
4
Technological and Biological Changes and the Future of Pest Management
As a biological process, the agricultural enterprise is profoundly affected by the physiological, biochemical, ecological, and genetic attributes of the organisms involved. Anthropogenic activities within and beyond the agricultural enterprise have the potential to affect agriculture through their effects on the biology of organisms, including those in production (crops and livestock) and pests associated with them. In this chapter, we examine technological changes that have introduced new sources of mortality for pest organisms and evaluate their potential role in contributing to pest management in the future. We also examine how human activities have effected changes in the ecological milieu within which pest-crop interactions take place and the selection regime under which pests evolve, and we relate the changes to prospects for pesticide use in the future.
GLOBAL PESTICIDE MARKET TRENDS
Chemical-Pesticide Market
The global chemical-pesticide market is about $31 billion. It is a mature market with a growth of about 1-2% per year. Breakdowns by product category, crop, and by category for region for global and US markets are shown in Table 4-1 (Agrow 1998) and Table 4-2 (Aspelin and Grube, 1999).
TABLE 4-1 Global Chemical Pesticide Market (1997 Sales)
|
Product |
Sales, billions of dollars |
% |
|
Herbicides |
14.7 |
47.6 |
|
Insecticides |
9.1 |
29.4 |
|
Fungicides |
5.4 |
17.5 |
|
Others |
1.7 |
5.5 |
|
Total |
30.9 |
100.0 |
|
Crop |
Sales, billions of dollars |
% |
|
Fruits, nuts, vegetables |
6.5 |
21.0 |
|
Home and garden, turf, and ornamentals |
5.25 |
17.0 |
|
Oil crops |
1.75 |
5.7 |
|
Cotton |
1.5 |
4.9 |
|
Cereals |
4.0 |
12.9 |
|
Maize |
2.5 |
8.1 |
|
Rice |
2.5 |
8.1 |
|
Sugarbeet |
1.0 |
3.2 |
|
Other |
5.9 |
19.1 |
|
Total |
30.9 |
100.0 |
|
Region |
Sales, billions of dollars |
% |
|
North America |
9.2 |
29.8 |
|
Western Europe |
7.8 |
25.2 |
|
East Asia |
7.1 |
23.0 |
|
Latin America |
3.7 |
12.0 |
|
Rest of World |
3.1 |
10.0 |
|
Worldwide Total |
30.9 |
100.0 |
|
Source: Agrow, 1998. |
||
Biopesticide Market
Biologically based pesticide products (also known as biorational products) generate sales of about $700 million per year (including transgenic crops) worldwide. The market for these products is expected to expand by over 20% over the next 5 years. Table 4-3 shows the sales figures for the global biopesticide market in the last few years and projected for 2004. These pesticides comprise living microorganisms or pheromones (animal-produced chemicals that serves as stimuli for behavioral responses in other individuals of the same species). The most successful biorational
TABLE 4-2 US Chemical Pesticide Market by Category (1997 Sales)
|
Product |
Sales, billions of dollars |
% |
|
Herbicides |
6.8 |
57.5 |
|
Insecticides |
3.6 |
29.9 |
|
Fungicides & Other |
1.5 |
12.6 |
|
Total |
11.9a |
100.0 |
|
aThis estimate for the US pesticide market is larger than other estimates for 1997 sales because it includes expenditures for some nonagricultural pesticide applications. This includes applications by owner/operators and custom/commercial applicators to industry, commercial and governmental facilities, buildings, sites, and land; and homeowner applications to homes and gardens, including lawns. Source: Aspelin and Grube, 1999. |
||
TABLE 4-3 Global Biopesticide Market (in millions of dollars).
|
Year |
|||||
|
Market |
1997 |
1998 |
1999 |
2004a |
% change (1999-2004) |
|
Microbial |
65 |
66 |
67 |
72 |
7.5 |
|
Transgenic plants |
405 |
429 |
455 |
610 |
34.1 |
|
Miscellaneous |
180 |
184 |
188 |
208 |
10.6 |
|
Total |
650 |
679 |
710 |
890 |
25.4 |
|
aestimated Source: Eppes, in press. |
|||||
pesticides are the Bacillus thuringiensis-based (Bt-based) microbial pesticides with current sales of about $140 million. More than 40% of Bt sales are in the United States. Rapid growth of Bt-based biopesticides is occurring as replacements of competitive chemical products that are being banned or phased out in environmentally sensitive areas, in consumer and export markets in which concerns about food residue is high, and in organic food production.
The Industry
Agricultural-Chemical Companies
Of sales revenue of agricultural-chemical companies 7–13% is spent on pesticide research and development. The 1997 pesticide sales of leading agrichemical companies are ranked in Figure 4-1.
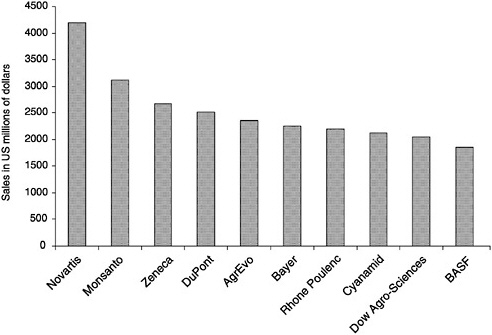
FIGURE 4-1 Pesticide sales of top 10 agrochemical companies, 1997.
Source: Panna 1998.
Major trends in the agrichemical industry today include
-
Consolidation of multinational agrichemical companies.
-
Rapid growth of transgenic-crop markets.
-
Vertical integration of agrichemical firms with seed companies and food processors.
-
Increase in generic pesticides (because chemicals are going off patent)
-
Increase in consolidation and transformation of input distributors.
Despite those trends, the different companies are investing their resources differently in a wide variety of technologies. Table 4-4 shows which companies are pursuing the technologies. Table 4-5 highlights the foci of the companies' pesticide programs.
Major consolidation of multinational companies is under way. A wave of consolidations took place in the 1980s. In the 1990s, the merger of Sandoz and Ciba (forming Novartis) raised the bar even higher, creating an agrichemical company $2 billion larger than the next largest ($5 billion versus $3 billion). Business leaders continue to create empires that can rival Novartis in size. For example, AgrEvo has merged with Rhone-
TABLE 4-4 Comparison of Technologies Pursued by the Pesticide Industry
|
Technology |
Companies |
Products |
Crops/Pest |
Date of Entry |
|
Transgenic seeds |
Monsanto/D&PL |
Bollgard (Bt) |
Cotton / bollworm and budworm |
1996 |
|
Monsanto/Dekalb |
Roundup Ready |
Soybean / weeds |
1996 |
|
|
Calgene/Stoneville |
BXN |
Cotton / weeds |
1996 |
|
|
Monsanto/D&PL |
Roundup Ready |
Cotton / weeds |
1997 |
|
|
Ciba, Dekalb, N.King |
Bt corn |
Corn / borer |
1996, 1997 |
|
|
Mycogen, Pioneer |
Bt corn |
Corn / borer |
1996, 1997 |
|
|
AgrEvo |
Liberty Link |
Canola / weeds |
1996 |
|
|
Dekalb, Ciba, Pioneer |
Liberty Link |
Corn / weeds |
1997 |
|
|
Asgrow |
Virus resistance |
Vegetables and fruits / viruses |
1996 |
|
|
Synthetic chemicals |
Bayer |
Admire/Provado |
Multiple crops / sucking insects |
1995 |
|
AgrEvo |
Applaud |
Cotton / whitefly |
1996 |
|
|
Valent/Sumitomo |
Knack |
Cotton / whitefly |
1996 |
|
|
Rohm & Haas |
Confirm/Intrepid |
Cotton, vegetables and fruits / caterpillars |
1996, 1997 |
|
|
American Cyanamid |
Pirate / Alert |
Cotton and vegetables / caterpillars |
1998 |
|
|
Rhone Poulenc |
Regent |
Cotton, corn, rice, and vegetables / sucking insects |
1998, 1995 |
|
|
Zeneca |
Abound, Heritage |
Multiple crops / fungicide |
1997 |
|
|
BASF |
Allegro |
Multiple crops / powdery mildew |
1997 |
|
|
Novartis |
Bion |
Vegetables / viruses and fungi |
1997 |
|
Natural products |
Thermo Trilogy |
Align |
Vegetables / caterpillars |
1995 |
|
Thermo Trilogy |
Neemix |
Greenhouse / mildew |
1995 |
|
|
Dow |
Tracer |
Cotton and vegetables / caterpillars |
1997 |
|
|
Novartis |
Proclaim |
Cotton and vegetables / caterpillars |
1997 |
|
|
Biocontrol agents |
AgraQuest |
LAGINEX |
Rice and noncrops / mosquito |
1997 |
|
Mycotech |
Mycotrol |
Fruits and vegetables / whitefly |
1996 |
|
|
Thermo Trilogy |
Spod-X |
Cotton / caterpillars |
1996 |
|
|
Ecogen |
AQ10 |
Grapes / powdery mildew |
1996 |
|
|
Thermo Trilogy |
SoilGard |
Greenhouse-potting mix / fungi |
1996 |
|
|
BioWorks |
BioTrek |
Field crops- / root rots |
1996, 1997 |
|
|
Eden Biosciences |
Gray Gold |
Greenhouse / gray mold |
1996, 1997 |
|
|
Liphatech/Gustafson |
UW85 |
Cotton and alfalfa seed / fungi treatment |
1997 |
|
|
Abbott |
DiTerra |
Multiple crops / nematodes |
1997 |
|
|
Abbott |
Trichodex |
Grapes / gray mold |
1997 |
|
|
Abbott |
Spherimos |
Rice and noncrops / Mosquito larvae |
1997 |
TABLE 4-5 Company Pesticide Programsa
|
Pesticidal Natural Products |
Bacillus thuringiensis |
Living Fungi or Bacteria |
Entomopathogenic Nematodes |
Insect Viruses |
Transgenic Crops |
Pheromones |
Synthetic Chemicals |
|
|
Abbott |
0 |
0 |
0 |
0 |
0 |
0 |
||
|
AgraQuest |
0 |
0 |
0 |
0 |
0 |
0 |
||
|
AgrEvo |
0 |
0 |
0 |
0 |
0 |
|||
|
Am Cy |
0 |
0 |
0 |
0 |
0 |
0 |
||
|
BASF |
0 |
0 |
0 |
0 |
0 |
|||
|
Bayer |
0 |
0 |
0 |
0 |
0 |
0 |
||
|
BioWorks |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
Consep |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
Dominion |
0 |
0 |
0 |
0 |
0 |
0 |
||
|
Dow |
0 |
0 |
0 |
0 |
0 |
|||
|
DuPont |
0 |
0 |
0 |
0 |
0 |
|||
|
Ecogen |
0 |
0 |
0 |
0 |
TABLE 4-6 Sales of Transgenic Crops and Chemical Pesticides, 1995-1997
|
Sales, millions of dollars |
||||
|
1995 |
1996 |
1997 |
Change, 1996-1997, % |
|
|
Herbicides |
14,280 |
15,050 |
14,700 |
−2.33 |
|
Insecticides |
8,750 |
8,745 |
9,100 |
4.06 |
|
Fungicides |
5,855 |
5,895 |
5,400 |
−8.40 |
|
Plant-growth regulators and others |
1,305 |
1,325 |
1,700 |
28.30 |
|
Total pesticides |
30,190 |
31,015 |
30,900 |
−0.37 |
|
Total transgenic crops |
75 |
235 |
650 |
176.60 |
|
Sources: Cultivar, 1997; Eppes, in press. |
||||
Poulenc to form Aventis (about the size of Novartis), and Novartis and AstraZeneca have merged to form Syngenta.
Another major trend in the market is the rapid growth of biotechnology products compared with chemical pesticides, stimulating the vertical integration of agrichemical companies with seed and food companies. These biotechnology products, chiefly transgenic-crop seed with pest-control attributes and herbicide tolerance, are dramatically changing market shares of agrichemical firms in soybeans (herbicide tolerance), cotton (insecticides for bollworm and budworm control), and corn (herbicide tolerance and insecticides for corn-borer control). Table 4-6 shows the total sales of transgenic crops relative to chemical pesticides in 1995–1997.
Agrichemical companies have invested billions of dollars to develop or access crop seed genetically engineered for caterpillar and disease control and tolerance to herbicide sprays. Monsanto is the industry leader, with reported investment that totals more than $8 billion to acquire Calgene, Delta PineLand, Cargill, Ecogen, DeKalb, Agracetus, Asgrow, Holden's, and PBI. Other examples are Dow's purchase of Mycogen and Pioneer's $30 million deal with Mycogen to obtain nonexclusive access to Bt genes for genetically engineered corn seed. Novartis and Mycogen partnered to launch a transgenic corn seed containing a Bt gene for corn borer control. A strategic alliance composed of Mycogen, Rhone Poulenc, and Dow was formed to develop crops with input and output traits. Other shifts in the industry include the purchase of Plant Genetic Systems for $750 million by AgrEvo (Hoechst-Schering-Roussel), DuPont's purchase of 20% of Pioneer for $1.7 billion, and the acquisition of Mogen by AstraZeneca to complement its share of seed companies (now called Advanta).
There is still heavy reliance on traditional chemicals for weed control, in part because of the widespread reliance on genetically engineered glyphosate-resistant crops. Other companies still depend on agrichemicals for primary income. These same multinationals are also entering into agreements with food companies to develop crops with value-added traits. Many agrichemical companies are shifting resources away from inputs (pesticides) and input traits (pesticidal genes) to output traits.
Examples of these traits are
-
Improvements in feed value (for example, corn seed engineered with phytase enzyme or high levels of lysine, an essential amino acid).
-
Higher-quality product (for example, fresh tomatoes with longer shelf-life or paste tomato with lower water content).
-
Greater nutritional value (for example, rice with higher vitamin A content).
The industry is investing billions of dollars in genomics to characterize the genes of entire organisms. Industrial leaders expect that advances in genomics will lead researchers to the precise location and sequence of genes that contain valuable input and output traits.
A shift in R&D resources from input to output traits probably would have a large impact on the future of plant protection. Will the cycle of innovation on the input side continue? Because of the high investment required for development of chemical pesticides and transgenic crops, will large agrichemical and life-science firms focus primarily on crops with large markets (such as row crops)? Whether companies will develop pesticides and input traits for minor use crops remains an open question.
A trend in agrichemical industry is the movement of many chemical pesticides off patent. As these chemicals become generic pesticides, manufacturers lose their monopolies on them. Large agrichemical companies are therefore aligning themselves with generic suppliers of chemical pesticides to reduce erosion in sales of the products that were formerly proprietary products. Several agrichemical companies have purchased outright or partially own generic companies. DuPont, for example, has entered into a joint venture and is now a 51% owner of Griffin. In May 1998, BASF purchased MicroFlo for the same reasons.
As more products become generic, profit margins erode for distributors, as well as for manufacturers. In response, distributors are consolidating and becoming “basic manufacturers” through acquisitions of proprietary products. For example, Gowan Corp., Griffin, and United AgriProducts (UAP) have taken this approach.
The consolidation of distributors can take two paths. In the first, a large distributor can acquire many smaller distributors. UAP, a practitio-
ner of this approach, is now as large as or larger than many basic manufacturing agrichemical companies and has revenues of $2 billion. The second path is the formation of consortiums of smaller distributors into a group with more influence than individual firms. In California, for example, a group of distributors joined to form a consortium called Integrated Agricultural Producers (IAP). The consolidation of distribution and agrichemical companies has created some concern for farmers. They are worried that they will have fewer choices and that prices will go up. However, the advent of the Internet is tearing down old structures, and the entire distributor-manufacturer relationship is expected to change for the benefit of farmers, who will be able to order all their products directly over the Internet.
One of the most important trends for agrichemical companies is the growing shift towards the development and registration of reduced-risk pesticides. In 1993, the Environmental Protection Agency (EPA) began a program of expedited review of what were classified as reduced-risk pesticides. Expedited reviews can reduce the time to registration by more than half (EPA 1998). Since the introduction of this program, the number registered as reduced-risk pesticides has steadily increased. Table 4-7 lists almost 20 reduced-risk pesticides that have been registered since 1994.
For a pesticide to be considered of reduced risk, it must have at least one or more of the following characteristics (EPA 1997a):
-
It must have a reduced impact on human health and very low mammalian toxicity.
-
It must have toxicity lower than alternatives (0.01–0.1 as much).
-
It displaces chemicals that pose potential human health concerns or reduces exposures to mixers, loaders, applicators, and re-entry workers.
-
It reduces effects on non-target organism (such as birds, honey bees, and fish).
-
It exhibits a lower potential for contaminating groundwater.
-
It lowers use or entails fewer applications than alternatives.
-
It has lower pest-resistance potential (that is, it has a new mode of action).
-
It has a high compatibility with integrated pest management (IPM).
-
It has increased efficacy.
The Food Quality Protection Act (FQPA) went further and mandated expedited registration of reduced-risk pesticides that could be expected to pose less risk to human health and the environment than other pesticides that meet existing safety standards. Since the enactment of FQPA, 62% of the 48 active ingredients registered have been considered “safer”
TABLE 4-7 Reduced-Risk Pesticides registered with US EPA since 1994
|
Chemical |
Pesticide type |
Use |
Year registered |
|
Acibenzolar-S-methyl |
Plant activator |
Tomato, lettuce, tobacco |
Pending |
|
Alpha-Metolachlor |
Herbicide |
All metolachlor uses |
1997 |
|
Azoxystrobin |
Fungicide |
Turf |
1997 |
|
Azoxystrobin |
Fungicide |
Almond, cucurbit, rice, wheat |
1997 |
|
Azoxystrobina |
Fungicide |
Grape, banana, peach, tomato, pecan, peanut |
1997 |
|
Bifenazate |
Insecticide |
Ornamentals |
1999 |
|
Cadre |
Herbicide |
Peanut |
1996 |
|
Carfentrazone |
Herbicide |
Wheat, corn |
1998 |
|
Cyprodinil |
Fungicide |
Stone fruit |
1998 |
|
Diflubenzurona |
Insecticide |
Below ground termite bait station |
1998 |
|
Diflufenzopyr |
Herbicide |
Corn |
1999 |
|
DPX-MP062 |
Insecticide |
Cotton, tomato, pepper, cole crops, lettuce, sweet corn, apple, pear |
Pending |
|
Fenhexamid |
Fungicide |
Grape, strawberry, ornamental |
1999 |
|
Fludioxonil |
Fungicide |
Corn |
1995 |
|
Fludioxonil |
Fungicide |
Various seed treatments |
pending |
|
Fludioxonil1 |
Fungicide |
Potato |
1997 |
|
Hexaflumuron |
Termiticide |
Below-ground bait station |
1994 |
|
Hexaflumurona |
Termiticide |
Above-ground bait station |
1997 |
|
Hymexazol |
Fungicide |
Sugar beets |
1995 |
|
Imazamox |
Herbicide |
Soybeans |
1997 |
|
Mefenoxam |
Fungicide |
Ornamentals, citrus, nuts, fruit trees |
1996 |
|
Methyl anthranilate |
Bird repellent |
Food crops |
1994 |
|
Pymetrozine |
Insecticide |
Potato, cucurbit, tomato, tobacco |
1999 |
|
Pyriproxyfen |
Insecticide |
Cotton |
pending |
|
S-Dimethenamid |
Herbicide |
Corn, soybean, peanut |
1999 |
|
Spinosad |
Insecticide |
Cotton |
1997 |
|
Tebufenozide |
Insecticide |
Walnuts |
1995 |
|
Tebufenozide |
Insecticide |
Pome fruit, cotton, leafy vegetables, cole crops, sugarcane, pecan, forestry, fruiting Vegetables, Ornamentals |
pending |
|
Trifloxystrobin |
Fungicide |
Pome fruit, grape, cucurbit, peanut, turf, banana |
1999 |
|
aNew uses of existing active ingredients. Source: adapted from Aspelin and Grube, 1999. |
|||
than conventional pesticides, and 77% of the new uses of pesticides have been considered “safer” (EPA 1999a.) In the year 2000, 10 of the 25 new chemicals seeking registration are classified as of reduced risk (EPA 2000). Those facts indicate that many of the new pesticides being registered meet one or more of the criteria stated above for reduced risk and that companies are responding to the expedited review of these pesticides by EPA. The consequence of this response is that future pesticides will pose even less risk to human health and the environment because they will be compared with pesticides that are now being registered as of reduced risk.
Biopesticide Companies
The most successful biopesticide is Bacillus thuringiensis. The market is dominated by Abbott (68% share), followed by Thermo Trilogy (26%), Ecogen (4%), and Mycogen (2%). Several small biopesticide companies (AgraQuest, BioWorks, Consep, Dominion Biosciences, Eden Biosciences and Mycotech) focus on biological pest control other than with Bt (table 4-5). New entrants in the last 5 years are AgBio Development, AgraQuest, BioWorks Inc., Dominion, and EcoSoils Systems. These companies focus primarily on minor-use crops, such as fruits, vegetables, wood products, and ornamentals. Some of the products that are getting a foothold in the marketplace are botanical extracts from the seed of the neem tree (NRC 1991), microorganism-based pesticides that include fungus-based insecticidees and fungicides, baculovirus, and beneficial nematodes. Many of the newer products are designed to prevent soil-based diseases from attacking crops and their roots (Dutky 1999). Because of the lower cost and shorter time to develop a biopesticide, as opposed to synthetic chemicals or transgenic crops, as seen in Figure 4-2, small biopesticide companies are able to enter this market. New startup firms develop alternative technologies for smaller markets that are neglected by larger agrichemical companies.
Use of Microbial Pesticides in Integrated Pest Management (IPM) Systems
The use of microorganisms as commercial pest-control agents was reviewed by Quarles (1996), Rodgers (1993), and Starnes et al. (1993). Bt by far has the longest history and is the most widespread microbial pesticide in use today. Strengths of microbial pesticides can include
-
Safety with respect to nontarget organisms.
-
Biodegradability.
-
Low cost of development.
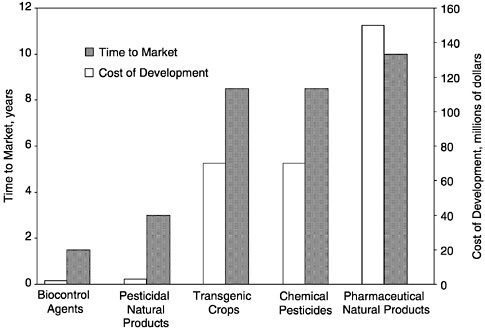
FIGURE 4-2 Cost to develop and time to market of various products.
-
Suitability for IPM systems.
Drawbacks of microbial pesticides can include
-
Short shelf-life of products.
-
Inconsistency of efficacy within and between ecosystems.
-
Short field residual life.
-
Difficulty of use.
Bacillus thuringiensis
Bt-based microbial pesticides are used extensively in vegetable production in California because of concern about chemical residues on harvested products. These pesticides are also used when there is a need to decrease selection pressure on diamondback moth and cabbage looper pests that evolves primarily from heavy applications of conventional pesticides in production of cole, lettuce, and tomato crops (Zalom and Fry 1992).
John Trumble and his colleagues at the University of California, Riverside have done considerable work on Bt in IPM vegetable programs
(Moar and Trumble 1987, Trumble 1985, Trumble 1990). Trumble reported higher profits in IPM plots in production of celery and tomatoes relative to traditional chemical-spray programs (Trumble 1989, Trumble 1991, Trumble and Alvarado-Rodriguez 1993).
The Hunt-Wesson and Campbell Soup companies have introduced IPM programs in a large tomato-growing area of Sinaloa, Mexico. Results of the field tests indicate increases in yield per acre, decreased costs per ton, and improvements in tomato quality (Moore 1991). Campbell Soup has reduced the amount of pesticides used by incorporating Bt and other nonchemical approaches into most of their vegetable growing (W. Reinert, University of California, Davis, April 8, 1997, personal communication).
Bt has been successfully applied to cruciferous crops for many years (Sears et al. 1983, Ferro 1993). The practicality of Bt for controlling cotton caterpillars—such as the Egyptian cotton leafworm, S. littoralis Boisduval and Helicoverpa spp.—has also been demonstrated (Broza et al. 1984, Daly and McKenzie 1986). With the advent of transgenic crops, these IPM systems are no longer useful. The diamondback moth, Plutella xylostella, which has evolved resistance to all chemical classes and also to Bt, can be managed with IPM (Metcalf 1989). In Taiwan, two larval parasitoids, pheromone traps, and Bt reduced pest population densities on cauliflower and broccoli to less levels than in neighboring, conventionally sprayed plots (Asian Vegetable Research and Development Center 1991).
A survey of tree-fruit researchers throughout the United States indicated that the integrated use of pheromone mating disruptants, low doses (one-tenth of the recommended label rate) of pyrethroids, and the full rate (1 lb.) of Bt is a useful IPM program for leafroller management (Tette and Jacobsen 1992). The peach twig borer (Anarsia lineatella Zeller) is a major pest of almonds in California that has been controlled through an IPM program (Dr. Frank Zalom, University of California, Davis, April 15, 1998, personal communication). As the organophosphate insecticides are phased out because of adverse effects on hawks in the almond orchards, Bt reportedly is an economical replacement for organophosphates.
Bt has been used operationally for controlling forest and tree caterpillar pests for many years (Bowen 1991, Cunningham 1988, Elliott et al. 1993). Bt kurstaki (Btk) is the most widely used insecticide for forest defoliators, such as gypsy moth, spruce budworm, western spruce budworm, forest tent caterpillar, fall cankerworm, and hemlock looper. Improving the efficacy of Bt sprays continues to be a subject of research, but Btk is a successful stand-alone product and already has replaced established insecticides—such as carbamates, organophosphate, and pyrethroid pesticides—that were once considered essential to the system. The displacement occurred because of a combination of efficacy, economics, and system-specific attributes.
Bt tenebrionis (Btt), active on coleopteran pests, has been shown to control Colorado potato beetle under field conditions (Ferro and Gelernter 1989, Ferro and Lyon 1991, Jaques and Laing 1989) and is now used commercially in IPM systems. Roush and Tingey (1991) reported the development of an IPM system for Colorado potato beetle in New York. The program includes scouting, border sprays, or barriers to trap migrating overwintering adults and sprays of Btt on small larvae, cryolite on large larvae, and endosulfan on adults. The system enhances predators and manages pesticide resistance in potatoes through the use of “soft” chemicals and Btt, and rotation of pesticides in different chemical classes.
Although Btt has commercial promise for control of Colorado potato beetle, the advent of the pesticide imidacloprid has reduced the use of Btt substantially. Imidacloprid can provide season-long control when applied as a potato seed treatment, whereas Btt must be applied weekly when there are small larvae. Therefore, the chemical is easier and more effective and has become the product of choice for most potato growers. However, Colorado potato beetle has become resistant to all chemicals used against it and is expected to develop resistance to imidacloprid as well.
The discovery of the beetle-active Btt fueled the search for strains with novel activity. Companies in the Bt arena boast collections of several thousand Bt isolates. Many new crystal types have been discovered with activity against nematodes, mites, corn rootworm (Diabrotica spp.), adult flies, and ants. The potential for use of these new Bt strains in IPM is unknown. Most of the attention is on finding new genes that can be engineered into plants.
Ecogen and Novartis scientists have used various molecular techniques (including electroporation and transconjugation) to develop products that combine genes from aizawai and kurstaki strains to increase activity against key lepidopteran pests, such as armyworm (Spodoptera spp.) and cotton bollworm (Helicoverpa spp.). Novo Nordisk used classical mutation to improve Btt. This strain produces a larger crystal and crystal size correlates directly with field activity. In addition, fusion of genes from Bt into baculovirus is being used to expand the host range of Bt. Ecogen has introduced CryMax, which is a Bt strain engineered to contain multiple copies of a Btk protein, improving efficacy on target caterpillar pests.
It is well known that Bt remains active against the pest for only several hours on plant foliage under typical field conditions because of UV degradation, rainfall, and other environmental perturbations. In the early 1980s, Monsanto developed a recombinant plant-colonizing pseudomonad for delivery of Bt genes, with the objective of improving residual activity and efficacy of Bt proteins. This concept was developed by Mycogen into the products MVP™ and M-Trak™. The Bt-bearing pseudomonad is killed (to avoid regulatory hurdles for registering recombinant
microorganisms) and sprayed on the crop as other Bt products are. The pseudomonad cell is reported to protect the Bt protein from environmental degradation, thus providing longer residual activity. These Bt products have had modest commercial success. A starch-encapsulation procedure for virus and Bt designed to improve the survival and efficacy of these microbial products in the field is under development by the US Department of Agriculture (USDA) Agricultural Research Service (ARS) in Peoria, Illinois.
Novo Nordisk discovered an enhancer of Bt, a natural substance produced in the Bt at a very low concentration (Manker et al. 1994, 1995). The same natural product was isolated previously by University of Wisconsin researchers as a fungicide, which they called zwittermicin A (He et al. 1994). When the compound is combined at higher concentrations with the Bt protein, efficacy against the most refractory caterpillars, such as Helicoverpa zea and Spodoptera exigua, is increased substantially in the field.
New genetically engineered and improved Bt products might provide more opportunities and choices for growers who use IPM programs. The most successful Bt products are ones that provide efficacy, ease of use, and consistency approaching traditional chemical pesticides. Improved armyworm ( Spodoptera) and bollworm (Helicoverpa) products are the most important developments in the use of Bt microbials in agriculture. Bt products could capture a larger market share and replace some established chemical products. In any case, Bt strains probably will remain important replacements for chemical insecticides in fruit, vegetable, and forestry IPM systems.
Baculoviruses
Although they have not had the commercial success of Bt, baculoviruses could have important potential for use in IPM programs. They have a number of advantages. Baculoviruses are ideal for IPM because as far as is known, they are safe for nontarget insects, humans, and the environment. Baculoviruses might, in some cases, be the only effective biocontrol agents available for controlling insect species (Cunningham 1988) and they provide an avenue for overcoming specific problems, such as resistance. It is important to have a selection of control agents when designing pest-management strategies. Because viruses are not likely to elicit cross resistance to chemicals, they should receive more attention from university and industrial researchers (Cunningham 1988). The use of multiple biological products has the advantage of lowering the potential for evolution of pest resistance. Although practical applications of viruses are beginning to develop in IPM programs in US agriculture, the
long-term value of viral approaches for widespread use has not been demonstrated.
Helicoverpa zea Boddie (cotton bollworm) nuclear polyhedrosis virus (NPV) was the first baculovirus to be marketed in the United States. It was developed by International Minerals and Chemical Corporation (IMC) but marketed by Sandoz under the tradename Elcar in 1976 after purchase of IMC 's biological products division. Interest in Elcar declined with the introduction of pyrethroids, which are effective, inexpensive broad-range insecticides. The properties of viruses suitable for IPM systems have been well studied (Ignoffo and Garcia 1992).
In Europe, a number of companies—including Kemira Oy (Finland), Oxford Virology (United Kingdom), and Calliope (France)—have introduced viral products for the insecticide market or are developing them. Viral products include Cydia pomonella L. (codling moth) granulosis virus (GV), Neodiprion sertifer (Geoffrey)(European pine sawfly) NPV, Spodoptera exigua (Hübner)(beet armyworm) NPV, and Autographa californica (Speyer) (alfalfa looper) NPV. The largest use of baculoviruses is in Brazil, where Anticarsia gemmatalis Hübner (velvetbean caterpillar) NPV protects 5.9 million hectares of soybeans against the velvetbean caterpillar.
In North America, the effort with baculoviruses has been led mainly by government agencies (Cunningham 1988, Podgwaite et al. 1991, Otvos et al. 1989). The US Forest Service (USFS) has registered NPVs to control Lymantria dispar (L.) (gypsy moth), Neodiprion sertifer, and Orgyia pseudotsugata (McDunnough) (Douglas fir tussock moth) in forestry. The Canadian Forest Service holds registrations for O. pseudotsugata NPV and Neodiprion lecontei (Fitch) (redheaded pine sawfly) NPV. US companies actively involved in baculovirus research are American Cyanamid, Thermo Trilogy, and DuPont.
Louis Falcon (University of California, Berkeley) has demonstrated the successful use of codling moth granulosis virus in pear, apple, and walnut IPM systems in California and Washington (personal communication, May 12, 1991). Organic growers pay $30 per acre-treatment of virus and make applications five to 10 times per season, in contrast with conventional growers who spray monthly (three times per season) the organophosphate insecticide Guthion™ at $7.50-10.00 per acre-treatment. According to Falcon, the cost of total chemical inputs (insecticides, fungicides, acaricides, bactericides) is approximately $360 per season and the total cost of all pesticides (fungicides, miticides, insecticides), including the virus for codling moth is $316 per season. In the virus-treated orchards, natural enemies can survive to control mite pests, thus eliminating the need for miticides, which are required in Guthion-treated orchards. Although Dr. Falcon's program has been successful for organic growers, mainstream fruit producers have not switched to it, because
Guthion™ is an effective alternative. With the recent development of Guthion-resistant codling moth populations after 20 years of use and restriction of the preharvest interval for Guthion by the California Department of Pesticide Regulation, growers have more incentive to adopt the virus-pheromone IPM program.
Application of baculovirus for control of beet armyworm, Spodoptera exigua (Hübner) has been well studied in greenhouse systems (Smits et al. 1987a). Such characteristics as dose-response curves, larval feeding behavior, application techniques (Smits et al. 1988), timing, and strain (Smits et al. 1987b) have been integrated into recommendations for operational use of virus with other control methods. On head lettuce in California, a beet armyworm NPV was field-tested for 3 years and compared with chemical insecticides (Gelernter et al. 1986); results indicated comparable control with methomyl and permethrin.
At the present stage in the development of baculovirus products, several limitations are associated with the viruses' use as insecticides. A major limitation in agricultural systems is the slow rate of kill, which results in feeding damage. Kill rate, however, is not as crucial in forest systems, where cosmetic damage is not as important. Reduction of kill time will rely on improvements in formulation and application in the immediate future. However, this limitation can be managed in the short term by using baculoviruses in combination with other insecticides through IPM.
Lower production costs are essential for both recombinant and wild-type baculoviruses to compete with classical insecticides. There are active research programs in both in vivo and in vitro production (Bonning 1996). Although viruses are less expensive to produce in vivo than in vitro, the cost still exceeds that of Bt. Viruses are formulated to be applied in the same fashion as Bt strains. However, for extensive use in IPM, dramatic improvements in formulation and application technology are needed. In formulation, knowledge of stability and shelf-life is required to optimize storage and distribution. In application, droplet size, density, dosage, and components in the tank mix (for example, stickers, and UV protectants) need to be optimized.
Another limitation of baculoviruses is their host specificity, which can reduce their commercial potential. However, the host specificity is viewed positively from the environmental and IPM standpoints. Two viruses with relatively broad host ranges are Autographa californica (alfalfa looper) NPV and Syngrapha falcifera (Kirby)(celery looper) NPV, each of which kills over 30 insect species. The celery looper virus is reported to have commercial potential in cotton IPM systems (Wood 1992). Host range can be broadened through molecular means or by mixing two viruses.
In the long term, the development of recombinant baculoviruses that
can kill rapidly will allow them to compete more effectively with classical pesticides. To increase the ability of baculoviruses to kill early, research to insert specific genes into the baculoviral genome is under way. These genes will serve as toxins or disrupters of larval development. Among the proteins being tested for exploitation are Bt endotoxin (which failed to improve the virus), juvenile hormone esterase, prothoracicotropic hormone (PTTH), melittin, trehalase, scorpion toxin, and mite toxin (Bruce Hammock, University of California, Davis, November 12, 1998, personal communication). The knowledge of the molecular biology of viruses has also promoted interest in modifying and improving baculoviruses with regard to host range and virulence.
The regulatory process that will be applied to recombinant baculoviruses is not yet clear. The recombinant virus system makes it possible to exploit a variety of proteins, including insect enzymes and hormones and proteins from other organisms. The recombinant viruses that will probably be commercialized first in the United States are the ones that carry genes expressing insect-selective nerve toxins, which are undergoing intensive safety and efficacy testing. In the United States public concern has not been voiced with regard to the safety of these viruses. However, objections have been raised in England as to the use of toxin genes in baculoviruses.
The growth and success of baculoviruses as commercial insecticides will depend on reducing production costs, developing practical and effective formulations, optimizing field performance, overcoming regulatory obstacles, and educating users and the public on their safety. To enable development of more economical and effective products, R&D efforts should focus on making improvements in baculovirus production, formulation, and application technologies in conjunction with genetic engineering of the viruses to enhance their kill rates and broaden their host range.
Entomopathogenic Fungi
Over 500 fungi are regularly associated with insects; some cause serious disease in their hosts, but few have been used commercially as control agents. Because of their dependence on specific environmental factors, such as relative humidity, fungi can be useful tools in IPM, especially as complements to other products. Fungi infect a broader range of insects than do other microorganisms, and infections of lepidopterans (moths and butterflies), homopterans (aphids and scale insects), hymenopterans (bees and wasps), coleopterans (beetles), and dipterans (flies and mosquitoes) are quite common. In fact, some fungi have very broad host ranges that encompass most of those insect groups. That is true of Beauveria
bassiana (Balsamo) Vuillemin, Metarhizium anisopliae (Metschnikoff) Sorokin, Verticillium lecanii (Zimmerman) Viegas, and Paecilomyces spp., all of which have worldwide distributions; these are the most commonly used insect pathogens developed for commercial pest-management products (McCoy 1990).
B. bassiana has been identified in many insect species in temperate and tropical regions and is used for pest control on a moderate scale in eastern Europe and China. Mycogen produced a B. bassiana-based bioinsecticide, which has been shown to be highly pathogenic in coleopterans. The fungus also is amenable to mass production of conidia by semi-solid fermentation. The product has been field-tested against citrus root weevil. Another B. bassiana strain, researched at the USDA ARS shows good control of rasping or sucking insects, such as thrips, whiteflies, and aphids.
M. anisopliae has been most extensively used in Brazil for control of spittlebugs on sugar cane. Using Metarhizium as the control agent, EcoScience Laboratories, Inc. has developed infection chambers in which insects (cockroaches and flies) brush against spores of the pathogen, which later germinate and infect the insect. However, this product has not been successfully commercialized.
V. lecanii is a pathogen that has demonstrated good control of greenhouse pests, such as Myzus persicae (Sulcer) aphids, on chrysanthemums. A distinct isolate of V. lecanii was obtained from whitefly and provided excellent control of greenhouse whitefly, Trialeurodes vaporariorum (Westwood), and of Thrips tabaci Lindeman on cucumber. V. lecanii was produced commercially as Vertalec for aphid control and Mycotal for control of whitefly from 1982 to 1986, and there is a resurgence of commercial interest in its use for control of aphids, whiteflies, and thrips because these greenhouse pests have developed resistance to chemical pesticides typically used for their control.
The fungal pathogen Entomophaga maimaiga has been recognized and used by USFS, states' departments of natural resources, and university personnel as a control for gypsy moth (Elkinton et al. 1991). Since its reappearance in the early 1990s, this pathogen largely has become self-perpetuating. It should be noted that public research agencies have played an important role in the development of gypsy moth pest-management strategies. In fact, the use of Entomophaga maimaiga with Bt and NPVs exemplifies the trend toward biologically based pest management in gypsy moth control in the last 10 years.
The effectiveness of fungi in controlling insect pests depends on the environmental conditions prevailing after application, particularly with respect to relative humidity. There is a need for research to develop moisture-retaining formulations that allow fungal growth at suboptimal rela-
tive humidity. Innovative biotechnology can also be used to engineer desirable traits into fungi and thus improve the effectiveness of some fungal pathogens. Transformation systems and recombinant-DNA techniques are now being used to study the mechanisms of pathogenicity and virulence at the molecular level (Carruthers and Hural 1990). Eventually, all this knowledge will enhance the ability to manipulate the genetics of these organisms.
Microbial and Natural-Product Fungicides
The use of microorganisms as fungicides was reviewed by Quarles (1996). There are many commercial microbial biopesticides for controlling plant pathogens, but they make up an insignificant portion of the fungicide market. Commercial adoption is hampered by the inconsistency of microbial fungicides in the field. However, as tools in IPM systems, microbial fungicides (Box 4-1) can be used in rotation with chemical products or stand-alone in conjunction with disease-forecasting models.
Natural products are organic compounds produced by microorganisms, plants, and other organisms. There are few natural-product fungicides for agriculture (Franco and Coutinho, 1991). One is validamycin, isolated from the bacterium Streptomyces hygroscopicus var. limoneus and used to control rice sheath blight (Yamamoto 1985). Extensive university research is focused on microbial control of plant pathogens (Adams et al. 1990, Taylor and Harman, 1990 Weller 1988). It has adressed microbial physiology or microorganism-plant interactions, especially for root patho-
|
BOX 4-1 Microbial Fungicides
Source: Quarles, 1996. |
gens. However, little is known of the identity of the plant pathogeninhibiting natural products produced by these microorganisms and it is an important subject for future research.
The use of Bacillus subtilis as a fungicidal treatment has been demonstrated on a number of diseases, including cornstalk rot (Fusarium roseum) (Kommedahl and Mew 1975), onion white rot (Sclerotium cepivorum) (Utkehede and Rahe 1983), potato charcoal rot (Macrophomina phaseolina) (Thirumalachar and O'Brien 1977), bean rust (Uromyces phaseoli) (Baker et al. 1985), apple blue mold (Penicillium expansum) (Sholberg et al. 1995), and peach brown rot (Monilinia fructicola) (Gueldner et al. 1988). The vast majority of the work with Bacillus subtilis has concentrated on treatment of seeds or soil to control pathogens; in general, the use of biocontrol as a foliar treatment is much less developed than in the soil-rhizoplane environment (Blakeman and Fokkema 1982). Foliage has been sprayed with strains of Bacillus spp. for plant-pathogen control. For example, Sharga (1997) screened 270 bacilli and found two that provided good in vitro control of Botrytis cinerea and B. fabae. However, once these microorganisms were applied in the field, a rapid decline in the population of introduced bacilli was observed within 24 hours and was concomitant with loss of protection against Botrytis. Sharga concluded that the protection observed in greenhouse tests was due to intense competition for exogenous nutrients between Bacillus and Botrytis. In a second study, Baker et al. (1985) sprayed bean cultivars in the field with Bacillus subtilis and observed substantial control of rust (Uromyces appendiculatus) for plants sprayed three times a week.
Bacilli are known to produce antifungal and antibacterial secondary metabolites (Korzybski et al. 1978). University of Wisconsin and Cornell researchers have identified a novel fungicidal compound, zwittermicin A, produced by Bacillus spp. (Stabb et al. 1994, He et al. 1994). The compound substantially inhibits elongation of germ tubes from Pythium medicaginis cysts (Silo-Suh et al. 1994). A second fungicidal metabolite produced by the same strain was recently identified as the known amino sugar kanosamine (Milner et al., 1996). Wisconsin field data on control of below-ground root diseases for soybean with the Bacillus strain are promising (Osburn et al. 1995). However, only one report demonstrated its use as an above-ground treatment; Smith et al. (1993) described suppression of cottony leak on cucumbers using B. cereus strain UW85.
Another group of previously described metabolites of Bacillus is the cyclic lipopeptides of the iturin class, some of which are potent fungicidal agents. These compounds consist of a cyclic octapeptide with seven alpha-amino acids and one beta-amino acid with an aliphatic side chain. There are several groups of iturins that differ in order and content of the lipopeptide side chain. Generally, a suite of related molecules is produced
with differences in the length and branching of the aliphatic amino acid residue. When tested against yeast, Saccharomyces cerevisiae, mycosubtilin was most active (LC50 = 10 µg/mL), followed by iturin-A and bacillomycin L (both 30 µg/mL) (Besson et al. 1979). Iturin-C is inactive against fungi, including Penicillium chrysogenum (Peypoux et al. 1978).
Researchers at USDA ARS have investigated the structure-activity relationship of the iturins by synthesizing a number of analogues differing in amino acid chain length. They reported that the activity of the iturins increased with the length of the fatty acid side chain and the terminal branching in the order iso > normal > ante-iso (Bland et al. 1995) and that the “amounts of iturins obtained from natural production are inadequate to be commercially viable” on the basis of their work with a number of iturin-producing strains of Bacillus. However, only a small number of potential Bacillus strains have been screened for fungicidal activity. AgraQuest is developing a naturally occurring Bacillus subtilis, which has shown excellent activity for some foliar and fruit diseases. Investigators isolated it in a screening program directed at new strains with superior antifungal activity.
EcoSoils Systems, San Diego, California, has a unique approach to address the short-shelf-life problem of many microbials, such as Pseudomonas-based biofungicides. Industrial scientists ferment on-site continuously for control of turf diseases on golf courses. The company is applying the Bioject system to agricultural crops, especially for control of root rot diseases, such as Phytophthora root rot of avocado.
Natural products are another major source of new leads for pesticides. Most companies generate natural-product extracts either internally or from external sources and test them against test organisms or in mechanism-based screens. There are several advantages in using natural products directly as pesticides or as leads for new pesticides:
-
Large diversity of chemical structures.
-
High potential for finding new mechanisms of action.
-
Lower risk to environment.
-
Lower risk of toxicity to nontarget organisms.
Many of the new chemistries in fungicides, insecticides, and herbicides can trace their origins to natural products. For example, glufosinate is a derivative of bialophos, a natural product derived from Streptomyces viridachromogens (Leason et al. 1982). The methoxy-acrylate fungicides arose from strobilurin A, a natural product extracted from a mushroom (Clough et al. 1992). The active ingredient in the new spinosad insecticides used in cotton is isolated from the naturally occurring soil organism Saccharopolyspora spinosa (Thompson et al. 1994, Adan et al 1996, EPA
1997b). The new pyrrole insecticides originated in dioxypyrrolomycin, a natural product derived from an extract of Streptomyces spp. (Black et al. 1994).
Hypovirulence
Another potential use of microorganisms to protect plants is through hypovirulence—infection of a plant with benign strains that protect hosts from later infection by virulent strains. Hypovirulence is caused by a double-stranded RNA virus that infects the fungus, and reduces its virulence relative to the crop plant. The hypovirulent state can be induced through infection with viruses and related materials. Hypovirulence has achieved its widest success in combating chestnut blight, a disease that radically altered the eastern forest landscape after introduction of an invasive fungus (NRC 1996).
Transgenic Crops
Choice of crop variety has always been a cornerstone of crop protection, especially for disease and insect control. There are numerous review articles on plant resistance to insects (Harris 1980, Kogan 1982, Hedin 1983). The sources of resistance to pests in crops have been classified as nonpreferred, antibiosis, and tolerance (Pfadt 1971). Insect preference for a host plant is related to physical structure of the surface (hairs, wax, and so on), color, taste, odor, and light reflection. A resistant variety might be nonpreferred by virtue of lacking one or more of the preferred factors or characteristics of the host plant. Antibiosis is the adverse effect of the plant on the pest, which can be due to a deleterious chemical or the lack of specific nutrients in the plant. Tolerance is the ability of certain plants to withstand pest attack by virtue of general vigor or ability to repair tissue damage caused by a pest attack.
Plant resistance as a pest-management factor has achieved some outstanding results (for example, against grape phylloxera, woolly aphid, Hessian fly, and wheat stem sawfly) (Kogan 1982). Desirable features of pest-resistant plants are specificity, cumulative effectiveness (with effect on the pest compounded in successive generations), and persistence and harmony with the environment, ease of adoption, and compatibility with other IPM tactics (Kogan 1982). Disadvantages of using pest-resistant plants for pest management include the long development time (3-15 years), genetic limitations (due to lack of available resistance genes), and evolution of pests that have overcome and are no longer controlled by the bred-in resistance. To overcome some of the problems associated with traditional resistance-breeding approaches, transgenic technologies have
been adopted. Inserting resistance genes from other genomes into a crop plant can reduce the problems associated with a lack of available resistance genes.
At present, resistance is the predominant defense against several plant diseases, such as rust diseases, that would otherwise reduce cereal-crop production in much of the world. In the case of rust diseases, a plant cultivar generally is resistant to only one race of a pathogen. Other races of the pathogen can infect the plant, and the shift in the distribution of the pathogen races leads to a boom-and-bust syndrome of the rust diseases. Strategies of resistance-gene deployment in which multiple cultivars (with different race-specific resistance genes) or single cultivars with multiple race-specific genes are planted in fields have been found to be effective against the syndrome. Race-specific resistance genes deployed in this manner can be quite successful in controlling plant diseases (NRC 1996).
Crop seed engineered with a single gene for virus or insect control or herbicide tolerance is commercially successful (Table 4-3). In 1998, more than 5 million acres were planted with cotton seed engineered to contain a Btk gene for caterpillar control; in the same year, 1 million acres were planted with dual-trait cotton seed (engineered to contain Bt and a gene expressing herbicide tolerance). Potato containing a Btt gene for controlling Colorado potato beetle is also on the market. Cotton, corn, and soybean crops engineered to resist the herbicide Roundup (were launched during the last 2 years; Roundup Ready (soybean seed was planted on 9 million acres in 1997. Also, Liberty Link (canola and corn seed engineered to resist glufosinate are on the market. Industrial leaders anticipate that plants containing chitinase genes and other genes for fungal and bacterial plant-pathogen control will be on the market in the near future; many of these newer genetically engineered plant varieties are already undergoing field tests (Figure 4-3).
Transgenic plants, whether engineered to contain an insecticidal protein, such as an endotoxin protein from Bt or a chitinase gene to control root rot pathogens, appear to have the same advantages as traditionally bred pest-resistant crop varieties. For example, cotton engineered with a Bt Cry IA(c) or (b) protein
-
Is selective to lepidopterans.
-
Increases the persistence of Bt for season-long control.
-
Is compatible with the environment (reducing the use of more toxic chemical pesticides).
-
Is compatible with other pest-management tactics, such as use of natural enemies or chemical pesticides, aimed at other pests.
In general, transgenic approaches also have the same disadvantages
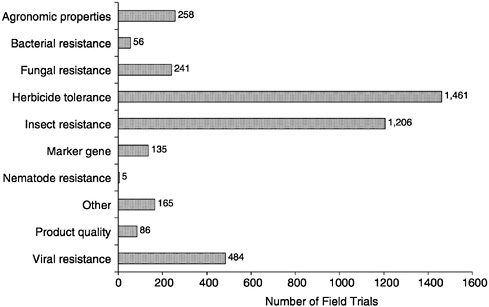
FIGURE 4-3 Agricultural-environmental biotechnology modifications, 1987-1998.
as traditionally bred crops. First, the period needed for development of transgenic plants—for such steps as finding appropriate genes, tissue-culture selection process, and backcrossing—is as long as traditional plant-breeding methods (John Callahan, AgraQuest, Inc., personal communication, May 22, 1998). Second, although much progress has been made in the discovery of new genes for introduction into plants, the ability to introduce the genetic material has surpassed the ability to discover new genes to engineer. Very few, if any, genes have been found for control of nematodes, sucking insects, and mites. In cotton, for example, transgenic plants with Bt genes will provide growers with another alternative for bollworm, but the need for products that control whiteflies, mites, and lygus bugs remains. Cholesterol oxidase, an enzyme from the microorganism Streptomyces and discovered by Monsanto Company, is toxic to cotton boll weevils and has been engineered into cotton plants; the enzyme is not effective on other insect pests.
The current choices of transgenic plants that contain single Bt genes are “first-generation” plants and will be followed by more sophisticated “second-generation” and “third-generation” plants with greater flexibility for use in IPM systems. These advances could include plants with inducible and tissue-specific expression systems whereby expression of the protein is “turned on” in response to insect feeding or some other stimulus. Also on the horizon are crops engineered with multiple genes
(“stacking”). This new advance has been undergoing extensive field trials over the last few years. Table 4-8 shows the number of field tests of genetically engineered crops containing one or more genes in 1987–1998.
For example, companies have already introduced crops stacked with Bt and herbicide tolerance. Crops with multiple genes for proteins with different modes of action (for example, gene 1 containing Bt is combined with gene 2 containing a protease inhibitor) will be useful to prevent or delay development of resistance by a pest. Some protease inhibitor and lectin genes have been engineered for caterpillar control. Macintosh et al. (1990) discovered that some protease inhibitors can be combined with Bt proteins for enhanced caterpillar kill.
A great deal of research has focused on the search for new genes for plant-pathogen control. In the next 10 years, agriculture will see the introduction of plants engineered to inhibit fungal and bacterial plant pathogens. Virus-resistant crops are already on the market.
Another limitation of both traditionally bred and transgenic crops is the evolution of pests that are unaffected by the bred-in resistance (the pest develops resistance to the plant resistance traits). This is a key issue for crops engineered with single-gene pest-control traits, such as Bt. Concerns about adoption of transgenic plants without IPM was expressed by Gould (1988), who wrote that, “the successful engineering of highly resistant crops could lead to the elimination of IPM techniques that aim at using intense, pest suppressive measures only when pests are likely to cause economically important damage.” Considering the advantages and the cost and time needed to develop new transgenic crops, it is important to sustain the life of these crops for a long period. Strategies designed to limit the development of resistant pests on the basis of understanding of pest population dynamics (such as studies on pest population establishment and growth, genetics, movement, behavior, number of generations required to develop resistance, and generation time) are critical for optimal and sustained use of transgenic plants in IPM systems.
Much research is still needed to develop effective resistance-management strategies and IPM systems incorporating Bt plants. There is a need for more research on pest genetics, resistance mechanisms, cross resistance, pest behavior and biology, and practical aspects of the implementation of refuges (for maintenance and development of susceptible insects) and areawide research on pest population levels and on detection and monitoring of resistance. The absence of USDA funding and coordination of public and private research on Bt-resistance management limits progress on resistance management. McGaughey et al. (1998) state that, “the challenge to US agriculture is to find ways to design and implement resistant management programs that will allow a realization of the benefits of this new agricultural biotechnology delivery system, while at the
TABLE 4-8 Number of Field Tests of Genetically Engineered Crops Containing Single or Multiple Genes
|
No. of Field Tests |
|||||||||
|
No. of Genetic Interventions |
|||||||||
|
Year |
One |
Two |
Three |
Four |
Five |
Six |
Seven |
Confidential Business Information |
Total |
|
1987 |
5 |
– |
– |
– |
– |
– |
– |
– |
5 |
|
1988 |
16 |
– |
– |
– |
– |
– |
– |
– |
16 |
|
1989 |
27 |
3 |
– |
– |
– |
– |
– |
– |
30 |
|
1990 |
36 |
12 |
1 |
1 |
– |
– |
– |
– |
50 |
|
1991 |
77 |
10 |
2 |
1 |
– |
– |
– |
– |
90 |
|
1992 |
131 |
20 |
3 |
5 |
– |
– |
– |
– |
159 |
|
1993 |
254 |
38 |
6 |
4 |
– |
– |
– |
– |
302 |
|
1994 |
505 |
49 |
16 |
– |
2 |
1 |
– |
– |
573 |
|
1995 |
629 |
63 |
14 |
4 |
4 |
– |
– |
– |
714 |
|
1996 |
501 |
88 |
10 |
1 |
8 |
1 |
2 |
611 |
|
|
1997 |
588 |
146 |
9 |
10 |
6 |
1 |
– |
2 |
762 |
|
1998 |
764 |
250 |
15 |
24 |
5 |
3 |
– |
2 |
1,063 |
|
Total |
3,533 |
679 |
76 |
50 |
25 |
5 |
1 |
6 |
4,375 |
|
Source: Stewart, in press. |
|||||||||
same time not jeopardizing the long-term benefits of this environmentally useful insecticide. ” Bt has been used for more than 30 years and is the most successful biopesticide ever developed. It is important to sustain its use as a biopesticide spray.
Genetic Engineering of Pests
Most research aimed at pest management based on genetic engineering has focused on genetic manipulation of crop plants, economically important tree species (Carozzi and Koziel 1997; Raffa 1989), and biocontrol agents (Hoy et al., 1997, Bonning and Hammock 1996). Less emphasis has been placed on engineering of weeds, pathogens, and insect pest species in ways that would decrease economic damage. However, the recent successes in stable transformation of mosquitoes based on the use of Hermes and Mariner transposons (Coates et al. 1998) has provided more impetus for genetic engineering of pests (Kidwell and Wattam 1998). The concept of modifying pests by use of classical genetic manipulations dates back at least to the 1940s, when Serebovsky (1940) and Vanderplank (1944) suggested that chromosomal abnormalities and hybrid sterility could be used for insect control. The most heralded success in using genetics for pest control is the eradication of the screwworm from areas of the United States (Baumhover 1966), Mexico (Krafsur et al. 1987), and Libya (Vargas-Teran et al. 1994) through release of irradiated, sterile insects. The genetically damaged males released in these programs mated successfully with native females, which laid eggs that could not develop properly. Repeated releases finally resulted in population extinction.
A considerable theoretical and empirical knowledge base about sterile-release techniques developed in the 1970s and 1980s (Asman et al. 1981, Whitten and Foster 1975; Davidson 1974, Prout 1978, Foster et al. 1988). More sophisticated genetic manipulation approaches, such as the use of conditionally lethal genes (Davidson 1974) and chromosomal translocations (Asman et al. 1981), also received attention during that time. Conditional lethals could spread into populations during favorable times but could induce a genetic load if they inhibited proper diapause initiation or hindered survival at high temperatures. Release of strains with single or double translocations could impose a genetic load on a native population while replacing native genes with those of the released strains. At least theoretically, a population of malaria-vector mosquitoes or plantpathogen-vector planthoppers could be replaced with an artificially developed nonvector strain (Curtis 1968). Because of the lack of enough simple success stories, support for this general kind of pest management declined in the late 1980s and early 1990s (but see Kerremans and Franz
1995). Nevertheless, the early theoretical and empirical studies determined the limits and potential of this species-specific control technology.
In the 1990s, entomologists and molecular biologists recognized that transposable elements, such as the P element in Drosophila, could be used in a manner similar to, but more efficient than, the use of translocations. That spurred new theoretical and empirical studies (for example, Curtis 1992, Ribeiro and Kidwell 1994, Meister and Grigliatti 1994). Although replacement of a native population with insects that had translocations required release of huge numbers of laboratory-reared insects (at least as many as in the native population if there was one translocation and no relative fitness reduction), it seemed theoretically possible to release a relatively small number of insects that had transposable elements (about 10% of the native population) to replace the native insect strain (Kidwell and Ribeiro 1992; Ribeiro and Kidwell 1994). A number of laboratory experiments with Drosophila have shown that a new P element introduced into a population spreads rapidly and is usually fixed in the population within 10 generations (Good et al. 1989). An important experiment by Meister and Grigliatti (1994) demonstrated that, when an alcohol dehydrogenase gene was artificially added to a P element, the P element was still able to invade a Drosophila population. Furthermore, the dehydrogenase gene was active in a number of cases.
If a transposon “loaded” with a gene for vector incompetence (inability to transmit malaria) (Olson et al., 1996) could be used to transform Aedes aegypti, the transformed strain could be released into the wild, and presumably the native malaria-transmitting strains of the mosquito would become refractory (Crampton et al. 1990, Hastings 1994, Curtis 1994, Pfeifer and Grigliatti 1996). The same approach could be taken for insect-transmitted plant diseases. This approach has not been without its critics, especially in the case of malaria control (Spielman, 1994, Pettigrew and O' Neill 1997). Some of the major concerns are about getting enough transmission reduction to substantially decrease disease severity (Spielman 1994); about the fact that a transposon that has invaded a native population cannot be used again; and about the potential for horizontal transmission of transposons between pest and nonpest species of animals (Robertson and Lampe 1995).
Another newly proposed approach for driving desirable genes into native populations involves use of rickettsial symbionts in the genus Wolbachia (Werren 1997). When a male insect carrying Wolbachia mates with a female that does not carry Wolbachia, no offspring are produced because of cytoplasmic incompatibility. The reciprocal cross is fertile, so the insects bearing the microorganism spread through the population (Sinkins et al. 1995). Because the Wolbachia is maternally inherited, any other maternally inherited trait present in the initial Wolbachia-carrying
insects will “hitch-hike” to fixation. Strategies have been developed that involve use of strains with Wolbachia and a virus that carries a gene for disease refractoriness (Sinkins et al. 1997). Other strategies involve isolation of the Wolbachia genes that code for cytoplasmic incompatibility; these genes are used to transform insect strains directly (Sinkins et al. 1997).
In addition to its use to develop novel control strategies based on transposons and microorganism-based incompatibility, genetic engineering could be used to improve the efficiency of classical genetic control strategies, such as introduction of conditional lethal genes or sex-ratio distortion genes (Gould and Schliekleman, in prep.). Researchers using classical genetic techniques have been severely limited in their ability to find genes that confer conditional lethality or sex-ratio distortion. Each time a project is developed for a specific pest, a new search must be undertaken to find useful genes or translocations. With the tools of genetic engineering, promoters and useful genes could be found that would be effective in many species. Once transformation of diverse insect species becomes more routine, it will be possible to transform a variety of species with appropriate genes.
Whether genetic engineering can be used for the direct control of pest species is an open question. In many cases, it seems easier to engineer crop plants for insect and disease resistance than it is to engineer the pest or vector. Most genetic control strategies to be used on pest species (such as the insertion of dominant lethal genes) work only for obligately sexual, outcrossing species (Atkinson and O'Brachta 1999, Thomas et al. 2000). Because many weeds are self-fertilized, they might not be amenable to this approach, and other weeds that currently outcross could become resistant to genetic control by increasing their frequency of inbreeding. Many pathogens and insect pests are parthenogenetic; these species would not be candidates for the new genetic control strategies.
Genetic control strategies are exceptionally specific in their action, so they are likely to be environmentally friendly. Once a genetic control strategy is put into place, it can maintain itself and not require continued expenditure of funds. Many obligately outcrossing pests (such as weeds and household and veterinary pests) could be difficult to control through plant engineering. There is a need to assess carefully what the niche might be for direct genetic control techniques. The niche and the environmental benefits might be larger if substantial funding is used immediately to support research. If research support is insufficient or too late, the niche for this environmentally benign control technology could become occupied by other more expensive and less benign techniques. Public concern about the use of genetically engineered insects could, however, overshadow any of the potential environmental benefits of this technology and could block its implementation.
Targets of Chemical Pesticides
In the last 50 years, there has been an intensive search for compounds that will control pests without damaging crops or the environment. During this time, the way the search for new pesticides has evolved with respect to how it is done and the criteria by which discoveries are judged. The current methods used to find and develop new pesticides are sophisticated and complex. A new compound has to be not only highly efficacious, but also environmentally acceptable. Thus, testing on the environmental behavior and toxicology of a candidate compound comes much earlier in the discovery process than it did in the past. If a candidate cannot meet the new criteria for environmental safety, as well as efficacy, it is not advanced.
The number of compounds that have to be tested before a viable candidate is found has increased from 2,000 to 40,000–50,000 (Stetter 1998). There are many reasons for the increase, including environmental regulation. For example, an upcoming material needs to “beat the standard”. That is, for a herbicide to be truly promising, it has to act at extremely low rates to outperform the materials alredy being used. Thus, many more compounds must be screened than would be needed simply to identify a compound with equivalent efficacy. Another problem is resistance. And, because many of the known target sites are no longer available, materials that contain novel modes of action are sought.
The current method of screening for new pesticides starts with high-throughput screens that can handle hundreds of thousands of compounds per year. These screens include miniaturized systems in which whole organisms are grown on microtiter plates or other systems but require microgram or nanogram quantities of material for tests, which can be run in a minimum of space and time (Jansson et al. 1997). There is also increased use of mechanism-based screens in either modified microorganisms or in vitro enzyme assays, where a particular molecular or enzymatic target is targeted. In most cases the molecular targets are ones that are peculiar to the pest and are not found in nontarget organisms (for example, essential amino acid biosynthetic pathways) (Abel 1996). The advantage of mechanism-based screens is that a search can be restricted to compounds that will selectively control the pest with less chance of nontarget toxicity. In addition, with the development of resistance to many current pesticides, compounds with new mechanisms of action have become more valuable. Mechanism-based screens potentially can find compounds that hit new target sites where there is no known resistance. A limitation is that activity detected with in vitro assays might not show activity on the whole organism.
Another change in the discovery process for new pesticides is the
time when research on the mechanism of action begins. In the 1950s to 1970s, research on determining the mechanism of action of new leads would not begin until after a compound that could lead to a product had been found or even registered for use. Now this research begins as soon as a new chemical is discovered to be biologically active. One reason for the change is to eliminate compounds with high potential for mammalian toxicity. Companies now start testing for mammalian (and other nontarget) toxicity early in the screening process. This has involved the development of high-throughput screens to test for nontarget toxicity. Another reason is to determine whether a new mechanism of action has been discovered. This information is used to develop new mechanism-based screens and to conduct computer modeling to guide the synthesis of analogues to locate better, more potent compounds. It is also used to determine the potential for resistance.
Combinatorial Chemistry
Combinatorial chemistry encompasses systems in which a large number of chemical compounds are made via multiple parallel synthesis. In this system, a set of building blocks (N) is coupled through repetitive steps (x) to synthesize a number (Nx) of different compounds. The process results in a chemical library of a few hundred to millions of compounds, depending on the number of building blocks and the number of repetitive steps (Lyttle 1995.) The technique was first used to make peptides and oligonucleotides, but more recently it has been adapted to make nonpolymeric small molecules (Warr 1997a). In a recent publication, it was reported that one person working 7 months was able to generate a 400-member library of heterocyclic carboxamides; this resulted in identification of a herbicidal compound with 4 times the activity of the original lead compound (Parlow et al. 1997.)
Combinatorial chemistry is often done on a solid support phase, which greatly simplifies automation, product isolation, and purification. The disadvantage of this approach is the limited size of the sample, the functionalization needed for attachment to the solid phase, and the chemistries needed for linkers and capping to prevent unwanted reactions (Warr 1997a). Another problem with combinatorial chemistry is the need for sufficient diversity in the chemicals being synthesized to optimize the chances of discovering useful compounds. The latter problem has been addressed by making “virtual” libraries via specialized software programs (Warr 1997b) and then synthesizing a small number of compounds with the highest diversity. This procedure was used successfully to find an isozyme-specific inhibitor of glutathione-S-transferase (Lyttle et al. 1994).
Combinatorial chemistry has been widely adopted by many pharmaceutical and agrichemical companies to increase the potential for finding new, profitable products. It has been estimated that a company needs to synthesize and screen 40,000–50,000 compounds to find a compound that can lead to a new product. Using traditional methods of synthesis, one chemist can produce only 50–100 compounds per year. Using combinatorial and parallel-synthesis methods, one chemist has the potential of making several thousand compounds per year (Warr 1997a). Combinatorial synthesis is used not only to find new leads, but also to optimize leads.
The compounds generated via combinatorial chemistry are processed by high-throughput screening techniques, described previously. The advantage of these techniques is the diversity of structures that can be synthesized and tested. One disadvantage is the size of the sample that is initially synthesized. The small sample limits the amount of initial testing that can be done, and in many cases mechanism-based screens or some simple in vivo initial screens are used (Lyttle 1995). Another disadvantage is that, by their very nature, compounds within a particular combinatorial library are related. Depending on the measure of diversity used, combinatorial chemistry might or might not be a good source of broadly diverse chemistry for lead generation.
Development and Commercialization of New Chemicals
Functional genomics serves as a source of bioassays, monitoring, and diagnostics. The efficacy of the pesticide is only one of many criteria used by industrial researchers to determine whether it will be developed. After finding a new lead area of chemistry that lends itself well to synthetic modification, additional screens (beyond whole organism screens) are put into place to determine ecological, toxicological, and environmental characteristics of compounds. New screens have been developed to determine the toxicity of compounds (Yamanaka et al. 1990, Swanson et al. 1997). Tests are also conducted early in the discovery process to assess the toxicity of potential leads to nontarget organisms, such as fish or aquatic invertebrates. This information is used to guide the synthesis of analogues to try to eliminate nontarget toxicity, if present, while maintaining activity against the target pest. If nontarget toxicity cannot be eliminated, the chemical structure is not pursued further.
Screens are commonly used for quickly determining the soil binding and degradation characteristics of compounds (Lamoreaux 1990; Daniels and McTernan 1994). These screens determine the leaching potential and persistence of compounds with the goal of finding compounds that have the lowest leaching potential and minimal persistence but that still control the target pest. Compounds that have very high leaching potential or
long soil persistence are eliminated early in the discovery process unless there are other applications for which the compounds can be useful (for example, household pests).
Once a compound with the efficacy needed to control the pest with minimal impact on the environment and nontarget organisms is found, the development phase can begin. This is the most expensive part of bringing a new product to market. Development, testing, and registration can typically take 8-12 years and cost over $50 million for each pesticide (IANR 1994). Registration involves at least 142 tests, most of which are aimed at determining the environmental and toxicological characteristics of new compounds. EPA will register only those compounds that meet strict criteria for human and environmental safety. Industrial scientists must use this same battery of tests to maintain the registration of existing compounds.
Pesticide research and development have come a long way from the procedures used to find earlier compounds, such as DDT and related compounds. There is now a much better understanding of the desirable and undesirable characteristics of pesticides. Companies have set up systems to eliminate, early in the discovery phase, compounds that do not meet the new standards, to ensure that time and resources are not wasted on compounds that will not be acceptable as pest-control agents.
Application Technology
Pesticides effectively control many insects, diseases, and weeds. However, to be effective, pesticides have to target the crop or animal of interest. Pimentel (1995) estimated that only 0.00001-1% of the pesticides sprayed actually reach the target pests. The reasons for the low rate of interception are many, including the distribution of the pest, the size of the pest population at the time of application, uncertainty as to how and when the pest will encounter the pesticide, the use of broadcast sprays, and drift. One way to reduce the risk posed by pesticides is to increase the amount of pesticide that reaches the target pest while decreasing the total amount of pesticide applied to the field. That can be accomplished only through improvements in application technology (see table 3-1).
Spray drift is one of the biggest concerns regarding the movement of pesticides to nontarget organisms. Off-target losses can range from 50 to 70% of the applied pesticide because of evaporation and drift (Hall and Fox 1997). Drift from aerial applications is greatest and that from ground applications is least (Hall and Fox 1997).
There are several ways to reduce drift. One way is to use spray additives that affect the drop size of sprays by increasing the number of large droplets and decreasing the number of small droplets (Hall et al. 1993).
However, those agents can also affect the spray pattern, and lead to inefficient placement of the pesticide. Another method to decrease the number of fine droplets during spraying is to use new nozzles that are designed to decrease the number of fine droplets. The nozzles work by increasing droplet size through a reduction in the velocity of the liquid just before it is discharged (Ozkan 1997).
Spray additives can also be used to decrease the rate of evaporation of droplets. It has been shown that the most rapid evaporation occurs during the first 20 seconds of flight (Hall et al. 1993). Additives can reduce the rate. For example, a conventional formulation of permethrin lost 55% of its original volume after falling 2 m at terminal velocity at 40°C and 17% at 25°C. The loss could be reduced to 40% and 9%, respectively, with the incorporation of an antievaporant in the formulation (Hall et al. 1993).
The use of aerial electrostatic sprays has the potential to increase the amount of pesticide that reaches the crop while reducing drift. Carlton et al. (1995) showed that using a bipolar-charge protocol (having an opposite charge on each end of the spray boom of an aircraft) increased deposits by a factor of 4.3 on cotton leaves. Penetration into the canopy and wraparound leaf effects also increased. However, the technology is limited by the lack of charging nozzles suitable for the commercial market (Carlton et al., 1995). In addition, the effectiveness of electrostatic sprayers depends on environmental factors, such as temperature and relative humidity, height of the spray boom over the canopy, and distribution of the pesticide spray throughout the canopy (electrostatic sprayers tend to deposit most of the pesticide material in the top of the canopy).
Another method of increasing pesticide efficacy while decreasing drift potential is through the use of air-assisted sprayers, which allow the use of ultra-low-volume applications of pesticides in oil-based diluents. Mulrooney et al. (1997) used such a system to increase the effectiveness of malathion while reducing the total amount of pesticide applied. Bohannan and Jordan (1995) were able to increase the effectiveness of postemergence applications of various grass herbicides (sethoxydim and clethodim) by applying them in oil diluents. Sethoxydim and clethodim applied at 25 and 16% of their recommended dose rates gave over 90% control of yellow foxtail (Setaria glauca). However, that was not the case with two other herbicides, imazethapyr and bentazon, possibly because of the lack of a suitable formulation that was compatible with oil.
Hooded sprayers, spray towers, and spray tunnels are examples of mechanical methods for increasing the amount of pesticide reaching the target while reducing off-target movement (Hall and Fox 1997). However, the effectiveness of these physical methods varies with environmental conditions, crop canopy, and other such factors. Research needs to con-
tinue to determine the most efficacious way to reduce drift using physical barriers.
Researchers are examining other methods for selective application of a pesticide that targets the pest while minimizing application to nontarget organisms. Such methods should decrease the volume of pesticide spray while maintaining efficacy. Weed-sensing sprayers that use red and near-infrared sensors to detect the presence of green tissue have been successfully used in chemical fallow and hooded sprayers to reduce herbicide use by 14-90% (Ahrens1994, Hanks and Beck 1998, Blackshaw et al. 1998b.) However, these sensors and accompanying analytical equipment are relatively expensive, and the sensors are limited by the size of the weed that can be detected and are affected by environmental conditions, such as soil type, and wind speed (Blackshaw et al. 1998a).
The use of baits in combination with a pesticide has the potential of reducing the amount of a pesticide applied while maintaining efficacy. For example, the amount of malathion insecticide needed for control of Mediterranean fruit fly (Ceratitis capitata) in Israel was reduced from 1 kg/hectare to 150 g/hectare through the use of baits (Grinstein and Matthews 1997).
One of the biggest reasons for overapplication of pesticides is variation in the speed of the application equipment. New technology that constantly monitors ground speed and adjusts the application of the pesticide accordingly is being developed. The primary means of adjusting application rate is to alter the pressure of the spray. However, because the performance of many spray nozzles is pressure-dependent, applicators are limited in how much they can alter the pressure without affecting the distribution pattern of the spray (Paice et al. 1996). Other systems under development include use of a direct-injection system in which the spray volume is kept constant but the amount of active ingredient is varied (Ghate and Perry 1994).
Precision-agriculture techniques are new technologies under development that can vary the amount of herbicide application. In the future, users might map a field to identify locations of various weed populations and then select the amount and type of herbicide spray that match the weed density and population for that area of the field (Paice et al. 1996). One of the main limitations in adopting these mapping and sensor technologies is the need for sprayer technologies with rapid response and rate controllers that can automatically adjust the amount of active ingredient. Injection-metering devices, twin fluid nozzles, pulsed-nozzle systems, and rotary-spray generators with prediluted herbicides can decrease response time to less than 1 second.
Finally, one of the major risks in pesticide application is exposure of the applicator during the mixing procedures and disposal of used con-
tainers. Closed systems are being developed for many pesticides through minibulk systems and innovative water-dissolvable bags, which virtually eliminate exposure of the applicator to the pesticide during mixing. Furthermore, many companies are providing returnable containers, such as the “minibulks” (refillable tanks that holds typically 50 to 100 gallons of pesticide) that greatly reduce disposal costs and risks associated with the disposable smaller 2.5 gallon pesticide containers (Frei and Schmid 1997). A closed system greatly reduces exposure to the pesticide because the pesticide flows from the storage tank directly into the application device (the sprayer) and only does so if the container is properly connected to the sprayer. Additional work needs to be done in these technologies to reduce the risks even further.
Precision Agriculture
Precision agriculture can be defined as a bundle of technologies designed to adjust input use to variations in environmental and climatic situations over space and time so as to reduce residues associated with input use. Many of these technologies rely on space-age communication and incorporate the use of global positioning systems. Modern irrigation technologies that adjust input use according to variability in soil and weather conditions and rely on weather stations and moisture-monitoring equipment are also examples of precision technologies. Precision technologies have the potential to increase input-use efficiency, increase yields, and reduce residues of chemicals that can contaminate the environment. In many cases they can lead to input saving; but in others, the yield effect can also entail increased input use (NRC 1997).
There has been substantial variation in rates of adoption of what can be generically defined as precision technologies. Some modern irrigation technologies have high rates of adoption in high-value crops. Some components of precision agriculture, such as yield monitors, are gaining acceptance among grain producers. But overall rates of adoption of many components of precision agriculture have not been high (NRC 1997). Although some of the technologies can be obtained through consultants, adoption can be hampered by the cost of investment. Furthermore, the management software needed to take advantage of this highly technical information has not been fully developed. Assuming that management software does become available, adoption of precision farming technologies is likely to accelerate as their costs decline and productivity increases are demonstrated.
Precision technologies will allow farmers to take advantage of a wider arsenal of pest-control tools, to adjust application rates, and to target areas where pest problems are most severe. Because agricultural pollu-
tion can result from pesticides that do not reach their intended targets, precision technologies have the potential to reduce pesticide emissions into the environment and to reduce the negative side effects of pesticide use. However, precision agriculture is early in its adoption phase, and environmental benefits are not proved. Future research should focus on environmental impacts of precision technologies in the field and watershed scales (NRC 1997). The emergence of precision technologies is contributing to the increased value of information in agriculture, and it could also contribute to the value and use of agricultural management consultants who have access to data and analytical capability and can provide effective management strategies that take advantage of large bodies of information.
Remote Sensing and Pest Management
Satellite and airborne remote sensing data, coupled with geographic information systems and global positioning satellites, are potentially powerful tools for monitoring pest infestations and their crop impacts. Remote sensing has been effective in identifying new colonies of weeds in low-access areas and monitoring the spread of metapopulations (Lass and Callihan 1997, Lass et al. 1996, Everitt et al. 1993, Carson et al. 1995; Anderson et al., 1993). Thus, this technology can help to set priorities for herbicide application and to direct treatment to the locations where it can have the greatest impact on weed populations. In addition, airborne sensors that detect multispectral reflectance differences between crop and other vegetation canopies can identify crop-vigor variation in response to insect pests and plant pathogens (UC Davis, 1997; Baldy et al., 1996; Hill et al., 1996; Kline et al., 1996; Liedtke, 1997; Vickery et al.; 1997; Ustin et al., 1997). As this technology is improved, it is likely that pesticide use will decrease with the change from whole-field to pest-location-specific applications. Remote sensing for pest management in forest ecosystems has been widely used for several decades. Proposed improvements in application of remote sensing for pest management include increased availability of sensors, improvements in quality of images, and increased resolution of the spectral information (UC Davis, 1997).
A large number of earth-observing satellites scheduled for launch in the next few years will enable specific monitoring of environmental conditions and natural-resource management. Some commercial multispectral satellites will be collecting hyperspatial data (with spatial resolution of 1-5 m), and many others will be in resoulution range of 5-100 m. Other satellites will collect multispectral data in hundreds of bands (NASA 1998). Remote sensing is likely to become an increasingly important tool
for identification of pests and their impacts and later for site-specific application of pesticides.
Increasing Knowledge of Pest Ecology
Population dynamics, the field of ecology most relevant to pest management, has become a mature field of inquiry in the last 20 years. Many of the broad generalizations made in the past have been reassessed because they failed to explain important patterns in the populations of plants and animals (Denno et al. 1995, Kareiva 1993). Recent advances in population dynamics, at both theoretical and empirical levels, are applicable to agricultural systems (Kareiva 1996, Turchin 1999).
Insects and early-successional plant species have figured prominently in many basic studies of population dynamics because they are prominent parts of the landscape and because their population fluctuations can be monitored and manipulated within a reasonable time and spatial scales. Scientists with basic and agricultural job responsibilities have cooperated in some major studies because the goals in both are similar: the prediction of future population densities. An intellectual synergism has often resulted from such cooperative studies (Murdoch et al. 1984).
Biological-control researchers can boast of many successes, but the factors that lead to success or failure are rarely understood. Work by basic and applied scientists led to the careful dissection of the population dynamics involved in successful biological control of the citrus red scale (Murdoch et al. 1995, Murdoch and Briggs 1996). Insights gained from that work could help in designing future biological-control efforts aimed at this and other pests. Recent work on interactions among predators of herbivores (Rosenheim 1998) has shown that two species of predators are not always better than one in decreasing population densities of herbivores. That is an important issue in designing biological-control systems. Finally, the finding that plants, in response to damage by a herbivore, produce specific compounds that then attract parasites and predators of that herbivore also offers new insights into patterns of insect population densities (Tumlinson et al. 1998). These and other new findings could be used in breeding crop cultivars that are more compatible with biological control (NRC 1996).
Herbicide costs are an important component of agricultural production in some crop systems. Knowledge that allows for decrease in herbicide use can improve agricultural efficiency in those systems. Without an understanding of weed population dynamics, the decision to forgo the use of a herbicide can be risky. Research on weed population dynamics (Cousens and Mortimer 1995) indicates that long-term predictions about weed densities might not be possible, but that shorter-term, more specific
predictions are. New crop-specific models (White and Coble 1997) have been developed and have been proved useful in predicting whether herbicide treatment is economically justifiable. In other systems, research is under way to determine whether certain crop cultivars or combinations of plantings can be developed that will regulate weed populations at low densities with little input of herbicide and cultivation (Jordan 1993).
One of the major drawbacks in using ecological knowledge in agriculture is the information-processing and communication barrier. It will take time before most farmers are routinely using on-line extension information and maintaining their own computer-based records, but the transformation is under way. As ecological knowledge of pest population dynamics improves and becomes more accessible to farmers, the risks inherent in decreasing (or tailoring) pesticide can be diminished. In the last decade, there has been a tremendous increase in Internet-accessible information on pest identification and pest biology. Web pages are also being developed to help farmers with identification of the beneficial organisms in their fields. It is challenging to link the basic Web pages to interactive programs that can offer alternative solutions to problems associated with an identified pest. As more information is available on the impact of specific pesticides on specific beneficial organisms, it should be possible for farmers to choose pesticides according to which beneficial organisms are in their crops. Studies of population dynamics of plant pathogens have been hampered by our inability to monitor fungi, bacteria, and viruses when they are at low densities.
Decision-Support Systems for Pest Management
A wide array of issues—including crop productivity, environmental protection, and profitability —creates a complex background against which agricultural producers or professional managers are expected to make decisions about optimal pest management. Decision support systems (DSS) can condense the knowledge base required to make decisions through the use of models and databases of expert knowledge (Wagner 1993). DSS are defined as knowledge-based systems that help producers and managers to make strategic and tactical management decisions on pest-control treatments, time of planting and harvesting, cultivar selection, and marketing options (Plant and Stone 1991). Moulin (1996) described two types of DSS. The first consists of models based on production functions calculated with empirical or mechanistic relationships between variables, such as soil properties, temperature precipitation, and yield. The models can be used to improve understanding of biotic and abiotic factor relationships with production or as management tools for decisions related to production. Models of the second type are used stra
tegically to provide supportive information for broad policy decisions or tactically to provide specific recommendations. Most pest-management DSSs are of the second type and are based on rules derived from expert knowledge; they are often focused on pesticide selection rather than a complete set of management practices for managing a particular pest. The low representation of alternative tactics in DSSs is a result of inability to predict the outcome of these tactics. Cultural and biological pest-management tactics rely on more subtle influences on pest populations; so more environmental factors play a role in determining population dynamics and there is greater variability in success rates. DSSs have not been extensively used by producers because of the sparseness of specific information input requirements (NRC 1997).
ECOLOGICAL CHANGES AFFECTING AGRICULTURE
The distribution and abundance of organisms on the planet have long been influenced by global environmental change. For the most part, such changes have taken place on geological time scales. In recent years, however, it has become apparent that anthropogenic changes in global environments can affect the distribution and abundance of organisms on ecological time scales. Over the last 2 centuries, industrialization, urbanization, and agricultural development have contributed to alterations in global environmental patterns that are historically unprecedented in their magnitude. Among the forms of environmental change most likely to affect American agriculture and forestry, several stand out: an increase in atmospheric carbon dioxide resulting from fossil-fuel combustion, and a concomitant increase in temperature; stratospheric ozone depletion linked to chlorofluorocarbon emissions, and a consequent increase in exposure to short-wave ultraviolet-B radiation; increases in the frequency of biological invasions associated with internationalization and globalization of trade; and an increase in extinction rates due to expansion of development, particularly in tropical regions. Each of those has the potential to alter the relationships among agricultural and forest pests, their hosts, and growers or producers.
Carbon Dioxide and Global Warming
Increases in rates of fossil-fuel consumption worldwide have steadily increased atmospheric concentrations of carbon dioxide over the last 100 years. Increased carbon dioxide has two potential impacts on agricultural productivity. First, as a critical resource for photosynthesis, carbon dioxide availability can affect growth rates, yields, and chemical composition of both crop plants and weeds. Generally, increases in carbon dioxide
concentrations are associated with increases in productivity, particularly for annual crops. In the case of annual crop plants exhibiting C3 photosynthesis, such productivity increases average about 30% with a doubling of contemporary carbon dioxide (Cure and Adcock 1986,Rogers and Dahlman 1993). Individual species vary, however, in their responsiveness (C4 plants—such as corn, sorghum, sugar cane, and millet—exhibit a considerably lower response). The available evidence suggests that some direct growth and physiological responses to increased levels of carbon dioxide would be less dramatic in woody plants than in annual crops (Watson ey al, 1996). Conversely, woody plants show greater alterations in secondary chemicals than do annual plants and thereby could show the greatest alterations in resistance to insects and pathogens (Penuelas and Estiarte 1998).
Differential impacts on weedy species and crops could affect the economic impacts of weeds. Outcomes are difficult to predict. C3 plants tend to benefit to a greater extent from the “carbon dioxide fertilization effect;” Watson et al. (1996) estimate that, although 80 of 86 of the plant species that contribute 90% of per capita supplies of food are C3 plants, only four of 18 of the worst weed species are C3 plants. One possible outcome of increased carbon dioxide, then, might be a change in the composition of the principal weed flora.
Changes in carbon dioxide can alter plant composition, as well as plant growth rates. Under conditions of increased carbon dioxide, the production of nonstructural carbohydrates in many species is increased and protein and mineral-nutrient concentrations decline (Mooney and Koch 1994). Exposure to increased carbon dioxide can also lead to an increase in root:shoot ratios. Such changes in tissue composition and biomass allocation can have important affects on the extent and distribution pattern of herbivore or pathogen damage. In particular, dilution of protein and mineral nutrients by increases in nonstructural carbohydrates can lead to compensatory increases in food intake by herbivores (Slansky 1992) and concomitant increases in tissue losses. Increases in starch content of weed species and increases in allocation of biomass to roots or rhizomes could potentially reduce herbicide efficacy, particularly in C3 weed species.
Increased carbon dioxide also has the ability to affect plant-pest relationships through by affecting global temperatures. Through the “greenhouse effect”, increased carbon dioxide is predicted to increase average global temperatures by 1.5–4.5°C. Such an increase in temperature can have profound effects on both crops and pests. Higher temperatures can lead to a variety of problems in crop plants. Many critical developmental processes have relatively narrow optimal temperature ranges and are sensitive to excesses. Increased temperatures can affect the quality of vari-
ous cool-season crops, which have winter chilling requirements for bud set (for example, apple, cherries, and other tree fruits) or require lower night-time temperatures to maximize quality (for example, spinach and broccoli). Excessively high temperatures can also affect pollen viability, as in maize (Schoper et al. 1987) or tuber initiation with potato (Kooman et al. 1996). Another manifestation of the greenhouse effect is increased variability in temperatures; extremes in weather conditions can cause losses in a wide array of crop types.
Global warming can increase pest problems in agriculture and forestry as a result of expansion of insect, weed, and pathogen ranges into hitherto inhospitable regions. Temperature can influence surrounding developmental rates, fecundity, and activity of individual pests, as well as the size, distribution, and continuity of populations of pests (Drake 1994). Cold temperatures limit the distribution of many insect pests, particularly those of tropical origins. At present, for example, the red imported fire ant (Solenopsis invicta), accidentally introduced into North America from Brazil, is effectively restricted to southern states, where they cause considerable damage (Callcott and Collins 1996). This distribution parallels that of the introduced Apis mellifera scutellata, the so-called African (“killer”) bee, initially brought to Brazil from Africa and now reported to occur in the US South and Southwest. Porter et al. (1991) predicted that, in Europe, a 3–6°C increase in European temperature by 2025-2070 could expand the range of the European corn borer Ostrinia nubilalis by 1,220 km northward; Mochida (1992) predicted that a 3°C increase in temperature in Japan would lead to expansion of the ranges of eight major pests of rice or soybean. Damage inflicted by agricultural and forest pests could potentially increase as a result of decreased development time and increased voltinism. An increase in the number of breeding cycles per season raises the possibility that resistance to insecticides could evolve more rapidly.
A reduction in winter temperatures could increase the severity of plant-disease outbreaks; disease problems tend to be exacerbated under such conditions. Presumably, pathogens could also experience range expansion as a result of warming trends. Disease incidence and severity are subject to change in animal agriculture. Increases in average temperatures are predicted to increase the activity of vector species, such as culicid mosquitoes, and microorganisms of tropical origin could make inroads at higher latitudes.
Constraining any definitive predictions of impacts of increased carbon dioxide is the acknowledged buffering capacity of genetic adaptation and technological change. There is considerable evidence of genetic variation in temperature tolerance among crop cultivars and it is well documented for such important crops as wheat, rice, maize, and soybeans
(Watson et al. 1996). Existing agronomic practices have the potential to ameliorate adverse effects of increased global temperatures; the greatest uncertainty lies in whether economic systems have the flexibility to incorporate the necessary technological innovations in a timely way.
Increased Ultraviolet-B Radiation
As a result of stratospheric ozone depletion, due principally to emissions of chlorofluorocarbons, levels of ultraviolet-B-radiation (UV-B) (280-315 nm) reaching the earth's surface have increased in all temperate latitudes (Madronich 1993). Increased UV-B radiation is an environmental stress factor that can alter the physiology and development of terrestrial plants in several ways (Caldwell et al. 1989, Tevini 1993), ranging from such cellular and subcellular effects as DNA damage and degradation of photosystem II proteins to such whole-plant effects as stunting, shortening of internodes, alterations in root:shoot biomass ratios (Tevini and Teramura 1989), and alterations in plant primary and secondary chemistry (Ros and Tevini 1995,;McCloud and Berenbaum 1994). At the ecological level, these developmental effects can manifest themselves as yield reductions in several crop species (Tevini 1993). Increased UV-B can also affect interspecific competition in plants (Gold and Caldwell 1983), reduce plant resistance to pathogens (Orth et al. 1990, Panagopoulous et al. 1991), affect pollinator-plant relationships (Collins et al. 1997, Feldheim and Connor 1996), and alter host-plant suitability for herbivores (McCloud and Berenbaum 1994).
Plants have the capacity to reduce damaging effects of UV-B in several ways. Among these is the production of compounds that absorb damaging wavelengths of UV radiation. In several plant species, increased UV-B exposure induces production of flavonoids and other phenolics (reviewed in Caldwell et al. 1989.). Plants can also ameliorate the oxidative damage brought about by UV exposure by enhanced synthesis of enzymatic and nonenzymatic antioxidants (Larson 1988). Alpha-tocopherol, ascorbic acid, and glutathione are all effective scavengers of reactive oxygen species and in several temperate crop species increase in response to UV exposure (Foyer 1993, Grace and Logan 1996).
Inasmuch as antioxidant defensive compounds also serve as essential nutrients for insects, UV-B exposure can potentially increase plant suitability for herbivores. Indeed, Hatcher and Paul (1994) found that elevated UV-B increased nitrogen content of Pisum sativum and digestibility of its foliage by Autographa gamma, a caterpillar. This enhanced nutritional suitability, however, appears to be counterbalanced in other plants by increases in phenolics, flavonoids, and other UV-screening compounds, many of which are toxic or deterrent to insects.
Predicting overall effects of increased UV-B levels on US agriculture is difficult because of the scarcity of appropriate studies. The impact might be minimal. Genetic variability in resistance to UV-B has been documented in crop species (Teramura 1983, Teramura and Sullivan 1991, D'Surney et al. 1993) and weed species (such as Plantago lanceolata; McCloud and Berenbaum 1994); this allows for the possibility of adaptation in the case of weeds and selective cultivar use in the case of crop plants. Moreover, the nearly universal adoption of the Montreal protocols for global reductions in chlorofluorocarbon production suggests that ozone depletion could diminish, rather than accelerate, in the future.
Increased Frequency of Biological Invasions
Biological invasions constitute a form of global change as devastating ecologically and economically as more widely recognized global climatic changes (Vitousek et al. 1996). The growth of agriculture, industry, and development over the last 200 years has led to redistribution of species in all major taxa. In the United States, for example, over 5,000 species of alien plants have been introduced and established, infesting over 100 million acres of terrestrial and aquatic habitats; the acreage occupied by these plants is increasing 8–20% per year. The proportion of the flora that is made up of alien weeds varies by state from less than 10% to, in the case of Hawaii, almost 50% (Pimentel et al. 2000).
The impacts of alien species on agriculture and forestry are enormous (Niemela and Mattson 1996). Many of the introduced species are themselves pests; life-history attributes that are conducive to colonizing new habitats—including exploitative reproductive traits, broad resource requirements, small bodies, and high vagility—predispose these species to pest status. That problem is compounded by the fact that many species that are not pests in their native range become important pests in their new, acquired range because factors that historically regulated their population, such as natural enemies, are not present. Moreover, introduced species can act as disease vectors and facilitate the entry of new pathogens into nonnative agroecosystemso
Introduced species can reduce yields by removing crop biomass directly (as in the case of alien insects) or by competing with crop plants for water, minerals, or light for photosynthesis. Introduced species can also cause problems by altering trophic dynamics and ecosystem structure.
Forest ecosystems in North America have been radically altered, and dominant species driven to near extinction, by such invasive pests as chestnut blight, gypsy moth, and balsam woolly adelgid (Niemela and Mattson 1996). On rangeland, toxic species can poison livestock (even, on occasion, pollinator species); in agricultural and forest ecosystems, intro-
duced species can alter drainage patterns and accumulation of biomass and create fire hazards. Cheatgrass, a fire-adapted annual, has invaded over 5 million hectares of shrub-steppe habitat in the Great Basin and, by eliminating native perennials, contributed to the frequency of fires, which increased by a factor of 20 (Vitousek et al. 1996).
Introduced species, via introgression and hybridization with native species, can also interfere with evolutionary processes (Darmency 1997), with many possible outcomes, including extinction of native species, introduction of pesticide-resistance genes into susceptible genomes, and transfer of agriculturally undesirable traits into native species. All told, the management of alien weeds alone in the United States exacts a cost of $3.6–4.5 billion per year (Westbrooks 1998).
As for the impact of biological invasions on the future of pesticide use in the United States, suffice it to say that some of the most controversial examples of pesticide use in the last half-century have involved attempts to eradicate introduced species. An ill-fated campaign launched in 1957 to eradicate Solenopsis invicta, the red imported fire ant, culminated in expensive failure, public concern, and political concern about the safety of the chemical control agents used (Kaiser 1978, Hinkle 1982). Efforts to contain Mediterranean fruit flies (Ceratitis capitata) in California in 1989-1990 with malathion-laced baits have also led to a public outcry (Myers et al. 1998). Pesticides have often been used initially to eradicate localized populations of introduced species because they kill more quickly than do most biological control methods. Such use, while often highly desirable in agricultural contexts, might be less appropriate for eradicating normative species in preserves, refuges, and wildlands, where use of chemicals is inconsistent with management practices and can pose unacceptable risks to nontarget species, and in urban green spaces, where pesticide use can present unacceptable risks of exposure to large numbers of people.
In view of the increasing globalization of trade, accidental introductions will probably increase in frequency in the near term. Possible consequences of this accelerated pace of invasion are increased frequency of pesticide failure due to resistance acquisition and increased demand for narrow-spectrum chemicals or biologically based approaches.
Loss of Biodiversity
Whether accelerated biodiversity loss contributes a separate form of global change or is the consequence of the combined effects of many human-caused forms of global change is debatable. What appears to be clear, however, is that extinction rates globally are higher than rates recorded in the recent past (Pimm et al., 1995; Purvis et al., 2000; Wilson 1993). Effects of biodiversity loss on agriculture vary with site and system. In general, how-
TABLE 4-9 Number of Papers Published in 1996 that Report on Biologically Active Natural Substances
|
Activity |
No. of Publications |
|
Anthelmintic |
13 |
|
Antibacterial |
176 |
|
Antifeedant |
19 |
|
Antifungal |
91 |
|
Insecticidal |
10 |
|
Molluscicidal |
7 |
|
Pheromonal |
52 |
|
Plant-growth modulating |
17 |
|
Source: Claeson and Bohlin, 1997. |
|
ever, potential effects include loss of critical germplasm for crop improvement, breakdown of such mutualistic associations as those between pollinator and plant and between plant and mycorrhizae, and decrease in availability of natural enemies for use in biological-control programs. With respect to direct effects of extinctions on pesticide use, in that natural products have often provided leads for development of chemical control agents (or have themselves served as control agents), biodiversity loss could eliminate potential sources of new chemicals that have novel modes of action or novel sites. In a review of all articles appearing in 1996 in over 130 major journals dealing with natural products, Claeson and Bohlin (1997) tabulated the number of publications that reported a wide range of types of biological activity. In that single year, over 150 publications documented pesticidal activity of some sort (insecticidal, antifungal, pheromonal) (Table 4-9). Many random-screening efforts focus on floras and faunas in regions of the world at greatest risk of biodiversity loss through extinction (such as tropical rain forest) and could be compromised as well.
EVOLUTIONARY CHANGES IN PESTS
Evolutionary interactions between pests and farmers predate conventional pesticides by thousands of years (Day 1974). Evidence of such interaction is best preserved in weed characteristics that enable specific genotypes to avoid effects of traditional cultural control practices. As a result of changes in their timing of reproduction and seed size, some weed populations display resistance to winnowing, a process used by prehistoric and modern farmers to eliminate weed seeds from grain seeds, including those being saved for the next season's planting (Gould 1991). Other genotypes produce seedlings that physically resemble crop seed
lings and thereby decrease the efficiency of hand and hoe weeding procedures (Barrett 1983).
The historical record of insect and microorganisms adaptation to traditional agricultural practices is harder to trace than that of weeds, but there is evidence that prehistoric crop breeding indirectly produced cultivars resistant to pest attack. In turn, pest genotypes that are capable of surviving on these traditional cultivars have evolved (for example, sheath blight fungal genotypes found on traditional Tibetan cultivars of rice). In recent history, a major corn pest, Diabrotica barberi (northern corn rootworm), has adapted to the yearly rotation of soybean and corn by maintaining a quiescent egg (diapause) for an additional year (Krysan et al. 1986, Fisher et al. 1994); then eggs hatch in the season that corn is planted. New data indicate that a closely related corn pest, the western corn rootworm Diabrotica virgifera, has evolved in a different way to avoid the effect of corn-soybean rotation (Sammons et al. 1997). It feeds on corn, but late in the season it lays many of its eggs in soybean fields, which with crop rotation, will become cornfields in the following year.
Even biological control is not immune to the effects of pest evolution. A number of studies show that pests can evolve resistance to their parasitoids (e.g. Petersen 1978, Henter and Via 1995) and pathogens (e.g., David and Gardiner 1960, Alexander et al. 1984). But one unique advantage possessed by self-perpetuating biocontrol agents is their ability to respond evolutionarily to changes in the pests (Sasaki and Godfray 1999, Fellowes et al. 1998).
As explained in chapter 2, there has recently been much well-deserved attention to the evolution of pest resistance to synthetic pesticides. Carefully maintained records show that over 500 insects, 200 weeds, and 160 pathogens have evolved resistance to at least one pesticide (Delp 1988; Bills and Whalon 1991; Ffrench-Constant 1994; Georghiou and Lagunes 1988; Heap 2000; Holt et al. 1993; NRC 1986; Warwick 1991). Many pests are documented to have become resistant to new pesticides in less than 3 years (Forgash 1984, Shelton and Wyman 1991, Sun et al. 1992, Schwinn and Morton 1990). Unlike biological control agents, synthetic pesticides cannot respond evolutionarily to pest adaptation. That response is the responsibility of modern agriculture. A major factor in the “pesticide treadmill” involves two responses to pesticide resistance. The first is to increase the dose and frequency of use of the less effective pesticide (Georghiou 1986); this typically results in higher levels of pest resistance and damage to natural enemies and the environment. The second response is to develop and commercialize a new pesticide. The treadmill concept assumes that this two-step process will continue until the pest meets a resistance-proof pesticide or until the supply of effective new pesticides is exhausted.
There are certainly cases in which specific pests seem to lack genetic variation for adapting to a particular pesticide (such as the boll weevil and the organophosphate Guthion). But there are also cases in which, in the absence of new pesticides, a resistant pest has caused the collapse of a cropping system (such as Canyete Valley and Nicaraguan cotton industries, Barducci 1972). A difficult question for the future is whether products and approaches can be developed to ensure efficacy against specific pests.
One generalization that emerges from agricultural history and evolutionary theory is that the greater the impact of control measures on pest populations, the more extreme are their evolutionary responses. For example, traditional winnowing procedures were not very efficient, so weeds could find their way into seed bags without great resemblance to the crop seed. Modern winnowing procedures are highly efficient, so only the weed genotypes that precisely mimic the crop escape detection (Gould 1991). It is the hope of modern agriculturists that they can make the requirements for pest adaptation so stringent that pest evolution by natural selection will fail. That was certainly the hope of some early advocates of the use of insect hormones and pheromones in control; some even went so far as to suggest that counteradaptation to hormones was unlikely or impossible (Williams 1967). Early on, that proved not to be the case, however and multiple studies have demonstrated that this group of control substances rapidly selects for resistance. Within a few years, simple laboratory experiments resulted in resistant insects (e.g., Shemshedini and Wilson, 1990).
In recent years, many synthetic pesticides have been developed that are more toxic to target pests or more selective in the spectrum of pests that they affect than older classes of compounds. For example, about 1 lb. of an organophosphate insecticide per acre was needed to control Heliothis virescens, a major cotton pest. The pyrethroids that replaced organophosphate insecticides provided control at 0.01–0.10 lb/acre. The newly registered insecticide can control this pest with only 0.04 lb/acre. The same trend has been seen with some fungicides and herbicides. Environmental and health benefits are often associated with these two trends, but the trends could lead to problems with evolutionary sustainability. The heightened target toxicity has often been the result of new compounds that affect specific biochemical targets in the pest. Whereas traditional fungicides affected a number of target sites (NRC 1986), some of the new herbicides affect only a single site. Organophosphates and pyrethroids attack components of the nervous system common to all insects, but some of the newer insecticides affect target sites that are only found in specific groups of insects. For example, the toxins from Bt are effective at very low concentrations when a target insect has a midgut receptor to which the
toxin can bind. However, the presence of these receptors is often limited to a few insect genera (Frankenhuyzen 1993).
When a target site, such as acetylcholinesterase in the nervous system is so highly conserved evolutionarily that it is used by insects and vertebrates, it can be relatively difficult for pests to evolve alterations in the molecule. When a fungicide affects many biochemical targets, it is unlikely that a single population of the pathogen will contain genetic variation needed to alter each of the targets. However, rapid resistance to fungicides that act at a single target site has arisen. In contrast, when a target site such as the midgut receptor for the Bt toxin CryIIIA is present in one beetle genus but absent in related genera, there is an expectation that the structure of these receptors is evolutionary labile and could easily change in response to heavy selection pressure. That expectation is not always met.
Higher levels of toxicity mean that each pest encounters a relatively small number of toxin molecules because the farmer uses less insecticide. Detailed evidence is lacking, and it will be difficult to predict the speed of resistance development. It might be easier for a pest to evolve alterations to metabolize the lower concentrations of the new synthetic compounds than larger concentration of older compounds. Alternatively, protein sequestration and metabolism of low concentrations of pesticides might emphasize evolution of improved affinity of the detoxifying protein for the pesticide compared with the maximal turnover rate or concentration of the protein. If the trend for developing more specific, highly toxic synthetic pesticides continues, it will be necessary to determe whether such pesticides are predisposed to rapid pest adaptation.
The new field of pesticide-resistance management, which has interwoven the disciplines of IPM and population genetics, concedes that pests have the potential to adapt to almost all control tactics. If future synthetic compounds are more prone to pest adaptation than older compounds, resistance management could become more critical. The premise of resistance management is that, through an understanding of pest genetics and ecology, approaches can be developed for using pest-control tools in ways that will diminish the rate of pest adaptation. One such approach involves presenting pests with a combination of a stringent control and a refuge from that control. EPA, agricultural companies, USDA, and university scientists are testing this approach on a large scale with transgenic cotton, corn, and potato cultivars that produce the insecticidal protein derived from Bt. Another tactic for avoidance of pest resistance is managing for the susceptible gene. It is the susceptible gene in a pest population that makes a pesticide useful as a control agent. Conversely, it is the absence or loss of the susceptible gene that makes a pesticide ineffective. With careful monitoring of susceptible types, location of refugia, and re
laxation of selection pressure, resistance can be avoided (Roush et al. 1990, Maxwell et al. 1990). If those approaches succeed, they and other resistance management approaches might gain broader acceptance and use.
Metcalf (1989) analyzed rates of insect adaptation to succeeding waves of new insecticide classes and concluded that the pace of insect adaptation to pesticides was increasing. That could be due, in part, to the factors discussed here, but Metcalf proposed an additional mechanism. He presented the hypothesis that, in addition to evolving mechanisms enabling them to cope with a specific toxin, pests are evolving a general type of genetic flexibility that “preadapts” them to dealing with future compounds. Metcalf did not offer a detailed explanation of the form of this preadaptation, but a number of scenarios are feasible (Gould 1995). When the ability to control a pest with a specific compound is lost, the organism typically only has 20–1,000 times lower sensitivity to the compound. One or a few genes typically account for most of the resistance. It is generally found that these genes also confer some resistance to related pesticides, and sometimes to chemically unrelated pesticides (for example, DDT and pyrethroids). We would be likely to study gene A if it conferred 100-fold resistance to the selecting pesticide Z and conferred 2- to 10-fold resistance to other compounds. We would be much less likely to study gene B, which might have evolved in response to selecting pesticide Z, if it conferred only 2-fold resistance to pesticide Z, because 2-fold resistance seems like a minor problem and is difficult to study experimentally. By ignoring genes that are difficult to study, such as gene B, we could miss important aspects of pest and pathogen evolution. There might be many genes, like gene B, that confer only minor resistance to the selecting toxin but confer an overall heightened resistance to most toxins. It is even possible that gene B would some day be found to confer 100-fold resistance to a new compound.
Another mechanism that could lead to preadapted pests would be alterations in a pest's metabolic flexibility. It is possible that some pest genotypes incur more physiological disruption than other genotypes because of alterations in metabolic pathways. As pests continue to be selected for altered metabolism, shifts in the general physiology of the organism that decrease the degree of disruption might be selected for. Work with the sheep blow fly (Lucilia cuprina) has shown that resistant populations have genes for altered target sites but also have a gene for ameliorating the fitness cost incurred by having the resistance-conferring form of the gene (McKenzie and Game 1987). If genes that decrease such fitness costs have general buffering impacts on pests with other resistance genes, pest populations might accumulate traits that allow them to withstand metabolic changes caused by major resistance genes.
Very few data are available for testing Metcalf's general hypothesis, but, given the potential consequences, it seems prudent to pay more attention to this possibility. Beyond the preadaptation hypothesis, there is a reason to expect future pests to have greater ability to respond to selection: the globalized economy. With globalization comes increased movement of pests. For example, molecular studies of mosquito resistance to organophosphates suggest that resistance arose in one generally isolated population in Africa and then was spread to Europe and the Western Hemisphere by airplanes (Raymond et al. 1991). Increased air travel and increased global shipping of agricultural products provide pathways for the spread of resistance genes. It can also increase the raw genetic variability possessed by the average pest and pathogen population via the continual influx of new genes from distant locations.
Environmental-protection groups have been raising another issue related to pest adaptation. They contend that as genetically engineered crops are released into the environment, genes from these crops will spread into weed and pathogen populations and will cause increased crop losses. Rigorously derived data prove that genes can be transferred between some crops and their weedy wild relatives, for example, sorghum and Johnsongrass, cultivated rice and wild rice, wild radish and cultivated radish (Klinger et al. 1992, Krimsky and Wrubel 1996). In such cases it is likely that a herbicide-resistance gene will move from a crop to a weed and could then make the weed impossible to control with the specific herbicide and related herbicides.
REFERENCES
Abel, L.M. 1996. Biochemical approaches to herbicide discovery: Advances in enzyme target identification and inhibitor design. Weed Sci. 44:734–742.
Adams, R. M., C. Rosenzweig, R. M. Peart, J. T. Ritchie, B. A. McCarl, J.D. Glyer, R. B. Curry, J. W. Jones, K.J. Boote, and L. H. Allen, Jr. 1990. Global climate change and US agriculture. Nature 345:219–224.
Adan, A., P. Del Estal, F. Budia, M. Gonzalez, and E. Vinuela. 1996 Laboratory evaluation of the novel naturally derived compound spinosad against Ceratitis capitata. Pestic. Sci. 48:261–268.
Agrow. 1998. 299. Surrey, UK: PJB Publications Ltd.
Ahrens, W.H. 1994. Relative costs of a weed-activated versus conventional sprayer in northern Great Plains fallow. Weed Tech. 8:50–57.
Alexander, H. M., J. Antonovics and M. D. Rausher. 1984. Relationship of phenotypic and genetic variation in Plantago lanceolata to disease caused by Fusarium moniliforme var. subglutinans. Oecologia 65:89–93.
Anderson, G. L., J. H. Everitt, A. J. Richardson, and D. E. Escobar. 1993. Using satellite data to map false broomweed (Ericameria austrotexana) infestations on south Texas rangelands. Weed Technol. 7:865-871.
Asian Vegetable Research and Development Center. 1991. Demonstration of IPM of diamondback moth on farmers' fields in the lowlands. Progress Report, Asian Vegetable Research and Development Center.
Asman, S. M., P. T. McDonald, and T. Prout. 1981. Field studies of genetic control systems for mosquitoes. Ann. Rev. Ent. 26:289–343.
Aspelin, A. L., and A. H. Grube. 1999. Pesticide Industry Sales and Usage. 733-R-99-001. Washington, DC: US Environmental Protection Agency, Office of Pesticide Programs.
Atkinson, P. W., and D. A. O'Brachta. 1999. Genetic transformation of non-drosophilid insects by transposable elements. Ann. Ent. Soc. Am: 92:930–936.
Baker, C. J., Stavely, J., Mock, N. 1985. Biocontrol of bean rust by Bacillus subtilis under field conditions. Plant Dis. 69:770–772.
Baldy, R., J. DeBenedictis, L. Johnson, E. Weber, M. Baldy, B. Osborn, and J. Burleigh. 1996. Leaf color and vine size are related to yield in a phylloxera-infested vineyard. Vitis 35(4):201–205.
Barducci, T. B. 1972. Ecological consequences of pesticides used for the control of cotton insects in Canyete Valley, Peru. Pp. 423–438 in The Careless Technology: Ecology and International Development, M. T. Farvar and J. P. Milton, eds. Garden City, N.Y.: The Natural History Press.
Barrett, S. C. H. 1983. Crop mimicry in weeds. Econ. Bot. 37:255–282.
Baumhover, A. H. 1966. Eradication of the screwworm fly as agent of myiasis. J. Am. Med. Assoc. 196:240–248.
Besson, F., F. Peypoux, G. Michel 1979 Antifungal activity upon Saccharomyces cerevisiae of iturin A, mycosubtilin, bacillomycin L and of their derivatives; inhibition of this anti-fungal activity by lipid antagonists. J. Antibiotics 32 (8):828–833.
Bills, P., and M. Whalon. 1991. Weeds: 216 species of weeds are resistant worldwide. In The Occurrence of Resistance to Pesticides in Arthropods. Rome: Food and Agriculture Organization.
Black, B.C., R. M. Hollingworth, K.I. Ahammadsahib, C. D. Kukel, and S. Donovan. 1994. Insecticidal activity and mitochondrial uncoupling activity of AC 303,630 and related halogenated pyrroles. Pest. Biochem Physiol. 50:115–128.
Blackshaw, R.E., L. J. Molnar, and C.W. Lindwall. 1998b. Merits of a weed-sensing sprayer to control weeds in conservation fallow and cropping systems. Weed Sci. 46:120–126.
Blackshaw, R.E., L.J. Molnar, D.F. Chevalier, and C.W. Lindwall. 1998a. Factors affecting the operation of the weed-sensing Detectspray system . Weed Sci. 46:127–131.
Blakeman, J.P., N. J. Fokkema. 1982. Potential for biological control of plant disease on the phylloplane . Ann. Rev. Phyt. 20:167–192.
Bland, J. M., A. R. Lax, M. A. Klich. 1995. Iturin-A, an antifungal peptide produced by Bacillus subtilis. Pp. 105-107 in Proceedings of Plant Growth Regulatory Society of America, 22nd.
Bohannan, D.R., and T.N. Jordan. 1995. Effects of ultra-low volume applications on herbicide efficacy using oil diluents as carriers. Weed Tech. 9:682–688.
Bonning, B. C. 1996. Development of recombinant baculoviruses for insect control. Ann. Rev. Ent. 41:191–210
Bonning, B. C. and B. D. Hammock. 1996. Development of recombinant baculoviruses for insect control. Ann. Rev. Ent. 41:191–210.
Bowen, A.T. 1991. Innovative IPM application technology. J. Arboculture. 17:138–140.
Broza, M., B. Sneh, A. Yawetz, U. Oron, and A. Honigman. 1984. Commercial application of Bacillus thuringiensis var. entomocidus to cotton fields for the control of Spodoptera littoralis Boisduval (Lepidoptera: Noctuidae), J. Econ. Ent. 77:1530–1533.
Caldwell, M. M., A. H. Teramura, and M. Tevini. 1989. The changing solar ultraviolet climate and the ecological consequences for higher plants. Trends Ecol. Evol. 4:363-367.
Callcott, A. M. and H. Collins. 1996. Invasion and range of imported fire ants (Hymenoptera Formicidae) in North America from 1918-1995. Fla. Ent. 79:240–251.
Carlton, J.B., L.F. Bouse, and I.W. Kirk. 1995. Electrostatic charging of aerial spray over cotton. Trans. Of ASAE. 38:1641–1645.
Carozzi, N., and M. Koziel, eds. 1997. Advances in insect control: the role of transgenic plants. London: Taylor & Francis.
Carruthers, R. I., and K. Hural. 1990. Fungi as naturally occurring entomopathogens. Pp. 115-138 in New Directions In Biological Control: Alternatives For Suppressing Agricultural Pests And Diseases, R. R. Basker and P. E. Dunn, eds. Proceedings of a UCLA symposia on molecular and cellular biology, series v. 112. Los Angeles, Calif.: University of Califonria, Los Angeles.
Carson, H. W., L. W. Lass, and R. H. Callihan. 1995. Detection of yellow hawkweed with high resolution digital images. Weed Technol. 9:477-483.
Claeson, P., and L. Bohlin. 1997. Some aspects of bioassay methods in natural-product research aimed at drug lead discovery. Trends in Biotech. 15:245–248.
Clough, J.M., P. J. De Fraine, T. E. M. Fraser, and C. R. Godfrey. 1992. Fungicidal betamethoxyacrylates: from natural products to novel synthetic agricultural fungicides. Pp. 372-383 in Synthesis and Chemistry of Agrochemicals III, D. R. Baker, J. G. Fenyes, and J. J. Steffens, eds. Symposium series 504. Washington, D.C.: American Chemical Society.
Coates, C. J., N. Jasinskiene, L. Miyashiro, and A. A. James. 1998. Mariner transposition and transformation of the yellow fever mosquito, Aedes aegypti. Proc. Natl. Acad. Sci. USA 95:3748-3751.
Collins, S., J. K. Conner, and G. E. Robinson. 1997. Foraging behavior of honey bees (Hymenoptera: Apidae) on Brassica nigra and B. rapa grown under simulated ambient and enhanced UV-B radiation. Ann. Ent.. Soc. Am. 90:102–106.
Cousens, M. and M. Mortimer. 1995. Dynamics of Weed Populations. England: Cambridge University Press.
Crampton, J. M., A. Morris, G. Lycett, A. Warren, and P. Eggleston. 1990. Transgenic mosquitoes: future vector control strategy? Parasitol. Today 6:31–36.
Crowe, A.S. and J.P. Mutch. 1994. An expert systems approach for assessing groundwater contamination by pesticides. Ground Water 32(3):487–498.
Cultivar 1997. The Impact of Transgenic Plants in the Markets. November 18-25.
Cunningham, J.C. 1988. Baculoviruses: their status compared to Bacillus thuringiensis as microbial insecticides. Outlook on Ag. 17:10–17.
Cure, J. D., and B. Adcock. 1986. Crop responses to carbon dioxide doubling: a literature survey. Ag. Forest Meterol. 38:127–45.
Curtis, C. F. 1968. Introducing vector incompetence. Nature 218:368–69.
Curtis, C. F.. 1992Selfish genes in mosquitoes. Nature 357:450.
Curtis, C. F. 1994. The case for malaria control by genetic manipulation of its vectors . Parasitol. Today 10:371–74.
Daly, J.C., and J. A. McKenzie. 1986. Resistance management strategies in Australia: the Heliothis and ‘Wormkill' programmes. Pp. 951-959 v. 3 of Proceedings of the British Crop Protection Conference - Pests and Diseases, November 17-20, 1986, Brighton, England.
Daniels, B.T., and W. F. McTernan. 1994. A screening method to identify the probabilities of pesticide leaching . Ag. Water Manag. 25:23–4
Darmency, H. 1997. Movement of resistance genes among plants. Pp 209-220 in T. M. Brown, ed. Molecular Genetics and Evolution of Pesticide Resistance, M. T. Farvar and J. P. Milton, eds. Symposium Series 645. Washington, D.C.: American Chemical Society.
David, W. A. L. and B. O. C. Gardiner. 1960. A Pieris brassicae (Linnaeus) culture resistant to a granulosis. J. Insect Pathol. 2:106–14.
Davidson, G. 1974. The Genetic Control of Insect Pests. London; New York: Academic Press.
Day, P. R. 1974. Genetics of Host-Parasite Interaction. San Francisco: W. H. Freeman.
Delp, C. J. 1988. Fungicide resistance problems in perspective. Pp. 4-5 in Fungicide Resistance in North America, C. J. Delp, ed. St. Paul, Minn.: APS Press.
Denno, R. F., M. S. McClure, and J. R. Ott. 1995. Interspecific interactions in phytophagous insects – competition reexamined and resurrected. Ann. Rev. Ent. 40:297–331.
Drake, V. A. 1994. The influence of weather and climate on agriculturally important insects – an Australian perspective. Australian J. of Ag. Res. 45(3):487–509.
D'surney, S.J., J. T. Tschaplinski, N. T. Edwards, and L. R. Shugart. 1993. Biological responses of two soybean cultivars exposed to enhanced UVB radiation. Env. Exp. Bot. 33:347–356.
Dutky, E. M. 1999. Plant Pathology Update. College Park: University of Maryland.
Elkinton, J. S., A. E. Haje, G. H. Boettner, and E. E. Simmons. 1991. Distribution and apparent spread of Entomophaga maimaiga (Zygomycetes: Entomophthorales) in gypsy moth (Lepidoptera: Lymantriidae) populations in North America. Env. Ent. 20:1601–1605.
Elliott, H.J., R. Bashford, A. Greener, and S. G. Candy. 1993. Integrated pest management of the Tasmanian eucalyptus leaf beetle, Chrysophtharta bimaculata (Olivier) (Coleoptera: Chrysomelidae). For. Ecol. Man. 53:29–38.
EPA (US Environmental Protection Agency). 1997a. Pesticide Regulation (PR) Notice 97-3. Available. [Online]: http://www.epa.gov/opppmsd1/pr_Notices/pr97-3.html.
EPA (US Environmental Protection Agency). 1997b. Pesticide Fact Sheet, Spinosad. Available. [Online]: http://www.epa.gov/opprd001/factsheets/spinsoad.htm
EPA (US Environmental Protection Agency). 1998. Staff Background Paper #2.4 – General Overview: Reduced Risk Pesticide Program. Available. [Online]: http://www.epa.gov/oppfead1/trac/safero.htm
EPA (US Environmental Protection Agency). 1999a. Implementing the Food Quality Protection Act. EPA 735-R-99001. Washington, D.C.: EPA, Office of Pesticide Programs.
EPA (US Environmental Protection Agency). 1999b. Fact Sheets on New Active Ingredients. [Online]. Available: http://www.epa.gov/opprd001/factsheets.
EPA (US Environmental Protection Agency). 2000. Interim Fiscal Year 2000 Work Plan. Office of Pesticide Programs. [Online]. Available: http://www.epa.gov/opprd001/workplan/fy2000.htm
Eppes, J. In press. The Future of Biopesticides. Norfolk, Conn.: Business Communications Corporation.
Everitt, J. H., D. E. Escobar, R. Villarreal, M. A. Alaniz, and M. R. Davis. 1993. Integration of airborne video, global positioning system, and geographic information system technologies for detecting and mapping two woody legumes on rangelands. Weed Tech. 7:981–987.
Feldheim, K., and Conner, J. K. 1996. The effects of increased UV-B radiation on growth, pollination success, and lifetime female fitness in two Brassica species. Oecologia 106:284–297.
Fellowes, M. D. E., A. R. Kraaijeveld, and H. C. J. Godfray. 1998. Trade-off associated with selection for increased ability to resist parasitoid attack in Drosophila melanogaster. P Roy Soc Lond B Bio 265(1405):1553–1558.
Ferro, D. N. 1993. Integrated pest management in vegetables in Massachsetts. Pp. 95-107 in Successful Implementation Of Integrated Pest Management For Agricultural Crops, A. R. Leslie, and G. W. Cuperus, eds.. Boca Raton, Fla: Lewis Publications.
Ferro, D. N. and S. M. Lyon. 1991. Colorado potato beetle (Coleoptera: Chrysomelidae) larval mortality: operative effects of Bacillus thuringiensissubsp. san diego. J. Econ. Entomol. 84:806–809.
Ferro, D.N. and Gelernter, W.D. 1989. Toxicity of a new strain of Bacillus thuringiensis to Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Ent.. 82:750–755.
Fisher, J. R., J. J. Jackson and A. C. Lew. 1994. Temperature and diapause development in the egg of Diabrotica barberi (Coleoptera: Chrysomelidae). Env. Ent. 23(2):464–471.
Forgash, A. J. 1984. History, evolution, and consequences of insecticide resistance. Pestic. Biochem. Physio. 22:178–186.
Foster, G. G., W. G. Vogt, T. L. Woodburn, and P. H. Smith. 1988. Computer simulation of genetic control. Comparison of sterile males and field-female killing systems. Theor. Appl. Genet. 76:870–879.
Foyer, C. H. 1993. Interactions between electron transport and carbon assimilation in leaves: coordination of activities and control. Pp. 199-224 in Photosynthesis: Photoreactions to Plant Productivity, Y. P. Abrol, P. Mohanty, and Govindjee, eds. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Franco, C. M. M., and L. E. L. Coutinho. 1991. Detection of novel secondary metabolites. Crit. Rev. in Bio. 11:193–276.
Frankenhuyzen, K. U. 1993. The challenge of Bacillus thuringiensis. Pp. 193-220 in Bacillus thuringiensis, an Experimental Biopesticide: Theory and Practice, P. F. Entwistle, J. S. Cory, M. J. Bailey, and S. Higgs, eds. Chichester, New York: John Wiley & Sons.
fFrench-Constant, R. H. 1994. The molecular and population genetics of cyclodience insecticide resistance. Insect Biochemistry and Molecular Biology. 24(4):335–345.
Frei, B., and P. Schmid. 1997. Development trends in pesticide formulation and packaging. Pp. 34-43 in. Pesticide Formulation and Adjuvant Technology, C.L. Foy and D. W. Pritchard, eds. Boca Raton, Fla.: CRC Press.
Gelernter, W. D., N. C. Toscano, K. Kido, and B. A. Federici. 1986. Comparison of a nuclear polyhedrosis virus and chemical insecticides for control of the beet armyworm (Lepidoptera: Noctuidae) on head lettuce. J. Ec.. Ent.. 79:714–717.
Georghiou, G. P. 1986. The magnitude of the resistance problem. Pp. 14-43 in Pesticide Resistance: Strategies and Tactics for Management, National Research Council, ed. Washington, D.C.: National Academy Press:
Georghiou, G. P., and A. Lagunes. 1988. The Occurrence of Resistance to Pesticides: Cases of Resistance Reported Worldwide Through 1988. Rome: Food and Agriculture Organization of the United Nations.
Ghate, S. R., and C. D. Perry. 1994. Ground speed control of pesticide application rates in a compressed air direct injection sprayer. Trans. of ASAE. 37:33–38.
Gold, W. G., and M. M. Caldwell. 1983. The effects of ultraviolet-B radiation on plant competition in terrestrial ecosystems. Physiological Plantarum. 58:435–444.
Good, A. G., G. A. Meister, H. W. Brock, T. A. Grigliatti, and D. A. Hickey. 1989. Rapid spread of transposable P elements in experimental populations of Drosophila melanogaster. Genetics 122:387–396.
Gould, F. 1988. Evolutionary biology and genetically engineered crops. BioScience 38:26–31.
Gould, F. 1991. Evolutionary potential of crop pests. Amer. Sci. 79:496–507.
Gould, F. 1995. Comparisons between resistance management strategies for insects and weeds. Weed Tech. 9:830–839.
Gould, F. and P. Schliekelman. In press. Reassess autocidal control. In New technologies for Integrated Pest Management, G. G. Kennedy and T. Sutton, eds. St. Paul, Minn: APS Press
Grace, S. C., and B. A. Logan. 1996. Acclimation of foliar antioxidant systems to growth irradiance in three broad-leaved evergreen species. Plant Phys. 112:1631–1640.
Grinstein, A., and G. A. Matthews. 1997. Preface. Phytoparasitics. 25:Suppl:9S–10S.
Gueldner, R., C. Reilly, L. Pusey, C. Costello, R. Arrendale, R. Cox, R., D. Himmelsbach, G. Crumley, H. Cutler. 1988. Isolation and identification of iturins as antifungal peptides in biological control of peach brown rot with Bacillus subtilis. J. Ag. Food Chem., 36:366–370.
Hall, F.R., and R. D. Fox. 1997. The reduction of pesticide drift. Pp. 209-240 in Pesticide Formulation and Adjuvant Technology, C. L. Foy and D. W. Pritchard eds. Boca Raton, Fla.: CRC Press.
Hall, F. R., A. C. Chapple, R. A. Downer, L. M. Kirchner, and J. R. M. Thacker. 1993. Pesticide application as affected by spray modifiers. Pesticide Science: 38(2/3):123–133.
Hanks, J. E., and J. L. Beck. 1998. Sensor-controlled hooded sprayers for row crops. Weed Tech. 12:308–314.
Harris, M. K., ed. 1980. Biology and breeding for resistance to arthropods and pathogens in agricultural plants. Texas Agric. Exp. Stn. Pub. MP-1451.
Hastings, I. M. 1994. Selfish DNA as a method of pest control. Proc. Trans. Roy. Soc. London B 344:313–324.
Hatcher, P. E., and N. D. Paul. 1994. The effect of elevated UV-B radiation on herbivory of pea by Autographa gamma. Ent. Exp. Et App. 71:227–233.
He, H., Silo-Suh, L.A., Handelsman, J., Clardy, J. 1994. Zwittermicin A, an antifungal and plant protection agent from Bacillus cereus. Tet. Letters 35:2499–2502.
Heap, I. 2000. International Survey of Herbicide Resistant Weeds. March 23, 2000. [Online]. Available: www.weedscience.com.
Hedin, P. A., ed. 1983. Plant resistance to insects. ACS Symposium series 208. Washington, D.C.: American Chemical Society.
Henter, H. J., and S. Via. 1995. The potential for coevolution in a host-parasitoid system 1.1. Genetic variation within an aphid population in susceptibility to a parasitic wasp. Evolution. 49(3):427–438.
Hill, M. J., G. E. Donald, P. J. Vickery, and E. P. Furnival. 1996. Integration of satellite remote sensing, simple bioclimatic models and GIS for assessment of pasture suitability for a commercial grazing enterprise. Australian J. of Exp. Ag. 36:309–321.
Hinkle, M. K. 1982. Impact of the imported fire ant control programs on wildlife and quality of the environment. Pp. 41-43 in Proceedings of the Symposium on the Imported Fire Ant, June 7-10, 1981, Atlanta, Georgia, S. L. Battenfield, ed. Washington, D.C.: US Department of Agriculture.
Holt, J. S., S. B. Powles and J. A. M. Holtum. 1993. Mechanisms and agronomic aspects of herbicide resistance. Ann. Rev. Plant Physiol. Plant Molec. Biol. 44:203–229.
Hoy, M. A., R. D. Gaskalla, J. L. Capinera, and C. N. Keierleber. 1997. Laboratory containment of transgenic arthropods. Am. Entomol. 43:206-209, 255–256.
IANR (University of Nebraska Institute of Agriculture and Natural Resources). 1994. From Lab to Label. [Online]. Available: HtmlResAnchor http://ianrwww.unl.edu/ianr/pat/hortpara/sld031.htm (9/22/98).
Ignoffo, C. M., and C. Garcia. 1992. Combinations of environmental factors and simulated sunlight affecting activity of inclusion bodies of the Heliothis (Lepidoptera: Noctuidae) nucleopolyhedrosis virus. Env. Ent. 21:210–213.
Jansson, R. K., W. R. Halliday, and J. A. Argentine. 1997. Evaluation of miniature and high-volume bioassays for screening insecticides . J. Ec. Ent. 90:1500–1507.
Jaques, R .P. and Laing, D. R. 1989. Effectiveness of microbial and chemical insecticides in control of the Colorado potato beetle (Coleoptera: Chrysomelidae) on potatoes and tomatoes. Can. Ent. 121:1123–1131.
Jordan, N. R. 1993. Prospects for weed control through crop interference. Ec. App. 3:84–91.
Kaiser, K. 1978. The rise and fall of Mirex. Env. Sci. Tech. 12:520–528.
Kareiva, P. 1993. Agriculture – transgenic plants on trial Nature. 363:580–581.
Kareiva, P. 1996. Contributions of ecology to biological control. Ecology. 77:1963–1964.
Kerremans, P. and G. Franz. 1995. Isolation and cytogenetic analyses of genetic sexing strains for the medfly, Ceratitis capitata. Theor. Appl. Genet. 9:255–261.
Kidwell, M. G. and A. R. Wattam. 1998. An important step forward in the genetic manipulation of mosquito vectors of human disease. Proc. Natl. Acad. Sci. USA 95:3349–3350.
Kidwell, M. G., and J. M. C. Ribeiro. 1992. Can transposable elements be used to drive disease refractoriness genes into vector populations? Parasitol. Today 8:325–329.
Kline, W., K. Holmstrom, and S. Walker. 1996. ECB and CEW Mapping for Vegetable Crop IPM. Rutgers Cooperative Extension/Grant F. Walton Center for Remote Sensing & Spatial Analysis is now Utilizing GIS/GPS/RS Technologies for Precision Agriculture. [Online]. Available: http://deathstar.rutgers.edu/projects/gps/gps.html#IPM
Klinger, T., P. E. Arriola and N. C. Ellstrand. 1992. Crop-weed hybridization in radish (Raphanus sativus): effects of distance and population size. Am. J. Botany 79(12):1431–1435.
Kogan, M. 1982. Plant resistance in pest management. Pp. 93-134 in Introduction to Insect Pest Management, R. L. Metcalf and W. H. Luckman, eds. New York: John Wiley and Sons.
Kommedahl, T., and Mew, I. 1975. Biocontrol of corn root infection in the field by seed treatment with antagonists. Phytopathol. 65:296–300.
Kooman P.L., M. Fahem, P. Tegera, and A. J. Haverkort. 1996. Effects of climate on different potato genotypes to dry matter allocation and duration of the growth cycle. Eur. J. of Agr. 5(3-4):207–217.
Korzybski, T., Z. Kowszyk-Gindifer, and W. Kurylowicz. 1978. Section C: Antibiotics isolated from the genus Bacillus (Bacillaceae). In Antibiotics - Origin, Nature and Properties, Volume III. Washington, D.C.: American Society for Microbiology..
Krafsur, E. S., C. J. Whitten, and J. E. Novy. 1987. Screwworm eradication in North and Central-America. Parasitology Today. 35:131–137.
Krimsky, S.,and R. Wrubel. 1996. Agricultural Biotechnology and the Environment. Chicago, Ill: University of Chicago Press.
Krysan, J. L., D. E. Foster, T. F. Branson, K. R. Ostlie, and W. W. Crenshaw. 1986. Two years before the hatch: rootworms adapt to crop rotation. Bull. Entomol. Soc. Am. 32:250–253.
Lamoreaux, R. J. 1990. Herbicide discovery and development: Emphasis on groundwater protection . Crop Protection 13:483–487.
Larson, R. A. 1988. The antioxidants of higher plants. Phytochemistry. 27:969–978.
Lass W. L., and R. H. Callihan. 1997. The effect of phenological stage on detectability of yellow hawkweed (Hieracium pratense) and oxeye daisy (Chysanthemum leucanthemum) with remote multispectral digital imagery. Weed Technol. 11:248–256.
Lass, L. W., H. W. Carson, and R. H. Callihan. 1996. Detection of yellow starthistle with high resolution multispectral digital images. Weed Technol. 10:466–474.
Leason, M., Cunfliffe, D., Parkin, D., Lea, P.J., and Miflin, B.J. 1982. Inhibition of pea leaf glutamine synthetase by methionine sulphoximine, phosphinothricin and other glutamate analogues. Phytochemistry 21:855–857.
Liedtke, J. 1997. High Resolution Imagery Will Enhance the Effectiveness of Precision Farming. Ag Innovator. [Online]. Available HtmlResAnchor http://www.agriculture.com/ 01/11/97.
Lyttle, M. H. 1995. Combinatorial chemistry: A conservative perspective. Drug Development Research. 35:230–236.
Lyttle, M. H., M. D. Hocker, U. C. Hui, C. G. Caldwell, D. T. Aaron, A. Engquist-Goldstein, J. E. Flatgaard, and K. E. Bauer. 1994 Isozyme specific glutathione-S-transferase inhibitors: Design and synthesis. J. Med. Chem. 37:189–194.
MacIntosh, S. C., G. M. Kishore, F. J. Perlak, P. G. Marrone, T. B. Stone, S. R. Sims, and R. L. Fuchs. 1990. Potentiation of Bacillus thuringiensis insecticidal activity by serine protease inhibitors. J. Agric. and Food Chem. 38:1145–1152.
Madronich, S. 1993. The atmosphere and UVB radiation at ground level. Pp. 1-39 in Environmental UVB Photobiology, A. R. Young. New York: Plenum Press.
Manker, D. C., R. L. Starnes, M. G. Beccio, W. D. Lidster, S. C. MacIntosh, J. K. Swank, D. R. Jiménez, and D. R. Edwards. 1995. A synergistic metabolite from Bacillus thuringiensis with potential for use in agricultural applications. Invited Symposia Pacifichem ‘95 Honolulu, Hawaii, December 1995. Washington, D.C.: American Chemical Society.
Manker, D.C., B. D. Lidster, S. C. MacIntosh, and R. L. Starnes. 1994. Potentiator of Bacillus pesticidal activity, US Patent Application Serial # 08/146,852, PCT # WO 9409630. (May 11)
Marrone, P. G. 1999. Microbial pesticides and natural products as alternatives. Outlook on Ag. 28(3):149–154.\par Maxwell, B. D., M. L. Roush, and S. R. Radosevich. 1990. Predicting the evolution and dynamics of herbicide resistance in weed populations. Weed Tech: 4(1):2–13.
McCloud, E. S., and M. R. Berenbaum. 1994. Stratospheric ozone depletion and plant-insect interactions: effects of UV-B radiation on foliage quality of Citrus jambhiri for Trichoplusia ni. J. Chem. Ecol. 20:525–539.
McCoy, C. W. 1990. Ventomogenous fungi as microbial pesticides. Pp.139-159 in New Directions In Biological Control: Alternatives For Suppressing Agricultural Pests And Diseases, R. R. Basker and P. E. Dunn, eds. Proceedings of a UCLA Symposia on Molecular and Cellular Biology , Series v. 112.
McGaughey, W., F. Gould, and W. Gelernter. 1998. Bt resistance management. Nature Biotech. 16:144–146.
McKenzie, J. A. and A. Y. Game. 1987. Diazinon resistance in Lucilia cuprina: Mapping of a fitness modifier. Heredity 59:371–381.
Meister, G. A. and T. A. Grigliatti. 1994. Rapid spread of a P element/Adh gene construct through experimental populations of Drosophila melaogaster. Genome 36:1169–1175.
Metcalf, R. L. 1989. Insect resistance to insecticides. Pest.. Sci. 26:333–358.
Milner, J. L., L. Silo-Suh, J. C. Lee, H. He, J. Clardy, J. Handelsman. 1996. Production of kanosamine by Bacillus cereus UW85. Appl. Env.. Microb. 62:3061–3065.
Moar, W. J., and Trumble, J. T. 1987. Biologically derived insecticides for use against beet armyworm. Calif. Agr.. Nov-Dec:13-15.
Mochida, O. 1992. Management of insect pests of rice in 2000. Pp. 67-81 in Pest Management and the Environment in 2000, A. Aziz, S.A. Kadir, and H. S. Barlow, eds. Seminar Kuala Lampur, Malaysia.
Mooney, H. A., and W. Koch. 1994. The impact of rising CO2 concentrations on the terrestrial biosphere . Ambio 23:74–76.
Moore, J. 1991. Insecticide program stresses B.t. products. Am. Veg. Grower. Jan:32-34.
Moulin, A. P. 1996. Decision support systems and computer models, new tools for contemporary agriculture. Can. J. P1. Sci. 76:1.
Mulrooney, J. E., K. D. Howard, J. E. Hanks, and R. G. Jones. 1997. Application of ultra-low-volume malathion by air-assisted ground sprayer for boll weevil (Coleoptera: Curculionidae) control. J. Econ. Ent.. 90:639–645.
Murdoch, W. W., J.D. Reeve, and C.B.Huffaker. 1984. Biological control of olive scale and its relevance to ecological theory. Am. Nat. 123:371–392.
Murdoch, W.W., R.F. Luck, and S.L. Swarbrick. 1995. Regulation of an insect population under biological control. Ecology 76:206–217.
Murdoch, W.W. and C.J. Briggs. 1996. Theory for biological control: recent developments. Ecology 77:2001–2013.
Myers, J. H., A. Savoie, and E. Vanreden. 1998. Eradication and pest management. Ann Rev. Ent. 43:471–491.
NASA, 1998. Data And Information Services For Global Change Research. Sioux Falls, S.Dak.: Earth Observing System (EOS), EROS Data Center.
Niemela, P. and W. J. Mattson. 1996. Invasion of North American forests by European phytophagous insects —Legacy of the European crucible. Bioscience. 46:741–753.
NRC (National Research Council). 1986. Pp. 100-110 in Pesticide Resistance: Strategies and Tactics for Management. Washington, DC: National Academy Press
NRC (National Research Council). 1991. Neem: A Tree for Solving Global Problems. Washington, DC: National Academy Press.
NRC (National Research Council). 1996. Ecologically Based Pest Management: New Solutions for a New Century . Washington, DC: National Academy Press
NRC (National Research Council). 1997. Precision Agriculture in the 21st Century: Geospatial and Information Technologies in Crop Management. Washington, DC: National Academy Press.
Olson, K. E., S. Higgs, P. J. Gaines, A. M. Powers, B. S. Davis, K. I. Kamrud, J. O. Carlson, C.D. Blair, and B. J. Beaty. 1996. Genetically engineered resistance to dengue-2 virus transmission in mosquitoes. Science 272:884–886.
Orth, A. B., H. Teramura, and H. D. Sisler. 1990. Effects of ultraviolet-B radiation of fungal disease development in Cucumis sativus. Am. J. of Botany. 77:1188–1192.
Osburn, R.M., Milner, J.L., Oplinger, E.S., Smith, R.S., Handelsman, J. 1995. Effect of Bacillus cereus UW85 on the yield of soybean at two sites in Wisconsin. Plant Dis. 79:551–556.
Otvos, I.S., J. C. Cunningham, and W. J. Knapp. 1989. Aerial application of two baculoviruses against the western spruce budworm, Choristoneura occidentalis Freeman (Lepidoptera: Tortricidae) in British Columbia. Can. Ent.. 121:209–217.
Ozkan, E, ed. 1997. Equipment Developments. Chemical Application Technology. Newsletter 1(1):6p. [Online]. Available: http://www.ag.ohio-state.edu/~catnews/catequi.htm.
Paice, M. E. R, P .C.H. Miller, and W. Day. 1996. Control requirements for spatially selective herbicide sprayers. Comp. and Elect. in Ag. 14:163–177.
Panagopoulus, I., J. F. Borman, and L. O. Bjorn. 1991. Response of sugar beet plants to ultraviolet-B (280-320nm) radiation and Cercospora leaf spot disease. Physiologia Plantarum 84:140–145.
PANNA (Pesticide Action Network North America). 1998. PANUPS: Pesticide Updates – Asia, October 16, 1998. Adapted from Agrow: World Crop Protection News, 6/26/98 and 9/18/98 .
Parlow, J. J., D. A. Mischke, and S. S. Woodard. 1997. Utility of complementary molecular reactivity and molecular recognition (CMR/R) technology and polymer-supported reagents in the solution-phase synthesis of heterocyclic carboxamides. J. Org. Chem. 62:5908–5919.
Penuelas, J., and M. Estiarte. 1998. Can elevated CO2 affect secondary metabolism and ecosystem function? Trends Eco. Evo. 13:20–24.
Peterson, J. J. 1978. Development of resistance by the southern house mosquito to the parasitic nematode Romanomermis culicivorax. Env. Ent. 7:518–520.
Pettigrew, M. M., and S. L. O'Neill. 1997. Control of vector-borne disease by genetic manipulation of insect populations: technological requirements and research priorities. Aust. J. of Ent.. 36:309–317.
Peypoux, F., F. Besson, G. Michel, L. Delcambe, and B. Das, B. 1978. Structure de l'iturine C de Bacillus subtilis. Tetrahedron 34:1147–1152.
Pfadt, R. 1971. Fundamentals of Applied Entomology. New York: The Macmillan Company.
Pfeifer, T. A. and T. A. Grigliatti. 1996. Future perspectives on inspect pest management: engineering the pest . J. Invert. Pathol. 67:109–119.
Pimentel, D. 1995. Amounts of pesticides reaching target pests: environmental impacts and ethics. J. Ag. Env. Ethics. 8:17–29.
Pimentel, D., L. Lach, R. Zuniga, and D. Morrison. 2000. Environmental and economic costs of nonindignous species in the United States. Bioscience: 50(1):53–65.
Pimm, S. L., G. J. Russell, J.L. Gittleman, T. M. Brooks. 1995. The future of biodiversity. Science. 269(5222):347–350.
Plant, R.E. and N.D. Stone. 1991. Knowledge-based systems in agriculture. Toronto, Ontario: McGraw-Hill, Inc.
Podgwaite, J. D., R. C. Reardon, D. M. Kolodny-Hirsch, and G. S. Walton. 1991. Efficacy ofground application of the gypsy moth (Lepidoptera: Lymantriidae) nucleopolyhedrosis virus product, Gypchek. J. Econ. Ent. 84:440–444.
Porter, J. H., M. L. Parry, and T. R. Carter. 1991. The potential effects of climatic change on agricultural insect pests . Agric. Forest Meterol. 57:221–240.
Prout, T. 1978. The joint effects of the release of sterile males and immigration of fertilized females on a density regulated population. Theor. Popul. Biol. 13:40–71.
Purvis, A., P.M. Agapow, J.L. Gittleman, and G. M. Mace. 2000. Nonrandom extinction and the loss of evolutionary history. Science. 288(5464):328-330.
Quarles, William. 1996. New Microbial Pesticides for IPM. IPM Pract. 18(8), August:5-10.
Raffa, K. F. 1989. Genetic engineering of trees to enhance resistance to insects. BioScience 39:524–534.
Raymond, M., A. Callaghan, P. Fort and N. Pasteur. 1991. Worldwide migration of amplified insecticide resistance genes in mosquitoes. Nature 350:151–153.
Ribeiro, J. M. C. and M. G. Kidwell. 1994. Transposable elements as population drive mechanisms: specification of critical parameter values. J. Med. Entomol. 31:10–16.
Robertson, H. M, and Lampe, D. J., 1995. Distribution of transposable elements in arthropods. Annu. Rev. Entomol. 40:333–357.
Robertson, S. T. 1980. History of gas odorization. Pp. 1-5(a). in: Odorization, F. H. Suchomel and J. W. Weatherly III, eds.. Chicago, Ill.: Institute of Gas Technologies.
Rodgers, P.B. 1993. Potential of biopesticides in agriculture. Pestic. Sci. 39:117-129.
Rogers, H. H. and R. C. Dahlman. 1993. Crop responses to CO2 enrichment. Vegetatio 104/105:117–131.
Ros, J.,and M. Tevini. 1995. Interaction of UV-radiation and IAA during growth of seedlings and hypocotyl segments of sunflower. J. Plant Physiol. 146:295–302.
Rosenheim, J. A. 1998. Higher-order predators and the regulation of insect herbivore populations . Ann. Rev. Entomol. 43:421–447.
Roush, R. T. and W. M. Tingey. 1991. Evolution and management of resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Resistance ‘91—Achievements and Developments in Combating Pesticide Resistance, Rothamsted Experimental Station, 15–17 July.
Roush, M. L., S. R. Radosevich, and B. D. Maxwell. 1990. Future outlook for herbicide-resistance research. Weed Tech.: 4(1):208–214.
Sammons, A. E., C. R. Edwards, L. W. Bledsoe, P. J. Boeve, and J. J. Stuart. 1997. Behavioral and feeding assays reveal a western corn rootworm (coleoptera, chrysomelidae) variant that is attracted to soybean. Env. Ent. 25(6):1336–1342.
Sasaki, A., and H. C. J. Godfray. 1999. A model for coevolution of resistance and virulence in coupled host-parasitoid interactions. P Roy Soc Lond B Bio 266(1418):455–463.
Schoper, J.B., R. J. Lambert, and B. L. Vasilas. 1987. Pollen viability, pollen shedding, and combining ability for tassel heat tolerance in maize. Crop Sci. 27:27–31.
Schwinn, F. J. and H. V. Morton. 1990. Antiresistance strategies: design and implementation in practice. Pp. 170-183 in: Managing Resistance to Agrochemicals From Fundamental Research to Practical Strategies, M. B. Green, H. M. LeBaron and W. K. Moberg, eds. Washington, DC: American Chemical Society.
Sears, M. K., Jaques, R. P. and Laing, J. E. 1983. Utilization of action thresholds for microbial and chemical control of lepidopterous pests (Lepidoptera: Noctuidae, Pieridae) on cabbage . J. Econ. Ent.. 76:368–374.
Serebovsky, A. S. 1940. On the possibility of a new method for the control of insect pests . Zool. Zh. 19:618–630.
Sharga, B. M. 1997. Bacillus isolates as potential biocontrol agents against chocolate spot on Faba beans . Can. J. Microbio. 43:915–924.
Shelton, A. M. and J. A. Wyman. 1991. Insecticide Resistance of Diamondback Moth (Lepidoptera: Plutellidae) in North America. Proc. Second Internatl. Diamondback Moth Workshop . Taiwan.
Shemshedini, L. and T. G. Wilson. 1990. Resistance to juvenile hormone and an insect growth regulator in Drosophila is associated with an altered cytosolic juvenile hormone-binding protein. Proc. Natl. Acad. Sci. USA. 87:2072–2076.
Sholberg, P.L., Marchi, A., Berchard, J. 1995 Biocontrol of postharvest diseases of apple using Bacillus spp. isolated from stored apples. Can. J. Microbiol. 41:247–252.
Silo-Suh, L.A., B. Lethbridge, S. J. Raffel, H. He, J. Clardy, and J. Handelsman. 1994 Biological activities of two fungistatic antibiotics produced by Bacillus cereus UW85. Appl. Envi.. Microb. 60:2023–2030.
Sinkins, S. P., C. F. Curtis, and S. L. O Neill. 1997. The potential use of symbionts to manipulate arthropod populations . In Influential Passengers: Inherited Microorganisms And Arthropod Reproduction , S. L. O Neill, A. A. Hoffman, and J. H. Werren, eds. New York: Oxford Univ. Press.
Sinkins, S. P., H. R. Braig, and S. L. O'Neill. 1995. Wolbachia superinfections and the expression of cytoplasmic incompatibility . Proc. Roy. Soc. Lond B Bio. 261(1362):325–330.
Slansky, F. S., Jr. 1992. Allelochemical-nutrient interactions in herbivore nutritional ecology . Pp. 135-174 in Herbivores: Their Interactions with Secondary Plant Metabolites, Volume 2, Ecological and Evolutionary Processes, G. Rosenthal and M. Berenbaum, eds. New York: Academic Press.
Smith, K. P., M. J. Havey, and J. Handelsman. 1993. Suppression of cottony leak of cucumber with Bacillus cereus strain UW85. Plant Dis. 77:139–142.
Smits, P. H., I. P. Rietstra, and J. M. Vlak. 1988. Influence of application techniques on the control of beet armyworm larvae (Lepidoptera: Noctuidae) with nuclear polyhedrosis virus. J. Econ. Ent. 81:470–475.
Smits, P. H., M. C. van Velden, M. van de Vrie, and J. M. Vlak. 1987b. Feeding and dispersion of Spodoptera exigua larvae and its relevance for control with a nuclear polyhedrosis virus. Ent. Exp. Appl. 43:67–72.
Smits, P. H., M. van de Vrie, and J. M. Vlak. 1987a. Nuclear polyhedrosis virus for control of Spodoptera exigua larvae on glasshouse crops. Ent.. Exp. Appl., 43:73-80.
Spielman, A. 1994. Why entomological antimalaria research should not focus on transgenic mosquitoes. Parasitol. Today 10:374–376.
Stabb, E. V., L. M. Jacobson, and J. Handelsman. 1994. Zwittermicin A producing strains of Bacillus cereus from diverse soils. Appl. Env. Microb. 60:4404–4412.
Starnes, R. L., C. L. Liu, and P. G. Marrone. 1993. History, use and future of microbial insecticides. Am. Ent. 39:83–91.
Stetter, J. 1998. Pesticide innovation: trends in research and development. Med. Fac. Landbouwww. Univ. Gent.: 63(2):135–164.
Stewart, P A., and A. Sorensen. 2000. Federal uncertainty or inconsistency? Releasing the new agricultural-environmental biotechnology into the fields. Politics and Life Sciences 19(1): in press.
Sun, C. N., Y. C. Tsai and F. M. Chiang. 1992. Resistance in the diamondback moth to pyrethroids and benzoylphenylureas . Pp. 149-167 in Molecular Mechanisms of Insecticide Resistance: Diversity Among Insects , C. A. Mullin and J. G. Scott, eds., Washington, DC: American Chemical Society
Swanson, M. B., G. A. Davis, L. E. Kincaid, T. Schultz, J. E. Bartimess, S. Jones, and E. L. George. 1997. A screening method for ranking and scoring chemicals by potential human health and environmental impacts. Env. Tox and Chem. 16:372–383.
Taylor, A. G., and G. E. Harman.. 1990. Concepts and technologies of selected seed treatments. Ann. Rev. Phyto. 28:321–339.
Teramura, A. H. 1983. Effects of ultraviolet-B radiation on the growth and yield of crop plants. Phys. Plant. 58:415–427.
Teramura, A.H., and J. H. Sullivan. 1991. Potential impacts of increased solar UV-B on global plant productivity . Pp. 625-634 in Photobiology, E. Riklis, ed. New York: Plenum Press.
Tette J. P. and B. J. Jacobsen. 1992. Biologically intensive pest management in the tree fruit system. Pp. 57-83 in Food, Crop Pests and the Environment, the Need and Potential for Biologically Intensive Integrated Pest Management, Zalom F.G. and Fry, W. E., eds., St. Paul, Minn.: APS Press.
Tevini, M. 1993. Effects of enhanced UV-B radiation on terrestrial plants. Pp. 125-153 in UV-B Radiation and Ozone Depletion: Effects on Humans, Animals, Plants, Microorganisms, and Materials, M. Tevini, ed. Boca Raton, Fla: Lewis Publications.
Tevini, M., and A. H. Teramura. 1989. UV-B effects on terrestrial plants. Photochem. Photobiol. 50:479–487.
Thirumalachar, M., and M O'Brien. 1977. Suppression of charcoal rot in potato with a bacterial antagonist . Plant Dis. Reptr. 61(7):543–546.
Thomas, D.A., C. A. Donnelly, R. J. Wood, and L. S. Alphey. 2000. Insect population control using a dominant, repressible, lethal genetic system. Science 287(5462):2474–2476.
Thompson, G.D., J. D. Busacca, O. K. Jantz, and H. A. Kirst. 1994. Spinosyns: an overview of new natural insect management systems. Proceedings of the ESA Annual Meeting, Dallas, 13-17 December 1994 .
Trumble, J .T., and B. Alvarado-Rodriguez. 1993. Development and economic evaluation of an IPM program for fresh market tomato production in Mexico. Agric. Ecosys.and Env. 43:267–284.
Trumble, J. T. 1985. Integrated pest management of Liriomyza trifolii: influence of avermectin, cyromazine, and methomyl on leafminer ecology in celery. Ag. Ecosys. and Env. 12:181–188.
Trumble, J. T. 1989. Leafminer and beet armyworm control on celery. Final Report for 1988-9, Project #9, University of California at Riverside.
Trumble, J. T. 1990. Vegetable insect control with minimal use of insecticides. Hort. Sci. 25:159–164.
Trumble, J. T. 1991. California Tomato Board Layman's Summary 13-O.
Tumlinson, J. H., C. M. Demoraes, W. J. Lewis, P. W. Pare, and H. T. Alborn. 1998. Herbivore-infested plants selectively attract parasitioids. Nature 393(6685):570–573.
Turchin, P. 1999. Population regulation: a synthetic view. Oikos. 84:153–159.
UC Davis (University of California, Davis). 1997. Proceedings of the Workshop on Remote Sensing for Agriculture in the 21st century, October 1996. Available. [Online]: http://cstars.ucdavis.edu/proj/ag-21/ag21cen.html.
Ustin, S. L., D. A. Roberts, and Q. J. Hart. 1997. Seasonal vegetation patterns in a California coastal savanna derived from advanced visible/infrared imaging spectrometer (aviris) data . In Remote Sensing Change Detection: Environmental Monitoring Applications and Methods, Elvidge, C.D., and Lunetta, R., eds. Ann Arbor, Mich: Ann Arbor Press.
Utkhede, R., and J. Rahe. 1983. Interactions of antagonist and pathogen in biological control of onion white rot. Phytopath. 73:890–893.
Vanderplank, F. L. 1944. Hybridization between Glossina species and suggested new method for control of certain species of tsetse. Nature 154:607–08.
Vargas-Teran, M., B. S. Hursey, and E. P. Cunningham. 1994. Eradication of the screw-worm from Libya using the sterile insect technique. Parasitol. Today 10:119–122.
Vitousek, P. M., C. M. D'Antonio, L. L. Loope, and R. Westbrooks. 1996. Biological invasions as global environmental change. Am. Sci. 84:468–478.
Wagner, P. 1993. Techniques of representing knowledge in knowledge-based systems. Ag. Sys. 41:53–76.
Warr, W. A. 1997a. Combinatorial chemistry and molecular diversity: An overview. J. Chem. Inf. Comput. Sci. 37:134–140.
Warr, W. A. 1997b. Commercial software systems for diversity analysis. Persp. Drug Disc. Design.. 7/8:115–130.
Warwick, S. I. 1991. Herbicide resistance in weedy plants: physiology and population biology . Ann. Rev. Ecol. Syst. 22:95–114.
Watson, R. T., M. C. Zinyowera, R. H. Moss, eds. 1996. Climate Change 1995. Contribution of Working Group II to the Second Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press.
Weller, M. 1988. Biological control of soilborne plant pathogens in the rhizosphere with bacteria. Ann. Rev. Phyto. 26:379–407.
Werren, J. H. 1997. Biology of Wolbachia. Ann. Rev. Ent. 42:587–609.
Westbrooks, R. G. 1998. Invasive Plants: Changing the Landscape of American: Fact Book. Washington, DC: Federal Interagency Committee for the Management of Noxious and Exotic Weeds.
White, A. D., and H. D. Coble. 1997. Validation of HERB for use in peanut (Arachis hypogaea). Weed Technol. 11:573–579.
Whitten, M. J., and G. G. Foster. 1975. Genetical methods of pest control. Ann. Rev. Ent. 20:461–476.
Williams, C. M. 1967. Third-generation pesticides. Sci. Am. 217:13–17.
Wilson, E. O. 1993. Is humanity suicidal. Biosystems. 31(2-3):235–242.
Wood, M. 1992. Microbes blow those hornworms away. Ag. Res. June:4–7.
Yamamoto, H. 1985. Development of validamycin, its controlling effect on rice sheath blight. Jpn. Pest. Inf. 17:17.
Yamanaka, S., M. Hashimoto, M. Tobe, K. Kobayasii, J. Sekizawa, and M. Nishimura. 1990. A simple method for screening assessment of acute toxicity of chemicals . Archives of Tox. 64:262–268.
Zalom, F. G., and Fry, W. E. 1992. Biologically intensive IPM for vegetables. Pp. 107-165 in Food, Crop Pests and the Environment, the Need and Potential for Biologically Intensive Integrated Pest Management, F. G. Zalom, and W. E. Fry, eds., St. Paul, Minn.: APS Press.


































































