
Engineering Resilience versus Ecological Resilience
C. S. Holling
Ecosystem Structure And Function
Ecological science has been shaped largely by the biological sciences. Environmental science, on the other hand, has been shaped largely by the physical sciences and engineering. With the beginning of interdisciplinary efforts between the two fields, some of the fundamental differences between them are generating conflicts caused more by misunderstanding of basic concepts than by any difference in social purposes or methods. Those differences are most vivid in that part of ecology called ecosystem science, for it is there that it is obvious that both the biota and the physical environment interact such that not only does the environment shape the biota but the biota transforms the environment.
The accumulated body of empirical evidence concerning natural, disturbed, and managed ecosystems identifies key features of ecosystem structure and function (Holling et al., 1995) that probably are not included in many engineers' image of ecology:
- Ecological change is not continuous and gradual; Rather it is episodic, with slow accumulation of natural capital such as biomass or nutrients, punctuated by sudden releases and reorganization of that capital as the result of internal or external natural processes or of man-imposed catastrophes. Rare events, such as hurricanes, or the arrival of invading species, can unpredictably shape structure at critical times or at locations of increased vulnerability. The results of these rare events can persist for long periods. Therein lies one of the sources of new options that biological diversity provides. Irreversible or slowly reversible states exist—that is, once the system flips into such a state, only explicit manage-
- ment intervention can return its previous self-sustaining state, and even then success is not assured (Walker, 1981). Critical processes function at radically different rates covering several orders of magnitude, and these rates cluster around a few dominant frequencies.
- Spatial attributes are not uniform or scale invariant. Rather, productivity and textures are patchy and discontinuous at all scales from the leaf to the individual, the vegetation patch, the landscape, and the planet. There are several different ranges of scales each with different attributes of patchiness and texture (Holling, 1992). Therefore scaling up from small to large cannot be a process of simple linear addition; nonlinear processes organize the shift from one range of scales to another. Not only do the large and slow variables control small and fast ones, the latter occasionally "revolt" to affect the former.
- Ecosystems do not have single equilibria with functions controlled to remain near them. Rather, destabilizing forces far from equilibria, multiple equilibria, and disappearance of equilibria define functionally different states, and movement between states maintains structure and diversity. On the one hand, destabilizing forces are important in maintaining diversity, resilience, and opportunity. On the other hand, stabilizing forces are important in maintaining productivity and biogeochemical cycles, and even when these features are perturbed, they recover rather rapidly if the stability domain is not exceeded (e.g., recovery of lakes from eutrophication or acidification, Schindler, 1990; Schindler et al., 1991).
- Policies and management that apply fixed rules for achieving constant yields (such as constant carrying capacity of cattle or wildlife or constant sustainable yield of fish, wood, or water), independent of scale, lead to systems that gradually lose resilience and suddenly break down in the face of disturbances that previously could be absorbed (Holling, 1986). Ecosystems are moving targets, with multiple potential futures that are uncertain and unpredictable. Therefore management has to be flexible, adaptive, and experimental at scales compatible with the scales of critical ecosystem functions (Walters, 1986).
The features described above are the consequence of the stability properties of natural systems. In the ecological literature, these properties have been given focus through debates on the meaning and reality of the resilience of ecosystems. For that reason, and because the same debate seems to be emerging in economics, I will review the concepts to provide a foundation for understanding.
The Two Faces Of Resilience
Resilience of a system has been defined in two different ways in the ecological literature. These differences in definition reflect which of two different aspects of stability are emphasized. I first emphasized the consequences of those different aspects for ecological systems to draw attention to the paradoxes be-
tween efficiency and persistence, or between constancy and change, or between predictability and unpredictability (Holling, 1973). One definition focuses on efficiency, constancy, and predictability—all attributes at the core of engineers' desires for fail-safe design. The other focuses on persistence, change, and unpredictability—all attributes embraced and celebrated by biologists with an evolutionary perspective and by those who search for safe-fail designs.
The first definition, and the more traditional, concentrates on stability near an equilibrium steady state, where resistance to disturbance and speed of return to the equilibrium are used to measure the property (O'Neill et al., 1986; Pimm, 1984; Tilman and Downing, 1994). That view provides one of the foundations for economic theory as well and may be termed engineering resilience.
The second definition emphasizes conditions far from any equilibrium steady state, where instabilities can flip a system into another regime of behavior—that is, to another stability domain (Holling, 1973). In this case the measurement of resilience is the magnitude of disturbance that can be absorbed before the system changes its structure by changing the variables and processes that control behavior. We shall call this view ecological resilience (Walker et al., 1969).
The same differences have also begun to emerge in economics with the identification of multistable states for competing technologies because of increasing returns to scale (Arthur, 1990). Thus, increasingly it seems that effective and sustainable development of technology, resources, and ecosystems requires ways to deal not only with near-equilibrium efficiency but with the reality of more than one equilibrium. If there is more than one equilibrium, in which direction should the finger on the invisible hand of Adam Smith point? If there is more than one objective function, where does the engineer search for optimal designs?
These two aspects of a system's stability have very different consequences for evaluating, understanding, and managing complexity and change. I argue that designing with ecosystems requires an emphasis on the second definition of resilience, that is, the amount of disturbance that can be sustained before a change in system control and structure occurs—ecological resilience. I do so because that interplay between stabilizing and destabilizing properties is at the heart of present issues of development and the environment—global change, biodiversity loss, ecosystem restoration, and sustainable development.
The two contrasting aspects of stability—essentially one that focuses on maintaining efficiency of function (engineering resilience) and one that focuses on maintaining existence of function (ecological resilience)—are so fundamental that they can become alternative paradigms whose devotees reflect traditions of a discipline or of an attitude more than of a reality of nature.
Those who emphasize the near-equilibrium definition of engineering resilience, for example, draw predominantly from traditions of deductive mathematical theory (Pimm,. 1984) where simplified, untouched ecological systems are imagined, or from traditions of engineering, where the motive is to design sys-
tems with a single operating objective (DeAngelis, 1980; O'Neill et al., 1986; Waide and Webster, 1976). On the one hand, that makes the mathematics more tractable, and on the other, it accommodates the engineer's goal to develop optimal designs. There is an implicit assumption of global stability, that is, that only one equilibrium steady state exists, or, if other operating states exist, they should be avoided (Figure 1) by applying safeguards.
Those who emphasize the stability domain definition of resilience (ecological resilience), on the other hand, come from traditions of applied mathematics and applied resource ecology at the scale of ecosystems. Examples include the dynamics and management of freshwater systems (Fiering, 1982), of forests (Holling et al., 1977), of fisheries (Waiters, 1986), of semiarid grasslands (Walker et al., 1969) and of interacting populations in nature (Dublin et al., 1990; Sinclair et al., 1990). Because these studies are rooted in inductive rather than deductive theory formation and in experience with the impacts of large-scale management disturbances, the reality of flips from one operating state to another cannot be avoided. Moreover, it becomes obvious that the variability of critical variables forms and maintains the stability landscape (Figure 2).
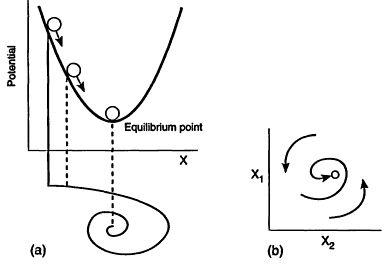
Figure 1
Two views of a single, globally stable equilibrium. (a) Provides a mechanical ball and topography analogy. (b) Provides an abstract state space view of a point's movement toward the stable equilibrium, with x1 and x2 defining, for example, population densities of predator and prey, or of two competitors. This is an example of engineering resilience. It is measured by the resistance of the ball to disturbances away from the equilibrium point and the speed of return to it.
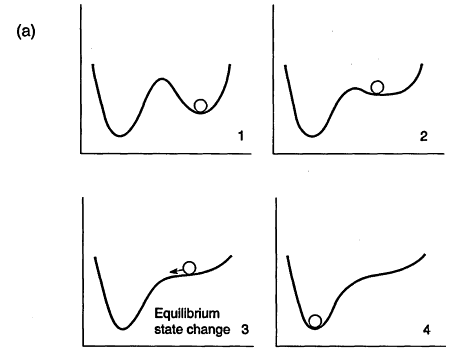
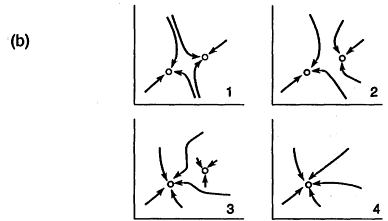
FIGURE 2
Topographic analogy and state space views of evolving nature. The system modifies its own possible states as it changes over time from 1 to 4. In this example, as time progresses, a progressively smaller perturbation is needed to change the equilibrium state of the system from one domain to the other, until the system spontaneously changes state. (a) Ball and topography analogy. (b) Equivalent state space representation.
Managing For Engineering Resilience
Management and resource exploitation can overload waters with nutrients, turn forests into grasslands, trigger collapses in fisheries, and transform savannas into shrub-dominated semideserts. One example, described by Walker et al. (1969) concerns grazing of semiarid grasslands. Under natural conditions in east and south Africa, the grasslands were periodically pulsed by episodes of intense grazing by various species of large herbivores. Directly as a result, a dynamic balance was maintained between two groups of grasses. One group contains species able to withstand grazing pressure and drought because of their deep roots. The other contains species that are more efficient in turning the sun's energy into plant material, are more attractive to grazers, but are more susceptible to drought because of the concentration of biomass above ground in photosynthetically active foliage.
The latter, productive but drought-sensitive grasses, have a competitive edge between bouts of grazing so long as drought does not occur. But, because of pressure from pulses of intense grazing, that competitive edge for a time shifts to the drought-resistant group of species. As a result of these shifts in competitive advantage, a diversity of grass species serves a set of interrelated functions— productivity on the one hand and drought protection on the other.
When such grasslands are converted to cattle ranching, however, the cattle have been typically stocked at a sustained, moderate level, so that grazing shifts from the natural pattern of intense pulses separated by periods of recovery, to a more modest but persistent impact. Natural variability is replaced by constancy of production. The result is that, in the absence of intense grazing, the productive but drought-sensitive grasses consistently have advantage over the drought-resistant species and the soil- and water-holding capacity they protect. The land becomes more productive in the short-term, but the species assemblage narrows to emphasize one functional type. Droughts can no longer be sustained and the system can suddenly flip to become dominated and controlled by woody shrubs. That is, ecological resilience is reduced. It is an example of what Schindler (1990, 1993) has demonstrated experimentally in lakes as the effect of a reduction of species diversity when those species are part of a critical ecosystem function.
There are many examples of managed ecosystems that share this same feature of gradual loss of functional diversity with an attendant loss of resilience followed by a shift into an irreversible state, such as occurs in agriculture and in forest, fish, and grasslands management (as summarized in Holling, 1986). In each case the cause is reduction of natural variability of the critical structuring variables such as plants, insect pests, forest fires, fish populations, or grazing pressure to achieve a social, economic, or engineering objective. The result is that the ecosystem evolves to become more spatially uniform, less functionally diverse, and more sensitive to disturbances that otherwise could have been ab-
sorbed. That is, ecological resilience decreases even though engineering resilience might be great. Short-term success in stabilizing production leads to long-term surprise.
Moreover, such changes can be essentially irreversible because of accompanying changes in soils, hydrology, disturbance processes, and species complexes that regulate or control ecological structure and dynamics. Control of ecosystem function shifts from one set of interacting physical and biological processes to a different set (Holling et al., 1995).
In the examples of resource management that I have explored in depth, not only do ecosystems become less resilient when they are managed with the goal of achieving constancy of production, but the management agencies, in their chive for efficiency, also become more myopic, the relevant industries become more dependent and static, and the public loses trust (Gunderson et al., 1995). This seems to define an ultimate pathology that typically can lead to a crisis triggered by unexpected external events, sometimes followed by a reformation of policy. I first saw the form of this pathology emerging in the early stages of testing and developing theories, methods, and case study examples of adaptive environmental assessment and management. Those cases and their diagnoses were summarized in Holling (1986).
Those cases involved a number of different examples of forest development, fisheries exploitation, semiarid grazing systems, and disease management in crops and people. We have greatly expanded and deepened the case studies and tests since then, adding examples that are presented in a new book that explores both the dynamics of ecosystems and the dynamics of the institutions that attempt to manage them (Gunderson et al., 1995). Two of the original examples continue to provide insights.
In those two examples, the initial diagnoses of the pathology as I saw it in the early 1970s were as follows:
- Successful suppression of spruce budworm populations during the 1950s and 1960s in eastern Canada, using insecticide, certainly preserved the pulp and paper industry in the short-term by significantly reducing defoliation by the insect so that tree mortality was delayed. This encouraged expansion of pulp mills but left the forest, and hence the economy, more vulnerable to an outbreak that would cause more intense and more extensive tree mortality than had ever been experienced before. That is, the short-term success of spraying led to moderate levels of infestation and partially protected foliage that became more homogeneous over larger areas, demanding ever more vigilance and control.
- Effective protection and enhancement of salmon spawning through use of fish hatcheries on the west coast of North America quickly led to more predictable and larger catches by both sport and commercial fishermen. That triggered increased fishing pressure and investment in both sectors, pressure that caused more and more of the less productive natural stocks to become locally extinct.
That left the fishing industry precariously dependent on a few artificially enhanced stocks, whose productivity began declining in a system where larger-scale physical oceanic changes contributed to unexpected impacts on the distribution and abundance of fish.
In both those cases, however, by the 1980s I began to realize that the phase of a growing pathology was transient and could be broken by a spasmodic readjustment, an adaptive lurch of learning that created new opportunity. It is that creation of something fundamentally novel that gives an evolutionary character to development of a region that might make sustainable development an achievable reality rather than an oxymoron.
The heart of these two different views of resilience lies in assumptions regarding whether multistable states exist. If it is assumed that only one stable state exists or can be designed to so exist, then the only possible definitions for, and measures of, resilience are near-equilibrium ones—such as characteristic return time. Prod that is certainly consistent with the engineer's desire to make things work, not to make things that break down or suddenly shift their behavior. But nature is different.
There are different stability domains in nature, and variation in critical variables tests the limits of those domains. Thus, a near-equilibrium focus seems myopic and attention shifts to determining the constructive role of instability in maintaining diversity and persistence and to designs of management that maintain ecosystem function in the face of unexpected disturbances. Such designs would maintain or expand ecological resilience. It is those ecosystem functions and ecological resilience that provide the ecological ''services'' that invisibly provide the foundations for sustaining economic activity.
Managing For Ecological Resilience
There is a puzzle in these examples and this analysis. It implies that efficient control and management of renewable resources in an engineering sense leads initially to success in managing a target variable for sustained production of food or fiber but ultimately to a pathology of less resilient and more vulnerable ecosystems, more rigid and unresponsive management agencies, and more dependent societies. But there seems to be something inherently wrong with that conclusion, implying, as it does, that the only solution is humanity's radical return to being "children of nature." The puzzle needs to be clarified to test its significance and generality.
The above conclusion is based on two critical points. One is that reducing the variability of critical variables within ecosystems inevitably leads to reduced resilience and increased vulnerability. The second is that there is, in principle, no different way for agencies and people to manage and benefit from resource development. Both points are explored in more detail in a new book on barriers and
bridges to ecosystem and institutional renewal (Gunderson et al., 1995), so here I will deal only with highlights.
Puzzles can sometimes be solved by searching for counterexamples. Oddly, nature itself provides such counterexamples of tightly regulated yet sustainable systems in the many examples of physiological homeostasis. Consider temperature regulation of endotherms (warm-blooded animals). The internal body temperature of endotherms is not only tightly regulated within a narrow band, but among present-day birds and mammals, the average temperature is perilously close to lethal. Moreover, the cost of achieving that regulation requires ten times the energy for metabolism that is required by ectotherms (cold-blooded animals). That would seem to be a recipe for not only disaster but a very inefficient one at that. And yet evolution somehow led to the extraordinary success of the animals having such an adaptation—the birds and mammals.
To test the generality of the variability-loss/resilience-loss hypothesis, I have been collecting data from the physiological literature on the viable temperature range within the bodies of organisms exposed to different classes of variability. I have organized the data into three groups ranging from terrestrial ectotherms, which are exposed to the greatest variability of temperature from unbuffered ambient conditions, to aquatic ectotherms, which are exposed to an intermediate level of variability because of the moderating attributes of water, to endotherms, which regulate temperature within a narrow band. The viable range of internal body temperature decreases from about 40 degrees centigrade for the most variable group to about 30 degrees for the intermediate, to 20 degrees for the tightly regulated endotherms. Therefore resilience, in this case the range of internal temperatures that separates life from death, clearly does contract as variability in internal temperature is reduced, just as in the resource management cases. I conclude, therefore, that reduction of variability of living systems, from organisms to ecosystems, inevitably leads to loss of resilience in that part of the system being regulated.
But that seems to leave an even starker paradox for management; seemingly successful control inevitably leads to collapse. But, in fact, endothermy does persist and flourish. It therefore serves as a revealing metaphor for sustainable development. This metaphor contains two features that were not evident in my earlier descriptions of examples of resource management.
First, the kind of regulation is different. Five different mechanisms, from evaporative cooling to metabolic heat generation, control the temperature of endotherms. Each mechanism is not notably efficient by itself. Each operates over a somewhat different but overlapping range of conditions and with different efficiencies of response. It is this overlapping "soft" redundancy that seems to characterize biological regulation of all kinds. It is not notably efficient or elegant in the engineering sense. But it is robust and continually sensitive to changes in internal body temperature. That is quite unlike the examples of rigid
regulation by management where goals of operational efficiency gradually isolated the regulating agency from the things it was regulating.
Examples of similar regulation of ecosystem dynamics in nature include the set of herbivorous antelope species that structure the vegetation of the savannas of East Africa at intermediate scales from meters to kilometers (Walker et al., 1969) or the suite of 35 species of insectivorous birds that, through their predation on insect larvae, set the timing for outbreaks of spruce budworm in the forests of eastern Canada (Holling, 1988). In these examples, each species performs its actions somewhat differently from others, and each responds differently to external variability because of differences in habitat preference and the scales of choice for its resources (Holling, 1992). As an example, some species of insectivorous birds exert modest predation pressure over a broad range of prey densities, whereas others exert strong pressure over narrow ranges of density and still others function between those extremes. The densities at which the predation impact is maximal also differ between species. Competition occurs among these species such that the aggregate predation effect is inefficient when predators are abundant and prey scarce and efficient when the reverse is true. As a consequence, the result of their joint action is an overlapping set of reinforcing influences that are less like the redundancy of engineered devices and more like portfolio diversity strategies of investors. The risks and benefits are spread widely to retain overall consistency in performance independent of wide fluctuations in the individual species. That is at the heart of the role of functional diversity in maintaining the resilience of ecosystem structure and function.
We chose the term functional diversity to describe this process, following the terms suggested by Schindler (1990) and by Holling et al. (1995). Such diversity provides great robustness to the process and, as a consequence, great resilience to the system behavior.
The second feature of nature's way of tightly regulating variability that is different from traditional management is the tendency to function near the edge of instabilities, not far away from them. That is where information and opportunity are the greatest. Again endothermy provides an example. Endothermy is a true innovation that explosively released opportunity for the organisms that evolved the ability to regulate their body temperature. Maintaining high body temperature, just short of death, allows the greatest range of external activity for an animal. Speed and stamina increase and activity can be maintained at both high and low external temperatures. A range of habitats forbidden to an ectotherm is open to an endotherm. The evolutionary consequence of temperature regulation was to open opportunity suddenly for dramatic organizational change and the adaptive radiation of new life forms. Variability is therefore not eliminated. It is reduced in one place and transferred from the animal's internal environment to its external environment as a consequence of allowing continual probes by the whole animal for opportunity and change. Hence the price of reducing internal resilience, maintaining high metabolic levels, and operating
dose to an edge of instability is more than offset by that creation of evolutionary opportunity. Nature's policy of ecological resilience, if we can call it that, seems far from those of traditional engineering safeguards or economic efficiency, where operating near an equilibrium far from an instability defines engineering resilience.
But ascribing that designation to engineering is to stereotype the field with only one face of its activities, just as ecological resilience represents only one face of ecology. At least some aspects of ecologically resilient control are equally familiar to the control engineer, for operation at the edge of instability is characteristic of designs for high-performance aircraft. Oddly, the result is opportunity. Effective control of internal dynamics at the edge of instability generates external options. Operating at the edge of instability generates immediate signals of changing opportunity.
That surely is at the heart of sustainable development—the release of human opportunity. It requires flexible, diverse, and redundant regulation, early signals of error built into incentives for corrective action, and continuous experimental probing of the changes in the external world. Those are the features of adaptive environmental and resource management. Those are the features missing in the descriptions I presented of traditional, piecemeal, exploitive resource management and its ultimate pathology.
Conclusion
There are indeed strong suggestions that management and institutional regimes can be designed to preserve or expand resilience of systems as well as provide developmental opportunity. It is a central issue that only now is beginning to be the focus of serious scholarship and practice. Of the cases I know well, management of the forests of New Brunswick seems most clearly to demonstrate the cycles of crisis and learning and the hesitant emergence of a more sustainable path.
In the New Brunswick example, one major crisis and several minor ones have occurred since the early 1950s. During this period, the new technologies of airplanes and pesticides developed in World War II were adapted for spraying operations and their use was progressively refined to achieve high mortality of insects while reducing environmental side effects. These procedures for pesticide control of budworm synergized with other technological developments in tree harvesting, pulp production chemistry, and mill construction and resulted in large investments in pulp production. Minor crises occurred when effects on human health were linked to the pesticides. Key pieces of integrated understanding of the natural system were achieved by the teams of Morals (1963) and the modelers of the 1970s (Clark et al., 1979). The brittleness that developed (defined by a loss of ecological resilience together with an increase in institutional efforts to control information and action) reflected the complacent belief among
agency staffs that budworm damage was controlled in an efficient and cost-effective manner and that there was plenty of wood available for harvest. In reality, the costs of using pesticides were rapidly increasing because of increases in oil prices and because of modification of pesticide application in response to public pressure. In addition, available stocks of harvestable trees were decreasing because of past harvests and because more and more mature stands over larger areas were gradually deteriorating from the pressure of moderate but persistent budworm defoliation. The major crises occurred during the late 1970s when a forest inventory report finally indicated that there would not be sufficient stock to support the current mills, thereby confirming an earlier prediction of the models. This led to a new law that restructured the licensing and forest management policies and freed the innovative capacity of local industries within a regional set of goals and constraints. A sequence of adaptive responses among the actors began to develop regional forest policy in a way that now engages local industrial, environmental, and recreational goals.
The examples of growing pathology are caused by the very success of achieving near equilibrium behavior and control of a single target variable independently of the larger ecosystem, economic, and social interactions. When that orientation or goal is abandoned, it happens suddenly, in response to perceived or real crises. The scale of the issues becomes redefined more broadly from a local to a regional setting and from short-term to long-term. The scientific understanding of the natural system becomes more integrated, and the issues themselves are not posed in response to needs to maximize constancy or productivity of yield, but to ones of designing interrelations between people and resources that are sustainable in the face of surprises and the unexpected. If there is such a thing as sustainable development, then that is it. The key features are integration of knowledge at a range of scales, engagement of the public in exploring alternative potential futures, adaptive designs that acknowledge and test the unknown, and involvement of citizens in monitoring and understanding outcomes. That is possible only in situations where ecological resilience and public trust have not been degraded. If they have, as in many situations, then the initial goal has to be the restoration of both resilience and trust.
References
Arthur, B. 1990. Positive feedback in the economy. Scientific American 262:92-99.
Clark, W. C., D. D. Jones, and C. S. Holling. 1979. Lessons for ecological policy design: A case study of ecosystem management. Ecological Modeling 7:1-53.
DeAngelis, D.L. 1980. Energy flow, nutrient cycling and ecosystem resilience. Ecology 61:764-771.
Dublin, H. T., A. R. E. Sinclair, and J. MeGlade. 1990. Elephants and fire as causes of multiple stable states in the Serengeti-mara woodlands. Journal of Animal Ecology 59:1147-1164.
Fiering, M.B. 1982. Alternative indices of resilience. Water Resources Research 18:33-39.
Gunderson, L. H., C. S. Holling, and S. Light. 1995. Barriers and Bridges to Renewal of Ecosystems and Institutions. New York: Columbia University Press.
Holling, C.S. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4:1-23.
Holling, C. S. 1986. Resilience of ecosystems; local surprise and global change. Pp. 292-317 in Sustainable Development of the Biosphere, W. C. Clark and R. E. Munn, eds. Cambridge, England: Cambridge University Press.
Holling, C. S. 1988. Temperate forest insect outbreaks, tropical deforestation and migratory birds . Memoirs of the Entomological Society of Canada 146:21-32.
Holling, C.S. 1992. Cross-scale morphology, geometry and dynamics of ecosystems. Ecological Monographs 62(4):447-502.
Holling, C. S., D. D. Jones, and W. C. Clark. 1977. Ecological policy design: A case study of forest and pest management. IIASA CP-77-6:13-90 in Proceedings of a Conference on Pest Management, October 1976, G. A. Norton and C. S. Holling, eds. Laxenburg, Austria.
Holling, C. S., D. W. Schindler, B. Walker, and J. Roughgarden. 1995. Biodiversity in the functioning of ecosystems: An ecological primer and synthesis. In Biodiversity Loss: Ecological and Economic Issues, C. Perrings, K-G Mäer, C. Folke, C. S. Holling, and B-O Jansson, eds. Cambridge, England: Cambridge University Press.
Morris, R. F. 1963. The dynamics of epidemic spruce budworm populations. Memoirs of the Entomological Society of Canada 21:332.
O'Neill, R. V., D. L. DeAngelis, J. B. Waide, and T. F. H. Allen. 1986. A Hierarchical Concept of Ecosystems. Princeton, N.J.: Princeton University Press .
Pimm, S.L. 1984. The complexity and stability of ecosystems. Nature 307:321-326.
Schindler, D.W. 1990. Experimental perturbations of whole lakes as tests of hypotheses concerning ecosystem structure and function. Proceedings of 1987 Crafoord Symposium. Oikos 57:25-41.
Schindler, D.W. 1993. Linking species and communities to ecosystem management. Proceedings of the 5th Cary Conference, May 1993.
Schindler, D. W., T. M. Frost, K. H. Mills, P.S.S. Chang, I. J. Davis, F. L. Findlay, D. F. Malley, J. A. Shearer, M. A. Turner, P. J. Garrison, C. J. Watras, K. Webster, J. M. Gunn, P. L. Brezonik, and W. A. Swenson. 1991. Freshwater acidification, reversibility and recovery: Comparisons of experimental and atmospherically-acidified lakes. Volume 97B: 193-226 in Acidic Deposition: Its Nature and Impacts, F. T. Last and R. Watling, eds. Proceedings of the Royal Society of Edinburgh.
Sinclair, A. R. E., P. D. Olsen, and T. D. Redhead. 1990. Can predators regulate small mammal populations? Evidence from house mouse outbreaks in Australia. Oikos 59:382-392.
Tilman, D., and J. A. Downing. 1994. Biodiversity and stability in grasslands. Nature 367:363-365.
Waide, J. B., and J. R. Webster. 1976. Engineering systems analysis: Applicability to ecosystems. Volume IV, pp. 329-371 in Systems Analysis and Simulation in Ecology, B.C. Patten, ed. New York: Academic Press.
Walker, B.H. 1981. Is succession a viable concept in African savanna ecosystems? Pp. 431-447 in Forest Succession: Concepts and Application, D.C. West, H. H. Shugart, and D. B. Botkin, eds. New York: Springer-Verlag.
Walker, B. H., D. Ludwig, C. S. Holling, and R. M. Peterman. 1969. Stability of semi-arid savanna grazing systems. Ecology 69:473-498.
Walters, C.J. 1986. Adaptive Management of Renewable Resources. New York: McGraw Hill.














