




Below is the uncorrected machine-read text of this chapter, intended to provide our own search engines and external engines with highly rich, chapter-representative searchable text of each book. Because it is UNCORRECTED material, please consider the following text as a useful but insufficient proxy for the authoritative book pages.
Riboflavin (Vitamin Be) Riboflavin was first isolated from egg white and was later isolated from milk and liver. The name was adopted after the compound was shown to contain the sugar alcohol, ribitol (Figure 12~. It is very slightly solu- ble in water (11 mg/100 ml) at a neutral or acid pH, but is highly soluble in alkaline solution. Riboflavin is synthe- sized by the gut microflora and, thus, may reduce the dietary need for the vitamin. Riboflavin is present at various concentrations in a wide range of feeds. It is present in significant quantities in fresh pasture but is low in cereals where it is complexed with proteins. Ribo- flavin supplements to diets are light-sensitive and may be subject to loss during storage. 1 NUTRITIONAL ROLE Dietary Requirements of Various Species Riboflavin is required in the diets of nonruminant ani- mals. Ruminants obtain sufficient amounts of the vita- min from that synthesized by the rumen microflora. Deficient intakes of the vitamin result in impaired growth. Chronic riboflavin deficiency can be fatal. Spe- cific signs of riboflavin deficiency include seborrheic dermatitis, cheilosis, conjunctivitis, and congenital mal- formations in rats and mice; curled toe paralysis, re- duced embryonic survival, and dermatitis in poultry; muscular weakness, ataxia, dermatitis, anemia, and cardiological changes in dogs and foxes; dermatitis, alo- pecia, ataxia, corneal degeneration, hemmorhagic adre- nals, and fatty degeneration of the kidneys in pigs; and dermatitis and normocytic hypochromic anemia in primates (Nutrition Foundation, 1984~. The riboflavin requirements for most species range from 3 to 7 mg/kg of diet. 53 Biochemical Functions Riboflavin functions in the intermediary transfer of electrons in metabolic oxidation-reduction reactions as two coenzymes, Gavin mononucleotide (FMN) and fla- vin adenine dinucleotide (FAD). The riboflavin coen- zymes function with a large number of oxidases and dehydrogenases important in normal metabolism. Those enzymes that use FMN include glucose oxidase, -amino acid oxidase, and lactate dehydrogenase. Those that use FAD include D-amino acid oxidase, cyto- chrome reductase, succinic dehydrogenase, the acyl- CoA dehydrogenases, ~-gulonolactone dehydrogenase, ~x-glycerophosphate dehydrogenase, and glutathione reductase. The activity of the last enzyme in the eryth- rocyte responds directly to changes in nutritional ribo- flavin status and is, therefore, used as a clinical parameter for that purpose. The riboflavin coenzymes transfer electrons to the pyridine dinucleotides of the mitochondrial electron transport chain. Due to this role in energy metabolism, deficient intakes of riboflavin result in impaired effi- ciency of respiratory energy production. This may result in increases in feed intake by 10 to 15 percent. Reduced electron transport in riboflavin deficiency also results in specific pathologies in those tissues with the greatest normal respiratory rates. FORMS OF THE VITAMIN The dietary form of this vitamin may be riboflavin or the coenzyme forms FMN and FAD, which are the pre- dominant forms in mammalian tissues. The supplemen- tary form for addition to diets is generally riboflavin, although some researchers have used sodium riboflavi- nate. This form is more soluble in water than riboflavin (2 percent versus 11 mg/100 ml).
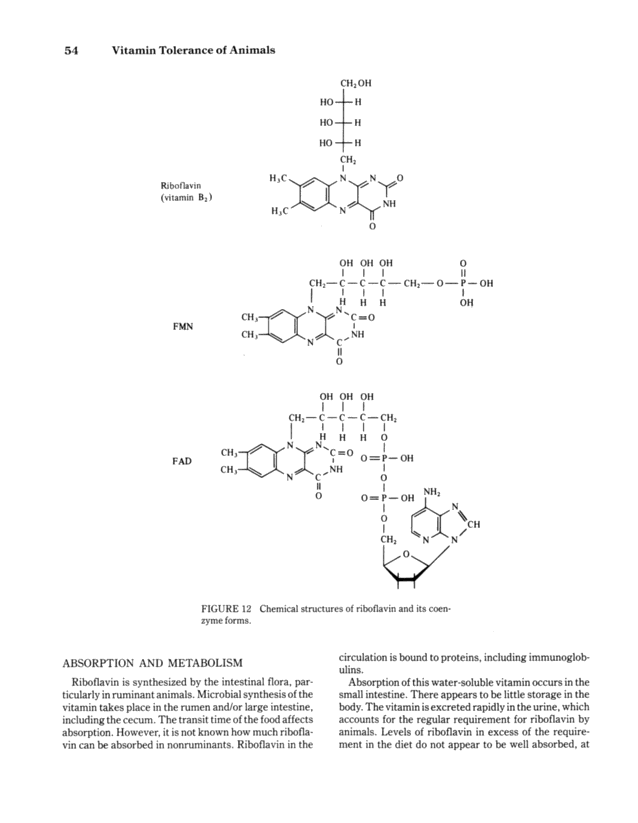
54 Vitamin Tolerance of Animals Ribof lavin (vitamin B2) CH2 OH HO- HO- HO- -H -H -H CH2 H 3 C ~ N ~ N ~ O H3C~ O OH OH OH 1 ~ ~I! CH2-C C-C CH2 O-P - OH 1 1 1 1 1 CH3~N N'C-O FMN l ~ ~ CH3-~ N ~ C ~ NH 1 FAD ABSORPTION AND METABOLISM o 11 o OH OH OH CH2 C C- C CH2 1 1 1 1 1 I H H H O CH3 - :' C=0 0 P OH CH3-IN C,NH 11 o o O-P OH NH2 CH2 AN ~0~ FIGURE 12 Chemical structures of riboflavin and its coen- zyme forms. Riboflavin is synthesized by the intestinal flora, par- ticularly in ruminant animals. Microbial synthesis of the vitamin takes place in the rumen and/or large intestine, including the cecum. The transit time of the food affects absorption. However, it is not known how much ribofla- vin can be absorbed in nonruminants. Riboflavin in the circulation is bound to proteins, including immunoglob- ulins. Absorption of this water-soluble vitamin occurs in the small intestine. There appears to be little storage in the body. The vitamin is excreted rapidly in the urine, which accounts for the regular requirement for riboflavin by animals. Levels of riboflavin in excess of the require- ment in the diet do not appear to be well absorbed, at
Riboflavin 55 least in dogs and rats. Unna and Greslin (1942) reported that only 0.1 percent of a dose of riboflavin (exact form not stated) was recovered in the urine of treated dogs within 24 hours following administration. The feces of rats fed large quantities of riboflavin showed an intense yellow color. Massive deposits of riboflavin were re- ported at the site of subcutaneous injections for as long as 10 days (Unna and Greslin, 1942~. Middleton and Grice (1964) studied riboflavin absorption in the intact rat from an orally administered dose of 10 ,ug and re- ported recoveries of about 27 percent in the urine and 10 percent in the feces. Axelson and Gibaldi (1972) esti- mated that about 20 percent of a 1,000-,ug oral dose of riboflavin was absorbed in the rat. About 4 to 10 percent was recovered in the urine. When the dose was adminis- tered intraperitoneally,47 to 51 percent was recovered. HYPERVITAMINOSIS A summary of the effects of riboflavin administration in animals is shown in Table 13. Seymour et al. (1968) conducted a series of experiments in which 5-week-old pigs were fed diets containing up to 8.8 mg riboflavin/kg at environmental temperatures ranging from -4°C to 32°C for 5 weeks. The minimum riboflavin requirement for maximal BW gain or efficiency of feed conversion was estimated at 3 to 4 mg/kg, with some evidence that it might be increased at low environmental tem~era- tures. Unna and Greslin (1942) found that 10-week-old dogs tolerated oral doses of 24 mg of riboflavin/kg of BW over a 5-month period. No adverse effects on growth or histopathology were reported. Unna and Greslin (1942) reported that the dog could tolerate oral doses of 2 g riboflavin/kg of BW with no evidence of toxicity. Riboflavin toxicity has been studied in laboratory ani- mals. Ellis et al. (1943) fed diets containing 3 to 10 mg riboflavin/kg to successive generations of rats from 28 to 700 days old. A dietary level of 3 mg/kg appeared to be adequate for breeding stock. No adverse effects on growth or reproduction were noted for higher levels of the vitamin. In fact, the level of 10 mg/kg was found to give the progeny slightly improved growth. Leclerc (1979a) fed diets containing 1 to 16 mg riboflavin/kg to lactating rats and reported an increased tissue concen- tration with a plateau at 8 mg/kg of diet for dams and 4 mg/kg for pups. Again, no adverse effects were noted at any level. Burch et al. (1956) fed weanling rats diets containing 0 to 15 mg riboflavin/kg for a 5- to 6-week period. They found that a level of 15 mg/kg resulted in increased FMN and FAD levels in the liver, kidney, and heart, but no adverse effects were noted. Unna and Greslin (1942) administered an oral dose of 10 mg of riboflavin/day for 140 days to weanling rats in each of three successive generations. They reported no adverse effects on growth or reproduction. Doses of up to 5 g/kg of BW were administered by various routes in the same investigation. It was reported that the LD50 for riboflavin or sodium riboflavinate administered orally was more than 10 g/kg of BW. For riboflavin or sodium riboflavinate administered by the intraperitoneal route, the LD50 was 0.56 g/kg of BW; when administered by the subcutaneous route it was 5 g/kg of BW for ribofla- vin and 0.79 g/kg of BW for sodium riboflavinate. Unna and Greslin, (1942) found that the rat tolerated oral doses of 10 g of riboflavin or sodium riboflavinate/kg of BW with no evidence of toxicity. Leclerc (1979b) fed diets containing l, 2, 4 or 6 mg riboflavin/kg to gestating rats and reported larger litter sizes at levels greater than l mg/kg. No effects on prog- eny whole body riboflavin pool or on dam BW, liver riboflavin concentration, or weight gain were noted. About 30 percent of the riboflavin consumed during gestation was recovered in the urine of each group, al- though this fell to about 20 percent just before parturi- tion. Schumacher et al. (1965) fed 0.4 or 10.4 mg of riboflavin/100 g of diet to female rats for 2 weeks prior to mating and through gestation and lactation. They re- ported that the reproduction of the high-riboflavin group was reduced significantly from 68 percent to 38 percent. These results have not been confirmed by other studies. Mean birth weight, number of pups in a litter, and mean pup BW at weaning were not affected significantly. Concentrations of the vitamin were also not affected in the fetal carcasses or in pup livers at wean~ng. PRESUMED UPPER SAFE LEVELS Insufficient data are available to support estimates of the maximum dietary tolerable levels. Because ribofla- vin does not appear to be well absorbed from the gut, it is unlikely to present a hazard to animals when included at high levels in the diet. Leclerc (1979a) reported that a dietary level exceed- ing 8 mg of riboflavin/kg in the rat (about 3 times the dietary requirement) resulted in a plateauing of the tis- sue contents of the vitamin. Other available data sug- gest that levels between 10 and 20 times the dietary requirement can be tolerated safely by rats. Riboflavin is more toxic when administered parenter- ally than when administered orally. Estimates of the rat LD50 for the intraperitoneal, subcutaneous, and oral routes are 0.56 g/kg, 5 g/kg, and more than 10 g/kg of BW, respectively.
56 a) 4= == to a E ._ ·= =: O she ~ ~; Hi: cot .= ~ 0,¢ ¢ ~ 0 ~ CQ A - _ _ _ ~at us ~_ _ _ _ _ _ _ ~5 0 ~ ~ - 41 3 E 0 ~ ~a O ° ° E e C O c c `e o ~= C .,, ~ o A, ~0 ~ 3 C ~ ~ ~ 3 ~ ~ o ~A 3 ~ C ~ I ~ 9- ~ - ' - ~ ~ ~ ~ ~ ~ ~ ~ · . 2 ~ j ~ Zo ~ Zo ~ ~ ~ ~ ·- ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ·- ~ .~ ~ ~ ~ Zo ~ ~ ~ Zo ~ ~ Zo ~ - - 4- ~4 ~4= ~ _ _ O O ~ O ~ ~cn O ~be ~ ~ ~ E .c 3 <,, ~3 0 ~ ~ O == .~.~= ~ ~ ~n cn ~c ~L ~C ~cn ~ cn CQ cn =. ~c .c .> , ~, ~E ~E ' c ° P:: P ~P ~cn° P:: cn B B B ~bO y y B B ~ y E E y y y ° E y y y E Y I , I I O ° ° j D P P P tn ~ ~ ~o 3 ~ ' B~ ~°= ~ ° ~3 ~P ~ ~o oo ~2 "1' ° oo ' ~° ~o ~o ~o b ~· ~tn tn C ~C ~cn CC co u' cn O O ~cn ~P~ ~ ~
Riboflavin 57 SUMMARY 1. Riboflavin is a water-soluble vitamin that is not absorbed well. It is essential in the diets of nonruminant animals. 2. A level exceeding ~ mg of riboflavin/kg of diet (about 3 times the nutritional requirement) results in a plateauing of the tissue contents of the vitamin in rats. Other available data with this species suggest that di- etary levels between 10 and 20 times the requirement (possibly 100 times) can be tolerated safely. 3. Riboflavin administered parenterally is more toxic than when given orally. Estimates of the rat I~D50 for the intraperitoneal, subcutaneous, and oral routes are 0.56 g/kg, 5 g/kg, and more than 10 g/kg of BW, respectively. REFERENCES Axelson, J. E., and M. Gibaldi. 1972. Absorption and excretion of riboflavin in the rat-An unusual example of nonlinear pharmaco kinetics. J. Pharm. Sci. 61:404. l Burch, H. B., O.H. Lowry, A. M. Padilla, and A. M. Combs. 1956. Effects of riboflavin deficiency and realimentation on Gavin en- zymes of tissues. J. Biol. Chem. 223:29. Ellis, L. N., A. Zmachinsky, and H. C. Sherman. 1943. Experiments upon the significance of liberal levels of intake of riboflavin. J. Nutr. 25:153. Leclerc, J. 1979a. Influence of the dietary supply of riboflavin on the vitamin nutritional status of the lactating rat and the litter. Int. J. Vit. Nutr. Res. 49:276. Leclerc, J. 1979b. Vitamin B2 nutritional status of pregnant rats and their offspring in relation to nutritional intake of riboflavin. J. Int. Vitaminol. Nutr. 49:51. Middleton, E. J., and H. C. Grice. 1964. Vitamin absorption studies. IV. Site of absorption of C'4-riboflavin and S35-thiamine in the rat. Can. J. Biochem. 42:353. Nutrition Foundation.1984. P.285 in Present Knowledge in Nutrition. 5th ed. Washington, D.C.: Nutrition Foundation. Schumacher, M. F., M. A. Williams, and R. L. Lyman.1965. Effect of high intakes of thiamine, riboflavin and pyridoxine on reproduction in rats and vitamin requirements of the offspring. J. Nutr. 86:343. Seymour, E. W., V. C. Speer, and V. W. Hays.1968. Effect of environ- mental temperature on the riboflavin requirement of young pigs. J. Anim. Sci.27:389. Unna, K., and J. G. Greslin.1942. Studies on the toxicity and pharma- cology of riboflavin. J. Pharmacol. Exp. Ther. 76:75.
