
2
How Can We Best Preserve Biological Diversity and Protect Endangered Ecosystems?
The extinction of plant and animal species is not only decreasing Earth’s biodiversity and depriving humans of potential resources for food, medicine, and simple enjoyment of nature; it is endangering the functioning of ecosystems and potentially precipitating a cascading effect of increased ecosystem loss and further erosion of biodiversity (Kinzig et al., 2001; Loreau et al., 2001, 2002). Ecosystems are critical to human welfare. They sequester carbon, produce oxygen, generate chemical energy from sunlight, and are integral to soil formation, nutrient cycling, food production, wood and fiber production, the regulation of water flow, and the transference of water to the atmosphere. Ecosystems have always been in flux, but as the Millennium Ecosystem Assessment (MEA) makes clear:
Over the past 50 years, humans have changed ecosystems more rapidly and extensively than in any comparable period of time in human history, largely to meet rapidly growing demands for food, fresh water, timber, fiber, and fuel. This has resulted in a substantial and largely irreversible loss in the diversity of life on Earth. (MEA, 2005:1)
Human activity is the likely cause of the extinction or disappearance of close to 850 species over the past 500 years (Baillie et al., 2004). The rates of plant and animal extinction today are conservatively estimated to be 100 times to more than 1,000 times greater than past average extinction rates calculated from fossil lineages (MEA, 2005). The causes of past and present extinctions include a number of human-driven factors such as habitat and ecosystem destruction and fragmentation, overexploitation of species through hunting and fishing, competition and predation by invasive species, and pollution (MacDonald, 2002; MEA, 2005; Loreau et al., 2006; Worm et al., 2006). At the global scale, the biosphere can be grouped into geographical regions called biomes in which climatic conditions and vegetation structure are internally similar, but differentiable from other biomes. The majority of some biomes, such as the temperate forests or Mediterranean woodlands and shrublands, have already been converted to cultural landscapes (Figure 2.1).
Today, tropical forest systems and wetlands are being hit particularly hard by habitat destruction and fragmentation (e.g., DeFries et al., 2005). On top of this, rates of extinction could increase catastrophically as species find themselves unable to adjust geographical ranges quickly enough in response to climate change. In an analysis of the impacts of a moderate climate warming scenario on more than 1,000 plant and animal species in Mexico, Australia, South America, and Africa, Thomas et al. (2004) concluded that between 15 percent and 37 percent of these species would be committed to extinction by 2050. Malcolm et al. (2006) estimated that up to 43 percent of species in some biodiversity hotspots may face extinction owing to changing climate and vegetation distributions caused by global warming.1
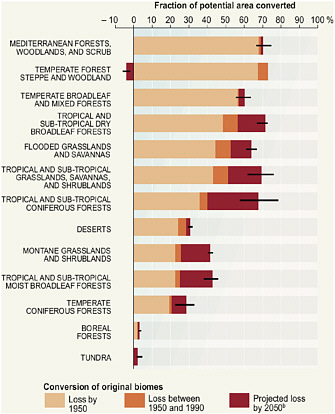
FIGURE 2.1 Past and projected conversions of major biomes to cultural landscapes and loss of original ecosystems. SOURCE: MEA (2005).
ROLE OF THE GEOGRAPHICAL SCIENCES
The geographical distribution of biodiversity, threats of biodiversity and ecosystem loss, and regions where conservation efforts should focus are not evenly distributed, but display distinct spatial patterning at all scales, from local to global (Brooks et al., 2006; Kremen et al., 2008). Species richness decreases from the equator poleward (Figure 2.2). Within this general pattern, certain geographical areas have notably high numbers of species, many found nowhere else in the world. These areas of high endemic species richness are referred to as biodiversity hotspots and are often regions prone to significant ongoing ecosystem alteration and loss (Figure 2.2). The role that location and geographical context play on biodiversity and ecosystem loss makes the geographical sciences integral to understanding this issue.
Through field studies, remote sensing, and ecological modeling, the geographical sciences document and explain biodiversity distribution and contribute to its preservation through strategies aimed at optimizing conservation (Church et al., 2003). Scientists are still seeking to determine how many species of plants and animals the planet supports. The lack of information is particularly notable in marine and freshwater systems, which have received less attention than their terrestrial counterparts (Richardson and Poloczanska, 2008).
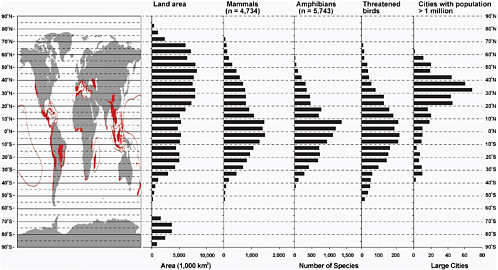
FIGURE 2.2 Map of Earth with the locations of major biodiversity hotspots in red (Myers et al., 2000), the latitudinal biodiversity gradients for mammals, amphibians, and threatened bird species (MEA, 2005), and cities with populations greater than 1 million (UN, 2009). SOURCE: Adapted from Myers et al. (2000); MEA (2005); and UN (2009). Used with permission of Island Press, Washington, D.C.
Geographical scientists have demonstrated that both biophysical and sociocultural dimensions are central to the causes and consequences of land-cover and landuse change, and they have advanced understanding of how human circumstances (e.g., social marginalization) and associated processes (e.g., policy changes) affect biodiversity or ecosystem loss. This has resulted in the development of the emerging interdisciplinary field of land change science (LCS), the goal of which is to develop integrated explanations of land change (Turner et al., 2007). The Global Land Project of the International Geosphere-Biosphere Programme represents an international effort to understand the interacting drivers, patterns, and impacts of such changes.
An exposition of the aims of LCS and insights into the role of the geographical sciences is provided by a recent special feature on the topic in Proceedings of the National Academy of Sciences (Turner et al., 2007). In one study particularly representative of the spatial and integrative nature of the geographical sciences, Irwin and Bockstael (2007) use geographical pattern metrics of land-use change in Maryland from 1973 to 2000 to document the spread of urban development and the resulting fragmentation of habitat (Figure 2.3). They drew three conclusions. First, contrary to earlier work on national patterns, urban growth is an ongoing phenomenon and is being underestimated in other studies because of insufficient attention to increasing low-density exurban development. Second, the increasing growth is often peripheral and low density and is leading to increased habitat fragmentation and loss. Third, the environmentally sensitive Chesapeake Bay region is actually experiencing increased development because of its commercial and recreational amenity value. The movement of people to economically and environmentally attractive urban areas causes habitat loss and fragmentation, but much remains to be done to assess the precise impacts of these demographic shifts on ecosystems and biodiversity. It follows that an important question for the geographical sciences, environmental sciences, sociology, economics, and environmental ethics is how to manage the growing
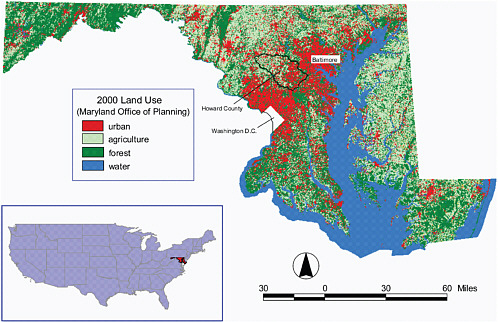
FIGURE 2.3 Land use in Maryland in 2000. Note the fragmentation of forested and agricultural land on the outskirts of the Baltimore-Washington metropolitan area, which Irwin and Bockstael show from longitudinal data has been ongoing since the 1970s. Also, the increasing concentration of urban development along the shores of the Chesapeake Bay reflects an influx of development because of the commercial and recreational amenities of the ecologically sensitive waterway. Urban growth is in this case affecting both terrestrial and estuarine ecosystems. SOURCE: Irwin and Bockstael (2007).
pressures for urban development in a manner that balances demands and biodiversity/ecosystem protection (see also Chapter 4). Understanding the geographical patterns and rates of such changes in land use through use of the geographical sciences can help elucidate the trade-offs involved.
RESEARCH SUBQUESTIONS
How is biodiversity distributed and controlled?
The number of species that comprise the biodiversity of the planet is unknown, with estimates ranging between 5 million and 30 million. Of this number, only about 2 million have been scientifically described (Groombridge and Jenkins, 2002; MEA, 2005). The spatial distributions of many of the known species remain poorly articulated. The geographical sciences can help address this lacuna through the integration of biological censuses with measurements of physical environmental variables. Field-based measurements can be obtained through traditional ecological census methods and physical geography approaches, but it is impossible to cover the expanse of the biosphere, particularly in remote and difficult-to-access tropical regions. Hence, estimating species distributions and biodiversity requires integrating the results of field research with remote sensing data, and using geographic information systems (GIS) to overlay and extrapolate biological and environmental variables (e.g., Cohen andand Goward, 2004; Gillespie et al., 2004, 2008; Goetz et al., 2007; Figure 2.4).
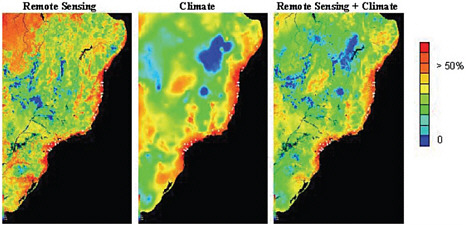
FIGURE 2.4 Results of a maximum entropy model provide an example of how remote sensing data on landscape/vegetation and climatic data can be combined to improve understanding of fine-scale features that influence the spatial distribution of species, in this case Carpornis melanocephala (black-headed berryeater) in Brazil. Higher values indicate optimal conditions for the species, whereas lower values indicate poor conditions. Geographically explicit and integrated studies of field observations, remotely sensed land-cover data, and other environmental data to estimate species distributions are an important application of the geographical sciences. SOURCE: Gillespie et al. (2008).
Geographical science approaches also contribute to the development of models that estimate potential biodiversity and aid in the development of conservation strategies on the basis of measured environmental parameters, human land-use patterns, species physiology, and species behavior. Coupled field and remote sensing modeling is being developed to estimate biodiversity over wide geographical areas. These approaches show much promise for documenting current conditions and establishing conservation strategies (Figure 2.5).
Understanding the geographical and environmental distribution of genetic diversity within species is necessary to anticipating how changes in population genetic structure could influence ecosystem functioning, extinction risk, and biodiversity (Reusch et al., 2005). As environmental changes affect a species, certain genotypes may be advantaged, and the genetic diversity of the species may change over time. Losses in genetic diversity can also occur through stochastic processes when species populations are isolated or become very small. The loss of genetic diversity may hamper a species’ ability to exist in certain environmental settings because the genetically controlled traits required for survival in those environments have been lost. Understanding and preserving the genetic diversity of species, which often demonstrates spatial patterning, are important components of the conservation applications of the geographical sciences in the future.
What are the spatiotemporal patterns and drivers of ecosystem and habitat loss?
The rates of global ecosystem loss and habitat fragmentation and the relationship of those processes to biodiversity loss are important topics of continued research for the geographical sciences. It can be difficult even to compare fragmentation measures from one country to another because of differences in data availability and the metrics used (Kupfer, 2006). Some of the most important insights into the magnitude, rate, and spatial distribution of habitat loss have come from the geographical sciences. A recent study of Amazonian deforestation in Brazil by Morton et al. (2006) provides an informative example (Figure 2.6). Between 2001 and 2004 more than 3.6 million hectares of Amazonian forest were cleared for intensive agriculture. Morton and colleagues used a time series of MODIS (Moderate Resolution Imaging Spectroradiometer)
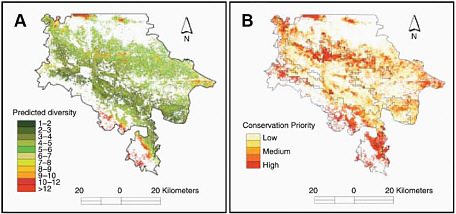
FIGURE 2.5 (A) Predicted tree species diversity and (B) suggested areas of conservation priority for the state of Chiapas, Mexico, based on combination and modeling of vegetation data, environmental data, human population and land-use data, and remote sensing imagery. Even in a highly fragmented and mountainous landscape, this research demonstrates the usefulness of integrated and spatially explicit approaches to documenting biodiversity and developing conservation strategies. SOURCE: Cayuela et al. (2006).
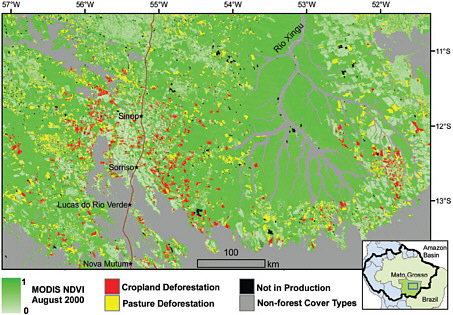
FIGURE 2.6 Clearance of Amazonian forest for pasture and cropland development in the state of Mato Grosso, Brazil, between 2001 and 2004 as detected from satellite imagery. Areas that were cleared for grazing were typically twice the size of patches cleared for crops. SOURCE: Morton et al. (2006).
satellite imagery to show significant deforestation caused by the clearing of more than 540,000 hectares for the development of both cropland and grazing land in the state of Mato Grosso, Brazil. They found that the destruction of forest land was not random. The satellite imagery showed that it occurred along the leading edges of previously cleared land. The study concluded that agricultural intensification and the development of permanent croplands and rangelands was accelerating rates of deforestation, and these rates might be expected to increase if crop prices rise in the future.
Habitat loss can further isolate current protected areas and make the conservation of species and ecosystems within such protected areas difficult. Morton et al. (2006) used 20 years of satellite data to assess changes in habitat around 198 of the world’s largest tropical forest conservation areas. They found that about half of the protected areas had experienced significant natural habitat loss (i.e., 5 percent or greater) over the past 20 years. In some cases the loss of natural habitat in the surrounding buffer areas was as high as 80 percent. The increasing isolation of these protected areas makes them less effective as conservation areas for maintaining species diversity. There is great potential to expand the scope of such studies to many different world regions and timescales.
Although the biophysical sciences and remote sensing can do much to aid in biodiversity and ecosystem conservation, understanding the underlying socioeconomic patterns and forces that drive particular land-use patterns and ecosystem losses requires close interaction between biophysical and social scientists. Gap analysis, which uses GIS to overlay threatened species distributions, their habitat types, human land use, and the status of legal protection for lands can be extremely useful for determining geographically based conservation strategies and land acquisition priorities to preserve biodiversity in an increasingly fragmented environment (e.g., Davis et al., 2004; Gleason et al., 2006).
Two challenges for integrating sociocultural measures of land-use and land-cover change deserve particular attention in the coming decade. First, remote sensing data do not provide direct measures of sociocultural variables such as income, political preference, and education, only indirect, or proxy, measures. This limitation of remotely sensed data means that scientists attempting to explain land-surface or coastal ecosystem processes need to assert a relationship between the remotely sensed data and underlying social processes only when an externally validated link between the remotely sensed data phenomenon and the social process has been established. Thus further research on where and when to establish such links is needed.
Second, and linked to the first challenge, data from most remote sensing datasets, such as Landsat satellite imagery, are typically too coarse to provide effective measures of the small-scale or diffuse land-use and land-cover processes under examination. Fortunately, in recent years the availability of data that have finer spatial resolution has increased, and the cost has decreased. Geographical scientists are thus in a good position to test and improve, where possible, the explanatory power of recent research on land-use and land-cover change using increasingly finer resolution data.
How will future climate change influence species’ distributions and biodiversity?
Information on climatic controls on geographical distribution and potential rates of species migration are critical to anticipating the impacts of future climate change on species. The climate and ecosystems of some present biodiversity hotspots may be so altered in the future as to make them unsuitable to support their current endemic species. Data are still too sparse, however, to predict confidently the impacts of climate change on future potential distributions and migration rates of most threatened species (Araújo et al., 2005; Pimm, 2007). Developing effective conservation policies depends on being able to predict where species will be able to exist under a changed climate and how fast they might be able to migrate to new regions as the climate changes. GIS-based collations and analyses of species distributional data and current and projected environmental conditions can be particularly important in linking field observations, climate forecasts, and ecological models of species distributions and potential migration rates in a spatially explicit manner to help with this task (e.g., Guisan et al., 2006). In terms of potential migration rates, the geographical sciences can contribute through both empirical studies based on observations of past and present organism dispersal patterns (e.g., Clark et al., 1998; Greene et al., 2004) and through mathematical simulations of dispersal (e.g., Malanson, 2003).
One example of a geographical approach to this problem—and a sobering view of the possible magnitude of climate change impacts—comes from a study by Malcolm et al. (2006). They examined 14 different computer models that produce estimates of the climatic changes likely to be caused by a doubling of CO2. They averaged the climatic estimates produced by those models and used the averaged estimates of climate as input for two different models that predict biome distributions on the basis of climatic conditions. They then used these results to predict the possible future shifts of major biodiversity hotspots based on the predicted shifts in climate and vegetation. They then considered potential migration rates of the biota in the current hotspots relative to where suitable habitat for those species would be found in the future. They concluded that the extinction rates resulting from geographical shifts in current hotspots would be regionally variable, ranging from less than 1 percent to 43 percent, with an average loss of 11.6 percent of the species. They found that the most vulnerable hotspots were the South African Cape Floristic Region, Caribbean, Indo-Burma, Mediterranean Basin, Southwest Australia, and Tropical Andes (see Figure 2.1). In these areas, plant extinctions could exceed 2,000 species. A particularly troubling conclusion from this study was that if these climate-driven changes occurred within the projected time frame of 100 years, the rate of tropical extinctions caused by global warming could well exceed the already high rates of extinction from deforestation and land-surface change.
Observational records of plant and animal populations and ecosystems are usually too short in duration to detect and interpret long-term trajectories or the response of biological systems to long-term environmental change. Extending the temporal depth of environmental analysis along with the spatial coverage is a central concern of the geographical sciences. Such paleoecological and paleoclimatic studies can provide empirical data on the response of species in terms of geographical distribution, population size, migration rates, and extinction by looking at the fossil records of species and community reactions to past episodes of climatic warming over the Quaternary Period and earlier (Botkin et al., 2007; Willis et al., 2007; MacDonald et al., 2008a). Such research can point out which species or communities may be relatively resilient and which may be more sensitive to future land-surface and climate change based upon their response to past episodes of environmental change.
Paleostudies entail reconstructing long histories of ecosystems to indicate their response to past environmental changes and the degree to which they have been altered by human activity in order to provide benchmarks or targets for ecosystem restoration. An example of such a study comes from the work of Millspaugh et al. (2000) based on analysis of a sediment core from Cygnet Lake in Yellowstone National Park (Figure 2.7). Through the analysis of fossil charcoal and pollen Millspaugh et al. (2000) showed that there was a positive relationship between fire frequency and increased temperatures and aridity. The hot and dry conditions were natural variations in climate caused by increased summer insolation related to changes in the orbital geometry of Earth. These results suggest that increased fire frequency is a reasonable expectation with future climate warming, but that there is a high degree of adaptation and resilience to varying fire frequencies in the vegetation of Yellowstone. Both observations are important for the management of the Park’s ecosystems in the face of climate change. More basic biogeographical research on species distributions, environmental relations, and dispersal capabilities, coupled with model development, can improve assessments of the impacts of climate change on biodiversity and endangered ecosystems.
How can we conserve biodiversity and ecosystems while sustaining human livelihoods?
Devising conservation strategies requires grappling with the complexity in indigenous as well as increasingly globalized land-use practices and differing cultural approaches to nature valuation, conservation practices, and compliance (e.g., Messina et al., 2006; Robbins et al., 2006; Walsh et al., 2008). The role of socioeconomic factors in habitat ecosystem modification and biodiversity is often complex and at times counterintuitive. Geographical scientists working at the interface of conservation ecology and the social sciences have shown, for example, that some human land-use patterns, such as certain forms of swidden
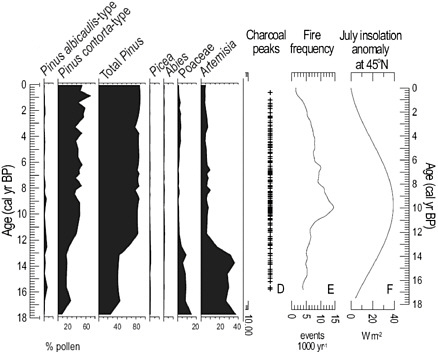
FIGURE 2.7 The history of regional vegetation and fire frequency in Yellowstone National Park over the past 17,000 years based on fossil pollen and charcoal. Since 11,000 years ago the vegetation has remained relatively stable despite significant long-term changes in fire frequency driven by climate change related to changes in July insolation related to changes in Earth’s orbital geometry. SOURCE: Millspaugh et al. (2000).
agriculture,2 may support local human populations and also serve to mimic natural disturbance regimes and thus help preserve biodiversity (Robbins et al., 2006; see also Chapter 5).
The geographical sciences have contributed to current debates about the premises behind widely used conservation strategies such as the best approach for ex situ biodiversity conservation.3 Although the concept of the modern park, or “Yellowstone model,” dates to the late 19th century, the related notion of a hunting preserve took form across the tropics during the colonial era (Cronon, 1996). These preserves endured over time, eventually becoming national parks that gained considerable international support following the rise of an environmental movement in high-income countries during the 1970s and 1980s. By the late 1980s, however, it was becoming clear that the park model was failing in many tropical countries. In most cases there was
a strong conviction that local people were destructive agents who should not live within the borders of parks (Cronon, 1996). This notion led to the removal of local peoples and the near or complete destruction of their livelihoods, in the process straining relations between park authorities and local residents (e.g., Peluso, 1993; Guha, 1997).
The failure of the park model led to a dynamic round of experimentation with community-based natural resources management (CBNRM) starting in the late 1980s. Here the core idea was that the government needed to give local people a stake in the success or failure of a park. The theory was that if local people profited from ecotourism, they would actively support efforts to conserve wildlife within and around the borders of a park. Associated with this flurry of experimentation was scholarship championing this approach (e.g., Metcalf, 1994) as quite positive whereas others, most notably Neumann (1997, 2002), saw CBNRM—and related approaches such as buffer zones—as just another disguised extension of a park model that ultimately constrained the livelihoods of local people. Still others saw the way in which CBNRM was implemented by governments as the major problem (e.g., Logan and Moseley, 2002).
By the late 1990s, there was a group of conservation biologists—most notably Terborgh (1999, 2000)—arguing that CBNRM had completely failed to conserve biodiversity and that there should be a return to a stricter and more robust form of the park model, sometimes referred to as fortress conservation. Subsequent scholarship critiqued the reemerging fortress conservation model, questioning whether it really served to conserve biodiversity (Robbins et al., 2006) and pointing out that it was still devastating for local livelihoods (Wilshusen et al., 2002). These studies highlight a growing body of work in the geographical sciences that underscores the importance of incorporating local perspectives and ideas in the development of biodiversity and ecosystem conservation strategies (Stevens, 2002; Kates et al., 2005).
Confronting these issues from an integrated natural and social sciences perspective is a major challenge that calls upon the integrative perspective and analytical tools of the geographical sciences (e.g., Turner et al., 1995; Liverman et al., 1998; Lambin et al., 1999; Fox et al., 2002; Walsh and Crews-Meyer, 2002). These integrated approaches are in the early stages of being formally incorporated into LCS (Turner et al., 2005, 2007) and should be examined for inclusion in the specific context of biodiversity and ecosystem conservation.
SUMMARY
Lessons learned by geographical scientists in the past two decades from attempts to model the process of land-use and land-cover change, and to project future distributions of land use and land cover, suggest that socially sensitive and integrated research approaches within the geographical sciences could greatly assist in the development and implementation of viable conservation strategies (e.g., Pontius et al., 2007). The ability of the geographical sciences to combine field studies, remote sensing data, climate data, and land-change models to understand ecosystem changes and biodiversity distribution will be critical to developing land-use policies and conservation strategies in the coming decade.










