-cancer screening programs. In the screening programs, prostate cancer was identified with PSA and digital rectal examination (DRE), and confirmed with ultrasonographically-guided needle biopsy of the prostate. The studies were based on the observation that prognosis is unequivocally related to the size of the tumor (McNeal et al., 1986), the stage of the disease, and the grade of the tumor. Tumors were organized into a hierarchy of risk for cancer death on the basis of division into 10 categories. Some 300 patients underwent whole-mount sections of the prostate with careful attention to the level of invasion and grade. At one end of the spectrum were patients with positive surgical margins, seminal-vesicle invasion, or lymphnode invasion (incurable). Only 4% of patients who came to radical prostatectomy were in the small-cancer or latent-cancer group, 35% were in the incurable group, and the rest were in the intermediate groups, whose cancers were potentially effectively controlled.
As longitudinal screening programs identify increased numbers of patients with organ-confined disease, patient survival will improve (Catalona et al., 1991, 1993, 1994). However, because of an increased number of multiple biopsies in these patients, a higher percentage of latent cancers will be detected. In some studies, only 5% of the patients with negative results of ultrasonography and DRE were in the group considered to have biologically inactive disease. That finding is supported by the observation that 1 g or 1 cm3 of prostatic tissue raises serum PSA by 3.5 ng/mL. Furthermore, there is no relationship between reported tumor volume and PSA of less than 0.4 ng/mL. When tumor size, grade, and stage were compared with those in autopsy groups, 65% of the autopsy cases were in the very-low-risk group. These studies illustrate several important points. First, PSA is probably not detecting a large number of cases of latent prostatic cancer. Second, a high percentage of patients are being detected when they are in the high-risk group and it is too late for cancer control. Third, the identification of new biologic markers is necessary to decrease the length-time bias and to detect earlier cases that are likely to progress rapidly to advanced disease. With the current markers, it is likely that the introduction of screening programs will lead to a shift toward detecting cases in the treatable category, in that people will be screened more often. The identification of markers of prostatic cancer with greater specificity and
sensitivity might be helpful in epidemiologic studies designed to determine the relationship between xenobiotic substances and the appearance or progression of prostatic cancer and to determine whether subsets of environmental and nutrient factors that predispose to the aggressive form of prostatic cancer can be identified.
It is worth asking whether prostatic cancer is being detected and treated in many patients who will ultimately die of other causes. Some studies indicate that most patients who are found to have prostatic cancer on the basis of increased PSA are those who should be effectively treated with radical surgical intervention. However, screening and treatment of this group must take several factors into consideration, including the cost of the evaluation, the morbidity associated with the tests themselves, and the morbidity of those who will not be cured, in contrast with those with progressive disease.
What would it mean for everyone with prostatic cancer to be treated? About 1–2% of men die of prostatic cancer, and 10–12% have clinically manifest disease. If all the men over the age of 50 with prostatic cancer had their cancers removed (30% of males), there would be an estimated 75,000 deaths due to surgery (assuming a complication rate of 1%), in contrast with the 30,000 who would have died from prostatic cancer. These screening programs do not take into consideration the morbidity of such an experiment, which would result in impotence in 250,000 men and 20,000 deaths in the first year alone (Hinman, 1991). Some argue that the surgical mortality is as low as 0.1%, but objective data from controlled trials are not available, and estimates consider mortality only in the immediate perioperative period (Catalona, 1994).
Debate continues with regard to the appropriateness of screening with PSA and the efficacy of surgical intervention, versus watchful waiting. A number of large clinical trials are being conducted to address these issues. The results could have an impact on individual risk assessment. Methods to assess an individual's natural life expectancy in relation to the biologic activity of the tumor are equally important.
Markers are useful to identify persons at risk, to detect preclinical disease, and to categorize clinical disease. Any reduction in mortality and morbidity achieved by the use of new markers must be superior to current diagnostic techniques. This includes cost assessments. Data are now available for comparing routine prostatic-cancer screening that uses DRE. In a study of 6,600 men comparing screening with DRE and PSA, Catalona et al. (1994) reported that DRE alone would miss 40% of prostatic cancers, and PSA alone would miss 23% of prostatic cancers. When PSA was added to DRE, there was a 68% increase in the detection of organconfined prostatic cancers. The results confirm the complementary nature of the two tests.
In initial screening programs with PSA and DRE, 9% of the patients underwent biopsy evaluation (Catalona et al., 1993). Those who were negative were monitored every 6 months for 3 years. The longitudinal screening indi-
cated that 40% of men will develop prostatic cancer and that about 80% of men with PSA over 10 ng/mL will have proven prostatic cancer. Screening doubles the identification of patients with organ-confined prostatic cancer. The identification of patients with organconfined disease and a PSA between 4 and 10 ng/mL is controversial. Only 40% of patients with PSA between 4 and 10 ng/mL will have prostatic cancer (Benson et al., 1992a,b). Should we use PSA concentration or rate of its change as a guide to biopsy? Available data do not support a delay in biopsy except in patients with a small prostate, and the consensus is that the rate of change in PSA correlates poorly with prostatic cancer detection. The continued search for improved methods of interpreting PSA results attests to the limitation of this marker.
Marker Development
Several major advances in marker research would improve the outcome of prostatic cancer marker detection. The ideal marker would specifically detect potentially biologically active prostatic cancer in its premalignant stages. The identification of a marker with a low false-positive rate that would reduce the number of false-positive findings in patients with PSA concentrations between 4 and 10 ng/mL would reduce patient evaluation costs. The identification of a marker that would detect patients with biologically active tumors capable of rapid progression would identify the subgroup most in need of surgical intervention.
The frequent occurrence of multiple biopsies of the prostate in response to high false-positive PSA results provides ample tissue samples for marker evaluation. Prostatic intraepithelial neoplasia (PIN) and adenomatous hyperplasia are two premalignant lesions associated with prostatic tumorigenesis, and a grading scale has been developed for these lesions. The term PIN I refers to dysplastic changes, but aggressive clinical actions such as rebiopsy of the prostate, are not usually taken. In contrast, when PIN II or III is identified, the prostate is commonly rebiopsied; these lesions have been associated with increased PSA Which of the premalignant lesions will develop into prostatic cancer is of current interest, and such markers as oncogenes, growth factors, and cytoskeletal markers are being evaluated (see Table 4-5). An understanding of the pathologic condition is important for proper interpretation of pathologic results in the development of new markers.
Scientists are searching for new markers that will improve the sensitivity and specificity of the current PSA test. PSA in blood predominantly forms a complex with alpha1-antichymotrypsin. The PSA might be irreversibly inactivated in vitro by two protease inhibitors, alpha1-antichymotrypsin and alpha2u-globulin.
To improve the sensitivity of the PSA test, new assays have been developed to detect the different molecular forms of PSA in serum. One assay detects binding of PSA to proteinase inhibitors, a
second assay detects PSA in complex with alpha1-antichymotrypsin, and a third detects nonbound PSA. Figure 6-3 depicts the three assays. Continuing studies indicate that the specificity of the PSA test might be improved by quantitating the various forms of bound and nonbound PSA. The ratio of PSA bound to alpha1-antichymotrypsin is higher in prostatic cancer patients; nonbound PSA is more commonly observed in patients with benign prostatic hyperplasia.
The development of monoclonal antibodies for other markers more specific for prostatic cancer is in progress. One major problem in prostatic cancer staging is seminal-vesical invasion. An assay that could specifically detect seminal-vesical invasion would be a useful prognostic indicator.
Neuroendocrine Cells in the Prostate
There are highly specialized neuroendocrine epithelial cells in the prostate. They have endocrine and paracrine functions and contain various peptides, including serotonin, calcitonin, and thyroid-stimulating hormone. Increasing evidence suggests that these cells are important during prostatic growth and differentiation. Cells with similar functions are found in the gastrointestinal tract and the respiratory system and are of local endodermal origin. The endocrine and paracrine cells in the prostate are slender apical processes that extend into the lumen and put out dendritic projections between adjacent epithelial cells.
The peptides are present in semen and are detectable with radioimmunoassay or high-performance liquid chromatography. The dendritic processes identified with electron microscopy might exert a paracrine function on adjacent epithelial cells. Of primary importance is that both afferent and efferent neurons have been observed in association with neuroendocrine cells. The potential of these molecules either as growth regulators or as markers of disease remains to be determined (see Table 4-5).
CANCER OF THE KIDNEY
This section discusses examples in urinary toxicology of the examination and use of biologic markers to determine the choice of relevant test species and aid in the interpretation of the results of toxicity studies.
Renal Tumors in Male Rats Mediated by Alpha2u-globulin
A structurally diverse group of organic chemicals has been shown to cause a renal syndrome in male rats that is manifested acutely by the accumulation of marker protein in renal proximal-tubule cells visible under the light microscope as hyaline droplets (Hard et al., 1993; Health Effects Institute, 1985, 1988; USEPA, 1991a,b). Mechanistic studies with a few select
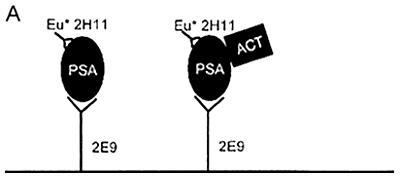
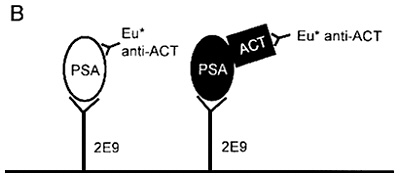
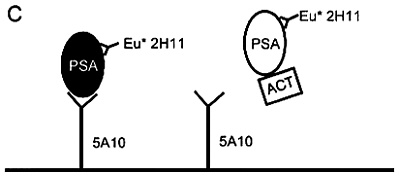
Figure 6-3 New assays developed to detect the different molecular forms of PSA in serum. Schematic representation of three sensitive two-site immunofluorometric procedures to measure PSA immunoreactivity in serum. (A) Assay T detects epitopes available on both the PSA molecule that did not form a complex and PSA that formed a complex with proteinase inhibitors such as alpha-antichymotrypsin; (B) assay C, using the same catching antibody as assay T, measures PSA that forms a complex with alpha 1-antichymotrypsin by a polyclonal tracing antibody against alpha 1-antichymotrypsin; and (C) assay F detects PSA that has not formed a complex, but has difficulty recognizing PSA that has formed a complex with proteinase inibitors such as alpha 1-antichymotrypsin. Source: Lilja, H., A.T.K. Cockett, and P-R. Abrahamsson. 1992. Reprinted with permission of Cancer.
chemicals have shown that the protein involved in the production of the hyaline droplets is alpha2u-globulin (Alden et al., 1984; Dietrich and Swenberg, 1991; Lehman-McKeeman and Caudill, 1992a; Lock et al., 1987), the major urinary protein excreted by adult male rats. Compared with the female rat and some other species, including humans, the male rat is physiologically proteinuric (Neuhaus and Lerseth, 1979; Olson et al., 1990; Shapiro and Sachchidananda, 1982; Vandoren et al., 1983). A large amount of the alpha2u-globulin is synthesized in the liver of the male rat under the control of several hormones, particularly androgen (Feigelson and Kurtz, 1977; Roy and Chatterjee, 1983; Roy and Neuhaus, 1967; Roy et al., 1983), but it has not been detected in the liver of normal female rats or in other species, including humans (MacInnes et al., 1986; Sippel et al., 1975). In addition to stimulating the synthesis of large amounts of alpha2u-globulin in the male rat, testosterone suppresses the degradation of alpha2u-globulin (Feigelson and Kurtz, 1977; MacInnes et al., 1986; Motwani et al., 1984; Roy and Chatterjee, 1983; Roy and Neuhaus, 1967; Roy et al., 1983; Sippel et al., 1975). The predominant hypothesis linking alpha2u-globulin to renal carcinogenesis is that with continued exposure to such chemicals as dlimonene and trimethylpentane, which bind to alpha2u-globulin and cause its accumulation, the acute protein overload progresses to renal-cell injury, which can be observed histologically as cell necrosis in the proximal tubule with the presence of granular casts in the outer medulla, chronic and progressive nephrosis, and papillary linear mineralization (Alden, 1986; Borghoff et al., 1990; Flamm and Lehman-McKeeman, 1991; Hard et al. 1993; Swenberg et al., 1989; USEPA, 1991b).
That rather well-defined progression of toxicity in male rats is referred to as alpha2u-globulin nephropathy (Alden, 1989; Borghoff et al., 1990; Swenberg et al., 1989). Ultimately, there is an increase in the incidence of tumors arising from the renal-tubule epithelial cells. It has been suggested that a chemical cannot induce alpha2u -globulin nephropathy without having the potential for inducing tumors at some dose (Dominick et al., 1991). Because the chemicals that produce this type of toxicity and carcinogenicity are nongenotoxic, it is suggested that the renal-tubule tumors seen after chronic exposure to alpha2u-globulin hyaline-droplet-inducing agents result from sustained target-organ toxicity that leads to increased renal-cell proliferation followed by promotion of spontaneously-initiated cells of increased opportunity for mutagenesis (Alden, 1989; Swenberg et al., 1989; Trump et al., 1984a); however, quantitative relationships between protein accumulation, renal disease, and sustained increases in cell proliferation are not understood (Melnick, 1992). An alternative hypothesis is that of a chemically mediated direct toxic effect on the kidney, where alpha2u-globulin influences the dose of toxic chemical that reaches the target organ (Melnick, 1992).
The histopathologic sequence of this form of toxicity is distinctive, and the differences in potency and species and
sex susceptibility suggest that these chemicals act by mechanisms different from classical renal carcinogens (Alden and Frith, 1991; Hard, 1987, 1990; Hard and Whysner, 1994; Lipsky and Trump, 1988). Nevertheless, renal-tu-bali tumors induced by alpha2u-globulin accumulators are structurally indistin-guishable from either spontaneous tumors or those induced by classical carcinogens (Hard, 1990; Lipsky and Trump, 1988; NTP, 1989, 1990). Although human renal-cell tumors are structurally similar to rodent tumors, the histopathology of the alpha2u-globulin-induced nephropathy in affected kidneys in males differs remarkably from that in human renal carcinoma (Bennington and Beckwith, 1975; Tannenbaum, 1971; USEPA, 1991).
The mechanistic data developed to understand the formation of male rat renal tumors mediated by alpha2u-globulin are reviewed below. As discussed elsewhere in greater detail (Melnick, 1992; USEPA, 1991b), gaps in data on specific mechanisms are considerable. For example, the crucial connections between cellular necrosis, increased cell proliferation, tubular hyperplasia, and renal-tubule tumors are not proved, and the possibility that similar proteins in humans lead to a common mechanism has not been completely ruled out. Chemical substances that induce renal-tubular tumors in male rats must be evaluated one by one. d-Limonene is used as an example of the accepted standard prototype alpha2u-globulin accumulator (Hard and Whysner, 1994).
The Environmental Protection Agency has reviewed the data on a number of chemicals that cause increases in renal-tubule tumors in male rats and might increase alpha2u-globulin hyaline-droplet nephropathy, and it has recommended criteria for the use of such information in risk assessment (USEPA, 1991b).
Acute Nephropathy
The renal effects of chemicals that induce alpha2u-globulin accumulation constitute a continuum of changes that seem to be initiated by the appearance of eosinophilic hyaline droplets in the proximal-tubule epithelial cells and that with chronic exposure progress to severe hyperplasia and tumorigenesis. The features of acute nephropathy include accumulation of hyaline droplets in the proximal tubules; necrosis and regeneration, particularly in the P2 segment of the proximal tubule; linear mineralization in the papilla; and granular cast formation, primarily at the junction of the inner and outer stripes of the outer medulla (Alden et al., 1984, Halder et al., 1984; Hard and Whysner, 1994; HEI, 1988; Swenberg et al., 1989).
Studies that have involved the administration of chemicals capable of inducing alpha2u-globulin accumulation in immature rats, aged rats, castrated male rats, male NCI Black Reiter (NBR) rats that do not synthesize alpha2u-globulin in the liver, male rats given estrogen, or female rats given exogenous estrogen and alpha2u-globulin show that
development of the nephropathy occurs only in the presence of alpha 2u-globulin (Chatterjee et al., 1989; Dietrich and Swenberg, 1990; Garg et al., 1988, 1989b; Hobson et al., 1986; Logothe-topoulos and Weinbren, 1955; Murty et al., 1988; Ridder et al., 1990; Roy and Neuhaus, 1967; Sippel et al., 1975),
A background of protein-droplet formation is seen in the male rat kidney (Goldsworthy et al., 1988a; Logothe-topoulos and Weinbren, 1955; Maunsbach, 1966a). The droplets consist of protein that is being degraded within the phagolysosomal compartment of proximal-tubule epithelial cells. It is estimated that male rat kidneys filter 40–50 mg of alpha2u-globulin each day and reabsorb about 25 mg of it into proximal-tubular cells (Lehman-McKeeman and Caudill, 1992b; Neuhauss et al., 1981). In contrast, there is no background incidence of hyaline droplets containing alpha2u-globulin in female rats, which synthesize less than one-hundredth as much alpha2u-globulin as male rats (Vandoren et al., 1983); this small amount of protein probably does not accumulate as histologically evident hyaline droplets. Thus, the physiologic processes underlying the spontaneous formation of hyaline droplets probably constitute a threshold phenomenon and result from the large quantities of alpha2u-globulin that the male rat reabsorbs each day.
Agents capable of inducing hyaline-droplet formation increase both the size and the number of droplets in the male rat kidney—a process that requires the interaction of either the parent chemical or a metabolite with alpha2u-globulin (Garg et al., 1989a; Short et al., 1987). The interaction is probably a rate-limiting step in the development of nephropathy. It has been shown that the binding of hyaline-droplet-inducing agents to alpha2u-globulin is reversible but highly specific, inasmuch as the dissociation constant for the ligand-alpha2u-globulin complexes is around 10-7 M (Borghoff et al., 1991; Lehman-McKeeman and Caudill, 1992b). It is the binding of a specific chemical to alpha2u-globulin that disrupts the normal handling of the protein in the kidney, particularly its lysosomal degradation, and the protein-chemical complex accumulates within phagolysosomes (Lehman-McKeeman et al., 1990a). For example, the dose-response curve for d-limonene-induced hyaline-droplet nephropathy includes doses below which no exacerbation can be demonstrated (USEPA, 1991b).
Chronic Renal-Cell Injury and Proliferation
In addition to the manifestations just described of acute toxicity associated with alpha2u-globulin nephropathy, chronic exposure to chemicals that cause the syndrome results in the development of more severe glomerular lesions, and prominent linear mineralization often occurs in the kidney (Alden, 1989; Alden and Frith, 1991; Bruner, 1984; Dominick et al., 1991; Trump et al., 1984b). The structural characteristics of the renal injury associated with alpha2u-globulin nephropathy have been
well described, but little has been learned about the mechanisms underlying the associated renal cell death (Melnick, 1992; USEPA, 1991). Nor has the intracellular concentration of alpha2u-globulin required to trigger cell death been determined. Whether a role exists for the nonbound chemical or metabolite and whether the accumulation of alpha2u-globulin is the actual cause of the renal toxicity have been questioned (Melnick, 1992). Renal injury can be initially identified as an impairment of proximal-tubule function with an increase in marker cellular casts or debris in the urine (Alden et al., 1984; Kanerva et al., 1987a; Short et al., 1987). In addition, a well-defined dose-response relationship indicates that a loss of renal function is observed only at dosages that cause alpha2u-globulin nephropathy. Renal toxicity in both acute and subchronic types of d-limonene-induced alpha2u-globulin nephropathy has been determined; it seems, in concordance with the hyaline-droplet response, that dosages of less than 5 mg/kg per day (for 91 days) do not after renal-cortical function, as assessed by transport of organic anions and cations (Webb et al., 1989). In direct contrast, dosages known to induce renal-tubule tumor formation—from 75 to 150 mg/kg per day—cause marked loss of renal-cortical function (Lehman-McKeeman et al., 1989).
The cytotoxicity and loss of renal function associated with alpha2u -globulin nephropathy cause a compensatory. increase in renal-cell proliferation. For chemicals evaluated by the National Toxicology Program (NTP) for subchronic and chronic toxicity, the histologic appearance of renal-cell hyperplasia is commonly reported. Additional studies that used more-specific methods for measuring cell proliferation showed that unleaded gasoline or 2,4,4-trimethylpentane (Short et al., 1987, 1989), d-limonene (Dietrich and Swenberg, 1991), and 1,4-dichlorobenzene (Charbonneau et al., 1989) increase cell proliferation, particularly in the P2 segment of the proximal tubule. The shape of the dose-response curve for renal-cell proliferation induced by alpha2u-globulin nephropathy indicates a nonlinear relationship. A quantitative dose-response relationship between protein accumulation, sustained cell proliferation, and tumorigenesis needs to be explored further.
Renal Neoplasia
Renal neoplasia occurs spontaneously in rats at a very low rate; tubule-cell adenoma and adenocarcinoma are not apparent until the age of 2 years and even then are observed in only 0.2 and 0.1%, respectively, of often studied strains of male rats and less than 0.1 and 0.1% of the female rats. If followed over the entire life span of the rat, those incidences increase to 0.2 and 0.4% of male rats and 0.4 and 0.6% of female rats (Solleveld et al., 1984). (See also National Toxicology Program (NTP) historical control data, e.g., USEPA, 1991c; NTP, 1990) Known genotoxic renal carcinogens, such as substituted
nitrosamines, do not exhibit acute hyaline-droplet nephropathy or sex or species specificity (Alden and Frith, 1991; Hard, 1987). Moreover, renal tubule-cell tumor incidence after exposure to a genotoxic renal carcinogen is considerably higher, often approaching 100%. In direct contrast, the increased incidence of renal tumors associated with an alpha2u-globulin hyaline-droplet inducer are generally low (the highest is 30%) even at maximum tolerated doses.
Genotoxicity
The mutagenicity of hyaline-dropletinducing agents (both parent chemical and some of the metabolites that bind to alpha2u-globulin) has been tested in a variety of assays, and these chemicals as a class are nongenotoxic (Flamm and Lehman-McKeeman, 1991; Hard et al., 1993; Hard and Whysner, 1994). For d-limonene, the major metabolite that binds to alpha2u-globulin is d-limonene 1,2-epoxide. The identification of a potentially reactive epoxide intermediate that binds to alpha2u-globulin might raise some concern that this epoxide could also react with DNA and be responsible for the renal carcinogenicity of d-limonene. However, that appears to be unlikely, inasmuch as the 1,2-epoxide is a very stable moiety that binds reversibly to alpha2u-globulin, without forming covalent adducts (Lehman-McKeeman et al., 1989). In addition, this epoxide is not mutagenic in Salmonella species, does not induce sister-chromatid exchange in V79 cells, and causes no unscheduled DNA synthesis in rat hepatocytes (Flamm and Lehman-McKeeman, 1991; Vonder Hude et al., 1989; Watanabe et al., 1980). Collectively, the lack of genotoxicity of these chemicals, and their metabolites directly involved in the interaction with alpha2u-globulin argues that ''nongenotoxic'' mechanisms must be involved in the chronic carcinogenicity associated with acute alpha2u globulin nephropathy.
Related Proteins and Their Role in Safety Evaluation
Some uncertainty remains about whether human subpopulations might be sensitive to similar proteins and, if so, the molecular identity, kinetics, and biology of these proteins in humans. The alpha2u-globulin found in male rats is structurally related to a group of transport proteins, many of which are synthesized in humans. The proteins in this group of about 20 low-molecular weight proteins, the alpha2u-globulin superfamily of proteins called lipocalins, are similar in molecular weight, certain sequence homology, and tertiary structure (where it is known) to the alpha2u globulin (Lehman-McKeeman and Caudill, 1992b; Pervaiz and Brew, 1987). They include protein I (a minor urinary protein in humans, more abundant in male than female urine), retinol-binding protein (synthesized in the liver by all mammals), apolipoprotein D (isolated from human plasma), pregnancy-associated endometrial alpha2-microglobulin (isolated from human pancreas), bovine
beta-lactoglobulin, and mouse urinary protein (MUP), the protein most closely resembling alpha2u-globulin (Åkerström and Lögdberg, 1990; Bernard et al., 1989; Brooks, 1987; Pervaiz and Brew, 1987; Pevsner et al., 1988; Snyder et al., 1988). The only member of this superfamily with a clearly defined physiologic function is retinol-binding protein, although all of them are thought to be carriers of lipophilic molecules. Although male rats have a much higher urinary protein concentration than humans (Flamm and Lehman-McKeeman, 1991), because concentrations of protein homologues in human urine are well below those of alpha2u-globulin in male rats (Berggard, 1970; Ekstrom and Berggard, 1977; Peterson and Berggard, 1971), it is highly unlikely that enough protein could accumulate in the normal human kidney to result in hyaline-droplet formation (Hard and Whysner, 1994). Abnormal accumulation of hyaline droplets can be seen in disease processes, however. For example, large amounts of proteins are produced in multiple-myeloma patients (Pirani et al., 1983), lysozyme is produced in human mononuclear-cell leukemia (Muggia et al., 1969), and hyaline-droplet accumulation is observed in patients with epidemic hemorrhagic fever who are infused with large amounts of concentrated human serum albumin (Oliver and MacDowell, 1958).
Although mouse urinary protein is more abundant in males, females excrete it. In addition, mouse urinary protein accounts for 4% of the total hepatic mRNA in mice, whereas alpha2u-globulin accounts for about 1% of the rat hepatic MRNA. Lehman-McKeeman and Caudill (1992a,b) reported that neither d-limonene 1,2-oxide nor the reactive metabolite of 2,4,4-trimethyl-2-pentanol binds to MUP or to protein 1. Binding of the reactive metabolite to the protein appears to be necessary for materials to induce a protein nephropathy related to protein accumulation. Differences in renal metabolite concentrations are not clearly due to binding, however, in that most of the metabolite in the kidney is not associated with alpha2u-globulin. If the phenomenon involves a direct chemical action, the delivery of the active metabolite to the target is important, so characterization of the binding, turnover, and transport properties of lowmolecular-weight, lipophilic ligand-binding proteins in humans might be more relevant than the quantities of protein.
Male Rat Renal Tumors in the Assessment of Risk to Humans
The preceding discussion has laid out the predominant view of the mode of action in the development of renal tumors caused by alpha2u-globulin. An alternative mode of action has been proposed by Melnick (1992), who suggested that other factors besides alpha2u globulin accumulation might be involved in the proliferative response. A nongenotoxic chemical that causes renal tumors in male rats but not in female rats or in other species is probably acting through the alpha2u-globulin mechanism if there is evidence of hyaline-
droplet formation and of alpha2u-globu-lin in the droplets, there is granular cast formation in the outer medulla and papillary linear mineralization, there is increased cell replication in the renal tubules, and there is no evidence of another plausible mechanism.
The renal carcinogens believed to work through the alpha2u-globulin mechanism fulfill the first two criteria; several also fulfill the third criterion, and some fulfill the fourth. But d-limonene and 2,2,4-trimethylpentane are the only two of these chemicals that have been sufficiently investigated to establish binding of their metabolites to alpha2u-globulin and, perhaps just as important, the absence of binding to other members of the superfamily of alpha2u-globulin lowmolecular-weight proteins (Lehman-McKeeman and Caudill, 1992a).
Tetrachloroethylene is an interesting example of a chemical that has been shown to cause alpha2u-globulin accumulation and to induce renal tumors at a low incidence in male rats. The hyaline droplets appear only at very high doses—only at concentrations above the high dose of the carcinogenicity bioassay (Goldsworthy et al., 1988a; Green et al., 1990; Parker et al., 1992; USEPA, 1991c). In addition to the postulated alpha2u-globulin mechanism, scientific evidence supports other alternative mechanisms. At least four different mechanisms of renal carcinogenesis have been postulated for tetrachloroethylene (Parker et al., 1992; USEPA, 1991c). Cytotoxic and potent mutagenic tetrachloroethylene metabolites that could contribute to the tumorigenesis of this chemical are formed in the kidney (Dekant et al., 1986; Green and Odum, 1985; Green et al., 1990; Vamvakas et al., 1989). Peroxisome proliferation occurs in the kidneys of rodents exposed to tetrachloroethylene and conceivably could contribute to the induction of renal tumors (Goldsworthy and Popp, 1989). In the case of tetrachloroethylene, the mode of renal turnorigenesis is not understood. Therefore, the renal tumors observed in male rats after tetrachloroethylene exposure would be considered as contributing to the overall weight of evidence in identifying potential cancer hazards to humans, although the tumors would not be used to derive a quantitative risk estimate because of the feasibility of their resulting in part from the alpha2u-globulin nephropathy.
d-Limonene is the example of a tested chemical that meets all the criteria for the alpha2u-globulin process of renal-tumor induction. The renal-tumor data on d-limonene do not indicate much likelihood of this chemical's producing human cancer, and it is appropriate to refrain from extrapolating the potential hazard and risk rate from the male rat renal tumors produced by d-limonene to humans (USEPA, 1991a,b; Hard and Whysner, 1994).
Studies have shown increased excretion of alpha2u-globulin after low to moderate lead exposure (Mathias et al., 1992) and have identified renal leadbinding proteins (Fowler and DuVal, 1991) as the kidney-specific cleavage product of alpha2u-globulin minus the first nine N-terminal residues. Recent
studies have demonstrated the presence of similar lead-binding proteins in human kidney, liver, and brain cytosolic fractions (Kahng et al., 1992). Further research is needed to determine the possible roles of these proteins in mediating the observed increase (Steenland et al., 1992) in renal cancer in persons occupationally exposed to lead.
SUMMARY
New technologies using modern advances in molecular biology, genetic engineering, in vitro tissue-culture systems, and animal-model systems provide a wide variety of new methods that can be applied to biologic-marker research. The new technologies are rapidly being integrated into strategies for biologicmarker research pertaining to the genitourinary tract and, in a broader perspective, to other organ systems as well. The potential number of biologic markers is enormous. Each newly identified marker will require careful evaluation before its use in human population studies.
Several important lessons have been learned from the use of PSA as a clinical biologic marker for prostatic cancer. The uncertainties associated with its use have emphasized the need for the cooperation of various disciplines—including biostatistics and epidemiology, the basic sciences, and clinical specialties—to review the utility of specific biologic markers before their integration into clinical practice.
Extensive studies have attempted to extrapolate data concerning alpha2u-globulin from animal studies to human conditions, such as nephrotoxicity and renal cancer. The results emphasize the need for caution in the extrapolation of in vitro and nonhuman in vivo studies to humans. At the same time, it is notable that potential biologic markers, such as alpha2u-globulin, can be studied in relation to environmental stress. Some markers are quantifiable at the singlecell level, and the interactions of various cells can be investigated. New technologies—such as differential PCR, quantitative in situ PCR, and quantitative fluorescence image analysis (QFIA)—facilitate the use of biologic markers to describe genotypic and phenotypic changes associated with xenobiotic exposure.
New technologies can also promote the study of biologic markers at the cellular level. Particularly important are the changes that occur in nucleic acid synthesis, which regulate cell growth and death and which enhance our understanding of carcinogenesis, nephrotoxicity, and other diseases of the genitourinary tract.


























