
2
Dynamic Habitability
Earth remains our only reference point for an inhabited world. The field of astrobiology, however, seeks to differentiate between the specific requirements for life on Earth (terran life; Box 2.1) and conditions on other planets that might enable and promote the emergence and persistence of life. Therefore, astrobiology distills what we know of terran life and its origins on early Earth into the broad planetary conditions necessary for the emergence and proliferation of life and the rise and evolution of its biosignatures. Ideally, those conditions identified are general enough to apply across various types of celestial bodies (rocky planets, icy moons, small bodies, or exoplanets) but also specific enough to provide useful search and discovery tools.
This chapter focuses on the generalized planetary conditions and characteristics that are thought to support the emergence of life and its persistence through time. In the spirit of systems science, these conditions arise from the fundamental and broadly applicable principles of thermodynamics. In addition to advancing a thermodynamic perspective on life’s origins, this chapter also explores the nature and extent of extant life and how these have influenced the recent focus on the concept of dynamic habitability (Box 2.2). Finally, this chapter demonstrates how biological ecosystems coevolve with key aspects of planetary dynamics and evolution and suggests that this coevolution might either encourage or inhibit life’s emergence.
A THERMODYNAMIC PERSPECTIVE ON LIFE’S ORIGIN
The 2015 Astrobiology Strategy (NASA 2015) addresses the origin of life in a manner that dates back a century to what has become known as the Oparin-Haldane hypothesis (e.g., Chang et al. 1983). This hypothesis focuses primarily on how life emerged and specifically postulates that life began as a set of chemical reactions starting from relatively simple molecules produced, for example, in the atmosphere by lightning or photochemistry, in the crust by water-rock reactions, or by aqueous alteration on the parent bodies of carbonaceous meteorites. The hypothesis posits that these prebiotic molecules subsequently reacted at water-atmosphere-mineral interfaces to produce more complex molecules, polymers (covalently bonded chains of small molecular units; for example, nucleotides polymerize to form nucleic acids, and amino acids polymerize to form proteins), and/or metabolic cycles—eventually leading to self-organized, autocatalytic protocells. (See Box 4.1 for a discussion of the role viruses may have played in the transition from abiotic to biotic processes.) The broad strokes of this paradigm are still recognizable in much of modern prebiotic chemistry, which is laid out in detail in Chapters 1 to 3 of the 2015 Astrobiology Strategy.
More recently, the conversation has shifted to investigate whether there are naturally occurring generalizable forces on a planet that, under the right circumstances, lead to life’s emergence. Smith and Morowitz (2016) suggest that such planetary conditions might better elucidate why life arises. They hypothesize that Earth’s biosphere, specifically its metabolisms, arose as a relaxation path for planetary-scale thermodynamic disequilibria that result from slow relaxation timescales in late-stage planet formation. Thus, the thermodynamic disequilibrium exhibited by the young Earth across its atmosphere, hydrosphere, and lithosphere led to the emergence of life in the form of the biosphere, or as Smith and Morowitz term it, the “fourth geosphere.” Within this fourth geosphere, they argue, the ecosystem, rather than the individual organism, is the organizational level at which hierarchical controls occur. These system-level controls enable both the emergence of life and the apparent continuous existence of a terran biosphere—albeit a dynamically evolving one—over more than 4 billion years.
Finding: Systems-level emergence of life research incorporating thermodynamic principles is important for understanding life as a planetary phenomenon.
A strength of the Smith and Morowitz hypothesis is that it suggests a universality of laws governing the emergence and evolution of life that transcends the manifestations of individual genomes and species. This, of course, does not guarantee that it is correct, but it represents a testable hypothesis.
Thermodynamic disequilibrium is also required to sustain life once it gets started. All known life takes advantage of thermodynamic disequilibrium that is globally powered either by incoming solar radiation or by Earth’s internal radiogenic or accretionary heat. Thus, thermodynamic disequilibrium is a necessary input for life. It is, however, also a potential byproduct of life and one that evolves with changing planetary and biosphere conditions (Krissansen-Totton et al. 2018). For example, the accumulation of O2 in Earth’s atmosphere is a byproduct of oxygenic photosynthesis (Meadows 2017). In the modern atmosphere, the coexistence of both O2 and CH4 produce a thermodynamic disequilibrium that is often regarded as a planetary biosignature (Simoncini et al. 2013). Although
NASA funded the initial research by James Lovelock in the late-1960s and early-1970s on disequilibrium as a life-detection strategy (e.g., Hitchcock and Lovelock 1967), it is now widely accepted that atmospheric disequilibrium is not necessarily a byproduct of life. Kasting (2014) and others have pointed out that CO and H2 are typically out of thermodynamic equilibrium in models of Earth’s early atmosphere as a consequence of the production of CO from photolysis of CO2 combined with escape of H2 to space. The fact that thermodynamic disequilibria at varying temporal and spatial scales can give rise to life, sustain life, and also be a byproduct of life highlights the challenge of differentiating conditions that may give rise to and sustain planetary habitability and also produce biosignatures detectable on a planetary scale.
THE DYNAMIC EARLY EARTH AND LIFE’S EMERGENCE
The search for habitable worlds inevitably leads to questions of habitability on Earth—the only planet on which life certainly arose and evolved and hence an essential pillar of astrobiology research. While the extent of life on Earth knows few bounds and inhabits almost all known extreme environments, it is not clear that life’s origins on this planet were as ubiquitous. A century following the Oparin-Haldane hypothesis (e.g., Chang et al. 1983), the potential chemical and environmental pathways that gave rise to life have been greatly expanded and remain hotly debated. In many cases, these evolved hypotheses bear little resemblance to the organic chemistry originally postulated, and despite decades of research, plausible prebiotic pathways within realistic early Earth environments remain elusive. One of the greatest challenges still facing the origins of life community is not only
a more accurate and complete understanding of dynamic and spatially variable early Earth conditions, but also the means to faithfully represent these conditions in prebiotic chemistry experiments.
The earliest signs of life on Earth indicate that the prebiotic chemistry that led to life’s origins most likely commenced during the Hadean Eon (prior to ~4 Ga), which is thought to be the most dynamic period in the planet’s history, but is also the period for which observations, proxies, and models are the most sparse. The earliest possible timing for sustainable prebiotic chemistry followed the Moon-forming event and replacement of the magma ocean by solid crust at ~4.5 Ga (Abe 1993; Elkins-Tanton 2008; Sleep et al. 2014; Harrison et al. 2008). In the ensuing half billion years, the onset of plate tectonics, formation of continents, and condensation of the atmosphere and oceans gave rise to the dynamic, diverse, and interconnected environments from which the chemistry that led to life’s origins arose.
The temporal relationships among these transitions are crucial to accurately describing and replicating early Earth environments in the laboratory and in modeling, and yet, the timing of only a few of these events is well constrained. For example, the presence of a liquid water ocean and crustal rocks with a bulk granitoid composition by ~4.4 Ga is borne out by the chemical and isotopic signatures of the Jack Hills Hadean zircons (Wilde et al. 2001; Mojzsis et al. 2001; Watson and Harrison 2005; Harrison 2009). However, the timing of other geological processes and planetary conditions are rarely discriminated throughout the Hadean Eon. The onset of plate tectonics is thought by many geologists to have commenced prior to ~3 Ga (Condie and Kröner 2008; Dhuime et al. 2012; Korenaga 2013; Laurent et al. 2014; Dhuime et al. 2015; Condie 2016; Tang et al. 2016; Nebel-Jacobsen et al. 2018). However, there is no consensus on this issue, nor is it clear that plate tectonics operated in the same manner as it does today. Furthermore, despite estimates of early bulk atmospheric and ocean conditions (Sleep et al. 2001; Zahnle et al. 2007; Abe et al. 2000; Kasting 2014), changes in atmospheric composition during the Hadean and Archean are less well defined. Therefore, specific and overlapping conditions are difficult to distinguish within the Hadean and early Archean. Furthermore, the broad understanding of Hadean conditions has not yet been translated to specific early Earth environments, their specific physiochemical conditions, the co-variation of these conditions, the flow dynamics, or the thermal and chemical gradients contained within them. Yet it is exactly these specific, co-varying, and dynamically interconnected environmental conditions that gave rise to life’s chemistry. The global extent and temporal ambiguity of these conditions further limit the extent to which prebiotic chemistry research can truly reflect early Earth environments.
The importance of constraining environmental conditions to better characterize early Earth prebiotic chemistry is emphasized in Chapters 1 and 2 of the 2015 Astrobiology Strategy (NASA 2015), and integration of plausible early Earth conditions into prebiotic chemistry experiments is becoming more common. Prebiotic pathways constituted within complete suites of early Earth environmental conditions, however, are still out of reach. It is important that advances in understanding the abiotic inventory and baseline are grounded in understanding evolving environmental conditions over time and space. The field of prebiotic chemistry was originally bifurcated along the “genetics-first” and “metabolism-first” hypotheses (e.g., Gilbert 1986; Wachtershauser 1990), however, these often-competing hypotheses overlap in their general objective: How do abiotic synthesis reactions eventually lead to the chemistry of life? The true divergence lies in the chemical targets: Did the primitive chemistry that led to life actually consist of metabolic precursors and primitive metabolic cycles, or the nucleic acids, proteins, and lipids that now comprise the central dogma of biology?
Specifically, origins-of-life research is largely focused on the abiotic reactions that lead to the initial reduction of CO2 to simple organic compounds, their synthesis into more complex monomers (e.g., nucleotides, amino acids) and/or metabolic precursors (e.g., pyruvate, acetic acid), and either the polymerization into functional polymers or a sequencing of reactions into prebiotic metabolic pathways. Laboratory experiments to demonstrate these processes have limited success under ideal conditions, and so the applicability of such experiments to early Earth is often limited to singular environmental parameters (e.g., mineralogy, temperature, etc.) instead of a full accounting of system-wide, co-varying conditions.
Recent investigations are targeting more plausible physicochemical conditions and sources of precursors and essential elements. For example, the role of minerals in prebiotic chemistry is a focus of much of the current experimental research. While the potential for mineral catalysis of RNA polymerization was identified early on (Ferris et al. 1996), a wide variety of minerals more representative of a heterogeneous early Earth are now
considered to be important for the concentration, adsorption, alignment, and polymerization of not only RNA, but a variety of nucleic acids, amino acids, and sugars (Anizelli et al. 2015; Estrada et al. 2015; Feuillie et al. 2015; Hashizume 2015; Pandey et al. 2015; Schwaminger et al. 2015; Bhushan et al. 2016; Gregoire et al. 2016; Iqubal et al. 2016; Nitta et al. 2016; Pedreira-Segade et al. 2016; Yang et al. 2016; Biondi et al. 2017; Fuchida et al. 2017; Georgelin et al. 2017; Guo et al. 2017; Iqubal et al. 2017; Kitadai et al. 2017; Pedreira-Segade et al. 2018; Villafane-Barajas et al. 2018; Yamaguchi et al. 2018). Mineral surfaces may serve as catalysts for the initial reduction of CO2 in both surface and subsurface environments (Varma et al. 2018; Steele et al. 2016; Braterman et al. 1983; Kim at et al. 2013; White et al. 2015; Martin et al. 2008; Herschy et al. 2014) and may give rise to biomolecular chirality (e.g., Hazen and Sverjensky 2010; Schrader 2017). Furthermore, specific minerals (e.g., schreibersite, chimney precipitates) are being invoked either as meteoritic or terrestrial sources of phosphorus or catalysts for phosphorylation (Pasek et al. 2015; La Cruz et al. 2016; Fernandez-Garcia et al. 2017; Pasek 2017; Holm 2014; Barge et al. 2014). Phosphorus may also have been available in the highly soluble form of phosphite, generated by reduction of phosphates through its reaction with Fe(II) (Herschy et al. 2018).
In addition to the variety of mineralogies available on the early Earth, the effects of individual parameters, including pH, salinity, fluid chemistry, and pressure on a range of prebiotic processes—from CO2 reduction to RNA polymerization—are being investigated to discriminate those physicochemical conditions that could or could not give rise to viable prebiotic chemistry (e.g., Cody et al. 2000; Lazar et al. 2015; Hazen et al. 2002; Kitadai et al. 2017; Pedreira-Segade et al. 2016; Pedreira-Segade et al. 2018; DaSilva et al. 2015; Nitta et al. 2016; Dalai et al. 2018; Maurer and Nguyen 2016; Maurer 2017). Even more traditional prebiotic synthesis experiments strive to identify precursors that are plausible for early Earth (e.g., Bada et al. 2016; Cafferty et al. 2016a; Powner et al. 2009; Sutherland 2016). In addition to system components, specific dynamics within geologic environments—including flow, mixing, and freeze-thaw and wet-dry cycles that concentrate and dilute solutions—are gaining prominence as important considerations for nucleotide formation, peptide and nucleic acid polymerization, and the rise of metabolic cycles (Forsythe et al. 2015; Damer and Deamer 2015; Ross and Deamer 2016; Ritson et al. 2018; McGlynn et al. 2012; Burcar et al. 2015; Baaske et al. 2007). Further, ultraviolet radiation is more commonly integrated into prebiotic synthesis experiments, bringing renewed interest to hypotheses about a surface emergence of life (e.g., Nuevo et al. 2012; Ritson et al. 2018; Patel et al., 2015, Fornaro et al. 2013; Bacchus-Montabonel 2013; Benner et al. 2010; and Pearce et al. 2017).
While formerly associated solely with the metabolism-first theory, the role that both marine and continental hydrothermal systems may play in the origin of life (Corliss 1990), and specifically in the formation, concentration, and polymerization of RNA (Milshteyn et al. 2018; Burcar et al. 2015; DaSilva et al. 2015; McGlynn et al. 2012; Damer and Deamer 2015), amino acids (Fuchida and Masuda 2015; Kitadai 2015; Zhang et al. 2017; Estrada et al. 2017), and other prebiotic scenarios (Yang et al. 2018; Niether et al. 2016; Damer and Deamer 2015), is becoming more widely considered. Of particular interest is the combination of physical and chemical parameters that produce thermal, pH, and other chemical gradients along with pore spaces that can drive chemical differentiation (Agerschou et al. 2017; Baaske et al. 2007; Keil et al. 2016; Kreysing et al. 2015; Möller et al. 2017; Herschy et al. 2014; Barge et al. 2014). Outside of the environmental parameters, alternative and multicomponent, multipolymer chemistries and processes are also challenging more long-established hypotheses and inspiring innovative research avenues (Jauker et al. 2015; Sproul 2015; Cafferty et al. 2016b; Griesser et al. 2017; Patel et al. 2015; Caetano-Anollés and Seufferheld 2013). Finally, the production of biotic precursors, specifically through proto-metabolic pathways during water-rock reactions, are reinvigorating ideas about the rise of metabolism (McGlynn et al. 2012; McDermott et al. 2015; Herschy et al. 2014; Varma et al. 2018; Westall et al. 2018). The chemistry of reductive homologation of hydrogen cyanide and other nitriles has been experimentally demonstrated to produce precursors of ribonucleotides, amino acids, and lipids. In this hypothesis, all necessary cellular subsystems could have arisen simultaneously through a common chemistry in reactions which require ultraviolet light and use hydrogen sulfide as the reductant (e.g., Powner et al. 2009; Patel et al. 2015). Sutherland, Powner, and colleagues continue to explore nitrile chemistry in holistic, systems approaches to the generation of all prebiotically important compound classes (Sutherland 2016; Islam and Powner 2017) advancing the early ideas of Oró (1960) and elaborated by Eschenmoser (2007). A similar systems approach is needed to explore various potential pathways that lead from chemistry to biology (Szostak 2017). Various elements, including divalent metal cations and iron-sulfur clusters, could have assisted in these prebiotic reactions and served as cofactors for the earliest catalysts.
Incongruities between experimental conditions and realistic early Earth parameters still challenge experimental progress. Recently, individual experimental parameters (e.g., specific chemical precursors or specific minerals) have been increasingly scrutinized for early Earth plausibility. However, it is rarely the case that the overall system conditions tested for prebiotic potential include multiple co-varying environmental parameters for a single early Earth environment or associations of early Earth environments. This is in large part due to an insufficient understanding of early Earth environmental conditions in specific geologic settings throughout the Hadean. The global constraints that do exist for Hadean conditions are not directly translatable to specific environments or parameters with the overlapping timing of key global parameters (e.g., onset of plate tectonics, decrease of atmospheric CO2, etc.). Furthermore, the subsequent impact of which environments and conditions were contemporaneously available have yet to be explored, although an attempt has been made to extrapolate this information from early Archean environments (Westall et al. 2018). Within the context of life’s origins and the requirements of prebiotic chemistry that might be tested on other planetary bodies and moons, the temporal and spatial scales of what we need and what we can look for are even more disparate.
Finding: Considering the coevolution of early Earth environments in the context of multiple parameters (e.g., temperature, pressure, and pH) and over a range of spatial and temporal scales advances the integration of prebiotic chemistry and origins of life research.
Moving from “Where Did Life Emerge?” to “Where Could Life Emerge?”
Constraining the specific location for the emergence of life on early Earth is a robust area of investigation and is still hotly debated. However, investigations of Earth’s potential prebiotic chemistry have highlighted the overall global planetary conditions and specific types of environments that might prove to be viable targets for the rise of life elsewhere. Smith and Morowitz (2016) suggest that the chemical potential of planetary-scale disequilibrium on Earth is focused “to an extreme degree on the rock/water interface and in the mixing chemistry of fluids and volatiles in and near the crust,” rendering these environments the most likely for life’s origins—an insight particularly pertinent for astrobiological exploration of ocean worlds (ROW 2017). However, it is still widely debated whether the emergence of Earth’s life required a strictly surface environment in direct contact with the atmosphere and incoming ultraviolet radiation, or if a subsurface hydrothermal system might have provided the disequilibria necessary for life’s origins. What is clear is that while planetary-scale disequilibrium might be the driving force for life’s emergence, it is the local-scale manifestations of disequilibrium that determine the outcomes for prebiotic synthesis and life’s coevolution with a planet. Both planetary-scale and local-scale disequilibria are important for prebiotic synthesis and life’s coevolution with the planet, therefore, consideration of both planetary-scale habitability and localized environments are important in designing successful strategies for searching for life beyond Earth.
What is known of the necessary planetary conditions for the emergence of life on early Earth is applicable to the search for life, if any, on other worlds. For the ocean worlds of the outer solar system, integrating the entire system into the discussion of habitability and the search for life allows the exploration of these worlds to be prioritized and planned. Over the past 20 years, the sophistication of understanding of these systems has grown dramatically. Knowledge of the system as a whole shifts focus from the search for water to the search for many parameters that are needed to support life, such as chemical energy and geologic activity. For example, the surfaces of the various ocean worlds have different ages, implying different levels of present day activity that span from vigorous resurfacing in the case of Europa (e.g., Pappalardo et al. 1998; Kattenhorn and Procter 2014; Schmidt et al. 2011; Collins and Nimmo 2009) to relative quiescence in the case of Callisto (e.g., Greeley et al. 2000). Based on understanding of terrestrial processes, geologic activity of any type—including, but not limited to, plate tectonics, resurfacing, geothermal activity, or fluid circulation—is part of the formula for habitability. Additionally, while the timing of the origin of life on Earth is not known, presumably there was a characteristic time when a balance of environmental stability and chemical and thermodynamic disequilibria was needed for life to begin. This perspective allows consideration of if and when such a system may have existed on the ocean worlds. A comprehensive understanding of the interplay of chemistry, geology, planetary structure, endogenic, and exogenic energy sources
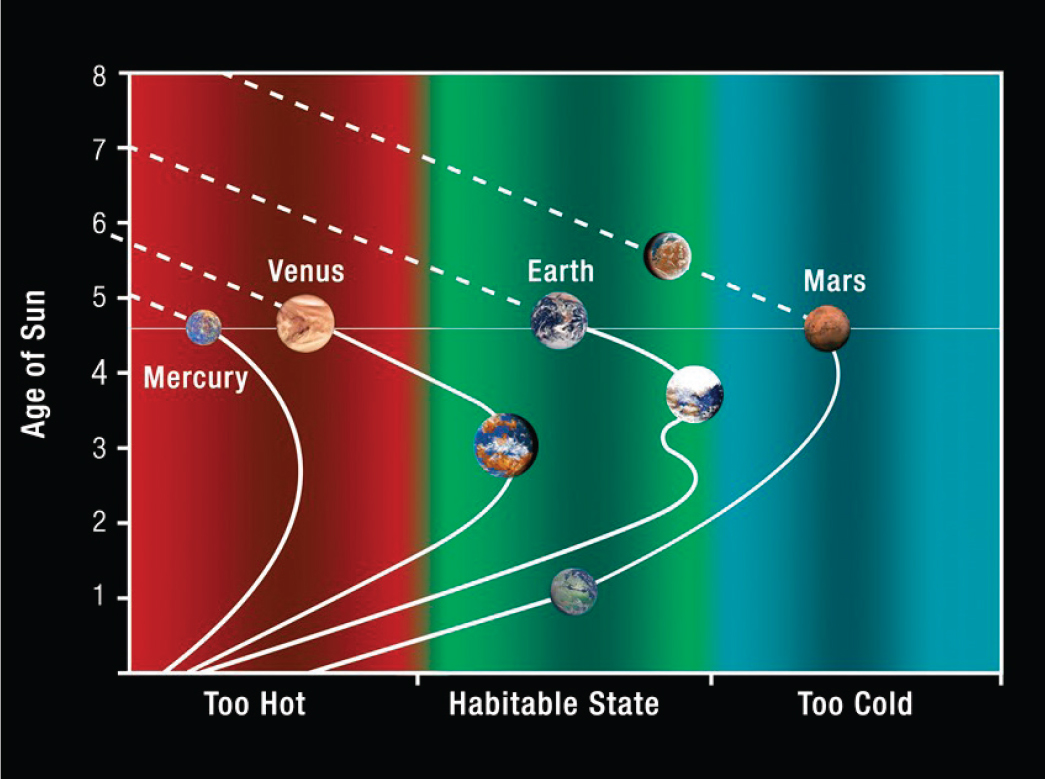
is required to understand how these worlds could operate as systems. Figure 2.1 illustrates an example of this interplay for the inner solar system’s rocky planets as dimensions of habitability evolve with the age of the Sun.
Finding: Planetary conditions that may be habitable today or in the past are not necessarily the same as those that could have fostered the emergence of life. Both are important for the search for life.
Finding: Due to the complexity of interactions between physical, chemical, and biological parameters and processes, habitability is not a binary property but is best thought of as a continuum of degrees of habitability.
Finding: Dynamic habitability and the coevolution of planets and life provides a powerful foundation upon which to integrate diverse astrobiology communities focusing on Earth, the solar system, stellar astronomy, and exoplanetary systems.
Recommendation: NASA and other relevant agencies should catalyze research focused on emerging systems-level thinking about dynamic habitability and the coevolution of planets and life, with a focus on problems and not disciplines—that is, by using and expanding successful programmatic mechanisms that foster interdisciplinary and cross-divisional collaboration.
EXAMPLES OF RECENT ADVANCES IN UNDERSTANDING OF DYNAMIC HABITABILITY
The definition of habitability is an environment’s ability, or inability, to support life, which can be misinterpreted as a simple yes/no proposition. Recent advances in dynamic habitability have blurred this black-and-white definition. First, dynamic habitability encompasses the transition of an environment from abiotic or prebiotic to biotic, or from biotic to abiotic (e.g., due to a catastrophic event such as large impact or stellar flares). Second, dynamic habitability addresses the spatial and temporal intersections of habitable environments with uninhabitable ones. Increasingly, astrobiologists have scrutinized these intersections by studying life living in extreme and often isolated environments on Earth. As in the case of refugia, such environments occur against the backdrop of conditions largely considered uninhabitable, and the isolated communities inhabiting these specialized niches demonstrate how habitability can be a local phenomenon in both space and time. In other cases, the extreme conditions are more generalizable to surface conditions but push the boundaries of conditions understood as being “habitable.”
Another area of significant advance in dynamic habitability has been the discovery of a previously unknown subsurface biome. While planetary surfaces receive abundant energy from the light of their host star, they are also subject to potentially sterilizing events such as giant impacts and stellar flares. Subsurface environments are potentially protected from such disruptive events. On Earth’s surface, sunlight drives photosynthetic ecosystems—those based upon the products of photosynthesis. In the subsurface, however, biological communities are increasingly found to exist beyond the direct influence of sunlight and can be entirely sustained by chemical energy—chemosynthetic communities. Examples of such communities include life in the deep Earth, in subsurface aquifers, caves, and life at and below the seafloor. Although many such organisms may have been common on early Earth (Westall et al. 2015a,b), as many of these subsurface ecosystems are explored for the first time on the modern Earth, their expressions at the surface and in the atmosphere remain unknown. Efforts to consider their impact are tied to a better understanding of the thermodynamic drivers of the ecosystems—whether abundant energy is available or not, and if the life exhibits vigorous, slow, or even dormant metabolic processes. With increasing interest in extreme surface and subsurface environments on other planets, understanding the role of these systems and their impact on planetary evolution and life detection is critical, making them a frontier of astrobiological research for which a number of notable examples are provided here.
Refugia and Ephemeral Habitats
Biological ecosystems develop to thrive in specific niches. Given the generally clement conditions of Earth’s surface today, it is common to think of niches as spatially large and connected and temporally long-lived. This need not be the case universally. Microbial life in the Atacama Desert of Chile provides a case in point. In the hyperarid core of the desert, photosynthesizing microbial communities exist on the undersides of translucent rocks. The rocks provide protection from desiccation and solar ultraviolet radiation while their translucence allows the microbes to access sufficient light for photosynthesis. These communities are rare and occur in small, isolated islands surrounded by microbially poor soil (Warren-Rhodes et al. 2006). Similarly, endolithic communities are found sparsely distributed throughout terrestrial hot spring systems (e.g., Rogers et al. 2014; Hynek et al. 2011; Walker et al. 2005) (Figure 2.2). Such examples highlight how finding microbial life in habitable, albeit extreme, environments requires an understanding of environmental controls and their interactions with biologic processes integrated over space.

These Earth examples increase the range of possible niches in which to search for signs of extinct or extant life on the surface of Mars. The search becomes not only for habitats that may have become inhabited as or after life emerged, but also for locations where life or signs of extinct life may persist despite widespread loss of surface habitability. Current understanding of the martian surface during the pre-Noachian and early Noachian (Box 2.3) suggests that life could have emerged on the planet (McKay 2010; Westall et al. 2015a). Life could not have emerged everywhere, however, and the locations that would have had the right conditions for hosting the emergence of life would have been isolated both in space and time (Westall et al. 2015a). Furthermore, life it seems could not emerge there today in any known surface environment. There exists the possibility, however, that viable cells on Mars could survive and maybe flourish for short periods in isolated, transient habitable niches at the surface (Rettberg et al. 2016). In addition to developing the hypothesis that it is not necessary for the entirety of the surface of an extraterrestrial body to be habitable for the emergence or persistence of life, this example demonstrates that integration of environmental controls and their interactions with biological processes is one that occurs over not just space, but time as well (Cabrol 2018).
The view of an ancient Mars surface dotted by habitable surface environments such as lakes and playas (Ehlmann et al. 2008; Squyres et al. 2004; Grotzinger et al. 2005; Wray et al. 2011; Michalski et al. 2013), and aquifers and hydrothermal systems (Skok et al. 2010; Ehlmann et al. 2011; Saper and Mustard 2013) has been enhanced by results from landed missions to Mars including Viking, Mars Pathfinder, Phoenix, the Mars Exploration Rovers, and Mars Science Laboratory, as well as the orbital missions Mars Global Surveyor, Mars Odyssey, Mars Express, Mars Reconnaissance Orbiter and Mars Atmosphere and Volatile Evolution mission (Arvidson 2016). Recently, findings of the ongoing Mars Science Laboratory mission have documented the potential for ancient Gale Crater sediments to provide nutrients and energy for life (Grotzinger et al. 2014); combined with the presence of water, the crater could have served as an ephemeral habitat for life. Since publication of the 2015 Astrobiology Strategy, however, attention has been turning increasingly to the idea of the modern martian surface as one existing on the edge of habitability (Ehlmann 2018). The identification of ephemeral martian surface habitats—existing from days, as in the case of the isolated environments above, to hundreds of thousands of years—is an area of increasing interest.
This interest has been driven in part by a rapidly evolving understanding of the martian cryosphere. Radar sounding of the south polar ice deposits has revealed volumes of carbon dioxide ice that, if released as a gas, could
double its atmospheric pressure (Phillips et al. 2011; Bierson et al. 2016). This would bring more of the martian surface above the triple point of water (Ehlmann 2018). Bierson et al. (2016) hypothesize that this has occurred as recently as within the last 1 Ma, possibly even multiple times. At the same time, the inventory of near-surface water ice reservoirs—including, but not limited to, pore-filling ice in the regolith (Boynton et al. 2002; Pathare et al. 2018), mid-latitude glaciers (Holt et al. 2008), and contiguous sheets of mid-latitude ice (Dundas et al. 2018)—is large (Carr and Head 2015) and increasing (Dundas et al. 2018; Pathare et al. 2018) (Figure 2.3). In concert with the possibility of periodic increases in atmospheric pressure, the extensive nature of near-surface water ice deposits raise the potential for formation of ephemeral surface water, thereby increasing chances for habitability on timescales of obliquity changes (Ehlmann 2018).
Over the past 15 years, discoveries from both spacecraft in orbit and landed on Mars have sparked a major revision in our thinking about the nature of diagenetic fluids and subsurface groundwater on the planet and their role in habitability over time (Ehlmann and Edwards 2014; Ehlmann et al. 2016, Grotzinger et al. 2015; Goudge et al. 2015). The recent report of strong seasonal variation in background levels of methane in the Mars atmosphere, and the resultant renewed investigation of sources(s) for that methane underscores the necessity to consider both surface and subsurface processes (Webster et al. 2018, 2015). Extending the lessons learned from terrestrial analogs about subsurface habitability, fluid history and water-rock reaction, rock-hosted life, and preservation to other space exploration targets such as Mars is valuable even while recognizing the challenges of accessing non-terran subsurface systems.
Saline and Hypersaline Environments
The martian example above alludes to the possibility of forming water through melting when atmospheric pressure is raised. Investigations on Earth and Mars, however, demonstrate that other means may exist for forming ephemerally habitable aqueous environments. For example, salts increase the range of environmental conditions under which liquid water may exist (Fischer et al. 2014). Salts have the following three impacts on liquid water that are pertinent when considering the possibility of life on other planets (Haberle et al. 2001; Chevrier et al. 2009; Rennó et al. 2009; Gough et al. 2011; Fischer et al. 2016):
- They lower the freezing temperature of water, allowing it to remain in a liquid state at temperatures below 0°C;
- They decrease the partial pressure of water vapor, in equilibrium with aqueous solutions, lowering evaporation rates in tenuous dry atmospheres; and
- They form crusts on the surfaces of liquids, which further inhibit evaporations.
These three effects increase the range of conditions in which brines can be stable.

On Earth, a diverse array of terrestrial microorganisms thrives in the hypersaline habitats created by brines (Boetius and Joye 2009). These environments (e.g., Figure 2.4) include subsurface saline aquifers, deep-sea brine pools, ancient subglacial brine reservoirs (Green and Lyons 2009; Mikucki et al. 2009; Joye and Samarkin 2009; Biddanda et al. 2009; Thomas and Dieckmann 2002), and surface bodies of water, like the Dead Sea (Arahal et al. 1996). Examining the communities in these environments expands the known limits of life. For instance, the discovery of sulfur- and iron-cycling microbial communities in the subglacial brine that episodically drains from Taylor Glacier in Antarctica’s Dry Valleys (Mikucki et al. 2009) is particularly relevant to understanding habitability on other worlds. The cycling of sulfur, methane, and iron in hypersaline habitats in the absence of sunlight underscores the metabolic flexibility of extremophiles (e.g., Boetius and Joye 2009; Li et al. 2016; Magnabosco et al. 2018). Microbes thriving in similar brine channels and brine inclusions in sea ice, which concentrates nutrients and organics, making them readily available for microbial consumption (Junge et al. 2001), also may apply to ocean world environments. In cold worlds like Europa, tidal heating could cause the formation of regions that could produce liquid brines and preserve them in cracks and crevasses like those found in terrestrial sea ice (Junge et al. 2001). If such regions exist in the subsurface of Europa and other icy worlds, they may be potentially habitable.
In another example of microbe resiliency and persistence in hypersaline environments, halophilic archaea isolated from Dead Sea samples more than 50 years after their collection (Arahal et al. 1996) indicate low mortality rates in briny environment (even up to 20 percent salt content; Boetius and Joye 2009). Notably, similar halophilic organisms are found in many branches of the tree of life, including archaea and bacteria. This suggests that either adaptation to saline environments occurred numerous times, or that lateral gene transfer occurred during their evolution.
On Mars salts capable of forming aqueous solutions at martian temperatures, such as perchlorates of calcium, magnesium, and sodium, were discovered in the polar and equatorial regions (e.g., Hecht et al. 2009; Glavin et al. 2013; Ming et al. 2014; Martin-Torres et al. 2015). Such perchlorates may be responsible for the formation of ephemeral brine on the surface of Mars (e.g., Rennó et al. 2009; McEwen et al. 2011, 2014; Ojha et al. 2015). One proposed manifestation of modern brines on the surface are recurring slope lineae (RSL)—annually occurring, narrow lineae of darker albedo than underlying material that occur on steep slopes and lengthen through the warm season (McEwen et al. 2011). By what process RSL form, however, is still ambiguous, as they may also form by dry granular flow (Stillman and Grimm 2018). However, if RSL are formed by brines, they are of interest because they could ephemerally host microbial cells (Rettberg et al. 2016). Moreover, RSL may point to more stable environments in the subsurface.
Continental Subsurface
Deep marine, chemosynthetic life has long been of interest. Investigations of such communities are now expanding to include subsurface continental settings (Suzuki et al. 2017; Kietavainen et al. 2017; Magnabosco et al. 2018). Studies in Precambrian cratons have transformed our understanding of the habitability of Earth’s subsurface in fracture water located deep within crystalline rocks that are billions of years old. Precambrian (and specifically Archean) settings have been the focus of many analog studies for Mars. Deep groundwaters in rocks more than 3 Ga old reveal preservation of fluids with residence times ranging from millions to billions of years (Lippmann-Pipke et al. 2011; Holland et al. 2013; Warr et al. 2018). Serpentinization relates oxidation of FeO in the crust to H2 production (Sleep and Bird 2007). Other moderate- to low-temperature processes of water-rock reaction producing hydrogen-, methane-, and sulphate-rich fracture fluids include radiolysis (Lin et al. 2005a,b; Li et al. 2016) and a range of other hydration reactions acting on mafic and ultramafic minerals (Sherwood Lollar et al. 2014; Mayhew et al. 2013). Such processes have also been invoked in the context of understanding production of methane on Mars (Webster et al. 2018; Atreya et al. 2007; Chassefiere et al. 2016) and the role of water-rock interactions in potential habitability of ocean worlds and icy bodies such as Europa and Enceladus (Bouquet et al. 2017; Hand et al. 2016; Waite et al. 2017).
Expanded understanding of subsurface habitability on Earth has implications for the search for life on Mars, where ionizing radiation and oxidation of the upper 1.5 to 3 m pose two great challenges to the existence of life on the surface (Eigenbrode 2018). Speculation about potential habitability arising from subsurface water at depth (Ehlmann 2018) or life in shielded lava tubes or caves (Leveille and Datta 2010; Blank et al. 2018) has motivated

the search for life in the subsurface of that planet. To this end, the Roscosmos and European Space Agency joint ExoMars 2020 rover, whose goal is to search for signs of past or present life, will carry a drill able to reach 2 m below the surface of the planet. Samples taken from this depth will be retrieved and analyzed for signs of organics or possible visible biosignatures using the rover’s instrument suite (Vago 2018). The NASA Mars 2020 (see Box 6.1) rover will also carry a drill for the recovery of shallow (i.e., to a depth of approximately 5 cm) subsurface samples; however, those samples will be cached on the surface for return to Earth at an unspecified time in the future (Ehlmann 2018). The sites to be sampled will be identified using instruments on the rover, but detailed studies will be done once the samples have returned to Earth.
Full consideration of in situ astrobiological analyses and future missions targeting the shallow subsurface of Mars is absent in the NASA portfolio and needs considerable development in the upcoming decade. Shallow planetary subsurfaces are easily accessible and extremely interesting as a possible microbial habitat when they are wet. They are shielded from radiation, and mixing by a variety of surface processes that impact the shallow subsurface could provide the nutrients necessary for sustaining microbial life. On Mars, aeolian abrasion is one of the most important, currently active, weathering processes. Chemical reactions induced by aeolian process could also have important implications for life and habitability (e.g., Atreya et al. 2006; Kok and Rennó 2009).
The growth of salt crystals in ice cracks and rock pores is another important weathering process. There is also evidence that this process is currently active on Mars. Salts have the potential of creating pockets of wet brines in the shallow subsurface of Mars (Fischer et al. 2014, 2016). Comprehensive measurements of regolith wetness and the possible detection of brines could elucidate habitability and aqueous weathering processes not only on Mars, but also on other potentially habitable worlds. Measurements of dry weathering processes, such as abrasion by moving sand grains, could elucidate the supply of nutrients necessary for sustaining microbial life in shallow planetary subsurfaces. Small, power-efficient instruments for studying aeolian processes, weathering, and regolith wetness will be useful for enabling the collection and analysis of samples of the most astrobiologically relevant areas.
Marine Subsurface and Ocean Worlds
Deep submarine oases, such as hydrothermal vents (Figure 2.5), highlight the potential for extensive ecosystems in highly dynamic tectonic settings. Active microbial communities, however, have also been found throughout the more widespread and less dynamic crystalline and sedimented oceanic crust (Edwards et al. 2012; Kallmeyer et al. 2012; Colwell and D’Hondt 2013). These microbial communities are both chemosynthetic- and photosynthetic-based, are supported by a variety of energy and carbon sources, and proliferate over a broad range of temperature, salinity, pH, and redox conditions. Yet fundamental questions remain unanswered: How much of the total planetary biomass is represented by these subsurface communities? What are the mechanisms for prokaryotic life’s dispersal and transport? How do biodiversity and total biomass vary with depth below surface? What role do the combined effects of temperature, pH, redox conditions, and pressure play in microbial adaptation and survivability?
The isolation of several high-pressure, or piezophilic, microbial strains as well as recent experiments showing that decompression of piezophiles causes cell death, underscore the need to include the effects of pressure when considering habitability and life detection. On Earth, piezophiles have been isolated from deep terrestrial boreholes and mines, deep ocean basins, and deep sea hydrothermal vents (Abe and Horikoshi 2001). Studies to understand the high-pressure biosphere have pushed the boundaries for high-pressure growth to 140 MPa (Kusube et al. 2017) and high-pressure survival up to 2 GPa (Sharma et al. 2002; Vanlint et al. 2011). Furthermore, growth at elevated pressures has also extended the known high temperature limit of life (Takai et al. 2008) to 122°C, as well as the high temperature limit of several individual species of hyperthermophiles (Zeng et al. 2009). The link between high-pressure and high-temperature organisms exemplifies the criticality of evaluating habitability through the lens of interconnected environmental parameters. In addition to fundamental questions about the nature and activity of life in the marine subsurface, recent research is advancing the frontiers of understanding of procedural best practices—for example, optimizing sensitivity of life detection and characterization in the context of challenges of contamination, related to both sample retrieval and sample handling (Sheik et al. 2018).
As understanding of extreme marine life on Earth is advancing, so too is the discovery that oceans in the outer solar system are relatively common (Khurana et al. 1998; Kivelson et al. 2000; Husmann et al. 2006; Thomas et al. 2007; Iess et al. 2012, 2014; Keane et al. 2016).

For many of these worlds, the most likely habitats are below the surface. On Europa (Figure 2.6), energy for metabolic processes at the seafloor could derive from tidal activity, radiogenic heating, and ongoing serpentinization that both release energy and could cause fractures to allow active seafloor alteration (Vance et al. 2016). Additional habitats within Europa’s ice shell may also exist. These range from veins along grain boundaries, as seen in glacial and accretion ice on Earth (Priscu and Christener 2004), to small distributed brine zones from partial melting (Collins et al. 2000; Pappalardo and Barr 2004), to larger subsurface melt pockets induced by tidal convection or diapirism, or eutectic melting (Showman and Han 2004; Sotin and Tobie 2004; Schmidt et al. 2011). Icy habitats may be linked to sub-ice ocean processes or carry with them a record of ocean conditions (Vance and Goodman 2009; Soderlund et al. 2014). Recent, but indirect, reported detections of plume activity at Europa (Sparks et al. 2016, 2017) are consistent with the release of water vapor by a process that does not necessarily depend upon tidal forcing (Roth et al. 2014a,b), which is insufficient to fracture Europa’s ice shell (Goldreich and Mitchell 2010). Therefore, it is possible that these plumes originate from within the ice shell rather than the ocean (Walker and Schmidt 2015). This suggests that understanding the interactions between surface, oceans, and the underlying subsurface are likely critical for understanding astrobiological implications (ROW 2017).
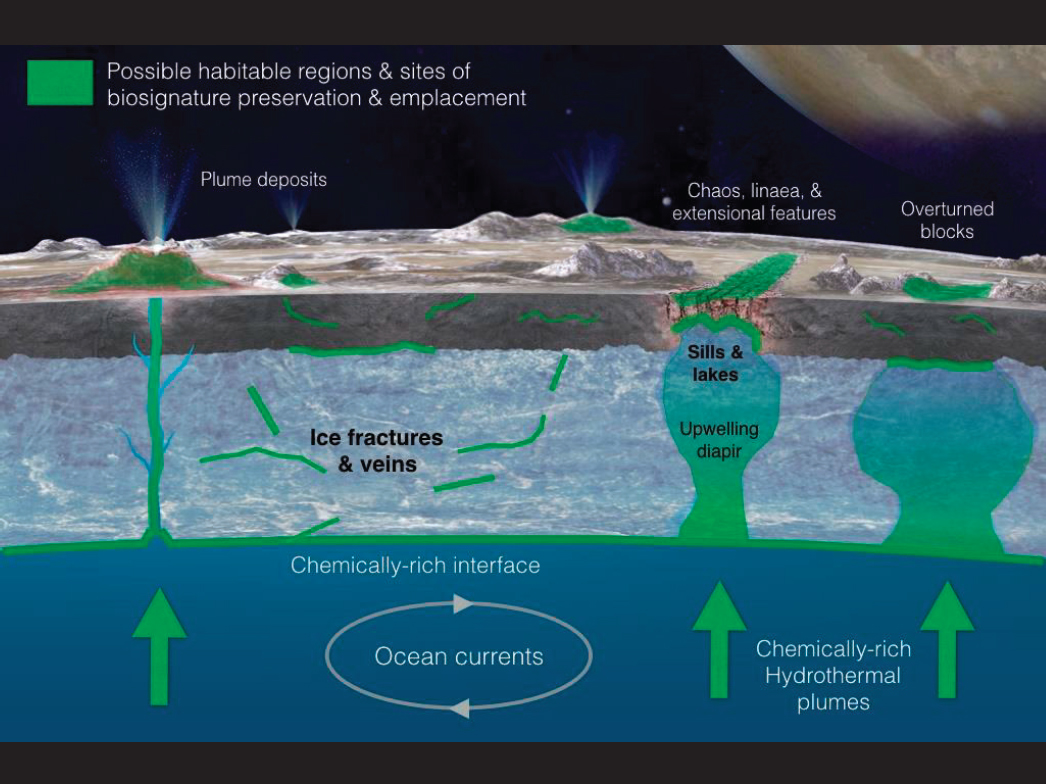
Enceladus, too, likely possesses a subsurface ocean (Iess et al. 2014) that is released by a south-polar plume consisting of many individual jets (Porco et al. 2006; Spencer et al. 2006). Enceladus has likely become active in the recent geologic past (Meyer and Wisdom 2007), and its activity may be punctuated (Nimmo and Pappalardo 2006; Meyer and Wisdom 2008; O’Neill and Nimmo 2010). Although Cassini data do not clearly articulate other regions of present day activity, the interpreted presence of crater relaxation (Bland et al. 2012) and thermal-infrared measurements by Cassini suggest high heat flow. The ice shell, ocean, and seafloor may all provide habitable environments, particularly given observations of plume chemistry that indicates hydrothermal activity and the production of hydrogen, which can serve as an electron donor for both energy transduction and the reduction of carbon dioxide for biosynthesis (Waite et al. 2006; Postberg et al. 2011; Hsu et al. 2015; Glein et al. 2015; Waite et al. 2017). Further, seafloor serpentinization (Hsu et al. 2015) may make life possible through the production of chemical energy in the form of methane and molecular hydrogen (Waite et al. 2017).
The plumes emanating from Enceladus, and potentially Europa, provide evidence of relatively intense cryoactivity reshaping their icy crusts, and modeling suggesting subsurface oceans has led to increased interest in the potential habitability of these two moons. Metabolisms fueled by natural radiogenic energy (i.e., radiolysis of water at the rock-water interface as used by sulphate-reducing bacteria deep in Earth’s crust; see Lin et al. 2006; Onstott
2016; Li et al. 2016) could occur on icy bodies. Further, the detection of hydrogen and methane—which indicate ongoing hydrothermal rock-water interactions like those favored in scenarios of the emergence of terran life—and formaldehyde suggest prebiotic conditions in Enceladus’ oceans (Waite et al. 2017). In light of these discoveries, there is debate as to whether remote plume characterization to detect biosignatures that consider molecular composition, organic complexity, and Gibbs free energy is sufficient for life detection, or if obtaining a plume sample in a flyby for onboard analysis and/or sample return is mandatory (see Chapter 6).
Titan’s surface reservoirs of liquid hydrocarbon are of interest to the astrobiology community (see Box 3.1), but the moon also harbors a deep subsurface global ocean (Iess et al. 2012). While Ganymede and Callisto also have subsurface global oceans, the extreme thickness of these moons’ ice shells impedes interactions between the surface and subsurface, which has its own implications for habitability. At Titan and Ganymede, dense, high-pressure ice layers may separate their oceans from their silicate mantles, which could inhibit water-rock reactions, such as serpentinization, postulated to be sources of chemical energy for life. Knowledge of serpentinizing systems on Earth is driving interest in the seafloor interactions on ocean worlds. Measurements of the properties of these oceans, while challenging to make, have begun a discussion about how to interpret chemical signatures of water-rock reactions. It is unclear whether Europa’s ocean pH (Glein et al. 2015, suggested a pH of 11 or 12) would foster habitability or indicates serpentinization gone to completion, fully reducing Europa’s ocean. Small silica particles observed in Enceladus’ plume (Hsu et al. 2015) were interpreted as evidence of habitability through ongoing hydrothermal activity, and the detection of molecular hydrogen would seem to reinforce this assumption, because products available for microbial metabolisms exist (Waite et al. 2017). While the very presence of this hydrogen may suggest it is not actively being consumed, the discovery of extant microbial communities of hydrogen-utilizing, sulfate-reducing bacteria in Earth’s subsurface, where ambient hydrogen concentrations are at the millimolar level, underscores the need to consider the effects of relative rates of production and storage of electron donors and acceptors (controlled by porosity and permeability as well as the rate of water-rock reaction), versus the potentially slow rate of metabolic consumption in subsurface environments (Lin et al. 2006; Hoehler and Jorgenson 2013; Sherwood Lollar et al. 2014; Li et al. 2016).
Finding: Expanded understanding of habitability of chemosynthetic subsurface environments, brine stability, and adaptations of life to saline fluids have widespread implications for the search for life in the solar system.
Fast versus Slow Life
The examples given above demonstrate that there are a range of potential environments for life, from the austere to those having rich natural resources and diverse local conditions. These environments provide a corresponding range of opportunities for life’s emergence, differentiation, and dispersal. The more complex environments provide the greatest opportunity for life but also give rise to more complex abiotic processes that raise background complexity and challenge-life detection studies. There is a gradient for both the richness of potential signatures of life and the degree of potentially obfuscating nonbiological processes, and it is not clear where on that gradient the best opportunities for life detection are located.
Further confounding the search for life is the interplay between metabolic rates and background processes. As previously mentioned, identifying signatures of so-called “fast life” existing in energy-rich environments may prove to be more or less challenging than finding “slow life” in energy-starved environments. The example from Enceladus highlights this conundrum, where hydrogen may be present either because it is not being actively consumed or because its rate of production exceeds that of slow metabolic consumption. Increasingly, research into slow, and even dormant, life on Earth opens the possibility of discovering such entities on other worlds.
Another example of slow-growing cells to take into account when considering planetary habitability are those entities residing in Earth’s crust or deep sediments. Deep-sea sediments host viable cells down to depths of nearly 2 km (Ciobanu et al. 2014; Jørgenson and Marshall 2016). In oligotrophic environments, the low availability of both nutrients and water severely limits cell viability. Cells in this environment have a remarkably slow metabolic turnover of about one cell division per thousand years (Ciobanu et al. 2014; Braun et al. 2017). The fractured crust is a somewhat different environment, where water that has infiltrated deep fractures can entrain microbes
over long periods of time at depths up to 3 to 4 km (Onstott 2016). Each fracture system, some as small as 1 cm, is an oasis for life (Sleep and Zoback 2007; Sherwood Lollar et al. 2007), and the microbial biomass in these subsurface habitats is low compared to that at the surface. Autotrophy and methane cycling are the main means of metabolic support for these deep microorganisms, with cell turnover times that range from one to hundreds of years (Onstott 2016).
Finding: “Slow” life that is barely able to survive in an austere environment may be detectable because the noise level is low. Whereas, “fast” life in a rich environment may be detectable because the signal is high. Assessing the relative signal-to-noise ratio of each type of population in its given environmental context would help identify corresponding biosignatures that are most relevant and distinctive.
Recommendation: NASA’s programs and missions should reflect a dedicated focus on research and exploration of subsurface habitability in light of recent advances demonstrating the breadth and diversity of life in Earth’s subsurface, the history and nature of subsurface fluids on Mars, and potential habitats for life on ocean worlds.
REFERENCES
Abe, Y. 1993. Thermal and chemical differentiation of the terrestrial magma ocean. Pp. 41-54 in Evolution of Earth and Planets (E. Takahashi, R. Jeanloz, and D. Weidner, eds.). Geophysical Monograph 74, IUGG 14, International Union of Geodesy and Geophysics and the American Geophysical Union, Washington, DC.
Abe, F., and K. Horikoshi. 2001. The biotechnological potential of piezophiles. Trends in Biotechnology 19(3):102-108.
Abe, Y., E. Ohtani, T. Okuchi, K. Righter, and M. Drake. 2000. Water in the early Earth. Pp. 413-433 in Origin of the Earth and Moon (R.M. Canup and K. Righter, eds.). University of Arizona Press, Tucson, AZ.
Agerschou, E.D., C.B. Mast, and D. Braun. 2017. Emergence of life from trapped nucleotides? Non-equilibrium behavior of oligonucleotides in thermal gradients. Synlett 28(01):56-63.
Anizelli, P.R., J.P.T. Bau, F.P. Gomes, A.C.S. da Costa, C.E.A. Carneiro, C.T.B.V. Zaia, and D.A.M. Zaia. 2015. A prebiotic chemistry experiment on the adsorption of nucleic acids bases onto a natural zeolite. Origins of Life and Evolution of Biospheres 45(3):289-306.
Arahal, D.R., F.E. Dewhirst, B.J. Paster, B.E. Volcani, and A. Ventosa. 1996. Phylogenetic analyses of some extremely halophilic archaea isolated from Dead Sea water, determined on the basis of their 16S rRNA sequences. Applied and Environmental Microbiology 62(10):3779-3786.
Arvidson, R. 2016. Aqueous history of Mars as inferred from landed mission measurements of rocks, soils, and water ice. Journal of Geophysical Research: Planets 121:1602-1626.
Atreya, S.K., A.-H. Wong, N.O. Rennó, W.M. Farrell, G.T. Delory, D.D. Sentman, S.A. Cummer, et al. 2006. Oxidant enhancement in martian dust devils and storms: Implications for life and habitability. Astrobiology 6(3):439-450.
Atreya, S.K., E.Y. Adams, H.B. Niemann, J.E. Demick-Montelara, T.C. Owens, M. Fulchignoni, F. Ferri, and E.H. Wilson. 2006. Titan’s methane cycle. Planetary and Space Science 54(12):1177-1187.
Atreya, S.K., P.R. Mahaffy, and A.S. Wong. 2007. Methane and related trace species on Mars: Origin, loss, implications for life, and habitability. Planetary and Space Science 55:358-369.
Baaske, P., F.M. Weinert, S. Duhr, K.H. Lemke, M.J. Russell, and D. Braun. 2007. Extreme accumulation of nucleotides in simulated hydrothermal pore systems. Proceedings of the National Academy of Sciences U.S.A. 104(22):9346-9351.
Bacchus-Montabonel, M.-C. 2013. Looking at radiation damage on prebiotic building blocks. Journal of Physical Chemistry A 117(51):14169-14175.
Bada, J.L., J.H. Chalmers, and H.J. Cleaves II. 2016. Is formamide a geochemically plausible prebiotic solvent? Physical Chemistry Chemical Physics 18:20085-20090.
Barge, L.M., I.J. Doloboff, M.J. Russell, D. VanderVelde, L.M. White, G.D. Stucky, M.M. Baum, et al. 2014. Pyrophosphate synthesis in iron mineral films and membranes simulating prebiotic submarine hydrothermal precipitates. Geochimica et Cosmochimica Acta 128:1-12.
Benner, S.A., H.-J. Kim, M.-J. Kim, and A. Ricardo. 2010. Cold Spring Harbor Perspectives in Biology 2:a003467.
Bhushan, B., A. Nayak, and Kamaluddin. 2016. Catalytic role of manganese oxides in prebiotic nucleobases synthesis from formamide. Origins of Life and Evolution of Biospheres 46(2-3):203-213.
Biddanda, B.A., S.C. Nold, S.A. Ruberg, S.T. Kendall, T.G. Sanders, and J.J. Gray. 2009. Great Lakes sinkholes: A microbiogeochemical frontier. EOS Transactions 90(8):61-62.
Bierson, C.J., R.J. Phillips, I.B. Smith, S.E. Wood, N.E. Putzig, D. Nunes, and S. Byrne. 2016. Stratigraphy and evolution of the buried CO2 deposit in the martian south polar cap. Geophysical Research Letters 43:4172-4179.
Biondi, E., Y. Furukawa, J. Kawai, and S.A. Benner. 2017. Adsorption of RNA on mineral surfaces and mineral precipitates. Beilstein Journal of Organic Chemistry 13:393-404.
Bland, M.T., K.N. Singer, W.B. McKinnon, and P.M. Schenk. 2012. Enceladus’ extreme heat flux as revealed by its relaxed craters. Geophysical Research Letters 39:L17204.
Blank, J.G., T.L. Roush, C.L. Stoker, A. Colaprete, S. Datta, U. Wong, M. Deans, et al. 2018. “Planetary Case as Astrobiology Targets.” White paper submitted to the Committee on an Astrobiology Science Strategy for the Search for Life in the Universe.
Boetius, A., and S. Joye. 2009. Thriving in salt. Science 324(5934):1523-1525.
Bouquet, A., C.R. Glein, D. Wyrick, and J.H. Waite. 2017. Alternative energy: Production of H2 by radiolysis of water in the rocky cores of icy bodies. Astrophysical Journal Letters 840:L8.
Boynton, W.V., W.C. Feldman, S.W. Squyres, T.H. Prettyman, J. Bruckner, L.G. Evans, R.C. Reedy, et al. 2002. Distribution of hydrogen in the near surface of Mars: Evidence for subsurface ice deposits. Science 297(5578):81-85.
Braterman, P.S., A.G. Cairns-Smith, and R.W. Sloper. 1983. Photooxidation of hydrated Fe+2— Significance for banded iron formations. Nature 303:163-164.
Braun, S., S.S. Mhatre, M. Jaussi, H. Røy, K.U. Kjeldsen, C. Pearce, M.S. Seidenkrantz, B.B. Jørgensen, and B.A. Lomstein. 2017. Microbial turnover times in the deep seabed studied by amino acid racemization modelling. Science Reports 7(1):5680 doi:10.1038/s41598-017-05972-z.
Burcar, B.T., L.M. Barge, D. Trail, E.B. Watson, M.J. Russell, and L.B. McGown. 2015. RNA oligomerization in laboratory aalogs of alkaline hydrothermal vent systems. Astrobiology 15(7):509-522.
Cabrol, N.A. 2018. The coevolution of life and environment on Mars: An ecosystem perspective on the robotic exploration of biosignatures. Astrobiology 18(1):1-27.
Caetano-Anollés, G., and M.J. Seufferheld. 2013. The coevolutionary roots of biochemistry and cellular organization challenge the RNA world paradigm. Journal of Molecular Microbiology and Biotechnology 23:152-177.
Cafferty, B.J., D.M. Fialho, J. Khanam, R. Krishnamurthy, and N.V. Hud. 2016a. Spontaneous formation and base pairing of plausible prebiotic nucleotides in water. Nature Communications 7:1-8.
Cafferty, B.J., C. Musetti, K. Kim, E.D. Horowitz, R. Krishnamurthy, and N.V. Hud. 2016b. Small molecule-mediated duplex formation of nucleic acids with ‘incompatible’ backbones. Chemical Communications 52:5436-5439.
Carr, M.H., and J.W. Head. 2015. Martian surface/near-surface water inventory: Sources, sinks, and changes with time. Geophysical Research Letters 42:726-732.
Chang, S., D. DesMarais, R. Mack, S.L. Miller, G.E. Strathearn, and J.W. Schopf. 1983. Prebiotic organic syntheses and the origin of life. Pp. 53-92 in Earth’s Earliest Biosphere: Its Origin and Evolution (J.W. Schopf, ed.). Princeton University Press, Princeton, NJ.
Chassefiere, E., J. Lasue, B. Langlais, and Y. Quesnel. 2016. Early Mars serpentinization-derived CH4 reservoirs, H2-induced warming and paleopressure evolution. Meteoritics and Planetary Sciences 51:2234-2245.
Chevrier, V.F., J. Hanley, and T.S. Altheide. 2009. Stability of perchlorate hydrates and their liquid solutions at the Phoenix landing site, Mars. Geophysical Research Letters 36:L1020.
Ciobanu, M.-C., G. Burgaud, A. Dufresne, A. Breuker, V. Redou, S.B. Maamar, F. Gaboyer, et al. 2014. Microorganisms persist at record depths in the subsea floor of the Canterbury Basin. The ISME Journal 8:1370-1380.
Cody, G.D., N.Z. Boctor, T.R. Filley, R.M. Hazen, J.H. Scott, A. Sharma, and H.S. Yoder Jr. 2000. Primordial carbonylated iron-sulfur compounds and the synthesis of pyruvate. Science 289:1337-1340.
Collins, G., and F. Nimmo. 2009. Chaotic terrain on Europa. Pp. 259-281 in Europa (R.T. Pappalardo, W.B. McKinnon, and K. Khurana eds.). University of Arizona Press, Tuscon, AZ.
Collins, G.C., J.W. Head, R.T. Pappalardo, and N.A. Spaun. 2000. Evaluation of models for the formation of chaotic terrain on Europa. Journal of Geophysical Research 105(E1):1709-1716.
Colwell, F.S., and S. D’Hondt. 2013. Nature and extent of the deep biosphere. Reviews in Mineralogy and Geochemistry 75:547-574.
Condie, K.C. 2016. A planet in transition: The onset of plate tectonics on Earth between 3 and 2 Ga? Geoscience Frontiers 9(1):51-60.
Condie, K.C., and A. Kröner. 2008. When did plate tectonics begin? Evidence from the geologic record. Pp. 281-294 in When Did Plate Tectonics Begin on Planet Earth? Geological Society of America Special Paper 440 (K.C. Condie and C. Pease, eds.). Geological Society of America, Boulder, CO.
Corliss, J.B. 1990. Hot springs and the origin of life. Nature 347:624.
Da Silva, L., M.C. Maurel, and D. Deamer. 2015. Salt-promoted synthesis of RNA-like molecules in simulated hydrothermal conditions. Journal of Molecular Evolution 80:86-97.
Dalai, P., P. Ustriyana, and N. Sahai. 2018. Aqueous magnesium as an environmental selection pressure in the evolution of phospholipid membranes on early Earth. Geochimica et Cosmochimica Acta 223:216-228.
Damer, B., and D. Deamer. 2015. Coupled phases and combinatorial selection in fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life 5(1):872-887.
Dhuime, B., A. Wuestefeld, and C.J. Hawkesworth. 2015. Emergence of modern continental crust about 3 billion years ago. Nature Geoscience 8(7):552-555.
Dhuime, B., C.J. Hawkesworth, P.A. Cawood, and C.D. Storey. 2012. A change in the geodynamics of continental growth 3 billion years ago. Science 335:1334-1336.
Dundas, C.M., A.M. Bramson, L. Ojha, J.J. Wray, M.T. Mellon, S. Byrne, A.S. McEwen, et al. 2018. Exposed subsurface ice sheet in the martian mid-latitudes. Science 359(6372):199-201.
Edwards, K.J., K. Becker, and F. Colwell. 2012. The deep, dark energy biosphere: Intraterrestrial life on Earth. Annual Review of Earth and Planetary Sciences 40:551-568.
Ehlmann, B.L., and C.S. Edwards. 2014. Mineralogy of the martian surface. Annual Review of Earth and Planetary Science 42:291-315.
Ehlmann, B.L., California Institute of Technology. 2018. “Mars Astrobiology: Recent Findings and Future Approaches.” Presentation to the Committee on an Astrobiology Science Strategy for the Search for Life in the Universe, January 17.
Ehlmann, B.L., F.S. Anderson, J. Andrews-Hanna, D.C. Catling, P.R. Christensen, B.A. Cohen, C.D. Dressing, et al. 2016. The sustainability of habitability on terrestrial planets: Insights, questions, and needed measurements from Mars for understanding the evolution of Earth-like worlds. Journal of Geophysical Research 121(10):1927-1961.
Ehlmann, B.L., J.F. Mustard, C.I. Fassett, S.C. Schon, J.W. Head, D.J. Des Marais, J.A. Grant, and S.L. Murchie. 2008. Clay minerals in delta deposits and organic preservation potential on Mars. Nature Geoscience 1:355-358.
Ehlmann, B.L., J.F. Mustard, S.L. Murchie, J.-P. Bibring, A. Meunier, A.A. Fraeman, and Y. Langevin. 2011. Subsurface water and clay mineral formation during the early history of Mars. Nature 479:53-60.
Eigenbrode, J., NASA Goddard Space Flight Center. 2018. “Preservation of Biotic/Abiotic Signatures Beyond Earth.” Presentation to the Committee on an Astrobiology Science Strategy for the Search for Life in the Universe, March 8.
Elkins-Tanton, L.T. 2008. Linked magma ocean solidification and atmospheric growth for Earth and Mars. Earth and Planetary Science Letters 271(1-4):181-191.
Eschenmoser, A. 2007. The search for the chemistry of life’s origin. Tetrahedron 63(52):12821-12844.
Estrada, C.F., D.A. Sverjensky, M. Pelletier, A. Razafitianamaharavo, and R.M. Hazen. 2015. Interaction between L-aspartate and the brucite [Mg(OH)2]-water interface. Geochimica et Cosmochimica Acta 155:172-186.
Estrada, C.F., I. Mamajanov, J. Hao, D.A. Sverjensky, G.D. Cody, and R.M. Hazen. 2017. Aspartate transformation at 200°C with brucite [Mg(OH)2], NH3, and H2: Implications for prebiotic molecules in hydrothermal systems. Chemical Geology 457:162-172.
Fernandez-Garcia, C., A.J. Coggins, and M.W. Powner. 2017. A chemist’s perspective on the role of phosphorous at the origins of life. Life 7(3):31.
Ferris, J.P., A.R. Hill, R. Liu, and L.E. Orgel. 1996. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 381(6577):59-61.
Feuillie, C., D.A. Sverjensky, and R.M. Hazen. 2015. Attachment of ribonucleotides on α-alumina as a function of pH, ionic strength, and surface loading. Langmuir 31(3):240-248.
Fischer, E., G.M. Martínez, H.M. Elliott, and N.O. Rennó. 2014. Experimental evidence for the formation of liquid saline water on Mars. Geophysical Research Letters 41(13):4456-4462.
Fischer, E., G.M. Martínez, and N.O. Rennó. 2016. Formation and persistence of brine on Mars: Experimental simulations throughout the diurnal cycle at the Phoenix landing site. Astrobiology 16(12):937-948.
Fornaro, T., J.R. Brucato, E. Pace, M.C. Guidi, S. Branciamore, and A. Pucci. 2013. Infrared spectral investigations of UV irradiated nucleobases adsorbed on mineral surfaces. Icarus 226:1068-1085.
Forsythe, J.G., S.-S. Yu, I. Mamajanov, M.A. Grover, R. Krishnamurthy, F.M. Fernandez, and N.V. Hud. 2015. Ester-mediated amide bond formation driven by wet-dry cycles: A possible path to polypeptides on the prebiotic Earth. Angewandte Chemie—International Edition 54(34):9871-9875.
Fuchida, S., and H. Masuda. 2015. Effect of temperature on the dissolution and thermal alteration of combined amino acids fixed in natural sediment under simulated hydrothermal conditions. Organic Geochemistry 87:1-8.
Fuchida, S., H. Naraoka, and H. Masuda. 2017. Formation of diastereoisomeric piperazine-2, 5-dione from DL-alanine in the presence of olivine and water. Origins of Life and Evolution of Biospheres 47(1):83-92.
Georgelin, T., M. Akouche, M. Jaber, Y., Sakhano, L. Matheron, F. Fournier, C. Methivier, G. Martra, and J.-F. Lamert. 2017. Iron(III) oxide nanoparticles as catalysts for the formation of linear glycine peptides. European Journal of Inorganic Chemistry 1:198-211.
Gilbert, W. 1986. Origin of life: The RNA world. Nature 319(6055):618.
Glavin, D.P., C. Freissinet, K.E. Miller, J.L. Eigenbrode, A.E. Brunner, A. Buch, B. Sutter, et al. 2013. Evidence for perchlorates and the origin of chlorinated hydrocarbons detected by SAM at the Rocknest aeolian deposit in Gale Crater. Journal of Geophysical Research: Planets 118(10):1955-1973.
Glein, C.R., J.A. Baross, and J.H. Waite. 2015. The pH of Enceladus’ ocean. Geochimica et Cosmochimica Acta 162:202-219.
Goldreich, P.M., and J.L. Mitchell. 2010. Elastic ice shells of synchronous moons: Implications for cracks on Europa and non-synchronous rotation of Titan. Icarus 209(2):631-638.
Goudge, T.A., J.F. Mustard, J.W. Head, C.I. Fassett, and S.M. Wiseman. 2015. Assessing the mineralogy of the watershed and fan deposits of the Jezero crater paleolake system, Mars. Journal of Geophysical Research 120:775-808.
Gough, R.V., V.F. Chevrier, K.J. Baustian, M.E. Wise, and M.A. Toblert. 2011. Laboratory studies of perchlorate phase transitions: Support for metastable aqueous perchlorate solutions on Mars. Earth and Planetary Science Letters 312:371-377.
Greeley, R., J.E. Klemaszewski, R. Wagner, and the Galileo Imaging Team. 2000. Galileo views of the geology of Callisto. Planetary and Space Science 48(9):829-853.
Green, W.J., and W.B. Lyons. 2009. The Saline Lakes of the McMurdo Dry Valleys, Antarctica. Aquatic Geochemistry 15(1-2):321-348.
Gregoire, B., V. Erastova, D.L. Geatches, S.J. Clark, H.C. Greenwell, and D.G. Fraser. 2016. Insights into the behavior of biomolecules on the early Earth: The concentration of aspartate by layered double hydroxide minerals. Geochimica et Cosmochimica Acta 176:239-258.
Griesser, H., P. Tremmel, E. Kervio, C. Pfeffer, U.E. Steiner, and C. Richert. 2017. Ribonucleotides and RNA promote peptide chain growth. Angewandte Chemie—International Edition 56(5):1219-1223.
Grotzinger, J.P. D.Y. Sumner, L.C. Kah, K. Stack, S. Gupta, L. Edgar, D. Rubin, et al. 2014. A habitable fluvio-lacustrine environment at Yellowknife Bay, Gale Crater, Mars. Science 343(6169).
Grotzinger, J.P., R.E. Arvidson, J.F. Bell, W. Calvin, B.C. Clark, D.A. Fike, M. Golombek, et al. 2005. Stratigraphy and sedimentology of a dry to wet eolian depositional system, Burns formation, Meridiani Planum, Mars. Earth and Planetary Science Letters 240:11-72.
Grotzinger, J.P., S. Gupta, M.C. Malin, D.M. Rubin, J. Scheiber, K.Siebach, D.Y. Summer, et al. 2015. Deposition, exhumation, and paleoclimate of an ancient lake deposit, Gale crater, Mars. Science 350(6257):acc7575.
Guo, C., J.S. Jordan, J.L. Yarger, and G.P. Holland. 2017. Highly efficient fumed silica nanoparticles for peptide bond formation: Converting alanine to alanine anhydride. ACS Applied Materials and Interfaces 9(20):17653-17661.
Haberle, R.M., C.P. McKay, J. Schaeffer, N.A. Cabrol, E.A. Grin, A.P. Zent, and R. Quinn. 2001. On the possibility of liquid water on present-day Mars. Journal of Geophysical Research 106(E10):23317-23326.
Hand, K.P., A.E. Murray, J.B. Garvin, and the Europa Lander Mission Concept Team. 2016. NASA Europa Lander Study 2016 Report: Europa Lander Mission Pre-Phase A. JPL D-97667, Task Order NNN16D011T. https://europa.nasa.gov/resources/58/europa-lander-study-2016-report/.
Harrison, T.M. 2009. The Hadean crust: Evidence from 4 Ga zircons. Annual Review of Earth and Planetary Sciences 37(1):479-505.
Harrison, T.M., A.K. Schmitt, M.T. McCulloch, and O.M. Lovera. 2008. Early (≥4.5 Ga) formation of terrestrial crust: Lu–Hf, δ18O, and Ti thermometry results for Hadean zircons. Earth and Planetary Science Letters 268(3-4):476-486.
Hashizume, H. 2015. Adsorption of nucleic acid bases, ribose, and phosphate by some clay minterals. Life 5(1):637-650.
Hazen, R.M., and D.A. Sverjensky. 2010. Mineral surfaces, geochemical complexities, and the origins of life. Cold Spring Harbor Perspectives in Biology 2(5):a002162.
Hazen, R.M., N. Boctor, J.A. Brandes, G.D. Cody, R.J. Hemley, A. Sharma, and H.S. Yoder Jr. 2002. High pressure and the origin of life. Journal of Physics: Condensed Matter 14:11489-11494.
Hecht, M.H., S.P. Kounaves, R.C. Quinn, S.J. West, S.M.M. Young, D.W. Ming, D.C. Catling, et al. 2009. Detection of perchlorate and the soluble chemistry of martian soil at the Phoenix landing site. Science 325(5936):64-67.
Herschy, B., A. Whicher, E. Camprubi, C. Watson, L. Dartnell, J. Ward, J.G.R. Evans, and N. Lane. 2014. An origin of life reactor to simulate alkaline hydrothermal vents. Journal of Molecular Evolution 79(5-6):213-227.
Herschy, B., S.J. Chang, R. Blake, A Lepland, H. Abbott-Lyon, J. Sampson, Z. Atlas, T.P. Kee, and M.A. Pasek. 2018. Archean phosphorus liberation induced by iron redox geochemistry. Nature Communications 9:1346.
Hitchcock, D. R., and J.E. Lovelock. 1967. Life detection by atmospheric analysis. Icarus 7(1-3):149-159.
Hoehler, T.M., and R.B. Jorgenson. 2013. Microbial life under extreme energy limitation. Nature Reviews Microbiology 11:83-94.
Holland, G., B. Sherwood Lollar, L. Li, G. Lacrampe-Couloume, G.F. Slater, and C.J. Ballentine. 2013. Deep fracture fluids isolated in the crust since the Precambrian era. Nature 497:357-360.
Holm, N. 2014. Glasses as sources of condensed phosphates on the early Earth. Geochemical Transactions 15:8.
Holt, J.W., A. Safaeinili, J.J. Plaut, J.W. Head, R.J. Philips, R. Seu, S.D. Kempf, et al. 2008. Radar sounding evidence for buried glaciers in the southern mid-latitudes of Mars. Science 322:1235-1238.
Hsu, H.-W., F. Postberg, Y. Sekine, T. Shibuya, S. Kempf, M. Horanyi, A. Juhasz, et al. 2015. Ongoing hydrothermal activities within Enceladus. Nature 519:207-210.
Husmann, H., F. Sohl, and T. Spohn. 2006. Subsurface oceans and deep interiors of medium-sized outer planet satellites and large trans-neptunian objects. Icarus 185(1):258-273.
Hynek, B.M., T.M. McCollom, and K.L. Rogers. 2011. Cerro Negro Volcano, Nicaragua: An assessment of geological and potential biological systems on early Mars. Pp. 279-285 in GSA Special Papers 483: Analogs for Planetary Exploration. (W.B. Garry and J.E. Bleacher, eds.). Geological Society of America, Boulder, CO.
Iess, L., D.J. Stevenson, M. Parisi, D. Hemingway, R.A. Jacobson, J.I. Lunine, F. Nimmo, et al. 2014. The gravity field and interior structure of Enceladus. Science 344:78-80.
Iess, L., R.A. Jacobson, M. Ducci, D.J. Stevenson, J.I. Lunine, J.W. Armstrong, S.W. Asmar, et al. 2012. The tides of Titan. Science 377(6093):457-459.
Iqubal, Md.A., R. Sharma, and Kamaluddin. 2016. Surface interaction of ribonucleic acid constituents with spinel ferrite nanoparticles: A prebiotic chemistry experiment. RSC Advances 6:68574-68583.
Iqubal, Md.A., R. Sharma, S. Jheeta, and Kamaluddin, 2017. Thermal condensation of glycine and alanine on metal ferrite surface: Primitive peptide bond formation scenario. Life 7(2):15.
Islam, S., and M.W. Powner. 2017. Prebiotic system chemistry: Complexity overcoming clutter. Chem 2(4):470-501.
Jauker, M., H. Greisser, and C. Richert. 2015. Spontaneous formation of RNA strands, peptidyl RNA, and cofactors. Angewandte Chemie—International Edition 54(48):14564-14569.
Jørgensen, B.B., and I.P.G. Marshall. 2016. Slow microbial life in the seabed. Annual Review of Marine Science 8:311-332.
Joye, S.B., and V.A. Samarkin. 2009. Metabolic variability in seafloor brines revealed by carbon and sulphur dynamics. Nature Geoscience 2(5):349-354.
Junge, K., C. Krembs, J. Deming, A. Stierle, and H. Eicken. 2001. A microscopic approach to investigate bacteria under in situ conditions in sea-ice samples. Pp. 304-310 in Annals of Glaciology Volume 33: Selected Papers of the International Symposium on Sea ice and Its Interactions with Ocean, Atmosphere, and Biosphere (M.O. Jeffries and H. Eicken, eds.). International Glaciological Society, Fairbanks, AK.
Kallmeyer, J., R. Pockalny, R.R. Adhikari, D.C. Smith, and S. D’Hondt. 2012. Global distribution of microbial abundance and biomass in subseafloor sediment. Proceedings of the National Academy of Sciences U.S.A. 109(40):16213-16216.
Kasting, J.F. 2014. Atmospheric composition of Hadean-early Archean Earth: The importance of CO. Pp. 19-28 in Earth’s Early Atmosphere and Surface Environment (G.H. Shaw, ed.). Geological Society of America, Boulder, CO.
Kattenhorn, S.A., and L.M. Procter. 2014. Evidence for subduction in the ice shell of Europa. Nature Geoscience 7:762-767.
Keane, J.T., I. Matsuyama, S. Kamata, and J.K. Steckloff. 2016. Reorientation and faulting of Pluto due to volatile loading within Sputnik Planitia. Nature 540:90-93.
Keil, L., M. Hartmann, S. Lanzmich, and D. Braun. 2016. Probing of molecular replication and accumulation in shallow heat gradients through numerical simulations. Physical Chemistry Chemical Physics 18:20153-20159.
Khurana, K.K., M.G. Kivelson, D.J. Stevenson, G. Schubert, C.T. Russell, R.J. Walker, and C. Polanskey. 1998. Induced magnetic fields as evidence for subsurface oceans in Europa and Callisto. Nature 395(6704):777-780.
Kietavainen, R., L. Ahonen, P. Niinikoski, H. Nykanen, and I.T. Kukkonen. 2017. Abiotic and biotic controls on methane formation down to 2.5 km within the Precambrian Fennoscandian Shield. Geochimica et Cosmochimica Acta 202:124-145.
Kim, J.D., N. Yee, V. Nanda, and P.G. Falkowski. 2013. Anoxic photochemical oxidation of siderite generates molecular hydrogen and iron oxides. Proceedings of the National Academy of Sciences U.S.A. 110:10073-10077.
Kitadai, N. 2015. Energetics of amino acid synthesis in alkaline hydrothermal environments. Origins of Life and Evolution of Biospheres 45(4):377-409.
Kitadai, N., H. Oonishi, K. Umemoto, T. Usui, K. Fukushi, and S. Nakashima. 2017. Glycine polymerization on oxide minerals. Origins of Life and Evolution of Biospheres 47(2):123-143.
Kivelson, M.G., K.K. Khurana, C.T. Russell, M. Volwerk, R.J. Walker, and C. Zimmer. 2000. Galileo magnetometer measurements: A stronger case for a subsurface ocean at Europa. Science 289(5483):1340-1343.
Kok, J.F., and N.O. Rennó. 2009. Electrification of wind-blown sand on Mars and its implications for atmospheric chemistry. Geophysical Research Letters 36(5):L05202.
Korenaga, J. 2013. Initiation and evolution of plate tectonics on Earth: Theories and observations. Annual Review of Earth and Planetary Sciences 41(1):117-151.
Kreysing, M., L. Keil, S. Lanzmich, and D. Braun. 2015. Heat flux across an open pore enables the continuous replication and selection of oligonucleotides towards increasing length. Nature Chemistry 7:203-208.
Krissansen-Totton, J., S. Olson, and D.C. Catling. 2018. Disequilibrium biosignatures over Earth history and implications for detecting exoplanet life. Science Advances 4(1):eaao5747.
Kusube, M., T.S. Kyaw, K. Tanikawa, R.A. Chastain, K.M. Hardy, J. Cameron, and D.H. Bartlett. 2017. Colwellia marinimaniae sp nov., a hyperpiezophilic species isolated from an amphipod within the Challenger Deep, Mariana Trench. International Journal of Systematic and Evolutionary Microbiology 67:824-831.
La Cruz, N., D. Qasim, H. Abbott-Lyon, C. Pirim, A.D. McKee, T. Orlando, M. Gull, D. Lindsay, and M.A. Pasek. 2016. The evolution of the surface of the mineral schreibersite in prebiotic chemistry. Physical Chemistry Chemical Physics 18:20160-20167.
Laurent, O., H. Martin, J.F. Moyen, and R. Doucelance. 2014. The diversity and evolution of late-archean granitoids: Evidence for the onset of “modern-style” plate tectonics between 3.0 and 2.5 Ga. Lithos 205:208-235.
Lazar, C., G.D. Cody, and J.M. Davis. 2015. A kinetic pressure effect on the experimental abiotic reduction of aqueous CO2 to methane from 1 to 3.5 kbar at 300°C. Geochimica et Cosmochimica Acta 151:34-48.
Leveille, R.J., and S. Datta. 2010. Lava tubes and basaltic caves as astrobiological targets on Earth and Mars: A review. Planetary and Space Science 58(4):592-598.
Li, L., B.A. Wing, T.H. Bui, J.M. McDermott, G.F. Slater, S. Wei, G. Lacrampe-Couloume, and B. Sherwood Lollar. 2016. Sulfur mass-independent fractionation in subsurface fracture waters indicates a long-standing sulfur cycle in Precambrian rocks. Nature Communications 7:13252.
Lin, L.-H., J. Hall, J. Lippmann-Pipke, J.A. Ward, B. Sherwood Lollar, M. DeFlaun, R. Rothmel, et al. 2005a. Radiolytic H2 in continental crust: Nuclear power for deep subsurface microbial communities. Geochemisty, Geophysics, Geosystems 6:Q07003.
Lin, L.-H., G.F. Slater, B. Sherwood Lollar, G. Lacrampe-Couloume, and T.C. Onstott. 2005b. The yield and isotopic composition of radiolytic H2, a potential energy source for the deep subsurface biosphere. Geochimica Cosmochimica Acta 69(4):893-903.
Lin, L.-H., P.-L. Wang, D. Rumble, J. Lippmann-Pipke, E. Boice, L.M. Pratt, B. Sherwood Lollar, et al. 2006. Long-term sustainability of a high-energy, low-diversity crustal biome. Science 314(5798):479-482.
Lippmann-Pipke, J., B. Sherwood Lollar, S. Niedermann, N.A. Stroncik, R. Naumann, E. van Heerden, and T.C. Onstott. 2011. Neon identifies two billion year old fluid component in the Witwatersrand Basin. Chemical Geology 283(3-4):287-296.
Magnabosco, C., P.H.A. Timmers, M.C.Y. Lau, G. Borgonie, B. Linage-Alvarez, O. Kuloyo, R. Alleva, et al. 2018. Fluctuations in populations of subsurface methane oxidizers in coordination with changes in electron acceptor availability. FEMS in press, doi:10.1093/femsec/fiy089.
Martin, W., J. Baross, D. Kelley, and M.J. Russell. 2008. Hydrothermal vents and the origin of life. Nature Reviews Microbiology 6:805-814.
Martin-Torres, F.J., M.-P. Zorzano, P. Valentin-Serrano, A.-H. Harri, M. Genzer, O. Kemppinen, E.G. Rivera-Valentin, et al. 2015. Transient liquid water and water activity at Gale crater on Mars. Nature Geoscience 8:357-361.
Maurer, S. 2017. The impacts of salts on single chain amphiphile membranes and implications for the location of the origin of life. Life 7(4):44.
Maurer, S., and G. Nguyen. 2016. Prebiotic vesicle formation and the necessity of salts. Origins of Life and Evolution of Biospheres 46(2-3):215-222.
Mayhew, L.E., E.T. Ellison, T.M. McCollom, T.P. Rrainor, and A.S. Templeton. 2013. Hydrogen generation from low-temperature water-rock reactions. Nature Geosciences 6:478-484.
McDermott, J.M., J.S. Seewald, C.R. German, and S.P. Sylva. 2015. Pathways for abiotic organic synthesis at submarine hydrothermal fields. Proceedings of the National Academy of Sciences U.S.A. 112:7668-7672.
McEwen, A.S., C.M. Dundas, S.A. Mattson, A.D. Toigo, L. Ojha, J.J. Wray, M. Chojnacki, S. Byrne, S.L. Murchie, and N. Thomas. 2014. Recurring slope lineae in equatorial regions of Mars. Nature Geoscience 7:53-58.
McEwen, A.S., L. Ojha, C.M. Dundas, S.S. Mattson, S. Byrne, J.J. Wray, S.C. Cull, et al. 2011. Seasonal flows on warm martian slopes. Science 333(6043):740-743.
McGlynn, S.E., I. Kanik, and M.J. Russell. 2012. Peptide and RNA contributions to iron-sulphur chemical gardens as life’s first inorganic compartments, catalytsts, capacitors, and condensers. Philosophical Transactions of the Royal Society A 370:3007-3022.
McKay, C.P. 2010. An origin of life on Mars. Cold Spring Harbor Perspectives in Biology 2010(2):a003509, doi:10.1101/cshperspect.a003509.
Meadows, V.S. 2017. Reflections on O2 as a biosignature in exoplanetary atmospheres. Astrobiology 17:1022-1052.
Meyer, J., and J. Wisdom. 2007. Tidal heating in Enceladus. Icarus 188(2):535-539.
Meyer, J., and J. Wisdom. 2008. Tidal evolution of Mimas, Enceladus, and Dione. Icarus 193(1):213-223.
Michalski, J.R., J. Cuadros, P.B. Niles, J. Parnell, A.D. Rogers, and S.P. Wright. 2013. Groundwater activity on Mars and implications for a deep biosphere. Nature Geoscience 6:133-138.
Mikucki, J.A., A. Pearson, D.T. Johnston, A.V. Turchyn, J. Farquhar, D.P. Schrag, A.D. Anbar, J.C. Priscu, and P.A. Lee. 2009. A contemporary microbially maintained subglacial ferrous ocean. Science 324(5925):397-400.
Milshteyn, D., B. Dmaer, J. Havig, D. Deamer. 2018. Amphiphilic compounds assemble into membranous vesicles in hydrothermal hot spring water but not in seawater. Life 8(2):11.
Ming, D.W., P.D. Archer, D.P. Glavin, J.L. Eigenbroide, H.B. Franz, B. Sutter, A.E. Brunner, et al. 2014. Volatile and organic compositions of sedimentary rocks in Yellowknife Bay, Gale Crater, Mars. Science 343(6169).
Mojzsis, S.J., T.M. Harrison, and R.T. Pidgeon. 2001. Oxygen-isotope evidence from ancient zircons for liquid water at the Earth’s surface 4,300 Myr ago. Nature 409(6817):178-181.
Möller, F.M., F. Kriegel, M. Krieβ, V. Sojo, and D. Braun. 2017. Steep pH gradients and directed colloid transport in a microfluidic alkaline hydrothermal pore. Angewandte Chemie—International Edition 56(9):2340-2344.
NASA (National Aeronautics and Space Administration). 2015. NASA Astrobiology Strategy 2015. NASA Astrobiology Program. Washington, DC.
NASEM (National Academies of Sciences, Engineering, and Medicine). 2017. Searching for Life Across Space and Time: Proceedings of a Workshop. The National Academies Press, Washington, DC.
Nebel-Jacobsen, Y., O. Nebel, M. Wille, and P.A. Cawood. 2018. A non-zircon Hf isotope record in Archean black shales from the Pilbara Craton confirms changing crustal dynamics ca. 3 Ga ago. Scientific Reports 8(1), doi:10.1038/s41598-018-19397-9.
Niether, D., D. Afanasenkau, J.K.G. Dhont, and S. Wiegand. 2016. Accumulation of formamide in hydrothermal pores to form prebiotic nucleobases. Proceedings of the National Academy of Sciences 113(16):4274-4277.
Nimmo, F., and R.T. Pappalardo. 2006. Diapir-induced reorientation of Saturn’s moon Enceladus. Nature 441:614-616.
Nitta, S., Y. Furukawa, and T. Kakegawa. 2016. Effects of silicate, phosphate, and calcium on the stability of aldopentoses. Origins of Life and Evolution of Biospheres 46(2-3):189-202.
NRC (National Research Council). 2007a. An Astrobiology Strategy for the Exploration of Mars. The National Academies Press, Washington, DC.
NRC. 2007b. The Limits of Organic Life in Planetary Systems. The National Academies Press, Washington, DC.
Nuevo, M., S.N. Milam, and S.A. Sandford. 2012. Nucleobases and prebiotic molecules in organic residues produced from the ultraviolet photo-irradiation of pyrimidine in NH3 and H2+NH3 ices. Astrobiology 12(4):295-314.
O’Neill, C.N., and F. Nimmo. 2010. The role of episodic overturn in generating the surface geology and heat flow on Enceladus. Nature Geoscience 3:88-91.
Ojha, L., M.B. Wilhelm, S.L. Murchie, A.S. McEwen, J.J. Wray, J. Hanley, M. Masse, and M. Chojnacki. 2015. Spectral evidence for hydrated salts in recurring slope lineae on Mars. Nature Geoscience 8:829-832.
Onstott, T.C. 2016. Deep Life: The Hunt for the Hidden Biology of Earth, Mars, and Beyond. Princeton University Press, Princteon, NJ. January 9.
Oró, J. 1960. Synthesis of adenine from ammonium cyanide. Biochemical and Biophysical Research Communications 2(6):407-412.
Pandey, P., C.K. Pant, K. Gururani, P. Arora, N. Pandey, P. Bhatt, Y. Sharma, J.S. Negi, and M.S. Mehata. 2015. Affinity of smectite and divalent metal ions (Mg2+, Ca2+, Cu2+) with L-leucine: An experimental and theoretical approach relevant to astrobiology. Origins of Life and Evolution of Biospheres 45(4):411-426.
Pappalardo, R.T., and A.C. Barr. 2004. The origin of domes on Europa: The role of thermally induced compositional diapirism. Geophysical Research Letters 31(L01701), doi:10.1029/2003GL019202.
Pappalardo, R.T., J.W. Head, R. Greeley, R.J. Sullivan, C. Pilcher, G. Schubert, W.B. Moore, et al. 1998. Geological evidence for solid-state convection in Europa’s ice shell. Nature 391:365-368.
Pasek, M., B. Herschy, and T.P. Kee. 2015. Phosphorous: A case for mineral-organic reactions in prebiotic chemistry. Origins of Life and Evolution of Biospheres 45:207-218.
Pasek, M.A. 2017. Schreibersite on the early Earth: Scenarios for prebiotic phosphorylation. Geoscience Frontiers 8:329-335.
Patel, B.H., C. Percivalle, D.J. Ritson, C.D. Duffy, and J.D. Sutherland. 2015. Common origins of RNA, protein, and lipid precursors in a cyanosulfidic protometabolism. Nature Chemistry 7(4):301-307.
Pathare, A.V., W.C. Fedman, T.H. Prettyman, and S. Maurice. 2018. Driven by excess? Climatic implications of new global mapping of near-surface water-equivalent hydrogen on Mars. Icarus 301:97-116.
Pearce, B.K.D., R.E. Pudritz, D.A. Semenov, and T.K. Henning. 2017. Origin of the RNA world: The fate of nucleobases in warm little ponds. Proceedings of the National Academy of Sciences U.S.A. 114:11327-11332.
Pedreira-Segade, U., C. Feuillie, M. Pelletier, L.J. Michot, and I. Daniel. 2016. Adsorption of nucleotides onto ferromagnesian phyllosilicates: Significance for the origin of life. Geochimica et Cosmochimica Acta 176:81-95.
Pedreira-Segade, U., L.J. Michot, and I. Daniel. 2018. Effects of salinity on the adsorption of nucleotides onto phyllosilicates. Physical Chemistry Chemical Physics 20:1938-1952.
Phillips, R.J., B.J. Davis, K.L. Tanaka, S. Byrne, M.T. Mellon, N.E. Putzig, R.M. Haberle, et al. 2011. Massive CO2 ice deposits sequestered in the south polar layered deposits of Mars. Science 332(6031):838-841.
Porco, C.C., P. Helfenstein, P.C. Thomas, A.P. Ingersoll, J. Wisdom, R. West, G. Neukum, T. Denk, R. Wagner. 2006. Cassini observes the active south pole of Enceladus. Science 311:1393-1401.
Postberg, F., J. Schmidt, J. Hillier, S. Kempf, and R. Srama. 2011. A salt-water reservoir as the source of a compositionally stratified plume on Enceladus. Nature 474:620-622.
Powner, M.W., B. Gerland, and J.D. Sutherland. 2009. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 459:239-242.
Priscu, J.C., and B.C. Christner. 2004. Earth’s icy biosphere. Pp. 130-145 in Microbial Diversity and Bioprospecting (A. Bull, ed.). American Society for Microbiology Press, Washington, DC.
Ramirez, R.M. 2018. A more comprehensive habitable zone for finding life on other planets. Geosciences 8:280-328.
Rennó, N.O., B.J. Bos, D. Catling, B.C. Clark, L. Drube, D. Fisher, W. Goetz, et al. 2009. Possible physical and thermodynamical evidence for liquid water at the Phoenix landing site. Journal of Geophysical Research: Planets 114:E00E03.
Rettberg, P., A.M. Anesio, V.R. Baker, J.A. Baross, S.L. Cady, E. Detsis, C.M. Foreman, et al. 2016. Planetary protection and Mars Special Regions—A suggestion for updating the definition. Astrobiology 16(2):119-125.
Ritson, D.J., C. Battilocchio, S. V. Ley, and J. D. Sutherland. 2018. Mimicking the surface and prebiotic chemistry of early Earth using flow chemistry. Nature Communications 9:1821.
Rogers, K.L., T.M. McCollom, and B.M. Hynek. 2014. “Models of Metabolic Community Structure in Martian Habitable Environments: Contraints from a Terrestrial Analog Acid-Sulfate Fumarole Environment, Cerro Negro Volcano, Nicaragua.” Abstract P32A-02 presented at 2014 Fall American Geophysical Union Meeting. American Geophysical Union, Washington, DC.
Ross, D., and D. Deamer. 2016. Dry/wet cycling and the thermodynamics and kinetics of prebiotic polymer synthesis. Life 6(3):28.
Roth, L., J. Saur, K.D. Retherford, D.F. Strobel, P.D. Feldman, M.A. McGrath, and F. Nimmo. 2014a. Transient water vapor at Europa’s south pole. Science 343:171-174.
Roth, L., K.D. Retherford, J. Saur, D.F. Strobel, P. Feldman, M.A. McGrath, and F. Nimmo. 2014b. Orbital apocenter is not a sufficient condition for HST/STIS detection of Europa’s water vapor aurora. Proceedings of the National Academy of Sciences U.S.A. 111(48): E5123-E5132.
ROW (Roadmaps to Ocean Worlds). 2017. Goals, Objectives and Investigations for Ocean Worlds. Outer Planet Assessment Group. June. https://www.lpi.usra.edu/opag/ROW/Priorities-Missions-Scenarios-0218.pdf.
Saper, L., and J.F. Mustard. 2013. Extensive linear ridge networks in Nili Fossae and Nilosyrtis, Mars: Implications for fluid flow in the ancient crust. Geophysical Research Letters 40:245-249.
Schmidt, B., D. Blankenship, G. Patterson, and P. Schenk. 2011. Active formation of chaos terrain over shallow subsurface water on Europa. Nature 479(7374):502-505.
Schrader, M.E. 2017. Polypeptide formation on polar mineral surfaces: Possibility of complete chirality. International Journal of Astrobiology 16(1):10-13.
Schwaminger, S.P., P.F. Garcia, G.K. Merck, F.A. Bodensteiner, S. Heissler, S. Gunther, and S. Berensmeier. 2015. Nature of interactions of amino acids with bare magnetite nanoparticles. Journal of Physical Chemistry C 119(40):23032-23041.
Sharma, A., J.H. Scott, G.D. Cody, M.L. Fogel, R.M. Hazen, R.J. Hemley, W.T. Huntress. 2002. Microbial activity at gigapascal pressures. Science 295:1514-1516.
Sheik, C.S., B.K. Reese, K.I. Twing, J.B. Sylvan, S.L. Grim, M.O. Schrenk, M.L. Sogin, and F.S. Colwell. 2018. Identification and removal of contaminant sequences from ribosomal gene databases: Lessons from the census of deep life. Frontiers in Microbiology doi:10.3389/fmicb.2018.00840.
Sherwood Lollar, B., K. Voglesonger, L.-H. Lin, G. Lacrampe-Couloume, J. Telling, T.A. Abrajano, T.C. Onstott, and L.M Pratt. 2007. Hydrogeologic controls on episodic H2 release from Precambrian fractured rocks—energy for deep subsurface life on Earth and Mars. Astrobiology 7(6):971-986.
Sherwood Lollar, B., T.C. Onstott, G. Lacrampe-Couloume, and C.J. Ballentine. 2014. The contribution of the Precambrian continental lithosphere to global H2 production. Nature 516:379-382.
Showman, A.P., and L. Han. 2004. Numerical simulations of convection in Europa’s ice shell: Implications for surface features. Journal of Geophysical Research: Planets 109(E01010).
Simoncini, E., N. Virgo, and A. Kleidon. 2013. Quantifying drivers of chemical disequilibrium: Theory and applications to methane in the Earth’s atmosphere. Earth System Dynamics 4:317-331.
Skok, J.R., J.F. Mustard, B.L. Ehlmann, R.E. Milliken, and S.L. Murchie. 2010. Silica deposits in the Nili Patera caldera on the Syrtis Major volcanic complex on Mars. Nature Geoscience 3: 838-841.
Sleep, N.H., and D.K. Bird. 2007. Niches of the pre-photosynthetic biosphere and geologic preservation of Earth’s earliest ecology. Geobiology 5(2):101-117.
Sleep, N.H., and M. Zoback. 2007. Did earthquakes keep the early crust habitable? Astrobiology 7(6):1023-1032.
Sleep, N.H., K. Zahnle, and P.S. Neuhoff. 2001. Initiation of clement surface conditions on the earliest Earth. Proceedings of the National Academy of Sciences U.S.A. 98(7):3666-3672.
Sleep, N.H., K.J. Zahnle, and R.E. Lupu. 2014. Terrestrial aftermath of the Moon-forming impact. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 372(2024).
Smith, E., and H.J. Morowitz. 2016. The Origin and Nature of Life on Earth: The Emergence of the Fourth Geosphere. Cambridge University Press, Cambridge, U.K. March 31.
Soderlund, K.M., B.E. Schmidt, J. Wicht, and D.D. Blankenship. 2014. Ocean-driven heating of Europa’s icy shell at low latitudes. Nature Geoscience 7:16-19.
Sotin, C., and G. Tobie. 2004. Internal structure and dynamics of the large icy satellites. Comptes Rendus Physique 5(7):769-780.
Sparks, W.B., B.E. Schmidt, M.A. McGrath, K.P. Hand, J.R. Spencer, M. Cracraft, and S.E. Deustua. 2017. Active cryovolcanism on Europa? The Astrophysical Journal 839:L18.
Sparks, W.B., K.P. Hand, M.A. McGrath, E. Bergeron, M. Cracraft, and S.E. Deustua. 2016. Probing for evidence of plumes on Europa with HST/STIS. The Astrophysical Journal 829(2):121.
Spencer, J.R., J.C. Pearl, F.M. Lasar, A. Mamoutkine, P. Romani, B.J. Buratti, A.R. Hendrix, L.J. Spilker, and R.M.C. Lopes. 2006. Cassini encounters Enceladus: Background and the discovery of a south polar hot spot. Science 311(5766):1401-1405.
Sproul, G. 2015. Abiogenic synthesis of lipoamino acids and lipopeptides and their prebiotic significance. Origins of Life and Evolution of Biospheres 45(4):427-437.
Squyres, S.W., J.P. Grotzinger, R.E. Arvidson, J.F. Bell, W. Calvin, P.R. Christensen, B.C. Clark, et al. 2004. In situ evidence for an ancient aqueous environment at Meridiani Planum, Mars. Science 306(5702):1709-1714.
Steele, A., F.M. McCubbin, and M.D. Fries. 2016. The provenance, formation, and implications of reduced carbon phases in martian meteorites. Meteoritics and Planetary Science 51(11):2203-2225.
Stillman, D.E., and R.E. Grimm. 2018. Two pulses of seasonal activity in martian southern mid-latitude recurring slope lineae (RSL). Icarus 302(1):126-133.
Sutherland, J.D. 2016. The origin of life—Out of the blue. Angewandte Chemie—International Edition 55:104-121.
Suzuki, S., S. Ishii, T. Hoshino, A. Rietze, A. Tenney, P.L. Morrill, F. Inagaki, J.G. Kuenen, and K.H. Nealson. 2017. Unusual metabolic diversity of hyperalkaliphilic microbial communities associated with subterranean serpentinization at The Cedars. The ISME Journal 11:2584-2598.
Szostak, J.W. 2017. The narrow road to the deep past: In search of the chemistry of the origin of life. Angewandte Chemie International Edition 56(37):11037-11043.
Takai, K., K. Nakamura, T. Toki, U. Tsunogai, M. Miyazaki, J. Miyazaki, H. Hirayama, et al. 2008. Cell proliferation at 122°C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proceedings of the National Academy of Sciences U.S.A. 105:10949-10954.
Tang, M., K. Chen, and R.L. Rudnick. 2016. Archean upper crust transition from mafic to felsic marks the onset of plate tectonics. Science 351(6271):372-375.
Thomas, D.N., and G.S. Dieckmann. 2002. Antarctic sea ice—A habitat for extremophiles. Science 295(5555):641-644.
Thomas, P.C., J.A. Burns, P. Helfenstein, S. Squyres, J. Veverka, C. Porco, E.P. Turtle, et al. 2007. Shapes of the saturnian icy satellites and their significance. Icarus 190(2):573-584.
Vago, J., European Space Agency ExoMars Program. 2018. “ExoMars: Search for Life on Mars.” Presentation to the Committee on an Astrobiology Science Strategy for the Search for Life in the Universe, March 7.
Vance, S., and J. Goodman. 2009. Oceanography of an ice-covered moon. Pp. 459-484 in Europa (R.T. Pappalardo, W.B. McKinnon, and K. Khurana, eds). University of Arizona Press, Tuscon, AZ.
Vance, S.D., K.P. Hand, and R.T. Pappalardo. 2016. Geophysical controls of chemical disequilibria in Europa. Geophysical Research Letters 43(10):4871-4879.
Vanlint, D., R. Mitchell, E. Bailey, F. Meersman, P.F. McMillan, C.W. Michiels, and A. Aertsen. 2011. Rapid acquisition of gigapascal-high-pressure resistance by Escherichia coli. mBio 2(1):e00130-10.
Varma, S.J., K.B. Muchowska, P. Chatelain, J. Moran. 2018. Native iron reduces CO2 to intermediates and end-products of the acetyl-CoA pathway. Nature Ecology and Evolutions doi:10.1038/s41559-018-0542-2.
Villafane-Barajas, S.A., J.P.T. Bau, M. Colin-Garcia, A. Negron-Mendoza, A. Heredia-Barbero, T. Pi-Puig, and D.A.M. Zaia. 2018. Salinity effects on the adsorption of nucleic acid compounds on Na– montmorillonite: A prebiotic chemistry experiment. Origins of Life and Evolution of Biospheres doi:10.1007/s11084-018-9554-9.
Wachtershauser, G. 1990. Evolution of the first metabolic cycles. Proceedings of the National Academy of Sciences U.S.A. 87(1):200-204.
Waite, J.H., C.R. Glein, R.S. Perryman, B.D. Teolis, B.A. Magee, G. Miller, J. Grimes, et al. 2017. Cassini finds molecular hydrogen in the Enceladus plume: Evidence for hydrothermal processes. Science 356:155-159.
Waite, J.H., M.R. Combi, W.-H. Ip, T.E. Cravens, R.L. McNutt, W. Kasprzak, R. Yelle, et al. 2006. Cassini ion and neutral mass spectrometer: Enceladus plume composition and structure. Science 311(5766):1419-1422.
Walker, C.C., and B.E. Schmidt. 2015. Ice collapses over trapped water bodies on Enceladus and Europa. Geophysical Research Letters 42:712-719.
Walker, J.J., J.R. Spear, and N.R. Pace. 2005. Geobiology of a microbial endolithic community in the Yellowstone geothermal environment. Nature 434(7036):1011-1014.
Warr, O., B. Sherwood Lollar, J. Fellowes, C.N. Sutcliffe, J.M. McDermott, G. Holland, J.C. Mabry, and C.J. Ballentine. 2018. Tracing ancient hydrogeological fracture network age and compartmentalization using noble gas. Geochimica Cosmochimica Acta 222:340-362.
Warren-Rhodes, K.A., K.L. Rhodes, S.B. Pointing, S. Ewing, D.C. Lacap, B. Gómez-Silva, R. Amundson, E.I. Friedmann, and C.P. McKay. 2006. Hypolithic cyanobacteria, dry limit of photosynthesis and microbial ecology in the hyperarid Atacama Desert. Microbial Ecology 52(3):389-398.
Watson, E.B., and T.M. Harrison. 2005. Zircon thermometer reveals minimum melting conditions on earliest Earth. Science 308(5723):841-844.
Webster, C.R., P.R. Mahaffy, S.K. Atreya, G.J. Flesch, M.A. Mischna, P.Y. Meslin, K.A. Farley, et al. 2015. Mars methane detection and variability at Gale Crater. Science 347:415-417.
Webster, C.R., P.R. Mahaffy, S.K. Atreya, J.E. Moores. G.J. Flesch, C. Malespin, C. McKay, et al. 2018. Background levels of methane in Mars’ atmosphere show strong seasonal variations. Science 360(6393):1093-1096.
Westall, F., F. Foucher, N. Bost, M. Bertrand, D. Loizeau, J.L. Vago, G. Kminek, et al. 2015a. Biosignatures on Mars: What, where and how? Implications for the search for martian life. Astrobiology 15:998-1029.
Westall, F., K.A. Campbell, J.G. Bréhéret, F. Foucher, P. Gautret, A. Hubert, S. Sorieul, N. Grassineau, and D.M. Guido. 2015b. Archean (3.33 GA) microbe-sediment systems were diverse and flourished in a hydrothermal context. Geology 43(7):615-618.
Westall, F., K. Hickman-Lewis, N. Hinman, P. Gautret, K.A. Campbell, J.G. Bréhéret, F. Foucher, et al. 2018. A hydrothermal-sedimentary context for the origin of life. Astrobiology 18(3):259-293.
White, L.M., R. Bharita, G.D. Stucky, I. Kanik, and M.J. Rusell. 2015. Mackinawite and greigite in ancient alkaline hydrothermal chimneys: Identifying potential key catalysts for emergent life. Earth and Planetary Science Letters 430:105-114.
Wilde, S.A., J.W. Valley, W.H. Peck, and C.M. Graham. 2001. Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago. Nature 409(6817):175-178.
Wray, J.J., R.E. Milliken, C.M. Dundas, G.A. Swayze, J.C. Andews-Hanna, A.M. Bladridge, M. Chojnaki, et al. 2011. Columbus crater and other possible groundwater-fed paleolakes of Terra Sirenum, Mars. Journal of Geophysical Research 116:E01001.
Yamaguchi, N., S. Anraku, E. Paineau, C.R. Safinya, P. Davidson, L.J. Michot, and N. Miyamoto. 2018. Swelling inhibition of liquid crystalline colloidal montmorillonite and beidellite clays by DNA. Scientific Reports 8:4367.
Yang, Y., I.R. Gould, L.B. Williams, H.E. Hartnett, and E.L. Shock. 2018. Effects of iron-containing minerals on hydrothermal reactions of ketones. Geochimica et Cosmochimica Acta 223:4746-4754.
Yang, Y., S. Wang, J. Liu, Y. Xu, and X. Zhou. 2016. Adsorption of lysine on Na– montmorillonite and competition with Ca2+: A combined XRD and ATR-FTIR study. Langmuir 32(19):4746-4754.
Zahnle, K., N. Arndt, C. Cockell, A. Halliday, E. Nisbet, F. Selsis, and N.H. Sleep. 2007. Emergence of a habitable planet. Space Science Reviews 129(1-3):35-78.
Zeng, X., J.L. Birrien, Y. Fouquet, G. Cherkashov, M. Jebbar, J. Querellou, P. Oger, et al. 2009. Pyrococcus CH1, an obligate piezophilic hyperthermophile: Extending the upper pressure-temperature limits for life. The ISME Journal 3:873-876
Zhang, X., G. Tian, J. Gao, M. Han, R. Su, Y., Wang, and S. Feng. 2017. Prebiotic synthesis of glycine from ethanolamine in simulated Archean alkaline hydrothermal vents. Origins of Life and Evolution of Biospheres 47(4):413-425.



























