
4
Characteristics of Sites for Astrobiological Investigation
Earth is the only planet with irrefutable evidence of life. When Earth formed some 4.55 billion years ago, it was devoid of life, but probably within the first billion years it went through a transition from a planet featuring solely abiotic processes to one featuring both abiotic and biotic processes. This transition did not occur with any certainty on Mars or on any other planetary body. However, astrobiology investigations of Mars seek to determine if life is or ever was present there, and if not, why not. Consequently, astrobiological studies of Mars require investigations that target both abiotic and putative biotic processes (including the possibility that martian life originated elsewhere in the solar system or beyond, as has been suggested by some researchers). Studies of abiotic processes help establish a baseline against which all putative biotic processes can be evaluated. Of particular interest are those abiotic processes that could lead to organic synthesis, polymerization, and the emergence of life—a progression of processes generally called prebiotic chemistry. The focus of this chapter is on the characteristics of sites and samples that are most likely to yield robust evidence of prebiotic chemistry, the preservation of biosignatures from past life, and the existence of present life.
THE SEARCH FOR SIGNS OF PREBIOTIC CHEMISTRY
As noted in Chapter 3, chemistries that may have led to life (prebiotic chemistry) include Miller-Urey type atmospheric electric discharges, serpentinization reactions of Fe-Mg minerals (e.g., olivine), Strecker synthesis, Fischer-Tropsch synthesis, and FeS-driven synthesis. The fundamental step in prebiotic chemistry is the formation of compounds with C-C bonds from single carbon molecules (e.g., CO, CO2, CH4). Therefore, the best sites to search for clues about prebiotic chemistry would be those where organic carbon is most likely to be preserved. However, as discussed below, no rich deposits of inorganic carbon have been identified on Mars.
The next section describes those sites that may be the best targets to search for biosignatures of past life, including organic carbon. The proposed target mineralogies and lithologies, which include putative hydrothermal systems, evaporites, sediments, iron oxides, and clay minerals, are the best targets to find possible evidence of
prebiotic chemistries. Of these potential sites to preserve biosignatures, the most likely sites are sediments, evaporites, and hydrothermal systems based upon observations of terrestrial rock systems.
THE SEARCH FOR PAST LIFE
The committee approaches a consideration of martian sites from a process-oriented perspective and discusses first the processes most likely to generate and preserve biosignatures. As pointed out in Chapter 3, biosignatures can be molecular, isotopic, morphological, mineralogical, or elemental in nature. This section finishes with a discussion of the characteristics of specific sites where biosignatures of past life might be found.
Processes That May Lead to the Preservation of Biosignatures on Mars
Entombment
Entombment involves processes characterized by rapid mineralization which preserve microorganisms and organic molecules against degradation. Such processes are especially important for reduced compounds preserved in highly oxidizing environments. Examples of processes leading to entombment include the following:
-
Evaporation. Evaporation leads to mineralization driven by increases in solute concentrations as water is removed. Minerals often crystallize on nuclei provided by microbial cells or organic particles. The relatively rapid precipitation that can occur in evaporative settings often captures particles and compounds within the mineral matrix. In some cases, such entrapped particles can provide evidence of biogenicity.
-
Freezing. Concentration of brine solutions by freezing is also a likely process of evaporite salt formation and entombment of particles. Its effectiveness as a preservation mechanism may be enhanced by the low temperatures experienced on Mars.
-
Temperature and pressure changes. Supersaturation due to cooling often leads to precipitation in and around hot springs. Supersaturation can also occur because of changes in pressure. The rate at which precipitation occurs can be rapid enough to entomb living cells or to protect biomolecules. For example, some micro- and macrostructures in and around hot spring deposits show morphological relationships that are uniquely biogenic.1 The precipitation that occurs owing to supersaturation can also occur in the subsurface where hydrothermal flow is driven by a buried heat source. The emplacement of igneous intrusives such as dikes infiltrating sedimentary rocks can generate subsurface hydrothermal flow and enhance mineralization reactions. For example, chert, a common authigenic mineral formed by hot springs in igneous terrains, can often preserve biosignatures.
-
Diffusion-driven reactions. Concretions are precipitates that occur within a stratigraphic horizon in response to variations in the concentrations of solutes. While the precipitation of the concretion is controlled by the degree of supersaturation and the diffusional gradients, the kinetics of concretion formation are not well known. Precipitation processes related to concretions can entomb potential biosignatures.
Enrichment of Organic Biomarkers
Enrichment processes include all phenomena that lead to enhanced concentrations of compounds or particles. The search for biosignatures on Mars may be limited to sites with local enrichment of biosignatures, given that analytical equipment may be hampered by detection limits. Sedimentary rocks in general often enrich biomarkers on Earth, while iron oxides and clays, wherever they are found, can incorporate organic molecules.
-
Sedimentary rocks. Sedimentation may enhance biosignature preservation in a number of ways. Sediments can provide an environment in which biomass accumulates and is buried. When sedimentation occurs as a pelagic process, settling particles may concentrate microbes that are scavenged from the water column.2 Sediments may also preserve morphological biosignatures: Trace fossils, wrinkle marks, stromatolites, and microbialites are identifiable sedimentary forms thought to involve the interaction between sediments and microbiota.
-
Iron oxides. Iron oxides are common minerals formed at the surface of Earth and Mars that are likely to enrich for specific types of organic molecules.3 Organic acids may be enriched on the surface of these particles. Furthermore, organics adsorbed to iron oxides may be stable in low-pH environments.
-
Clays. Clays preferentially adsorb organic molecules at both their surface and within the interlayer. Absorption can occur in low- or high-pH systems.
Sites and Samples for Biosignatures of Past Life
The processes that lead to preservation of biosignatures on Earth are far from fully understood. Although researchers expect their martian counterparts to follow the same general sets of processes, the conditions on Mars that may have led to biosignature preservation are poorly constrained, and hence, the matrix of target environments is necessarily varied, including sites and samples with an uncertain potential for success.
Sites Targeted for Entombment of Biosignature Molecules
A strategy to investigate martian biosignatures may focus on sites where the protection of biomolecules from degradation is expected. Three types of systems and specific sites are discussed below:
-
Hydrothermal deposits. Because hot springs are commonly characterized by mineral supersaturation and deposition, they would be good target sites for preservation of biosignatures. However, such hydrothermal systems have not been definitively identified on Mars via remote sensing or in situ imaging. Nonetheless, proposed sites of hydrothermal activity include areas near impact craters, slopes of volcanic structures such as Hecates Tholus and Ceraunius Tholus,4 and locations where ice melting may be occurring or have occurred.5,6 In addition, rift systems, such as the Cerberus Fossae, may have erupted both water and lava.7 These locations may also be sites favorable for the process of rapid mineralization and preservation of biomolecules.
-
Evaporites. Evaporation is known to occur on Mars, as evidenced by the sulfate-rich deposits analyzed by the rover Opportunity in Meridiani Planum8 and by detection from orbit of several thick deposits of sulfate minerals such as kieserite and gypsum, particularly in Valles Marineris.9 Evaporite deposits might be likely sites for entombed organic particles.
-
Concentrations of concretions. Concretions (termed “blueberries”) are well documented at Meridiani from investigations by the rover Opportunity.10 The dominant mineral of these concretions is hematite, an iron oxide that may have nucleated on organic centers and/or may have precipitated rapidly from concentrated solutions. These concretions have been concentrated as a lag deposit on the surface, where the more soluble evaporitic salts were removed by weathering processes. Sites with concretions should be considered as potential sites for biosignatures.
Sites Targeted for Enrichment of Biosignature Molecules
Sites of interest include carbon-bearing rocks and soils, layered sedimentary rocks, clays, weathered terrains, and iron oxide deposits.
-
Carbon-bearing rocks and soils. Deposits enriched in organic carbon are prime targets for detailed analysis, either with measurements in situ or on returned samples. Organic biomarker studies on Earth typically require carbon-rich samples, and there is no reason to believe that a strategy for Mars would be different. However, to date, no carbon-rich materials have been identified on Mars.
-
Layered sedimentary rocks. Ample evidence suggests that both surface water and groundwater existed on Mars, and that sediments accumulated from flowing surface waters and from lakes or oceans. Sedimentary rocks on Mars are found with various scales of stratification ranging from tens of meters (Figure 4.2) to very fine laminations visible in images from the rover Opportunity. Where deposits of sediments occurred in lakes or in a global ocean, biomolecule enrichment could have occured. Possible examples are deltas in the Holden crater.11
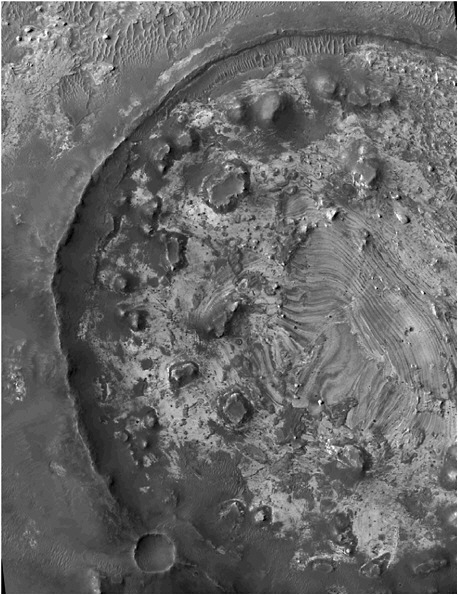
FIGURE 4.2 Layering in an unnamed crater in Meridiani Planum. The crater was once partly buried by layered deposits that have since been largely removed, leaving remnants of the former cover as mesas within the crater. The field is about 380 m across and is illuminated from the left. Image obtained by the High Resolution Imaging Science Experiment on the Mars Reconnaissance Orbiter spacecraft and provided courtesy of NASA/JPL/University of Arizona.
-
Clays. Clays are the finest fraction in a sedimentary deposit and generally reflect deposition in waters that have minimal current or wave turbulence. The surface charge and interlayering of clay minerals enhance adsorption of organic matter; hence clays may concentrate biomolecules from the solution in which they settled. However, clays are also transported by wind and have been mixed and deposited around the planet.
-
Weathered terrains. Bibring et al. have suggested that regions on the surface of Mars may show evidence for phyllosilicate minerals indicative of mineral weathering.12 If these older sections of the martian crust contain clays formed at low temperature, they may be likely sites to find organic molecules remaining from cells that were living when water was present. Minerals formed during the weathering of igneous rocks are indicative of the chemistry, pH, and redox conditions of the weathering solution.
-
Iron oxides deposits. Iron oxides are an abundant constituent of martian dust, and hematite concretions are closely associated with sulfate deposits at Meridiani. Concentrations of iron oxides, formed by geochemical processes, weathering, or Aeolian processes, could provide significant targets of astrobiological interest.
Sites Targeted for Other Biosignatures
Life harvests the available chemical energy in a rock-water-gas system or the light energy from the Sun at the same time that it extracts nutrients from the environment. A strategy for detection of extant or past life on Mars might therefore target sites where chemical energy is or was available, or sites showing evidence that nutrient extraction has occurred. Chemical systems of interest with respect to energy and nutrient needs for life include the following:
-
Carbon redox. A likely target for any in situ or sample-return analytical mission will be the detection and analysis of carbon in both the organic and inorganic species. Methane represents C in its most reduced form, and its presence within the martian atmosphere could be a signal of biologically mediated reactions that drive C to this redox state; methane may also be an oxidizable energy source for life on this planet. Point sources of methane should be considered as targets for future investigation.
-
Sulfur redox. The oxidation of sulfides and the reduction of sulfates are both biologically mediated reactions on Earth. A search for juxtaposed mineralogies containing the reduced and oxidized forms of S and Fe could occur at a variety of martian sites, including putative hydrothermal weathered deposits. Boundary effects may be particularly important. That is, interfaces between different minerals or fluids that are not in equilibrium may be appealing biological niches. An example of a potential target of interest is Noachian rock that may have weathered under less oxidizing conditions than immediately adjacent younger deposits.
-
Iron redox. The abundance of iron minerals on Mars, including igneous olivine, iron oxide dust, and sedimentary hematite, provide potential redox reaction sites that may support life and may leave mineralogical evidence of past life. A target for this exploration might include sites where especially high amounts of energy are available in the form of, for example, reduced iron minerals juxtaposed with likely oxidants. The rocks of the Columbia Hills, for example, have a wide range of ferrous/ferric ratios, with ferrous-rich primary minerals in intimate contact with ferric alteration products.13
Morphological Biosignatures
A morphological biosignature such as a stromatolite is only likely to be found at sites where water was present and where preservational mineralogy was favorable. The formation of stromatolites in shallow aqueous settings on Earth may suggest an investigation of paleo-shorelines on Mars. Lakes were likely common when the valley networks formed in the Noachian uplands,14 and large bodies of water must have been left at the ends of the large, post-Noachian outflow channels.15 However, the precise location of shorelines around these former bodies of water remains controversial. Other morphological biosignatures would be most likely to form in a sediment-depositing environment, such as a former lake or paleo-ocean.
Nonredox Gradients
The formation of gradients in elemental concentration due to biological exudates may be important at the meter scale in soil horizons or regolith and at the micron scale on mineral surfaces or in endolithic communities. The assimilation of trace elements at low concentrations by microorganisms or the sequestration of toxic elements into biologically mediated precipitates could also create distributions or gradients in trace elements that record the prior presence of biota in regolith or in sedimentary environments.
SITES AND SAMPLES FOR RECENT LIFE
All biochemical reactions that sustain life require water. Hence, water (and by extension ice) is the primary focal point in site selection for the search for present life. Sites where liquid water may have been present recently at the surface include numerous gullies,16 the ends of geologically recent outflow channels,17 and sites of recent glaciation.18 Water is, however, not the only target in the search for present life. Gas anomalies are also of great interest. For example, metabolic end products, including methane, carbon dioxide, and hydrogen sulfide, might indicate the presence of active local microbial communities.
Sites Containing Liquid Water
On Earth, life is active in aqueous systems over broad ranges of physicochemical conditions. Therefore, the presence of liquid water is a strong signal to follow in searching for present life.
Near the Surface
To date, there is no evidence for standing water on the surface of Mars. In fact, water is unstable under present surface conditions. However, it may persist in the shallow subsurface as part of an extensive groundwater system. Target environments where subsurface water—and hence present life—may be most likely include the following:
-
Sediments and sedimentary rocks where porosity and permeability are characteristic of aquifers;
-
Areas below surface features related to recent aqueous processes (e.g., gullies and young outflow channels); and
-
Areas with significant concentrations of minerals such as clay or evaporates that are diagnostic of low-temperature aqueous processes and that appear to be very recent.
Surface Water in the Recent Past
Recent outflow channels and recent gullies, if they formed by groundwater seepage, could have brought subsurface organisms to the surface. In this scenario, sediments and/or ice associated with gully and channel formation could harbor dormant life as spores or cells in vegetative states. On Earth, for example, spores have been shown to survive for very long periods of time.19 Sites of particular interest are fluvial deposits, evaporites, rocks, and minerals indicative of hydrothermal systems (e.g., hematite), and permafrost.20 Deposits at the ends of gullies, common in mid-southern latitudes, and deposits at the ends of young outflow channels, such as those in Cerberus and to the southeast of Olympus Mons, are thus possible sites to search for dormant, but extant, life.
Water Ice
Recent Earth-based studies have demonstrated that microorganisms can survive, grow, and metabolize in frozen sediments and ice cores, where pockets of liquid water are maintained.21 This could occur if salts are concentrated in liquid water or at the grain contacts where the freezing temperature is depressed. Relevant water-ice deposits include the following:
-
Ground ice. Ground ice appears to be ubiquitous a few centimeters below the surface at latitudes above ~50°N and S.22 While martian permafrost temperatures are much lower than terrestrial permafrost temperatures, these ice-rich environments have some similarities to permafrost environments found on Earth that contain many metabolizing microorganisms.
-
Glacial ice. Deposits left by cold-based glaciers have been identified on Mars, particularly on the northwest flanks of the large Tharsis volcanos.23 They probably formed in recent geological times during periods of high obliquity and might therefore still contain dormant lifeforms or their remnants.24
-
Polar ice caps. Thick (up to several km), layered sediments of dust and ice are exposed in the polar ice caps, which extend to ~80°N and S latitude (Figure 4.3). Owing to chaotic oscillations of Mars’s obliquity,25 the

FIGURE 4.3 Layered deposits near Mars’s north pole. The intricate patterns in the middle of the image are caused by gypsum-rich dunes exposed in a cliff. The smooth areas to the upper left and lower right are more typical of the deposits that surround the north pole. Image obtained by the High Resolution Imaging Science Experiment on the Mars Reconnaissance Orbiter spacecraft and provided courtesy of NASA/JPL/University of Arizona.
-
polar regions may experience periodic temperatures near the melting point of water ice, making the cap environments potentially habitable to extremely psychrophylic (cold-loving) microorganisms.
Localized Gas Fluxes
The metabolisms of many microorganisms create gaseous end products. These include CH4, CO2, H2S, N2, NH3, H2, and O2. CH4 is of particular interest with respect to Mars, owing to recent claims of its detection.26–28 On Earth, most of the atmospheric methane is biogenic, produced by aceticlastic or chemolithoautotrophic archaea; a recent study also ascribes the production of methane to plants.29 However, processes of abiogenic methane production also need to be considered. These include metamorphism/reduction of carbonates, serpentinization of olivine and subsequent abiotic reduction of CO2 with the generated H2, and recycling/thermal cracking of deeply buried complex organic matter (e.g., kerogen). Despite the ambiguity of methane as a biomarker,30 sites on Mars where methane is clearly detected are high-value targets in the search for present (presumably subsurface) life.
SUMMARY
There are a wide range of sites on Mars that merit astrobiological investigation. These include sites of possibly active water at the present, sites that have had recent water and could conceivably harbor dormant organisms, and sites that had ancient water. It is difficult to prioritize these from a scientific perspective, as they each represent viable and valid regions for study, and each will contribute greatly to astrobiological understanding of Mars. In addition, the technological requirements for exploring each are not at present well defined, such that it is difficult to know which will be most fruitful for the next one to two decades. However, a general statement is that the highest priority should go to sites where liquid water is or was present and where abundant energy is available that can drive metabolism.
REFERENCES
1. B.W. Fouke, J.D. Farmer, D.J. Des Marais, L. Pratt, N.C. Sturchio, P.C. Burns, and M.K. Discipulo, “Depositional Facies and Aqueous-Solid Geochemistry of Travertine-Depositing Hot Springs (Angel Terrace, Mammoth Hot Springs, Yellowstone National Park, U.S.A.),” Journal of Sedimentary Research 70:265-285, 2000.
2. L.E. Collins, W. Berelson, and D. Hammond, “Microbial Hitchhikers: The Flux of Bacteria on Particles Raining Through the Oxygen Minimum Zone, San Pedro Basin, CA,” Eos 87(36), Ocean Sciences Meeting Supplement, Abstract OS23H-06, 2006.
3. B. Gu, J. Schmitt, Z. Chen, L. Liang, and J.F. McCarthy, “Adsorption and Desorption of Natural Organic Matter on Iron Oxide: Mechanisms and Models,” Environmental Science and Technology 28:38-46, 1994.
4. M.H. Carr, The Surface of Mars, Cambridge University Press, New York, N.Y., 2006.
5. J. Farmer, “Hydrothermal Processes on Mars: An Assessment of Present Evidence,” in Evolution of Hydrothermal Ecosystems on Earth (and Mars ?), G. Bock and J. Goode (eds.), J. Wiley and Sons, N.Y., 1996.
6. J.A. Rathbun and S.W. Squyres, “Hydrothermal Systems Associated with Martian Impact Craters,” Icarus 157:362-372, 2002.
7. D.M. Burr, A.S. McEwen, and S.E. Sakimoto, “Recent Aqueous Floods from the Cerberus Fossae, Mars,” Geophysical Research Letters 29:1, doi:10.1029/2001Gl013345, 2002.
8. S.W. Squyres, R.E. Arvidson, J.F. Bell, J. Bruckner, N.A. Cabrol, W. Calvin, M.H. Carr, P.R. Christensen, B.C. Clark, L. Crumpler, D.J. Des Marais, C. d’Uston, T. Economou, J. Farmer, W. Farrand, W. Folkner, M. Golombek, S. Gorevan, J.A. Grant, R. Greeley, J. Grotzinger, L. Haskin, K.E. Herkenhoff, S. Hviid, J. Johnson, G. Klingelhöfer, A.H. Knoll, G. Landis, M. Lemmon, R. Li, M.B. Madsen, M.C. Malin, S.M. McLennan, H.Y. McSween, D.W. Ming, J. Moersch, R.V. Morris, T. Parker, J.W. Rice, Jr., L. Richter, R. Rieder, M. Sims, M. Smith, P. Smith, L.A. Soderblom, R. Sullivan, H. Wänke, T. Wdowiak, M. Wolff, and A. Yen, “The Opportunity Rover’s Athena Science Investigation at Meridiani Planum, Mars,” Science 306:1698-1714, 2004.
9. J. Bibring, Y. Langevin, J.F. Mustard, F. Poulet, R. Arvidson, A. Gendrin, B. Gondet, N. Mangold, P. Pinet, F. Forget, and the OMEGA team M. Berthé, J. Bibring, A. Gendrin, C. Gomez, B. Gondet, D. Jouglet, F. Poulet, A. Soufflot, M. Vincendon, M. Combes, P. Drossart, T. Encrenaz, T. Fouchet, R. Merchiorri, G. Belluci, F. Altieri, V. Formisano, F. Capaccioni, P. Cer-
roni, A. Coradini, S. Fonti, O. Korablev, V. Kottsov, N. Ignatiev, V. Moroz, D. Titov, L. Zasova, D. Loiseau, N. Mangold, P. Pinet, S. Douté, B. Schmitt, C. Sotin, E. Hauber, H. Hoffmann, R. Jaumann, U. Keller, R. Arvidson, J.F. Mustard, T. Duxbury, F. Forget, and G. Neukum, “Global Mineralogical and Aqueous History Derived from OMEGA/Mars Express data,” Science 312:400-404, 2005.
10. S.W. Squyres, R.E. Arvidson, J.F. Bell, J. Bruckner, N.A. Cabrol, W. Calvin, M.H. Carr, P.R. Christensen, B.C. Clark, L. Crumpler, D.J. Des Marais, C. d’Uston, T. Economou, J. Farmer, W. Farrand, W. Folkner, M. Golombek, S. Gorevan, J.A. Grant, R. Greeley, J. Grotzinger, L. Haskin, K.E. Herkenhoff, S. Hviid, J. Johnson, G. Klingelhöfer, A.H. Knoll, G. Landis, M. Lemmon, R. Li, M.B. Madsen, M.C. Malin, S.M. McLennan, H.Y. McSween, D.W. Ming, J. Moersch, R.V. Morris, T. Parker, J.W. Rice, Jr., L. Richter, R. Rieder, M. Sims, M. Smith, P. Smith, L.A. Soderblom, R. Sullivan, H. Wänke, T. Wdowiak, M. Wolff, and A. Yen, “The Opportunity Rover’s Athena Science Investigation at Meridiani Planum, Mars,” Science 306:1698-1714, 2004.
11. M.C. Malin and K. S. Edgett, “Evidence for Persistent Flow and Aqueous Sedimentation on Early Mars,” Science 302:1931-1934, 2003.
12. J. Bibring, Y. Langevin, J.F. Mustard, F. Poulet, R. Arvidson, A. Gendrin, B. Gondet, N. Mangold, P. Pinet, F. Forget, and the OMEGA team M. Berthé, J. Bibring, A. Gendrin, C. Gomez, B. Gondet, D. Jouglet, F. Poulet, A. Soufflot, M. Vincendon, M. Combes, P. Drossart, T. Encrenaz, T. Fouchet, R. Merchiorri, G. Belluci, F. Altieri, V. Formisano, F. Capaccioni, P. Cerroni, A. Coradini, S. Fonti, O. Korablev, V. Kottsov, N. Ignatiev, V. Moroz, D. Titov, L. Zasova, D. Loiseau, N. Mangold, P. Pinet, S. Douté, B. Schmitt, C. Sotin, E. Hauber, H. Hoffmann, R. Jaumann, U. Keller, R. Arvidson, J.F. Mustard, T. Duxbury, F. Forget, and G. Neukum, “Global Mineralogical and Aqueous History Derived from OMEGA/Mars Express Data,” Science 312:400-404, 2006.
13. S.W. Squyres, R.E. Arvidon, D.W. Blaney, B.C. Clark, L. Crumpler, W.H. Farrand, S. Gorevan, K.E. Herkenhoff, J. Hurowitz, A. Kusack, H.Y. McSween, D.W. Ming, R.V. Morris, S.W. Ruff, A. Wang, and A. Yen, “The Rocks of the Columbia Hills,” Journal of Geophysical Research 111:E02S11, doi:10.1029/2005JR002562, 2006.
14. N.A. Cabrol and E.A. Grin, “Distribution, Classification and Ages of Martian Impact Crater Lakes,” Icarus 142:160-172, 1999.
15. S.M. Clifford and T.J. Parker, “The Evolution of the Martian Hydrosphere: Implications for the Fate of a Primordial Ocean and the Current State of the Northern Plains,” Icarus 154:40-79, 2001.
16. M.C. Malin and K.S. Edgett, “Evidence for Recent Groundwater Seepage and Surface Runoff on Mars,” Science 288:2330-2335, 2000.
17. J.W. Head, L. Wilson and K.L. Mitchel, “Generation of Recent Water Floods at Cerberus Fossae, Mars by Dike Emplacement, Cryosphere Cracking and Confined Aquifer Groundwater Release,” Geophysical Research Letters 30(11):1577, doi:10.1029/2003GL017135, 2003.
18. J.W. Head, D.R. Marchant, M.C. Agnew, C.I. Fassett, and M.A. Kreslavsky, “Extensive Valley Glacier Deposits in the Northern Mid-Latitudes of Mars: Evidence for Late Amazonian Obliquity-Driven Climate Change,” Earth and Planetary Science Letters 241:663-671, 2005.
19. See, for example, W.L. Nicholson, N. Munakata, G. Horneck, H.J. Melosh, and P. Setlow, “Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments,” Microbiology and Molecular Biology Reviews 64(3):548-572, 2000; and references therein.
20. M.R. Walter and D.J. DesMarais, “Preservation of Biological Information in Thermal Spring Deposits: Developing a Strategy for the Search for Fossil Life on Mars,” Icarus 101:129-143, 1993.
21. See, for example, J.C. Priscu and C. Christner, “Earth’s Icy Biosphere,” pp. 130-145 in Microbial Diversity and Prospecting, A.T. Bull, ed., ASM Press, Washington, D.C., 2004.
22. W.C. Feldman, T.H. Prettyman, S. Maurice, J.L. Plaut, D.L. Bish, D.T. Vaniman, M.T. Mellon, A.E. Metzger, S.W. Squyres, S. Karunatillake, W.V. Boynton, R.C. Elphic, H.O. Funsten, D.J. Lawrence, and R.L. Tokar, “The Global Distribution of Near Surface Hydrogen on Mars,” Journal of Geophysical Research 109(E9):E09006, doi:10.1029/2003JE02160, 2004.
23. J.W. Head and D.R. Marchant, “Cold Based Mountain Glaciers on Mars: Western Arsia Mons,” Geology 31:641-644, 2003.
24. B.M. Jakosky, R.M. Haberle, and R.E. Arvidson, “The Changing Picture of Volatiles and Climate on Mars,” Science 310:1439-1440, 2005.
25. J. Laskar, B. Levrard, and J.F. Mustard, “Orbital Forcing of the Martian Polar Layered Deposits,” Nature 419:375-377, 2002.
26. V. Formisano, S. Atreva, T. Encrenaz, N. Ignatiev, and M. Giuranna, “Detection of Methane in the Atmosphere of Mars,” Science 306:1758-1761, 2004.
27. V.A. Krasnopolsky, J.P. Maillard, and T.C. Owen, “Detection of Methane in the Martian Atmosphere: Evidence for Life?,” Icarus 172:537-547, 2004.
28. M.J. Mumma R.E. Novak, M.A. DiSanti, B.P. Bonev, and N. Dello Russo, “Detection and Mapping of Methane and Water on Mars,” Bulletin of the American Astronomical Society 36:1127, 2004.
29. F. Keppler, J.T.G. Hamilton, M. Brass, and T. Roeckmann, “Methane Emissions from Terrestrial Planets under Aerobic Conditions,” Nature 439:187-191, 2006.
30. M. Allen, B. Sherwood Lollar, B. Runnegar, D.Z. Oehler, J.R. Lyons, C.E. Manning, and M.E. Summers, “Is Mars Alive?,” Eos 87:433-439, 2006.













