
6
What Determines Susceptibility to Nutrient Over-Enrichment?
KEY POINTS IN CHAPTER 6
To plan effective strategies for managing coastal nutrient over-enrichment, managers need to understand how different types of estuaries respond to nutrient inputs. This chapter reviews the wide variety of processes controlling the susceptibility of coastal systems to nutrient enrichment and discusses existing approaches to estuarine classification that may be useful in assessing susceptibility. It finds:
-
a widely accepted estuarine classification scheme is a prerequisite for a systematic approach to extending lessons learned and successful management options from one estuary to others;
-
such a classification scheme should allow categorization of relatively poorly known systems on the basis of a minimum suite of measurements;
-
quantitative classifications that provide insights into the relative importance of the different factors controlling estuarine dynamics have the most potential for predictive analysis; and
-
a high priority should be the development of a national framework of “index sites” within which there would be an integration and coordination of environmental monitoring and research, with the goal of developing a predictive understanding of the response of coastal systems to both nutrient enrichment and nutrient reduction.
Coastal zone managers strive to accommodate human actions while minimizing negative impacts on coastal ecosystems. Successful management requires considerable information at a variety of levels, including an understanding of systems in their natural, pristine condition as well as how natural systems respond to human
activities (Karr and Chu 1997). Coastal waters are particularly complex because different types of estuaries, embayments, and shelf systems differ in their responses to nutrient enrichment. This means that varying levels of nutrient input can cause very different responses in different systems.
This chapter examines classification schemes that could enhance attempts to understand, predict, and manage eutrophication and other impacts of nutrient over-enrichment in the nation’s coastal waters. It reviews existing and developing estuarine classifications. While the emphasis is on estuaries, where impacts of nutrient loading are most acute, the approach is equally appropriate for coastal systems in general, including those in the nearshore region of the continental shelf.
The diverse physical settings (defined by a number of parameters including geology, soil type, climatic setting, and topology) of estuaries and their watersheds give rise to different types of estuarine systems. While there are numerous similarities between all estuaries, there are also some basic differences. For example, both a drowned river valley estuary, such as Chesapeake Bay, and a bar-built estuary, such as Plum Island Sound in northeastern Massachusetts, have temporal and spatial patterns of salinity that reflect seasonal variations in freshwater discharge. However, while Chesapeake Bay is a deep-water, plankton-dominated system where waters have a long residence time, Plum Island Sound is a shallow, emergent, marsh-dominated system where waters have a short residence time. The expected quantitative values for indicators of ecological health or for susceptibility to nutrient over-enrichment are not the same for these two systems, even though many of the same biological or ecological attributes may work as indicators in these disparate situations. Knowledge of the physical setting and the undisturbed ecosystem condition must underpin any monitoring and management effort to restore a coastal system impaired by nutrient over-enrichment.
MAJOR FACTORS INFLUENCING ESTUARINE SUSCEPTIBILITY TO NUTRIENT OVER-ENRICHMENT
Certain key characteristics appear to be of primary importance in determining estuarine response to nutrient enrichment. These factors range from biotic factors to physical setting to hydrodynamic regime. Twelve of the most important factors are:
-
Physiographic setting. Characterization of the physiographic setting could include a geomorphic descriptor of an estuary (e.g., inverted continental shelf estuary like the Mississippi River plume, coastal embayment, and drowned river valley), a descriptor of the
-
major biological community(ies) (e.g., mangrove swamp, emergent marsh macrophyte, rocky intertidal, coral reef, and planktonic system), and a description of the biogeographic province as used by Hayden and Dolan (1976), Briggs (1974), and Gosner (1971). Physiographic setting largely determines the primary production base.
-
Primary production base. The term primary production base refers to various primary producers that have unique temperature, substrate, light, and nutrient requirements and thus respond differently to nutrient loading. Susceptibility will vary across estuaries with different primary production bases. Examples of major types of primary producer communities include: emergent marshes and swamps, attached intertidal algae, benthic microalgae, drifting macroalgae, seagrasses, phytoplankton, and coral.
-
Nutrient load. Nutrient load is the total amount of various nutrients contributed by the upstream landscape and atmosphere (Figure 6-1A&B). Coastal systems are among the most heavily loaded ecosystems on Earth. Even small nutrient losses per unit area of the terrestrial landscape become immense when scaled over the entire expanse of terrestrial watersheds.
-
Dilution. Dilution of watershed-derived nutrients occurs due to a variety of mixing processes upon entry into an estuary. It makes a difference whether a given nutrient load is distributed over 1 km2 versus 1,000 km2 or mixed into 106 versus 1010 m3. Dilution is one of the dominant factors used to predict lake eutrophication, and Nixon (1992) showed a strong relationship between areal nutrient load, which partially accounts for dilution, and primary production in a wide variety of estuaries. Areal nutrient load is the magnitude of the nutrient load (e.g., kg yr−1) scaled to the surface area of the receiving estuary (e.g., m2, thus kg m−2 yr−1). More recently the National Oceanic and Atmospheric Administration (NOAA) has incorporated estimates of dilution into their susceptibility classifications, using dissolved concentration potential and estuarine export potential.
-
Water residence time, TR, and flushing. Steady state conditions in a waterbody are affected by the fluxes into and out of the system. Residence time of water in an estuary or part of an estuary is an important temporal scale for relating physical phenomena to
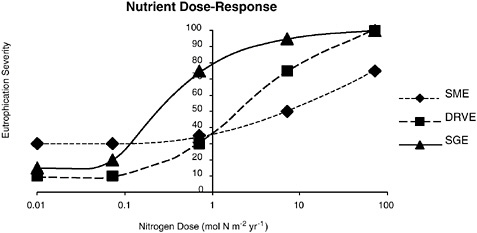
FIGURE 6-1 As nutrient loading is increased over the range of globally observed levels, it is hypothesized that different types of estuaries differ in their susceptibility to eutrophication. (A) This figure shows hypothetical Dose-Response Curves for three major types of coastal systems: Salt Marsh Dominated Estuary (SME), Plankton Dominated Drowned River Valley Estuary (DRVE), Seagrass Dominated Estuary (SGE). At the lowest levels of nitrogen loading (0.01 to 0.01 moles N m−2 yr−1), each of the systems is likely to be oligotrophic (low level of productivity). Salt marsh estuaries are naturally more productive than seagrass dominated and plankton dominated drowned river valley estuaries. Seagrass dominated estuaries are likely to be the most sensitive to nutrient enrichment, shown by the rapid rise in eutrophication severity as nitrogen loading is increased from 0.1 to 1.0 moles N m−2 yr−1. Salt marsh estuaries are expected to be the least sensitive to nutrient enrichment, which is illustrated by the slow rise in eutrophication severity only after nitrogen loading exceeds 1.0 moles N m−2 yr−1 (unpublished figure by C. Hopkinson).
-
ecological processes related to nutrient loading1 (Malone 1977; Cloern et al. 1983; Vallino and Hopkinson 1998; Howarth et al. 2000) (Box 6-1). For example, phytoplankton blooms can occur only when the plankton turnover time is shorter than the water
|
1 |
Many estuaries can be described as hypersaline (Chapter 5). Thus, some consideration was given to including salinity as one discriminator of estuarine class. However, because both dilution and water residence time play a role in determining salinity, it was felt that adding salinity as a specific discriminator of estuarine class would be redundant. |
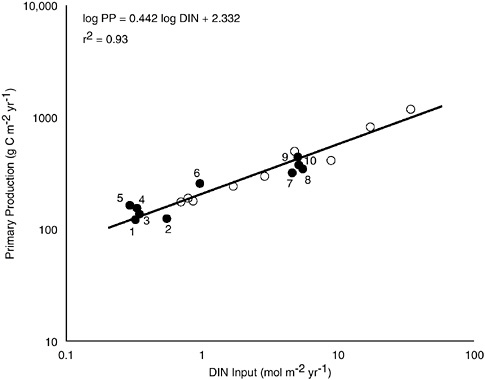
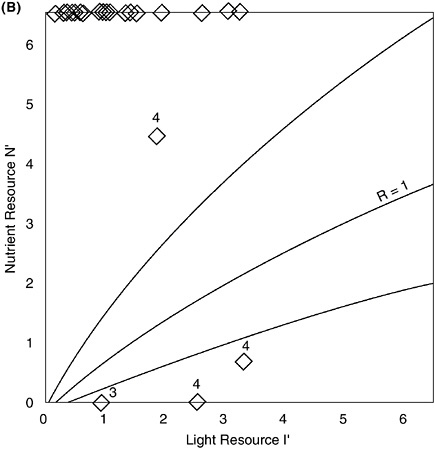
(B) Primary production by phytoplankton (14C uptake) as a function of the estimated rate of input of dissolved inorganic nitrogen (DIN) per unit area in a variety of marine ecosystems. The open circles are for large (13 m3, 5 m deep) well-mixed mesocosm tanks at the Marine Ecosystem Research Laboratory (MERL) during a multi-year fertilization experiment (Nixon et al. 1986; Nixon 1992). Natural systems (solid circles) are: (1) Scotian shelf, (2) Sargasso Sea, (3) North Sea, (4) Baltic Sea, (5) North Central Pacific, (6) Tomales Bay, California, (7) continental shelf off New York, (8) outer continental shelf off southeastern U.S., (9) Peru upwelling, (10) Georges Bank (modified from Nixon et al. 1996).
-
residence time. If both water residence time and phytoplankton turnover time are one day, there is no chance of a bloom; algae are flushed from the system as fast as they multiply. Alternatively, if the residence time is seven days and phytoplankton turnover time is one day, phytoplankton can double seven times prior to being exported and an initial algal population of 5 μg chl-a l−1 can become a 640 μg l−1 bloom, given no other losses. There are other ecological processes whose time scales also can be compared to residence time to determine their potential influence on eutrophication. The
|
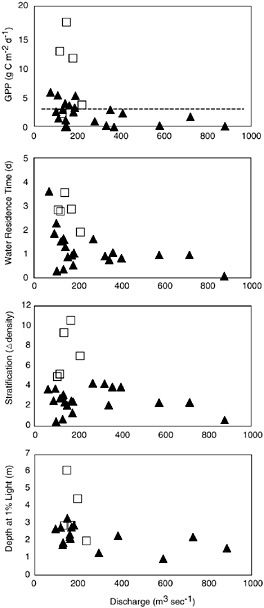
BOX 6-1 Estuaries vary greatly in their susceptibility to eutrophication (Bricker et al. 1999). The Hudson River estuary receives extremely high inputs of nutrients, both from wastewater treatment plants in New York City and environs and from nonpoint sources in the watershed. However, several studies in the estuary during the 1970s showed fairly low rates of primary production (Malone 1977; Sirois and Fredrick 1978). The low production, despite high nutrients, resulted from short water residence times and perhaps light limitation from relatively deep mixing of the water column. During many summers in the 1990s, the freshwater discharge into the estuary was less than seen during the 1970s (Howarth et al. 2000). This increased the water residence time, increased the stratification in the estuary, and also led to greater water clarity due to less input of sediment and/or lessened resuspension of bottom sediments (Figure 6-2). Thus, not only did phytoplankton have longer to grow before being advected out of the estuary, but light limitation was lessened, increasing growth rates. The result is the estuary became much more productive. During many times in the 1990s, rates of production were high enough to classify the estuary as highly eutrophic (Howarth et al. 2000). Thus, climatic variation can make an estuary more or less susceptible to eutrophication. Future climate warming in the northeastern United States is likely to result in lessened freshwater discharge (Moore et al. 1997), aggravating eutrophication in the Hudson and similar estuaries with short water residence times (Howarth et al. 2000). |
-
turnover time of organic nitrogen (i.e., conversion to inorganic nitrogen) in comparison to residence time can indicate whether this might be an important source of nitrogen fueling phytoplankton growth. For example, if residence time is seven days, organic nitrogen compounds with a lability (i.e., able to be decomposed or remineralized) or turnover time greater than 14 days are not likely to be remineralized to the inorganic form, and be available to phytoplankton while within the estuary. The fraction of total nitrogen input to estuaries from land and the atmosphere that is exported varies as a function of water residence time in the system, as is the fraction of input that is denitrified in estuaries (Nixon et al. 1996). NOAA has incorporated estimates of flushing in their development of an estuarine susceptibility index.
-
Stratification. Stratification is an important physical process affecting eutrophication. Stratification can maintain phytoplankton in the nutrient rich, photic zone (Malone 1977; Howarth et al. 2000) and isolate deeper waters from reaeration. Most hydrodynamic classifications include a measure of stratification intensity (Hansen and Rattray 1966). NOAA considers stratification to be an important component of their developing estuarine susceptibility index.
-
Hypsography. Hypsography describes the relative areal extent of land surface elevation, and might be a useful indicator of estuarine susceptibility to nutrient enhanced eutrophication. Knowledge of the relationship between estuarine area and elevation/depth will indicate the percentage of area potentially colonizable by emergent marsh, intertidal flats, submerged aquatic vegetation, phytoplankton, macroalgae, etc. Overlaid with measures of water turbidity and stratification, it might be possible to illustrate the spatial extent of sites potentially susceptible to a variety of eutrophication symptoms.
-
Grazing of phytoplankton. Grazing by benthic filter feeders acts to clear particles from the water column, and can limit the accumulation of algal biomass (Cloern 1982). Alpine and Cloern (1992) showed that filter feeding benthos in San Francisco Bay effectively decreased the estuarine response to nutrient loading (in terms of phytoplankton production). There is some conjecture as to the importance of what were once vast filter feeding oyster populations in Chesapeake Bay and whether these acted to decrease the intensity of phytoplankton blooms in the past (Newell 1988). Zoo-plankton grazing can exert a strong influence on phytoplankton
-
blooms and eutrophication symptoms in lakes, but this phenomenon remains relatively unexplored in coastal systems (Ingrid et al. 1996). Likewise, the feeding activity of top predators, which can “cascade” down to influence zooplankton sizes, abundances, and grazing pressure on phytoplantkon in estuaries (Carpenter and Kitchell 1993), remains poorly understood.
-
Suspended materials load and light extinction. Suspended load and light are two important factors related to nutrient over-enrichment (Box 6-2). Light is a primary factor controlling primary production. Researchers have shown that light can play a critical role in determining the response of estuarine systems to nutrient loading (e.g., Cloern 1987, 1991, 1996, 1999). In northern San Francisco Bay, high turbidity from watershed sediment erosion reduces light levels to such an extent that primary production is light-limited year round. A new conceptual model of coastal eutrophication (Cloern 1999) suggests that it is the interaction of nutrient loading and other stressors/factors that determines estuarine response.
-
Denitrification. Denitrification is the process whereby nitrate is converted to gaseous nitrogen and N2O, and thereby made biologically unavailable. Denitrification provides a sink for nitrogen in estuarine systems; it essentially counteracts allochthonous nutrient inputs to estuaries and thereby can reduce eutrophication responses. Denitrification has been shown to be proportional to the rate of organic nitrogen remineralization in sediments (Seitzinger 1988), which is coupled with the magnitude of primary production that is oxidized by the benthos (Nixon 1981; Seitzinger and Giblin 1996). The relationship between denitrification and eutrophication is not simply linear. There are potential indirect effects of eutrophication that limit denitrification. For example, bottom water anoxia limits nitrification and hence denitrification in sediments and bottom waters. High sulfide concentrations, which are also associated with anoxic conditions, inhibit nitrification as well (Joye and Hollibaugh 1995). Knowledge of the magnitude of denitrification can help predict the eutrophication response of an estuary because nitrogen that is denitrified is largely unavailable to support primary production.
-
Spatial and temporal distribution of nutrient inputs. Distribution of nutrient inputs varies along the expanse of an estuary (Vallino and Hopkinson 1998). The potential effect of nutrient
|
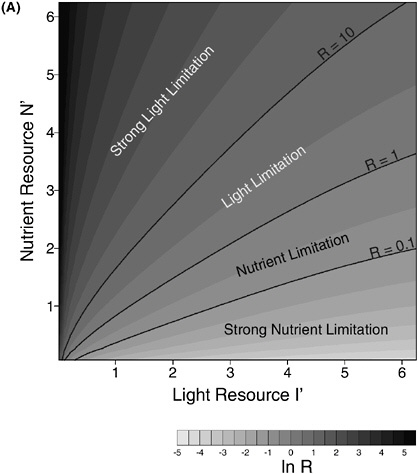
BOX 6-2 The quantity of nutrients into an estuarine system is not the only factor affecting susceptibility. Many estuaries maintain low algal biomass and low primary production under nutrient-rich conditions, such as North San Francisco Bay. How is it that some highly nutrient loaded systems do not show symptoms of eutrophication? Through research aimed at understanding the mechanisms controlling bloom dynamics in the San Francisco Bay system, Dr. James Cloern and colleagues at the U.S. Geological Survey (USGS) in Menlo Park, California, have developed an index of the sensitivity of a particular estuarine system to changes in nutrient concentration (Cloern 1999). The index is based on a model of phytoplankton population growth, where growth is the product of the carbon assimilation rate and the ratio of chlorophyll to carbon. The model includes functional responses of population growth to photosynthetic efficiency, light availability, temperature, photo-adaptation and nutrient availability. Because light energy can be a major resource that can and often does limit algal growth and production in estuarine ecosystems, a phytoplankton resource limitation plot, with light resource plotted against nutrient resource, indicates whether phytoplankton growth rate is more sensitive to changes in light or nutrients. Where the ratio of growth-rate sensitivity to light and nutrients, R, is greater than one (R>1) growth rate is strongly limited by light availability; ratios less than 1 (R<1) indicate strong nutrient limitation (Figure 6-3A&B). The boundary line between light and nutrient limitation shifts with changes in the physiological state of the phytoplankton. Thus it is possible to use measures of the light and nutrient resources in an estuary to make judgments about the relative strength of light and nutrient limitation over time and space. The light-nutrient limitation index is appealing for its simplicity. Using this approach, assessments of the sensitivity of an estuary to nutrient loading can be done using measurements of turbidity, light, and nutrient concentrations. The approach has been validated in applications to numerous estuaries in North America and Europe that demonstrate a range of temporal and spatial variations in the magnitude of nutrient and light limitation. Assessment results could be used to guide management strategies for individual systems. For instance, management strategies that emphasize nutrient reductions would be high priority in estuaries classified as nutrient sensitive; other strategies would be more important in estuaries classified as nutrient insensitive. The light-nutrient sensitivity index is but one tool available to assess and control the eutrophication threat to coastal ecosystems (Cloern 1999). Other important factors include the effects of residence time and phytoplankton grazing. Furthermore, the index gives no information about harmful algae, macroalgae, or seagrass epiphytes. Because of its simplicity, robustness, and ease of application, however, even with its limitations this index makes a significant contribution to developing a classification of estuarine sensitivity to nutrient enrichment that can be used to help manage nutrient over-enrichment problems. |
-
inputs will also vary depending on the location of the input. Seasonal variation in agricultural activity (such as fertilizer application, fossil fuel combustion, or precipitation) results in changes in nutrient loads that a recovery body may see during the year. Furthermore, variation in the load contributed by one of many tributaries may also vary, resulting in seasonal and geographic variability.
-
Allochthonous organic matter inputs. Organic matter contributes directly to eutrophication. The relative magnitude of inorganic versus organic nitrogen load influences the balance between autotrophic and heterotrophic metabolism (Hopkinson and Vallino 1995). The relative magnitude of dissolved versus particulate organic matter loads influences residence time of inputs, as particles are preferentially trapped by processes operating in the estuarine turbidity maximum and by gravity. The carbon:nitrogen stoichiometry of organic matter remineralized by the benthos and denitrification further influence the balance between autotrophic and heterotrophic processes in estuaries. Algal blooms are an example of an autotrophic process and net oxygen uptake is an example of a heterotrophic process.
Recognizing that there are basic differences in estuarine susceptibility to nutrient over-enrichment, the development of a conceptual framework or classification scheme for organizing how to think about susceptibility will enable scientists and managers to better understand and predict the effects of human activities on estuarine and coastal ecosystems, and thus to more effectively manage human activities. Variations in the 12 factors discussed above result in different responses. Hence, systematic variation in these 12 factors can result in systematic responses. Thus, coupling an understanding of how these factors vary from estuary to estuary of known load-response behavior can lead to a predictive framework or classification scheme.
It is not adequate to understand eutrophication and other processes in a few, well-studied coastal systems. Useful understanding will require a systematic means of extending the results from one coastal system to others that have not been studied extensively. There are too many estuaries in North America to carry out comprehensive ecosystem studies of all those undergoing nutrient enrichment, and scientists and managers do not presently have a broad enough understanding of estuarine and coastal processes to choose representative systems for detailed analysis. Development of a scheme for classification of estuarine and coastal systems is a prerequisite to understanding and reducing the effects of nutrient over-
enrichment. A classification system would provide a language with which to describe the similarities and differences between systems. By enabling researchers to apply more effective and rigorous hypothesis testing, such a classification scheme could provide greater guidance for focused and effective research.
COASTAL CLASSIFICATION
Scientists and resource managers have used classification systems for decades to organize information about ecological systems. Yet the classification of estuarine and coastal systems remains a difficult topic because they exhibit such dynamic changes in time and space. As discussed earlier, each estuary or coastal system possesses a set of characteristics (e.g., morphology, river flow, tidal range, circulation, productivity, etc.) that are controlled, to a large degree, by local geology and climate. A classification scheme should have the ability to:
-
encompass broad spatial and temporal scales,
-
integrate structural and functional characteristics under different disturbance regimes,
-
convey information about mechanisms controlling estuarine or coastal features, and
-
accomplish its goal at low cost with a high level of uniform understanding among resource managers.
A useful classification scheme should allow classification of relatively unknown systems on the basis of a minimum suite of measurements (Jay et al. 1999) obtainable from climate records, maps, remote sensing, or ocean monitoring.
To envision how a useful scheme might be constructed, an examination of existing schemes and their value for understanding estuarine response to nutrient loading is warranted. In reviewing existing estuarine classification systems, it is important to bear in mind that only in the past decade or so, however, have classification systems been developed to increase our understanding and ability to predict the effects of enhanced nutrient delivery to coastal ecosystems.
Most classifications of estuaries are based upon physical parameters and geomorphic characteristics. Three basic types of estuarine classification include geomorphic, hydrodynamic, and habitat enumeration approaches.
GEOMORPHIC CLASSIFICATION
Pritchard (1952, 1967) and Dyer (1973) used a geomorphic approach to classify estuaries. From a geomorphological standpoint, Pritchard identified four primary subdivisions of estuaries: 1) drowned river valleys (e.g., Chesapeake Bay), 2) fjord-type (e.g., Penobscot Bay), 3) bar-built (e.g., Laguna Madre), and 4) estuaries produced by tectonic processes (e.g., San Francisco Bay). This approach has intuitive appeal, but it generally lacks a quantitative framework allowing further elaboration. Thus, this approach is of limited usefulness in understanding, predicting, and managing estuary response to nutrient loading.
HYDRODYNAMIC CLASSIFICATION
The hydrodynamic approach to estuarine classification focuses on the interaction in narrow estuaries of tidal currents and river flow. Tidal currents provide energy for mixing while river flow is a source of stratification or buoyancy. Stommel and Farmer (1952) divided estuaries into 4 categories based on stratification: 1) well mixed, 2) partially mixed, 3) fjord-like, and 4) salt wedge. This simple classification was made quantitative by defining a stratification number G/J, where G is energy dissipation over a defined channel length and J is the rate of gain of potential energy of water moving through the estuary over the same length (Ippen and Harlemann 1961; Prandle 1986). By incorporating Richardson number and critical depth criteria, Fischer (1976), Simpson and Hunter (1974), and Nunes Vaz and Lennon (1991) have made variations on this classification.
The one-parameter (e.g., G/J) classifications capture one important aspect of estuarine circulation but they have no direct relation to the various estuarine types (Jay et al. 1999). Accordingly, two-parameter classifications were developed that describe the interaction of geomorphology, fresh water, and tides. The most widely known two-parameter scheme (Figure 6-4; Hansen and Rattray 1966) employs two parameters to classify estuarine circulation: 1) a stratification parameter, ∂S/S0 and 2) a circulation parameter, Us/UF. The first parameter describes stratification as the ratio of the top-to-bottom salinity difference to mean salinity over the section. The circulation parameter is a ratio of the net surface current to the mean freshwater velocity through the section. Numerous clarifications, modifications, and additions have been made to this approach, most focused on providing a closer connection between the density field and tidal processes (Fischer 1976; Officer 1976; Oey 1984).
Hansen and Rattray (1966) identified seven types of estuaries, basically following the conventional usage of Stommel and Farmer, but
further differentiating physically significant differences of regime. Type 1(a+b) estuaries have net flow seaward at all depths and upstream salt transfer controlled by diffusion (1a−well-mixed with slight salinity stratification, 1b−strong stratification). Type 2(a+b) estuaries have net flow reversal at depth with both advection and diffusion contributing to upstream salt flux (2a−slight stratification, 2b−strong stratification). Type 3(a+b) is distinguished from type 2 primarily by the dominance of advection in accounting for salt flux. (Type 3b estuaries are so deep that the salinity gradient and circulation do not extend to the bottom) (i.e., fjords). In Type 4 estuaries (salt wedge), the stratification is still greater and the flow grades from a thick upper layer flowing over a thin lower layer to a shallow surface layer flowing with little influence over a deep lower layer. The Hansen and Rattray (1966) circulation-stratification diagram illustrates that a range of circulation patterns and estuarine morphologies is possible for a certain degree of stratification (Box 6-3 and Figure 6-4).
HABITAT CLASSIFICATION
There is a long history of classifying environments on the basis of their plant community composition or sediment characteristics. In the early 1900s there were several attempts to classify wetlands, especially peatlands of Europe and North America, on the basis of the combined chemical and physical conditions of the wetland along with the vegetative community description. These early classifications served as models for more inclusive classifications developed by the U.S. Fish and Wildlife Service in the 1950s and 1970s.
In the 1950s, the U.S. Fish and Wildlife Service developed a classification scheme to inventory the distribution, extent, and quality of remaining wetlands in relation to their value as wildlife habitat. Twenty types of wetlands were described, including “coastal saline areas.” Four overall categories were arranged by life forms of vegetation in order of increasing water depth or frequency of inundation. The scheme was elegantly simple, with salinity being the sole chemical criteria. The classification became known as the Circular 39 Classification (Shaw and Fredine 1956).
In 1979, the National Wetlands Inventory classification was adopted as the primary wetland classification scheme for U.S. wetlands. Developed by the U.S. Fish and Wildlife Service, the classification scheme included deepwater habitats and wetlands (Cowardin et al. 1979). The classification and following inventory was intended to describe ecological taxa, to arrange them in a system useful for resource managers, to provide units for mapping, and to provide uniformity of concepts and terms. The classification has a hierarchical approach that uses systems, subsystems, classes, subclasses, dominance types, and special modifiers to more pre-
|
BOX 6-3 Hansen and Rattray (1966) developed a classification scheme with relatively simple parameters and good predictive ability to deal with salt transport mechanisms. The scheme allows calculation of the relative importance of diffusive (tidal) salt transport relative to total landward salt flux needed to maintain the salt balance. This kind of predictive ability is important for understanding relationships between nutrient loading and eutrophication processes. Since the original work was done, others have improved the parameterization of tidal processes. Jay and Smith (1988) directly represented the forcing of residual circulation. Friederichs and Madsen (1992) suggested a modification to address the influence of tidal flats. Hearn (1998) considered the effect of surface heating and evaporation on narrow Mediterranean estuaries. Because of the types of forcing factors chosen as parameters, classification schemes based on Hansen and Rattray are restricted to narrow embayments, fjords, and river-estuaries where these factors predominate. Systems dominated by wind forcing, such as broad, shallow embayments, and plumes, are unsuitable because they are too wide to have substantial river flow per unit width or they have no lateral boundaries. The Hansen and Rattray (1966) approach has been criticized because it fails to incorporate the inherent variability within individual estuaries and in the same estuary at different times. As new hydrodynamic classification schemes are developed, they should incorporate variability. Examples of such variability include spring-neap variability of stratification and variability associated with freshwater inputs. One measure of variability might include the ratio of cross-estuary to along-estuary salinity gradients (Geyer et al. 1999). By addressing estuarine variability as part of estuarine classification, such systems will better articulate the connections between estuarine structure and estuarine processes. |
cisely define wetlands and deepwater habitats. A stated goal is to eventually inventory the wetlands across the United States at a scale of 1:24,000.
The National Wetlands Inventory classification includes four levels, three of which include coastal habitats: marine, estuarine, and riverine. Subsystems that are of interest to coastal scientists and managers include subtidal and intertidal regions. The lowest hierarchical level, the modifier level, more precisely describes the water regime, salinity, pH, and soil.
A few habitat-based classification schemes have included forcing function criteria. Odum et al. (1974) developed a classification and functional description of coastal ecosystems that included major forcing functions and stresses that influence the distribution of systems. A hierarchical approach was used as opposed to a quantitative approach, and thus the forcing functions and stresses were not parameterized.
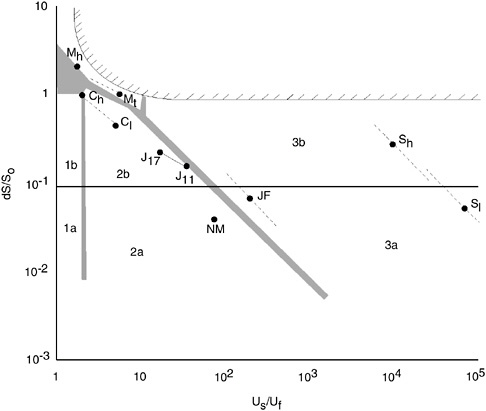
FIGURE 6-4 An example of a physically-based estuarine classification. Hansen and Rattray developed a classification for differentiating estuaries on the basis of factors controlling circulation. The first factor describes the degree of estuarine stratification, which is related to the difference in surface and bottom water salinity (∂S/S0), and the second factor describes the relative importance of freshwater flow on net flow (Us/Uf). This figure shows how several North American estuaries differ according to these criteria. There is a gradient of increasing stratification from Type 1 to Type 4 estuaries. The Columbia River (Ch and Cl) is a type 1 or 2 estuary depending on time of year and freshwater runoff; the James (J17 and J11) and Narrows of the Mersey (NM) estuaries are Type 2 estuaries; the Strait of Juan de Fuca (JF) and Silver Bay (Sh and Sl) systems are Type 3; and the Mississippi River (Mh and Ml) is a Type 4 estuary. Subscripts h and l refer to high and low river discharge conditions, respectively. Recent research on estuarine susceptibility to eutrophication suggests that stratification is a major determinant in estuarine response to nutrient loading. The Hansen and Rattray classification enables stratification to be quantified and thus may be useful in developing an eutrophication classification (modified from Hansen and Rattray 1966).
The objective of most habitat type classification schemes is to impose boundaries on natural ecosystems for the purposes of inventory, evaluation, and management. This approach works well for aiding in the inventory and evaluation of wetlands as well as estuarine habitats, but it generally lacks a logical or quantitative framework. The use of the approach as a tool to assess susceptibility to nutrient loading is unproven.
HYBRID CLASSIFICATION
Jay et al. (1999) borrowed from a highly effective hierarchical geomorphic classification scheme with a proven predictive ability for fluvial systems (Montgomery and Buffington 1993) to develop a geomorphic estuarine classification with a hierarchical structure. Several recent fluvial classification systems are based on a hierarchical ranking of linkages between the geologic and climatic settings, the stream habitat features, and the biota (Hawkins et al. 1993). The geomorphic and climatic processes that shape the abiotic and biotic features of streams provide a conceptual and practical foundation for understanding the structure and processes of fluvial systems. Furthermore, an understanding of process allows streams to be viewed in a larger spatial and temporal perspective, and to infer the direction and magnitude of potential changes due to natural and human disturbances. A stream classification system based on patterns and processes and how they are expressed at different temporal and spatial scales can aid successful management (Rosgen 1994).
The Jay et al. (1999) classification for estuaries provides a means to identify environments found in various types of estuarine subsystems, relates estuarine types to dominant sediment transport processes, and allows a prediction as to whether sediment transport is limited by transport capacity or sediment supply for coarse and fine sediments. Linkage with hydrodynamic classification schemes is through the non-dimensional hydrodynamic parameters associated with each sediment transport forcing mode. Six transport processes are parameterized: 1) net motion of river flow (QR), 2) oscillatory tidal flow (QT), 3) internal circulation or buoyancy forcing (QI), 4) atmospherically forced circulation (QA), 5) transport and resuspension by wind waves and swell (QW), and 6) transport by sea ice (QH). Each forcing process has a representative time scale and descriptive non-dimension hydrodynamic parameters responsible for distinctive modes of sediment transport. Extension of this hierarchical geomorphic classification to address issues of ecological importance such as eutrophication has not been attempted, but the inclusion of particle trapping and residence time processes suggests that this may be a profitable avenue for further investigation.
THE NATIONAL OCEANIC AND ATMOSPHERIC ADMINISTRATION’S NATIONAL OCEAN SERVICE CLASSIFICATION SCHEMES
NOAA’s National Ocean Service have been working to develop methods to gauge the susceptibility of estuaries to nutrient over-enrichment for over a decade. The first index developed, the “dissolved concentration potential” (DCP), integrated nutrient loads with an estimate of estuarine dilution and flushing. The dilution parameter is proportional to estuarine volume and the flushing parameter is calculated with the Ketchum (1951) fractional freshwater method, which is derived from the replacement of the freshwater component of the total system volume by river flow.
where Qf is freshwater discharge and Vf is freshwater volume in the estuary, Vt is total estuarine volume and ![]() is mean nutrient load for all estuaries.
is mean nutrient load for all estuaries.
The DCP provides an estimate of average nutrient concentration throughout an estuary assuming there is no biological processing. Systems with a high DCP tend to concentrate nutrient inputs, while systems with a low DCP strongly dilute or flush nutrients. NOAA categorizes estuaries as having low, medium, and high susceptibility to nutrient loading on the basis of DCP concentration, less than 0.1 mg l−1, 0.1 to 1.0 mg l−1, and greater than 1.0 mg l−1, respectively. It would be interesting to determine the degree to which a measure of eutrophication severity such as phytoplankton production or standing crop correlates with measures of nutrient load and DCP. This index has not been quantitatively compared to the Nixon model (Nixon 1992, 1997; Nixon et al. 1996), which is based solely on areal nutrient loading, in its ability to predict eutrophication.
DCP provides a quantitative measure of estuarine susceptibility to nutrient loading and is based on physical criteria including estuarine volume, volume of freshwater, and freshwater inputs. An underlying assumption is that the system is vertically homogenous (i.e., does not account for stratification). It could be applied at various times of the year to assess how susceptibility varies temporally. DCP relies exclusively on freshwater input as the mechanism for flushing. Flushing would be underestimated in systems where tides or winds are primary mechanisms controlling mixing, such as in many lagoonal systems. Spatial gradients in flushing cannot be accommodated with the DCP approach.
The “estuarine export potential” (EXP) is a second generation classifi-
cation developed by NOAA to predict estuarine response to nutrient loading. EXP defines the relative capacity of estuaries to dilute and flush dissolved nutrient loads. It addresses several deficiencies of the DCP index by incorporating aspects of stratification and tidal range. In its present configuration, EXP is not a quantitative index; rather it categorizes systems into low, medium, and high flushing and dilution potentials. The EXP index estimates dilution potential from measures of estuarine volume and the presence of stratification, and it estimates flushing potential from measures of tidal range and the ratio of river runoff to estuarine volume. The method uses a decision-rule process and a combination of qualitative and quantitative measures. This approach has intuitive appeal (as does the geomorphic classification scheme), but it generally lacks a formal quantitative framework allowing further elaboration. While the relative simplicity of the approach can be seen as a strength (i.e., data from a few estuaries can be applied to a large number of estuaries), the semi-quantitative parameterization limits the predictive ability.
NOAA staff, in conjunction with leading estuarine scientists, have applied the EXP scheme to the 138 estuaries included in the National Estuarine Eutrophication Assessment (Box 6-4). Overall, EXP was found to be useful in developing an assessment of eutrophication susceptibility, being in the “ballpark” for about 85 percent of the estuaries. Problems in prediction were focused mainly in a few Maine estuaries, small estuaries in southern California, and the Puget Sound estuaries. The next logical step would be to quantify the extent to which the ability to predict estuarine susceptibility has improved with further development of the EXP index. Analyses of the relationship between areal nutrient loading, EXP, and trophic state would illustrate whether predictive ability has improved relative to the first approximation presented by Nixon (1992).
Results produced using the EXP approach are currently being reviewed by experts, while the National Ocean Service pursues more rigorous coupling of EXP with National Estuarine Eutrophication Assessment results. At the same time, the National Ocean Service is refining the approach to help increase both spatial and temporal resolution. This will enable the National Ocean Service to evaluate the relative susceptibility of various regions within estuaries during a range of runoff conditions (Bricker et al. 1999).
EXP refinements under consideration include the addition of factors thought to influence estuarine susceptibility, including temperature (to address biotic differences between biogeographic provinces), the importance of wind mixing, inlet configuration, estuarine plume exchange with nearshore oceanic water, and the ratio of shoreline length to estuarine surface area (which is believed to correlate with the importance of intertidal wetlands). NOAA also plans to explore spatial and temporal vari-
|
BOX 6-4 Estuaries can be classified based on physical transport processes that, in part, determine their susceptibility to nutrient-related water quality conditions. An index to quantify the transport processes, EXP, was developed using physical and hydrologic data, assembled by NOAA’s National Ocean Service for 138 estuaries in the conterminous United States. As a first approximation, the EXP index classifies an estuary’s susceptibility to nutrient-related water quality concerns using two key physical factors: the dilution capacity of the water column and its flushing/retention time (as discussed earlier in this chapter, other factors can play an important role in determining susceptibility to eutrophication). Dilution capacity is determined by the volume of water available to dilute nutrient supplies. In vertically homogenous estuaries, the dilution volume is equal to the estuary volume. In contrast, for vertically stratified systems the dilution volume is limited to the upper layer of the water column. Flushing is the time required for freshwater inflow and tidal prism volume (modified by a re-entrainment coefficient) to replace the estuary volume. The index represents the average annual and system-wide conditions, providing an order-of-magnitude separation for the 138 coastal systems studied. Figure 6-5 provides some examples of results using this approach to classification. The results indicate that there are substantial differences among the 138 estuaries. Dilution volume ranges over five orders of magnitude and flushing time ranges just under five orders of magnitude. Systems with relatively large volumes and short flushing times, such as large river systems (e.g., Columbia and Mississippi Rivers) are less susceptible to eutrophication due to nutrient loading. Systems with moderate volumes and long flushing times, such as Chesapeake Bay, are more susceptible to eutrophication. One way to apply the susceptibility concept is to couple EXP with nutrient load estimates from each estuarine watershed (Figure 6-6). This provides a predicted nitrogen concentration in the water column that suggests, in a comparative sense, the potential for nutrient-related water quality symptoms. For example, higher nutrient concentrations imply the potential for more extreme expressions of nutrient-related symptoms. Coupling EXP with nutrient load estimates also has the potential to suggest how responsive the system may be to additional nutrient loads or nutrient abatement strategies. For example, estuaries in the upper left portion of Figure 6-6 would have to add or reduce comparatively more nutrients to affect water column concentrations than estuaries in the lower right. Likewise, this work may begin to describe how changes to an estuary’s physical environment could potentially alter its susceptibility to nutrient-related conditions. For example, the dilution or flushing components of EXP could be affected by alterations in freshwater inflow (e.g., diversions, impoundments, or consumptive loss) or tidal exchange (e.g., inlet modification, channel dredging). |
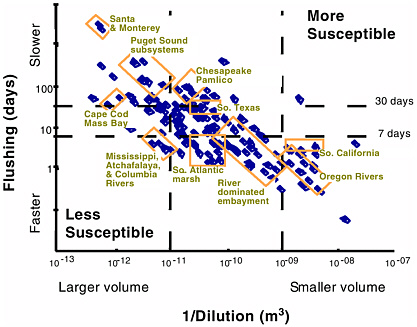
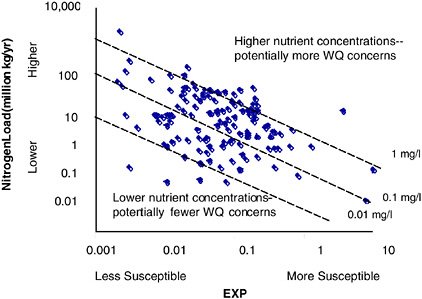
ability of EXP in single systems. These efforts, when coupled with efforts to model nutrient loading to estuaries and more rigorous quantification of tidal and stratification parameters should improve the capability to predict estuarine susceptibility to nutrient enrichment (Bricker et al. 1999).
Most recently, NOAA has been addressing some of the deficiencies of the DCP index to incorporate measures of tidal flushing and stratification. Tidal flushing is addressed by incorporating a measure of the tidal prism into the overall flushing calculation, and stratification is addressed by
calculating dilution (for stratified systems) on the basis of the freshwater fraction rather than total estuarine volume. Using updated information on nitrogen loading, calculated using USGS’s Spatially Referenced Regressions on Watersheds (SPARROW) model and estuarine volumes, some patterns have been revealed in plots of revised DCP versus nitrogen load for many of the estuaries included in NOAA’s National Estuary Eutrophication Assessment survey (Bricker et al. 1999; Box 6-4).
In this work, geographically and geomorphologically similar systems tended to cluster. For instance, estuaries in Maine fell out as a cluster as did large rivers, mid-Atlantic lagoons, and south-Atlantic marsh dominated estuaries. It is not obvious why these groupings occur, but this behavior may reflect similarities in stratification and circulation in local regions or it may reflect basic differences in physiography or primary production base (e.g., salt marsh dominated lagoons versus plankton dominated drowned river valleys). Initial comparisons to trophic state showed considerable deviations from predictions. For example, three clusters predicted to range from low to high susceptibility have been observed to be moderate to highly eutrophic. Again, an explanation for these patterns is not obvious and perhaps suggests further basic differences between various types of estuaries beyond that captured by measures of nutrient loading, dilution, and flushing. Perhaps a different set of rules governs the behavior of salt marsh and phytoplankton dominated estuaries. The DCP revisions summarized here represent preliminary results. They have not been thoroughly reviewed, but were presented to the committee to illustrate some of the directions NOAA’s National Ocean Service is pursuing to increase understanding of susceptibility to nutrient loading.
NEXT STEPS
NOAA’s DCP and EXP classification schemes are unique in that they were developed for the sole purpose of eutrophication understanding and prediction. However, neither of these schemes (including the revised DCP), have been tested rigorously to determine their ability to predict estuarine susceptibility to enhanced nutrient loading. Although physically based classification schemes have been useful in describing aspects of estuarine circulation, they have not been used systematically to understand responses to nutrient loading. For example, no classification scheme has been developed that encompasses the myriad of factors thought to be important eutrophication controls. While a statistical approach might be useful for identifying the suite of factors that best explain variability in eutrophication or other adverse impacts from nutrient over-enrichment across estuaries, such a relationship might not elucidate the actual mechanisms controlling eutrophication and thus we lose information perhaps
critical to managing and reversing eutrophication trajectories. Improved classifications need to be developed that can be generalized to a broader range of features and processes relevant to estuarine ecosystems, especially those affecting the susceptibility of various estuaries to nutrient over-enrichment.
A Proposal to Select and Use Coastal Index Sites
There are too many estuaries in the United States for the nation to conduct comprehensive ecosystem studies of all those affected by nutrient enrichment. Although the federal government conducts monitoring activities at more than 15,000 sites nationwide (Pryor et al. 1998), these efforts are not sufficiently coordinated to provide a predictive understanding of the causes and effects of nutrient enrichment. Because it is generally understood that different types of coastal systems differ in their response to nutrient enrichment, it would be extremely useful for managers to have a framework of dose-response curves for each of the major types of coastal systems. With such a tool, coastal managers could predict the effects of both increased and reduced nutrient inputs. Thus, a system is needed to classify coastal systems into a number of major types that are likely to respond similarly to nutrient enrichment. Resource managers could then apply these unique dose-response curves to their estuary’s particular conditions.
When considering questions about how to improve the integration of environmental monitoring and research across the nation’s many existing networks and programs, the Environmental Monitoring Team of the Committee on Environment and National Resources (a committee of the Executive Office’s National Science and Technology Council; Pryor et al. 1998) produced a three-tiered conceptual framework describing how federal environmental monitoring activities can fit together. The first tier includes inventories and remote sensing; the second includes national and regional surveys; the third tier includes intensive monitoring and research sites, or “index sites.” The goal was to integrate activities across tiers and thus provide the understanding that will enable sound evaluation of the status, trends, and future of the environment. (This approach also is similar to one proposed by NOAA, the Environmental Protection Agency [EPA], and USGS in their draft coastal research and monitoring strategy.)
The Committee on the Causes and Management of Coastal Eutrophication recommends adoption of this three-tiered framework as a way of better integrating monitoring and research in support of improved management of coastal ecosystems. Of primary importance, the committee suggests that monitoring and research be conducted at a sufficiently high spatial and temporal resolution at Tier III “index sites” to develop predic-
tive, cause-effect or dose-response models for the nation’s major types of coastal systems.
The establishment of a national framework of index sites where monitoring and research are closely integrated would lead to the development of a predictive understanding of coastal system responses to anthropogenic activities, especially nutrient enrichment. At index sites, intensive monitoring and research activities would lead to a broad understanding of how and why estuaries respond as they do to nutrient inputs. Research should not be restricted to increased nutrient loading scenarios, but should also examine responses to nutrient loading reductions. We expect a different set of dose-response curves for nutrient reductions that would incorporate time lags, hystereses, and non-linear responses of biological systems. Predictive models would be developed at index sites. Index sites should be established for each of the major types of estuaries; we expect unique dose-response curves for each estuarine type. Examples of possible Tier III index sites would be the coastal long-term ecological research sites. Coastal long-term ecological research sites are funded by the National Science Foundation (NSF) at Plum Island Ecosystem in northern Massachusetts, Baltimore Ecosystem Study, Virginia Coast Reserve, Santa Barbara Channel, Everglades, and Georgia Coastal. Each estuarine type should possess a unique dose-response curve that relates primarily to variations in the major factors controlling estuarine susceptibility to nutrient over-enrichment (i.e., loading, dilution, and flushing).
How best to identify the major types of coastal systems is elusive. Based on our analysis, this committee believes that a combination of physiographic province and primary production base could serve as the key criteria for selecting the major types of coastal systems. Following this thinking, and based on physiographic characteristics, the twelve major types of coastal systems are:
-
open continental shelf (e.g., Georgia Bight, Monterey Bay);
-
coastal embayment (e.g., Massachusetts Bay, Buzzards Bay, Long Island Sound);
-
river plume (inverted) estuary (e.g., Mississippi River plume);
-
coastal plain or drowned river valley estuary (e.g., Chesapeake Bay, Hudson River, Charleston Harbor);
-
coastal plain salt marsh estuary (e.g., Plum Island Sound, North Inlet, Duplin River);
-
lagoon (e.g., Padre Island, Pamlico Sound);
-
fjord estuary (e.g., Penobscot Bay);
-
coral reef system (e.g., Kaneohe Bay);
-
tectonically caused estuary (e.g., San Francisco Bay, Tomales Bay);
-
large river, non-drowned river estuary (e.g., Columbia River);
-
seagrass dominated estuary (e.g., Tampa Bay); and
-
rocky intertidal, macroalgae dominated estuary (e.g., Casco Bay).
How to identify research teams qualified to conduct process-oriented research at the index sites is equally elusive. To make major advances in understanding coastal systems and in predicting the effects of increased nutrient loading will require interdisciplinary research coordinated among investigators working within the index sites. Research should emphasize major ecological questions that stress linkages between terrestrial and coastal ecosystems. The research should seek to understand the causes of major ecological and environmental changes, including eutrophication and how populations, communities, and ecosystems of the coastal systems respond to these changes. Research at index sites should include experimental studies across a range of appropriate spatial and temporal scales. Comparative approaches encompassing parallel studies in different coastal systems are likely to provide important insight in how systems respond to nutrient enrichment. There should be close coupling between experimental, descriptive, and comparative research, with simulation modeling used to guide the research and to facilitate comparison with research in other systems. Finally, for the research to be of public value, there is a need for the detailed, process-based models to be abstracted to “simple” dose-response curves that can be easily applied by coastal resource managers at the local level.
Index site research teams should be selected on a peer-reviewed competitive basis, similar to that employed by NSF for the selection of recent coastal long-term ecological research sites. There are probably only 12 to 24 research groups around the country qualified to conduct this type of research, including academic and federal groups. For index site research to be successful, the highest selection priority should be on the originality and quality of the research proposal and research group, followed by representation of the major estuarine types, by research site characteristics and suitability for conducting eutrophication research, and finally by geographic spread.
Development of predictive, mechanistic models requires the integration of process-oriented research with comparative studies of estuaries (Geyer et al. 1999). It is not adequate to understand eutrophication processes in only the few, well-studied index estuaries; however, a systematic means of extending the results from one estuary to others that have not been extensively studied is also required. Understanding of processes resulting from the detailed studies in index estuaries can be tested and broadened through comparisons conducted in other estuaries of similar “type” but which represent the range of physical, hydrological, and biological characteristics. Within each “type,” responses are expected to vary
according to the major factors that control the response to nutrient addition such as dilution, freshwater input, flushing due to gravitational, tidal and wind driven circulation, stratification, water clarity/turbidity, denitrification, and biological control. Tier II coastal systems should be used as sites where comparative research can be conducted. Tier II systems might include estuaries such as those in NOAA’s National Estuarine Research Reserves and EPA’s National Estuary Programs. At these sites, research is conducted and data collected at much lower temporal and spatial resolution. Integration within a national program, however, would ensure collection of data necessary for testing the predictive models developed at index sites. Using information from existing programs can be cost-effective, but this is not always the ideal approach because many of these programs do not administer their own monitoring but instead rely on state agencies. Also, research projects can be short term, whereas a long-term perspective is of critical importance.
The result of monitoring, research, and modeling conducted within a three-tiered national framework would be a series of dose-response curves tailored to each major type of coastal system (Figure 6-5). Specific dose-response curves tailored to individual estuaries on the basis of their unique characteristics could then be applied by local and state resource managers hoping to control or reverse eutrophication trends.
ADDITIONAL QUESTIONS
An understanding of the response of coastal waters to nutrient loading is developing slowly. Following the scientific lead of freshwater ecologists modeling the trophic state of lakes, marine ecologists are developing a predictive understanding of some of the key parameters controlling estuarine response to nutrient loading. There are numerous models that relate eutrophication or primary production to single variables, such as the filter feeding benthos and light availability (Alpine and Cloern 1992; Cloern 1999). Nixon (1992) described a strong relationship between rate of estuarine phytoplankton production and areal rate of nutrient loading. However, the extreme variation in response to any level of loading clearly demonstrates the importance of other factors that determine differences between estuaries. The next level of understanding may well result from incorporation of additional factors into models. Several groups around the world are taking the next steps of incorporating measures of circulation, stratification, mixing, dilution, and turbidity into their eutrophication models. NOAA is updating its DCP and EXP measures of estuarine susceptibility with current data and comparing predictions to their national dataset of estuarine trophic state. By incorporating new measures for estuarine susceptibility, predictions of estuarine response to
|
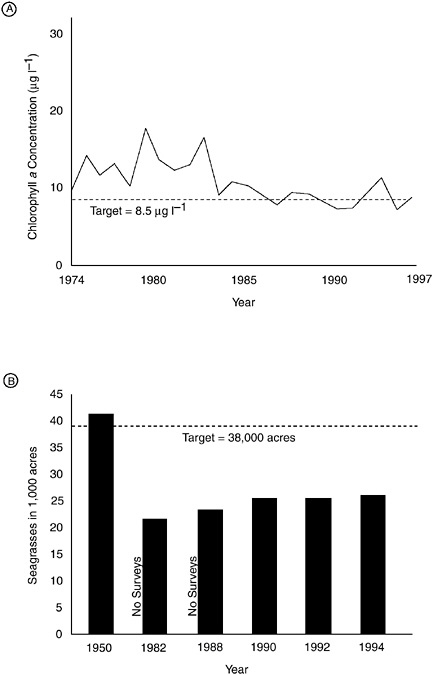
BOX 6-5 Tampa Bay, Florida, is a seagrass dominated estuary that has also experienced nitrogen source reductions and concomitant reversals in eutrophication. Impacts to Tampa Bay from increasing population and industrial development resulted in high algal biomass and large seagrass reductions during the 1960s and 1970s. By 1982, seagrass coverage was only 72 percent of earlier estimates. However, in 1996 seagrass coverage had increased by 25 percent since 1982 (Ries 1993; Johansson and Greening 2000). Nutrient loading reduction strategies were initiated in 1980, and since then, nitrogen inputs from sewage treatment plants have been reduced by 50 percent. As indicated in Figure 6-7, the pattern of eutrophication has been reversed in a time consistent with nutrient load reductions. Chlorophyll a concentrations began to decrease within three to five years of nutrient reductions and are now fluctuating close to targets set by resource managers. Seagrasses have taken longer to recover, lagging nutrient reduction by about eight years, but since 1988 coverage has been increasing about 200 hectares annually. With a management target of 15,378 hectares, recovery will take another 25 years to complete if recovery continues at present rates. The causes of lags are uncertain, but are thought to include continued release of nitrogen from internal nitrogen stores accumulated during earlier years of accumulation, and in the case of seagrass recovery, continued high epiphytic growth and high water column turbidity due to sediment resuspension in areas devoid of seagrass. It will be important to monitor Tampa Bay into the future to continue learning how the system responds, and how increased urban runoff from development, the growing role of atmospheric deposition from fossil fuel combustion, and complicating factors like dredging affect it. |
nutrient loading based on areal loading alone may be improved (Nixon 1992).
Scientists and managers are equally concerned about reversing the eutrophication trend observed in many of the nation’s estuaries. Whether the same classification schemes being developed to predict effects of increasing nutrients will work equally well in predicting reversals, if nutrient inputs are reduced, remains to be determined. As some studies have indicated, there are non-linearities and thresholds in eutrophication response to increased nutrient loading. There are undoubtedly additional non-linearities and thresholds governing the response of estuarine systems to reductions in nutrient loading. While loading, dilution, mixing, flushing, circulation, and stratification may be the key parameters governing the initial response of a system to nutrient increases, internal stores of organic matter and internal nutrient cycling processes might be important factors governing eutrophication reversal. There are few locations where nitrogen loading reductions have occurred and even fewer where resultant changes in estuarine trophic status have been chronicled.
The Himmerfjärden, an estuary of the Swedish central Baltic coast, is one example where eutrophication has been reversed following reductions in nitrogen loading. Present nitrogen loads are less than 10 percent of pre-treatment input levels. Primary production, chlorophyll concentrations, and water transparency have all improved since treatment began. In this system, the reversal showed little lag in response following source reductions, presumably reflecting the rocky nature of the coastal zone and small internal stores of organic nutrients (Granéli et al. 1990; Elmgren and Larsson 1997).
Tampa Bay, Florida, is another coastal system where local managers are reversing eutrophication. In this seagrass-dominated estuary, however, significant nutrient reductions have not resulted in eutrophication reversals to the extent observed in the Himmerfjärden. Time lags in recovery are hypothesized to result from large internal sources of detrital nitrogen accumulated over years of earlier eutrophic conditions. Recovery is likely to be slow until these historic stores of nitrogen are reprocessed and either flushed from the system or denitrified. Box 6-5 describes the Tampa Bay reversal in greater detail.
































