
FORMATION AND PRESERVATION OF BONA FIDE MICROFOSSILS
Sherry L. Cady
Department of Geology
Portland State University
Abstract
The discovery of microfossil-like objects in the martian meteorite ALH84001 underscores the principal challenge facing astrobiologists in the search for fossilized evidence of microbial life beyond Earth: How can bona fide microfossils be distinguished from carbonaceous or mineral pseudofossils? The same challenge faces paleontologists searching for the earliest signs of life on Earth. Criteria for assessing whether ancient microfossillike objects are remnants of microorganisms have been established,1 and only permineralized and non-mineralized cellular remains are accepted as bona fide microfossils. Although the exact mechanism and precise chemistry of permineralization are not known, permineralized cells retain enough morphological fidelity to be recognizable and, by definition,2 are carbonaceous (composed of complex organic biopolymers). As such, permineralized and nonmineralized cellular remains harbor multiple biosignatures that include cellular morphology. To detect and confirm the biogenicity of bona fide microfossils,it is necessary to use analytical instruments that not only reveal their presence, but also reveal the biochemical nature of their organic compounds. Whether permineralized and nonmineralized microfossils harbor definitive biosignatures depends upon the intrinsic characteristics of the microorganisms, the extrinsic characteristics of their environment, and the diagenetic transformations that alter the microfossils and the fine-grained mineral matrix in which they are preserved. Discussed here are ways in which these factors affect the formation and preservation of bona fide microfossils in mineralizing ecosystems—environments that produce the types of sedimentary deposits targeted in the search for life on Mars.3
Life itself arose as a self-perpetuating product of physical processes, and it is likely that the characteristics of Earth’s earliest organisms—their size, shape, molecular composition, and catalytic properties—bore a close resemblance to the products of physical processes that gave rise to biology. For this reason, detecting the remnants of early life in terrestrial rocks is difficult. In martian or other extraterrestrial samples, it is doubly challenging.
—Andrew H. Knoll4
Introduction
Evidence of early microbial life on Earth is preserved in chemically precipitated deposits that form from mineralizing fluids (e.g., hydrothermal deposits, evaporites, carbonates, and silicified carbonates) and detrital sediments deposited by water (e.g., clays, volcanic ash, siliciclastics). Although fossil-bearing strata are sparse in ancient rocks of Precambrian age, a continuous fossil record exists from about 2 billion years ago to the present. This fossil record has demonstrated that three-dimensionally preserved permineralized microbial cells are found primarily in deposits that formed in mineralizing environments, whereas two-dimensional compressions of cells, flattened by compaction during burial,are found primarily in detrital sediments.
Our understanding of early life on Earth is based upon the various types of biosignatures extracted from permineralized and nonmineralized cells and cellular remains. Permineralized microfossils retain the morphology the microbial cells had at the time of fossilization,as well as degradation-resistant (i.e., recalcitrant) cellular components such as cell walls and extracellular sheaths. Nonmineralized cells, while compressed, also retain morphological features and biochemical signatures. The structure and stereochemistry of individual organic
compounds that comprise the recalcitrant biomolecules of permineralized and nonmineralized cells may provide biomarker compounds that can be used to distinguish some of the major groups of microorganisms.5 The isotopic signatures of cellular remains may also reveal specific groups of organisms based on the degree to which they metabolically fractionated stable carbon isotopes.6 Some permineralized cellular remains also preserve anomalous concentrations of trace elements that were concentrated in vivo7 or post mortem. While the search for ancient microfossils has produced a relatively robust set of criteria for assessing the biogenicity of even the most ancient microfossil-like objects,810 proving that purported microfossils represent fossilized microorganisms is extremely difficult and time consuming, and numerous claims of the microfossils in ancient rocks remain contentious.
In those cases where primary biomolecules (or their diagenetic equivalent) are not preserved with a microfossil (e.g.,when complete mineral replacement occurs), proving the biogenicity of such objects is often impossible. Microfossil-like objects composed entirely of minerals that do not harbor characteristics indicative of having formed uniquely via the replacement of cellular remains must be interpreted with the utmost caution. Almost all controvernial objects purported to be early Precambrian microfossils are mineralic.11,12 As noted by Buick,13 unless compelling reasons for accepting noncarbonaceous objects as microfossils exist (i.e., the morphology of the object matches the morphology of a distinctive microbe, and it is unlike the morphology of any known nonbiogenic structure), such objects should be discounted as bona fide microfossils. Only when attributes that distinguish the biogenicity of such objects are demonstrated, and the purported microfossils are confidently distinguished from possible carbonaceous or mineralic non-biologically produced objects, can they be considered bona fide evidence of life.
Cherts, Important Paleobiological Repositories on Earth (and Mars?)
Although silica deposits harbor less than one-half of the ancient permineralized microorganisms, those preserved in primary and early diagenetic cherts display some of the most exquisite morphological fidelity.14 Siliceous rocks known as cherts consist primarily of opaline, micro-and macrocrystalline silica varieties (e.g., opalA,opal-CT, opal-C, microquartz, fibrous quartz varieties, and macroquartz). Cherts form in a range of environments via a number of different processes.15 Primary chert deposits form where biogenically or nonbiogenically precipitated silica grains accumulate. Early diagenetic cherts form as a result of the replacement of preexisting sedimentary phases such as opals, carbonates, or evaporites. Regardless of the origin (biogenic or nonbiogenic) and nature (hydrated opaline or microcrystalline silica varieties)of the primary silica phase, all cherts, given enough time, will eventually recrystallize to quartz cherts. The inevitable transformation of all authigenic silica phases to quartz often masks the exact sequence of events that led to the formation of a quartz chert deposit —hence, the need to place the deposit within the context of its regional and local geology. The long crustal residence time of cherts reflects the thermodynamic stability of quartz at Earth’s surface temperatures and pressures.
Few modern-day analogues exist for ancient primary Precambrian cherts, most of which precipitated nonbiogenically. The remains of silica-secreting eukaryotes (i.e.,sponges, radiolarians, diatoms) provide the source of silica for most younger, Phanerozoic cherts.Modern chert-forming environments that can be considered analogues for Precambrian chert-forming environments include silica-depositing thermal spring ecosystems.As mineralizing environments, hydrothermal systems have an enhanced potential to preserve biosignatures indicative of their prolific heat-loving microbial communities.1618 Hydrothermal systems are habitats for a variety of thermophilic and hyperthermophilic microbial communities.1921 As shown by Cady and Farmer,22 the microbial inhabitants of thermal springs and the sedimentary structures they produce are fossilized in metastable opaline silica by a variety of mechanisms.
Quartz cherts, including those precipitated from hydrothermal fluids, are likely to have been stable at the surface of Mars throughout its history. The predominantly monomineralic composition of cherts enhances the probability that such deposits on Mars could be detected via remote spectroscopy, provided the surface area of the deposit exceeds or approximates the pixel dimension of the spectrometer utilized. Primary hydrothermal cherts could have formed on Mars as surficial siliceous thermal spring deposits23 and as shallow subsurface siliceous epithermal deposits.24 Silica leached from near-surface silicate rocks can be redeposited at depth as silcretes,
forming extensive silica-rich soil horizons, or it can be redeposited along the edges of play a lakes and evaporative shallow-water basins forming primary or replacement cherts.25 Targets in the search for life on Mars, therefore, include hydrothermal deposits, hardpan subsoils, evaporites, and paleolake basin deposits.26 On Earth, these types of deposits are relicts of ecosystems that produced microfossils predominantly via the encrustation of cells by minerals precipitated in situ (i.e., authigenically).
If microbial life did emerge on Mars and thrived in any of the targeted mineralizing environments mentioned above, it would likely have been fossilized and some of its biosignatures preserved. Given the lack of extensive recrystallization of microfossil-like objects in the ancient carbonate globules found in the martian meteorite ALH84001,27 it appears likely that cell-sized objects, whether biological or nonbiological in origin, would have been preserved with higher morphological fidelity in ancient Mars rocks than their counterparts preserved in Earth’s Precambrian geological record.
Intrinsic Characteristics of Microorganisms and Fossilization
The intrinsic characteristics of microbes that affect fossilization have been studied at various structural levels with a relatively limited number of techniques. Differences in the distribution and composition of the various extracellular and cellular components, as well as the reactivity of the subcellular biomolecules within these components of the cells, can affect an organism’s susceptibility to fossilization. Modern analogue and experimental studies that utilize various types of electron microscopy techniques have revealed the types of preservational biases that occur during the earliest stages of microbial fossilization.
The example shown in Figure 1 illustrates a type of preservational bias introduced during the silicification of

FIGURE 1. Transmission electron microscopy photomicrograph. Cross-sectional view of a high-temperature hot spring microbial community partially entombed within an opaline silica matrix (dark matrix). This image illustrates how differences in the susceptibility of various microbial taxa to fossilization could lead to biases in paleobiological information during the earliest stages of preservation.
a microbial biofilm at life’s upper temperature limit. A biofilm containing a climax microbial community, which developed over several months on a substrate deployed by the author along the edge of a chert-forming hydrothermal spring located in Yellowstone National Park, was exposed subsequent to development to mineralizing fluid. Different microorganisms in the specimen, sectioned to electron transparency, are distinguished in the transmission electron photomicrograph on the basis of their ultrastructural characteristics. Note the variable amounts of extracellular matrix surrounding the different microorganisms. Some cells appear to be isolated within the opaline silica matrix, while other, more closely spaced cells,appear to form cohesive biofilm communities surrounded by opaline silica. It is clear that the proximity of opaline silica to the cell walls of the various organisms (one of the more robust cellular components during fossilization) differs as a function of the amount of extracellular matrix they produced. How such intrinsic differences in the amount and nature of the extracellular matrix of the different microorganisms affect their preservation cannot be predicted from such an image. Since postdepositional degradation processes will alter all cells to some degree, it is likely that the intrinsic differences displayed at this early stage of fossilization will lead to preservational biases.
Experimental silicification studies have revealed gross differences in the susceptibility of Gram-positive and Gram-negative bacteria.Gram positives silicify more rapidly than Gram negatives under controlled conditions.28 Gram positive and negative refer to the way in which the cell wall of a bacterium reacts to the Gram staining procedure (the reaction being a function of the structure and composition of the cell wall). A polymeric peptidoglycan layer, located between the plasma membrane and the outer cell wall membrane, contains numerous carboxyl and phosphoryl groups that serve as metal cation-binding sites that promote mineral nucleation and growth. The thicker peptidogly can layer in Gram-positive cell walls presumably contains a greater number of reactive sites for mineral nucleation. Based on these observations, Westall proposed that Gram-positive bacteria are more likely to be preserved in the geological record.29
Other experimental studies have focused on quantifying the type, density, and distribution of biomolecules that compose the cell walls and various extracellular components of a variety of microbial taxa.30,31 Investigations to date indicate that the exposed reactive chemical groups on microbial cell walls interact ionically, as a function of pH, with solutes in the milieu surrounding the cell. The organometallic complexes that precipitate at reactive biomolecular sites provide additional sites for the sorption of metal and nonmetal ions. Fortin et al. recently reviewed the principal types of biomolecules known to alter the reactivity of microbial cell walls and extracellular layers (e.g., capsules, S-layers).32
When microbial cells attach to surfaces, they produce exopolymeric substances containing reactive biomolecules that can bind a variety of ions, including metals. Anomalous concentrations of trace metals localized by microbial cells and biofilm matrices have been proposed as possible biosignatures, the distribution of metal ions revealing the former presence of microbial cells.33,34 Microbial taxa produce exopolymers under a range of environmental conditions, the composition of which can vary for various microbial species and for the same species under different environmental conditions.35 Important topics for future research include studies to determine how environmental perturbations alter the function and composition of cellular and extracellular components,and how these changes affect the susceptibility of microorganisms to fossilization.
Extrinsic Environmental Factors and Microbial Fossilization
Distribution of Microorganisms
Most microorganisms immersed in aqueous fluids attach to surfaces and form biofilms that consist of distinct communities of cells immersed within a hydrated matrix of exopolymeric substances. Dynamic studies regarding biofilm architecture and composition, and the distribution of microorganisms within biofilms,36 indicate that biofilms cannot be represented by a single, universal model. At the present time, with limited understanding of the complexities of microbial communities, models for biofilms are best developed on the basis of actualistic studies in modern ecosystems.
Biofilms develop in nearly every environment where water and available carbon and energy sources exist. Even at the upper temperature limit for life, hyperthermophilic biofilms develop on hydrothermal mineral precipi-
tates.37 It has been estimated that the numbers of prokaryotic cells in subsurface microbial populations exceed by at least an order of magnitude the numbers of cells found within soils and the open ocean.38 Subsurface biofilms in the deep subterranean biosphere have become the subject of intense study39 since microorganisms display a propensity to form biofilms on all available surfaces in environments that favor their proliferation. The presence of microfossil-like objects in the mineral-filled fractures of the martian meteorite ALH84001,40 a subsurface igneous rock, serves as a bellwether regarding the potential importance of subsurface paleobiological repositories.
The need to search for microbial biosignatures in any structural discontinuities in rocks from Mars through which mineralizing fluid has passed freely is exemplified by the discovery on Mars of recently young surficial effluents.41 Sedimentary deposits associated with surficial effluents on Earth are extremely fine grained, having precipitated rapidly from solutions supersaturated most commonly by cooling,evaporation,or fluid mixing. Any particulate matter, including microorganisms, entrained in the subsurface fluids that escape from such effluents is often sequestered in fine-grained mineral deposits. Even if evidence of a subsurface biosphere was not discovered in the deposits that precipitated around the effluents discovered on Mars, their study would provide detailed information about the geochemistry of the planet’s near-surface aquifers.
Effects of Solution Chemistry in Determining How Fossilization Occurs
In order to retain high cellular fidelity, permineralized microfossils must be preserved intra-and extracellularly within a fine-grained mineral matrix prior to extensive cellular degradation. Fine-grained mineral matrices form when numerous, aqueously precipitated crystal nuclei reach, nearly simultaneously, the critical dimensions needed for energetically favored crystal growth. Mineral nuclei of such critical dimensions can form by the random collisions of ions or atoms within a supersaturated solution (i.e., homogeneous nucleation) or on preexisting surfaces (i.e., heterogeneous nucleation). Since reactive sites that promote surface sorption and chemical bonding occur on microbial cell surfaces, heterogeneous precipitation of mineral nuclei at these sites can result in mineral encrustation of the cell.42 Whether encrustation will lead to the formation of a permineralized cell depends upon whether, and when, minerals precipitate on the inside of the cell wall, obviously post mortem, but prior to complete degradation of the cell wall. A limited amount of microbial cellular decomposition, accompanied by early mineralization, appears to have enhanced the preservation of cellular remains in the geological record.43 Post mortem fossilization of microorganisms favors the formation of fine-grained mineral matrices, since more reactive sites for mineral nucleation become available once organisms begin to decay.
Environments containing aqueous fluids likely to reach saturation for either heterogeneous or homogeneous mineral nucleation include (1) evaporated or rapidly cooled subaerial and subaqueous hydrothermal systems and the edges of play a lake basins, (2) locations where fluids of different composition mix (e.g., at the edges of all types of aquifers), and (3) sites where fluids are stratified across chemoclines and thermoclines (e.g., in stratified lake basins,in water-filled caves, and within biofilms and microbial mats). All of these environments should be considered in the search for paleobiological repositories on Mars.
Relative Timing of Fossilization, Cellular Degradation, and Post-fossilization Events
The amount of cellular fidelity displayed by permineralized and nonmineralized cells ultimately depends upon the amount of cellular degradation that occurs prior to fossilization. In their study of microbial preservation along outflow channels of mineralizing thermal spring ecosystems, Gerasimenko and Krylov demonstrated how the apparent paleobiodiversity preserved in the modern hot spring deposits resulted from the fossilization of morphologically similar taxa that were fossilized at various stages of cellular decomposition.44 The degree to which nonmineralized cells in detrital sediments retain their cellular fidelity depends upon whether they were buried to depths where anaerobic conditions prevailed prior to extensive cellular degradation. Preservational biases resulting from variation in the amount of cellular degradation have been quantified by Bartley,45 who established a rating system for analyzing statistically the effects of degradation on cell morphology. Post-fossilization events can either enhance or limit the potential for long-term preservation of permineralized microfossils or nonmineralized cellular remains. For example, preservation is enhanced in detrital sediments if they are cemented
by a fine-grained mineral assemblage prior to physical disruption, exposure to oxidants, or metamorphic alteration. On the other hand, preservation of cellular fidelity and long-term preservation are limited in mineralizing environments if intracellular and extracellular mineral growth continues unabated.46,47
Diagenesis and Preservation of Microbial Biosignatures
Once microorganisms are fossilized, the potential for long-term preservation of their cellular components depends upon the recalcitrance (resistance to biodegradation and major chemical transformations such as hydrolysis and oxidation) of the macromolecular biomolecules.48 The preservation potential of various types of organic biomolecules has been reviewed by Logan et al. and by Allison and Briggs.49,50 Factors that degrade organic matter and determine their recalcitrance have been studied in great detail.51 The organic remains of microorganisms source much of the world’s petroleum and gas reserves. Of the four principal classes of biomolecules, the various classes of lipids, especially glycolipids and lipopolysaccharides, are most resistant to degradation in all types of depositional environments. Lipids tend to resist chemical attack, are insoluble in water, and can be incorporated in kerogens, thereby increasing their preservation potential. Although the conversion of primary lipids to their diagenetic counterparts (i.e., geolipids) involves information loss through structural alteration (e.g., hydrogenation of double bonds, aromatization of rings, loss of functional groups), the original class of lipid can often be identified even after diagenetic changes.52
The diagenetic history of the minerals that entomb microfossils in paleobiological repositories ultimately determines the length of time microfossils can be preserved in the geological record. In general, diagenesis occurs when an increase in the temperature and/or pressure of a deposit rises to the point where it alters the structural configuration of the primary organic molecules and mineral phases. Microbial cells are usually preserved initially within thermodynamically metastable mineral assemblages, and their transformation to thermodynamically stable mineral assemblages occurs as a result of diagenetic processes that operate over time scales from tens to millions of years (e.g., burial and tectonic alteration). For example, mineral diagenesis alters chert deposits with time via the transformation of primary opaline silica phases to microcrystalline quartz varieties (microquartz, macroquartz, chalcedony, quartzine, “lutecite,” “moganite”) through intermediary cryptocrystalline opal phases.53,54
Importance of Chemical and Structural Discontinuities
Regardless of the mechanism by which a microbe is fossilized or the degree to which it is altered from its primary state, the microfossil must differ either in composition or in structural organization from the mineral matrix that surrounds it in order to be detected. For example, chemical and structural discontinuities occur between nonmineralized cells or permineralized microfossils and their mineral matrix. Compositional differences between a microfossil and its mineral matrix also develop when microorganisms concentrate —actively or passively, intra-or extracellularly —heavy metals or other ions from solution. Environmental perturbations can produce gradational compositional differences between microorganisms and their mineral matrix. The geochemical changes that accompany the evaporation of fluids can be preserved in the laminated crusts that develop around microbial cells. The geochemically predictable sequence of minerals around the microfossils preserves not only a cast of the microorganisms, but also paleoenvironmental and paleogeochemical information.
It is worth noting that if mineral-replaced microbial cells are preserved within a mineral matrix of the same composition, it will be extremely difficult to establish the biogenicity of the microfossil-like objects. Even when minerals display habits similar to the morphology of microbial cells, it will be necessary to demonstrate that such minerals could not have formed abiotically. As discussed by Buick,55 a certain degree of cellular elaboration must be displayed by a purported microfossil, although the level of morphological complexity continues to be debated. It could also be possible that structural defects or structural differences between the matrix minerals and microfossil-like objects exist. If so, however, they must be interpreted with caution —mineral microstructural characteristics may reflect only differences in the relative amounts of diagenetic processing.
Conclusions
At the present time, only bona fide microfossils provide the direct evidence needed to establish extraterrestrial and earliest Earth life. Nonmineralized and permineralized cells contain multiple biosignatures indicative of their biogenicity, thereby distinguishing them from pseudofossils. Although cellular permineralization is likely to occur in nearly all types of mineralizing environments, the rarity of permineralized microfossils in the geological record suggests that much remains to be learned about how diagenetic processes affect long-term preservation. As improvements are made in the resolution limits of analytical instruments that can simultaneously “image ” micro-fossil-like objects and detect and analyze the biochemical structure of their minute concentrations of carbonaceous compounds, we may find that permineralized remains are more common than heretofore realized.
Acknowledgments
The author gratefully acknowledges the NASA Exobiology Program (NASA Grant#NAG5-9579) and the National Science Foundation LExEn Program (NSF Grant #EAR-0096354) for support of basic research in areas described in this paper.
ELECTRON-BEAM TECHNIQUES FOR MICROFOSSIL CHARACTERIZATION
David McKay
NASA Johnson Space Center
Abstract
Electron-beam techniques are extremely valuable in searching for and characterizing microfossils of all kinds. Electron-beam techniques combined with optical microscopy, ion probe, and organic microanalysis techniques can usually provide definitive information on whether a candidate feature is truly a microfossil, even if it has been considerably altered by diagenesis. In our laboratory, we use scanning electron microscopy (SEM) and transmission electron microscopy (TEM) combined with detailed electron microprobe (EMP) analysis. Specialized techniques that may enhance textural data include ion etching of polished surfaces and small grains, acid etching of polished surfaces, and oxygen plasma etching of polished surfaces and small grains. Chemical mapping of polished surfaces with EMP can reveal subtle differences in chemistry and textures that may be associated with original biogenic features, even if the original biogenic features have been destroyed. Our general sequence of analysis is optical microscopy, followed by SEM petrography, EMP chemical mapping, quantitative EMP analysis of specific phases and areas, and TEM analysis. Examples for some of these techniques are illustrated.Some samples may then be analyzed in other laboratories by ion microprobe, time-of-flight (TOF) secondary ion mass spectroscopy (SIMS), or Raman spectroscopy.
Introduction
Indigenous microfossils in rocks are absolute proof that life was once active in the environment from which the rock came. Other than finding living organisms, finding true microfossils is perhaps the most reliable proof that life was present in the environment represented by the sample. However, some important pitfalls must be considered. The purported microfossil must be shown to be a true fossil of an organism or trace of an organism. This requires detailed textural information and often requires chemical information, sometimes including isotopic ratios. Next, the microfossil must be shown to be indigenous and not contamination. For some purposes, the microfossil must be shown to be contemporaneous with the enclosing rock, but for other purposes, finding microfossils added to the rock at a later time may be quite useful. Microfossils found in igneous rocks, for example, were clearly not present during the initial cooling and crystallization of the melt,but were added after the rock cooled,generally into cracks and pores of the original rock or by alteration of its phases.
Finding and characterizing microfossils in rocks can be greatly facilitated by using electron-beam techniques, particularly in combination with other techniques. Basic instruments used in our laboratories include SEM including attached energy-dispersive x-ray analyzer (EDXA), TEM, and EMP. Petrographic doubly polished thin sections of the rock samples are necessary for the preliminary examination by petrographic optical microscope. Images from this microscope serve as maps for the more detailed mapping and analysis by SEM or TEM. For samples that will be prepared for SEM or TEM, a polished thin section is not always necessary and small chips can be used. It is usually helpful to study the chips with a good binocular microscope.
The basic information desired is the texture of the rock and its included microfossils, the location of the microfossils, their morphology, and their dimensional measurements. Basic chemical information includes maps of major and minor elements and quantitative analyses of minerals, glasses, cements, and microfossils. Ideally, each suspected microfossil feature should have well-documented texture or fabric and feature morphology in two or three dimensions, quantitative chemistry, and chemical maps showing variation from place to place of each major and minor element. The chemistry of true microfossils is not likely to resemble the chemistry of the original organism; replacement or void filling may produce a composition such as iron oxide or SiO2 totally unlike the original reduced carbon, water, and other components of the living organism. Consequently, it is necessary to document and understand the chemical effects of diagenesis and replacement that create the fossilized version of the original organism. This, in turn, requires considerable experience, familiarity with the literature, or even experimental laboratory studies of fossilization.
Sample Preparation
In our laboratory, we typically prepare samples using the following processes:
-
A representative rock chip (generally 1 to 10 mm)is used for making one or more polished petrographic thin sections, typically 30 μm thick. We make these sections in the Curatorial Thin Section Laboratory here at Johnson Space Center. Such thin sections are used for optical petrographic mapping and mineral identification, followed by SEM petrographic mapping and EDXA analysis, EMP compositional mapping, and quantitative EMP mapping of particular phases, traverses, or features.
-
A small chip (0.1 to 2 mm) containing interesting features identified in a binocular microscope is mounted on an SEM stub and coated with a conductive coating of Cr, C, or Pt for SEM examination and photography. For some purposes, the sample is left uncoated and examined at low voltage.
-
Additional small chips (0.05 to 0.1 mm) are embedded in epoxy for microtome thin-section making, and several thin sections of ~50 to 80 nm are produced for TEM examination. The objective in TEM work is to document the mineralogy with electron diffraction, high-resolution imaging, and EDXA chemistry, and to determine the ultrastructure with low-to high-magnification images. Many TEMs have electron energy-loss spectrometers (EELS) attached, which can help with chemical and mineral identification.
-
Similarly,some polished thin sections may be etched in an ion beam at low angle to bring out textures related to ion erosion resistance.56,57 Ion etching may bring out extremely delicate textures not visible in optical, SEM secondary, or SEM backscattered images of polished samples. This technique is particularly useful for detecting amorphous material such as glass or polymerized gels that develop distinctive bumpy textures in the ion etching device.
-
For some applications, one of the polished thin sections is lightly etched in hydrofluoric acid fumes for a few minutes to bring out hidden textures.58
-
Another useful technique is to place the section or chip in an oxygen plasma-etching device for a few minutes. This technique removes many kinds of organic material from the exposed surface and may differentially etch away organics creating a multilevel surface. Before-and-after SEM images can then be used to locate etched organic material.
Analysis Objectives
Generally, we want to find features that might be the remains of microbes or might have been formed or influenced by microbial life. We wish to relate these features to their environment or surroundings. If the features are fossilized microbes, we try to determine the changes in texture, morphology, and chemistry that have taken place during diagenesis and fossilization. Next we want to relate the microbial remains or traces to the original environment and the original form and type of microbe. We try to determine whether nonbiologic processes that mimic the results of biologic processes could have made the feature. The value of a biomarker such as a microfossil is greatest for forms and features that cannot easily be made by inorganic processes. Finally, we try to determine whether features are indigenous or are contaminants added later. Every effort should be made to interpret the features as artifacts, inorganic products, and contamination. Only after such interpretations are eliminated can a microfossil origin be seriously considered.
Examination Steps
Petrographic optical microscopy of the polished thin section is generally the first step. This technique can rapidly locate areas of interest that may contain potential microfossils and can provide maps for subsequent, more detailed study with an electron microscope or electron microprobe. This technique can rapidly identify many minerals. Petrographic microscope examination can also show the relationship of the potential microfossils to the rest of the rock. We use optical images as maps to guide initial study by electron backscatter petrography in the SEM or EMP.
Next, SEM studies of polished thin sections are generally done with backscatter-and secondary-electron detectors. Backscatter-electron detectors can detect small differences in average atomic number and display them as brightness variations. If the polished surface has been etched by HF fumes, ion sputtering, or oxygen plasma, the secondary electrons will show the differences in topography caused by etching and will provide data complementary to the backscatter electrons.
SEM images and EDX analyses of features on small sample chips provide a simple way to search for possible biogenic morphologies. Features may include complete fossilized microbes or their colonies, fragments of microbes, products of microbes including fossilized biofilm, or traces of microbes including molds, casts, nucleation centers for mineral precipitation, and a number of other features.59
TEM imaging, chemical analysis, and mineral identification are usually done on separate small chips sliced by microtome. We sometimes use ion thinning rather than microtoming to prepare TEM thin sections. We have also developed a technique for coring a small interesting area from a polished thin section and making the core into a TEM thin section.
EMP analysis including chemical mapping and spot or traverse quantitative analysis of micrometer-size spots can provide much useful data on the minerals, the potential microfossils, and the matrix material. Quantitative EMP data using proper standards may provide very high quality data with a standard deviation of less than 1 percent for major elements. It can also detect and analyze trace elements down to a few tens of parts per million for many elements. Our microprobe is fully automated and computer driven, and can perform analyses and make chemical maps unattended over nights and weekends.
Occasionally we need other types of data that we cannot collect in our own laboratory, so we take the samples to another laboratory that has the necessary instruments such as an ion microprobe. For example, isotope ratios for some elements (e.g., S,C, O, H, N, and Fe) can sometimes be used to help discriminate between biogenic and nonbiogenic processes or to determine whether a feature formed on Mars or on Earth. Mass-dependent fractionation of S and C are among the most common indicators of biogenic fractionation for many kinds of terrestrial samples. Hydrogen fractionation providing deuterium-rich water appears to be a distinctive flag for indigenous martian water.
Organic species analysis of mapped features is another valuable technique that can be used on mapped thin sections or chips. We have used both double laser mass spectrometry and TOF-SIMS. Other useful techniques include gas chromatography-mass spectrometry (GC-MS); however, this is difficult to use on thin sectioned samples. Laser Raman spectroscopy provides some information on molecular species present. Three-dimensional relationships within the 30-μm polished thin sections can be determined using laser confocal microscope techniques. Staining with various organic stains and examination with a fluorescence microscope can be used to detect traces of DNA, cell walls, lipids, proteins, and other specific products of microorganisms.
Examples
Figures 1 through 5 provide examples of the use of some of the above-mentioned techniques.
The examples shown for the martian meteorites are not intended to be claims that the features are fossilized martian microbes. These examples simply illustrate features identified by electron-beam techniques, which — based on their morphologic similarity to known biogenic features in terrestrial samples —are candidates for additional investigation to collect detailed data on mineralogy,chemistry, isotopes, and ultrastructures. Only a full set of such data can provide convincing determination of either biogenetic origin or lack of it. The small size and the complexity of these features points out the need for additional types of microprobe instruments capable of isotopic and organic chemical analysis at the submicrometer scale.

FIGURE 1.(a) SEM view of ion-etched polished section of lunar volcanic ash (sample 74002). Here, the delicate thin branching textures of the ilmenite are clearly shown in high relief against the glassy matrix. The medium-gray crystals are olivines, and the bright crystals are spinels. Ion etching was used to characterize the mineralogy, textures, and cooling history of these volcanic particles60 and to reveal subtle matrix textures in impact-breccias.61 This technique may reveal structures below the resolution limits of ordinary backscattered electron mapping, and enables much higher spatial resolution secondary electron imaging to be used on polished thin sections. For lunar regolith samples, the technique revealed details not detectable by any other technique. (b) SEM view of a lightly ion-etched polished section of the martian meteorite, Nakhla. The U-shaped grain is iron sulfide (pyrite). Note the difference in relief among the various phases. A thin rim, somewhat lower than the calcium phosphate grain, completely surrounds it. This rim is not obvious on the unetched sample, but it may contain critical information on the change in conditions leading to incipient alteration of the pyrite.

FIGURE 2. Chemical maps, made by the electron microprobe, of a single carbonate globule on a fracture surface of the martian meteorite ALH84001. The chemical zoning is apparent for all elements, but in particular for the two magnetite-rich zones. Most investigators now agree that the complex zoning is consistent with precipitation of the carbonates from liquid water or brine at relatively low temperatures. Of particular importance is the presence of small high-Ca regions within the magnetite-rich zone. These high-Ca regions correspond to high P regions (not shown). This presence of Ca phosphate in the magnetite-rich carbonate may be an important constraint on models for formation of both the carbonate and the magnetite.

FIGURE 3.(a) SEM view of recent microbes in hot springs samples from Jemez Springs, New Mexico. These microbes are being replaced and fossilized by deposits of silica precipitating from the hot cooling water. The microbes are partially embedded in a biofilm, which is also being converted to silica. It has only recently been realized that biofilm may fossilize and be preserved, often better than microbial cells. (b) SEM view of coccoid bacteria from Jemez Springs, New Mexico. The bacteria were multiplying and forming chains. Both bacteria and surrounding biofilm are in the process of being replaced by silica. Images by Carlton Allen.

FIGURE 4.(a) SEM view of elongated bacteria form,which has been covered by a fine network of iron hydroxide fibers (ferrihydrite). The organism, which provided the form for the coating, has been lysed or dissolved away. Individual ferrihydrite fibers are only ~10 nm in diameter. This sample was from a laboratory experiment that grew and fossilized the microbes using basalt chips and groundwater from deep within the Columbia River basalt in Washington State.62 (b) Backscatter electron (BSE) image of a polished thin section of the martian meteorite Nakhla. It illustrates cracks in the olivine (ol and pyroxene (px grains filled with smectite-like clay mineral (cl, carbonates (carb, and a glass (gl . This image shows the ability of BSE to display subtle differences in average atomic number at a fine scale and to illustrate fine-grained textures. The spatial BSE resolution here is less than 50 nm at 15 kV. Several ~2-μm-round features (r are present which might be mineral concretions. However, these forms are candidates for possible fossil microbes. Additional analyses are indicated, including EMP carbon analysis and mapping, search for other possible biogenic traces such as P or N, search for organics using TOF-SIMs or fluorescent staining techniques, and possibly coring for TEM imaging and mineral identification. If carbon is present, isotopic analysis and mapping by ion microprobe may be indicated. Deuterium analysis of the clay matrix would likely determine whether the clay was formed on Mars (the current interpretation for these secondary phases in Nakhla) or on Earth.

FIGURE 5.(a) SEM view of a complex fracture surface from a chip of the martian meteorite, Nakhla. The features resemble in some ways the fossilized biofilms and bacteria cells illustrated in Figure 3. Composition data on the larger rounded features reveal that they are enriched in Fe and O compared to the substrate. The bright, filmy material is enriched in Si and O compared to the substrate. These features require further analysis to determine whether they are biogenic and indigenous to the Mars meteorite. (b) SEM view of another fracture surface of a chip of Nakhla. The fracture surface is full of rounded and bumpy forms in a complex three-dimensional matrix. The meteorite appears to break apart preferentially along preexisting fracture surfaces, which are often filled with secondary mineral deposits as illustrated in Figure 2. These features require further analysis to determine whether they are biogenic and indigenous to the Mars meteorite.
Summary
Electron-beam techniques combined with optical microscope surveys form a powerful method of finding and characterizing microfossils in almost any kind of rock. We and others are developing criteria for determining what features are truly microfossils and what features are inorganic structures that may mimic microfossils. In addition, detailed chemistry and mineralogy of the microfossil and its surrounding matrix may help determine whether the feature is indigenous. As the spatial and chemical resolution of electron-beam instruments improves, these instruments will become more valuable in finding, documenting, and understanding fossil organisms or their traces. If identified, such fossils and their traces constitute robust biosignatures in early terrestrial rocks and possibly in nonterrestrial rocks. The application of these techniques to samples from beyond Earth is just beginning, but promises to be a major part of future astrobiology. The complexity of the required characterization requires that samples be returned to terrestrial laboratories for detailed, multitechnique analysis.
ORGANIC DETECTION
Luann Becker
Department of Geological Sciences
University of California, Santa Barbara
Abstract
The search for biogenic organic matter on Mars and other moons and planets in our solar system is rapidly emerging as a result of technological advancements and the study of early “life ” on our own planet. As we have learned from our previous mission to Mars and the examination of martian meteorites, the criteria for establishing life require the appropriate strategy. Thus, a viable approach will require careful mapping of the surface from orbit for the selection of appropriate landing sites, robotic space missions equipped with several life detection in situ techniques for selection of samples, and sample return missions for additional verification of in situ results and laboratory measurements. The development of life detection techniques for future missions to Mars and beyond may require the appropriate Earth analogue to test the viability of these methods and further maximize our chances for scientific success.
Introduction
NASA is now entering a new phase of planetary exploration with the current scheduled missions to Mars and future missions to some of the moons of Jupiter and Saturn (e.g., Europa and Titan, respectively). Our interest in Mars is motivated by the recent studies of martian meteorites that suggest that the early history of the Red Planet was remarkably similar to the Earth’s, where life apparently arose 4 billion years ago. If this is indeed the case, then Mars was presumably a much warmer, wetter, planet than it is today. This hypothesis is further supported by orbital imagary returned by the Viking, Mariner, and Mars Global Surveyor (MGS) spacecrafts that all show compelling evidence that copious liquid water existed on the surface of Mars in the past. In fact, new images provided by MGS suggest that there may be current sources of liquid water at or near the surface of the Red Planet. Other MGS data suggest that an ocean may have once existed at high northern latitudes, and valley networks apparently once carried water into the northern basin. All of these data suggest that life could have arisen on Mars in liquid water environments.
The search for extinct organic matter (i.e., organic matter generated by now-extinct organisms, in rocks, sediments, and ices from Mars and other planets and moons in our solar system) is critical to the determination of where life existed. Missions to Mars have the potential to address whether life arose there in a separate origin and may further provide information about our own prebiotic evolution, a record that has all but been erased from Earth’s crust. Future missions to moons like Europa may enable us to search for clues of life in a liquid water environment. It seems clear that the potential for learning about life beyond our own planet is one of considerable interest to scientists and the general public alike. Yet, as we learned from the Viking missions, the search for life signs is problematic and requires an appropriate strategy that will maximize our opportunities to properly examine these compelling questions. This report discusses what is known about the organic matter in martian samples and further addresses techniques and strategies for future missions to Mars and beyond.
The Viking Missions
One of the major objectives of the 1976 Viking missions was to search for extinct organic compounds preserved in soils on the surface of Mars and to determine their structure and abundances.63 The determination of the structure of organic compounds would provide the information needed to assess both “abiotic ” ((meteoritic) and “biotic ” ((intact martian organics) syntheses of organic compounds. Thus, a gas chromatograph coupled to a mass spectrometer (GC-MS) was the method of choice selected to combine both sensitivity and structural specificity.
Developing a GC-MS instrument that would be capable of flight and still achieve the goal of detecting “intact ” organic compounds on Mars was a formidable one and required some modifications from the conventional
instrumentation. In particular, the chemical extraction of organics used to assess terrestrial samples was far too complex to miniaturize and operate reliably on the Viking landers. Therefore, thermal volatilization for the detection of organics and their degradation products was selected as the simplest and most flight-compatible approach. Another aspect of the original GC-MS design, which was later discarded to simplify the package, was a sample oven tied directly to the mass spectrometer for the detection of more complex, less volatile (kerogen-like) compounds.
The Viking GC-MS revealed that very little, if any, organic material is present in the soils sampled at the surface of Mars. The absence of organic carbon was attributed to the unusual oxidizing properties of the soil detected during measurements of the martian atmosphere. A recent assessment of this latter hypothesis demonstrated that nonvolatile salts such as benzenecarboxylic acids, oxalic acid, and acetic acid are metastable intermediates of meteoritic or martian organic debris under oxidizing conditions.64 Organics converted to carboxylic derivatives would be difficult, if not impossible, to detect using GC-MS. Thus, in evaluating the overall Viking strategy, future instrumentation for the detection of extinct organic matter should include the following:
- The capability to volatilize the more complex kerogen-like component (e.g., a high-temperature oven)— such an improvement may also address the composition of the minerals found in the martian surface materials, a point that we have yet to address with either the Pathfinder or the MGS missions; and
- Instrumentation that is capable of detecting the alteration products of meteoritic and intact biologically mediated martian organics.
Organic Compounds in Martian Meteorites
Although the Viking landers were unable to address the question of life on Mars, the orbiters did provide important measurements of the martian atmosphere that led to the identification of a unique suite of meteorites on Earth as martian in origin. To date, all that is known about the organic matter on Mars comes from the examination of these meteorites. Perhaps the most famous of the group (16 total) is the Allan Hills (ALH84001)meteorite that was reported to have evidence of both putative martian fossil remains and organics in the form of polycyclic aromatic hydrocarbons (PAHs).65 The announcement of the ALH84001 results has led to a reassessment of how to search for life on Mars and elsewhere.In fact,the techniques used to evaluate the origin of organic matter in ALH84001 and other martian meteorites are central to any discussion of the detection of extinct organic matter on Mars. Much of the controversy surrounding ALH84001 centers on stable carbon isotope studies (δ13C) of the carbonate and the associated organic matter. Thus, subsequent studies of ALH84001 have focused on determining the δ13C values of specific organic compounds isolated from various mineral phases to assess the sources (biotic versus abiotic) of this material.
Stable Isotope Measurements of Martian Organic Matter
Two independent investigations of the stable carbon isotopic compositions of the organic matter in ALH84001 indicate that a small portion (~50 out of 250 parts per million [ppm]) of this material has a δ13C value of –15 ‰. 66,67 A δ13C value of –15 ‰ would be unusual for martian organics since 13C//12C measurements of trapped gases in some martian meteorites indicate that two distinct carbon reservoirs exist on Mars:
-
An isotopically heavy component (atmosphere) enriched in δ13C (+36 ‰) and
-
A high-temperature igneous (i.e., mantle) component (δ13C –20 to –30 ‰).
On the other hand, a δ13C value of –15‰ is consistent with a “kerogen-like ” component in carbonaceous chondrites. Thus, some portion of the organic matter in ALH84001 is extraterrestrial in origin and likely derived from meteoritic or cometary debris that is exogenously delivered to the surface of Mars. Recent measurements of the organic matter isolated from the Nakhla meteorite also indicate that a portion of this material is extraterrestrial in origin with a δ13C value of –15 ‰.68 These new data further support the notion that extraterrestrial meteoritic
or cometary debris has been accumulating on the martian surface and sequestered in the crust over geologic time (Nakhla has been dated at ~1.2 billion years old). The study of extinct organic matter in martian meteorites brings some new considerations to bear on the selection of analytical techniques for future life detection missions. These considerations include the following:
- Extraterrestrial organic matter is being incorporated into the soils and rocks on Mars;
- Measurements of the δ13C compositions of organic compounds in martian samples could provide the information needed to distinguish abiotic and biotic sources of organic matter; and
- The detection of putative fossils in martian rocks (e.g., ALH84001) is not sufficient to determine that life evolved on Mars.
Future missions to Mars such as the European Mars Express will carry mass spectrometers capable of measuring stable carbon isotopic compositions in rocks and soils on Mars. The earliest of these measurements will come some three decades after the Viking landers produced null results regarding the existence of organic molecules at two places on the Martian surface.
Strategies in the Search for Extinct Organic Compounds on Mars and Beyond
The recognition of the martian meteorites has vastly expanded our knowledge of the chemistry, mineralogy, age, and isotopic composition of the martian crust. However, most of the meteorites we have are young, are igneous in nature, and appear to be from a similar location. In fact, isotopic dating of the Los Angeles meteorite, which was discovered in 1999, revealed an age of only 175 million years, contemporary in geologic terms. These new results suggest that Mars has been tectonically active in the recent past and may have a much younger surface then had previously been assumed (Meteoritical Society meeting, August 2000). As several studies have now indicated, however, none of these rock types appear to be conducive to the preservation and accumulation of organic matter.
In formulating a strategy to search for extinct organic matter on other moons and planets, we can draw some insight from our own geologic record. Experience gained from decades of searches for ancient evidence of past life on Earth suggests three major stages in the detection of extinct life on other planets. First comes the identification of specific sites of likely fossil preservation associated with the past presence of water; second is the selection of fossiliferous rocks for the studies of those locations; and third is the actual analyses of the rock themselves for structural, molecular, or isotopic evidence of past life. On other planets, site selection will depend upon geological and compositional observations made from orbit, rock selection based on chemical measurements made by landers or rovers, and either in situ analyses or sample return to Earth.
The most suitable lithologies for the preservation and accumulation of organic matter on Earth are sedimentary rocks that are typically fine grained and are characterized by well-defined aqueously derived mineral assemblages. We have yet to identify sediments on Mars or any other moon or planet; however, recent MGS images have provided new insight on locations that could offer the appropriate environment (aqueous) needed for the identification of these rock types. While it appears that evidence for fossil life cannot be resolved by orbital observations, future chemical investigations of the local environment may reveal the ecological imprint of biological activity. Ecological signatures, likely requiring broader spatial coverage to direct rovers and landers equipped with a network of instruments, may be reflected among local rocks in the chemical or mineralogical compositions or in gradients of these properties. On the other hand, depending on spatial resolution, orbital observations should be capable of detecting carbonate formations that are either directly or indirectly associated with biological activity on Earth, provided these rocks are not covered in dust that hinders their detection. Definitive evidence of fossil life, however, will require more direct and detailed observations of appropriately selected rocks. Rock selection serves two purposes:
-
To demonstrate that the sample formed in sedimentary environments, and
-
To show that it contains organic matter.
Although the absence of organic matter in a sedimentary rock would not preclude a biological imprint, a rock
containing organic matter would offer more lines of evidence to follow in establishing the presence of past life.
This strategy would, of course, require sample return to answer the more critical question of whether they contain biologically mediated Martian organic matter. It may, however, be possible to obtain additional information about the associated organic matter present in these mineral assemblages in a single measurement that is both capable of flight and is nondestructive to the sample. Such an approach must address the fundamental question of The presence or absence of organic matter, its source (abiotic versus biogenic), and the associated mineralogy, thus Fulfilling the requirement of establishing a Martian biota without sample return.
A potential candidate compound to search for would be amino acids. Amino acids play an essential role in Biochemistry as we know it and have properties such as chirality’s (handedness) that can be used to distinguish Between biotic and biotic origins.69 These compounds occur as racemes mixtures (D- and L-enantiomers) in carbonaceous chondrites, and only the L- are used in the proteins and enzymes in life on Earth. In fact, the detection of enantiomerically pure amino acids in a martian sample could be compelling evidence that life had occurred on Mars.
However, the identification of amino acids that seem to have all of the essential requirements needed to determine the origin of life on Mars, could be compromised if we do not sample properly. On Earth, temperature changes due to seasonal fluctuations and/or climate change result in an enhancement in racemization of amino acids. Recently, the effect of seasonal fluctuations in shallow lakes and Siberian permafrost on the rate of racemization was evaluated, and it was concluded that such changes were affecting the preservation of the amino acids.70
For sample selection sites on Mars where recent soil deposition is due to aeolian rather than fluvial or sedimentary processes, much of the soil, even at depth, would be considered well mixed with regard to exposure to high temperatures. Under these conditions, racemization of all amino acids would be complete even if life existed in the first billion years of martian history. However, in deep ancient (i.e., not gardened) rock and even in surface rocks at northern polar latitudes, the temperature maxima are currently low enough to allow incomplete racemization of most amino acids over martian history.
Thus, the search for viable organic matter depends, not only on the appropriate biomarker, but also on the proper location for sample selection and preservation of the organic signature. The detection of in situ organic matter may ultimately depend upon a variety of techniques with differing sensitivity, molecular structural information, sample preparation and degree of reliability in distinguishing biogenic and abiogenic production mechanisms.
Methods for the Detection of Extinct Organic Matter in Martian Samples
Raman Spectroscopy and Biomineralization
A major improvement in the study of complex biological systems has been the use of resonance Raman spectroscopy.71 Due to the rapid advancements in laser technology, most wavelengths between 190 and 1064 nm are available and easily implemented in a flight-compatible package. Although fluorescence can interfere with Raman spectra, most bacterial taxonomic markers (pigments) absorb in the fluorescence-free ultraviolet region (e. g. , detection limits of 1 ng/ml for a single bacterial cell). The application of Raman spectroscopy is being considered by several groups participating in future missions for the analysis of complex mineralogies such as meteorites and the detection of small quantities of biologically important chemical species (e. g., sulfur, carbon).
While the current wavelengths being studied (far red or near infrared) are not ideal for the detection of specific organic compounds (amino acids, lipids, etc.), they are suitable for material such as microbial pigments (biominerals) and their residues (e. g., phycocyanin in cyanobacteria at 530-560 excitation). For example, Raman spectra of deep-sea corals (Figure 1, left) closely resemble spectra for magnesium calcite with the exception of pink coral, which shows additional bands in the region 1010-3758 cm-1 that arise from a carotenoid pigment.72
More recently, Raman spectra (excited at 1064 nm) of epilithic Antarctic lichen encrustation embedded in Beacon sandstones (Figure 1, right) indicate vibrational bands for β -carotene (1136 cm-1 ) and α -quartz (463 cm-1 ).73 The Antarctic geobiological systems are the closest analogues we have on Earth to potential martian ecosystems. Thus, Raman spectroscopy offers a nondestructive approach suitable for the assessment of biomolecules of exobiological importance.
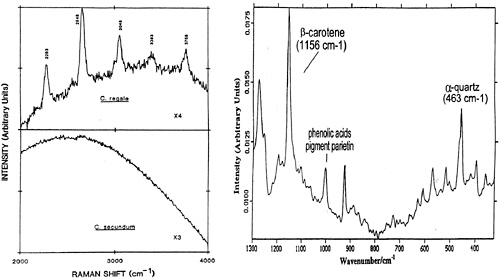
FIGURE 1. (left) Raman spectra (2000-4000 cm-1 ) of pink and white corals (J. Urmos, S.K. Sharma, and F.T. Mackenzie, “Characterization of Some Biogenic Carbonates with Raman Spectroscopy, ” American Mineralogist 76:641, 1991); (right) Fourier transform Raman of Xanthoria elegans lichens from Crater Cirque, Victoria Land, Antarctica (H.G.M.Edwards, D.W. Farwell, M.M. Grady, D.D.Wynn-Williams, and I.P. Wright, “Comparative Raman Microscopy of a Martian Meteorite and Antarctic Lithic Anologues, ” Planet. Space Sci. 47:353, 1999).
Laser Desorption Mass Spectrometry
One of the methods used for the detection of extinct organic matter in martian meteorites has been laser desorption mass spectrometry (LDMS).74,75 LDMS has the capability of examining organics on very small (~10 ìm) to large (millimeter) sized grains in low amounts (nanomoles) “intact ” without any further sample preparation. The very fine spot size of the laser allows for detailed analyses of an individual particle and a critical assessment of the distribution (heterogeneous versus homogeneous) and abundances (parts per billion [ppb] to ppm) of individual organic compounds. LDMS has also been used extensively for the detection of biomolecules and their fragments (DNA, proteins, peptides, etc.; see Session 3, “Detecting Extant Life ”).
The first organic compounds identified in ALH84001 were PAHs.These are ubiquitous in the universe, and on the Earth, they are the products of slow geochemical diagenetic reactions and the burning of biomass. The chemical architecture of PAHs, however, precludes any unique interpretation of their synthesis or source (i.e., biotic or abiotic). When coupled with the proper technique, however (e.g., carbon isotopes), LDMS is a useful approach for the detection and structural interpretation of intact organic compounds and their degradation products.
A recent examination of PAHs in the Nakhla meteorite (observed to fall in northern Egypt in 1911) revealed several peaks that were interpreted as the oxidative derivatives of PAHs.76 These degradation products (Figure 2) may be the first indication of the potent oxidant present in the martian regolith that converts most or all of the organic compounds to carbon dioxide in a relatively short period of time if left unprotected.77
Unlike Nakhla (Figure 2, bottom), the ALH84001 PAHs (Figure 2, top) do not indicate significant oxidation suggesting that these compounds were quickly sequestered into minerals and sediments where they were protected from the effects of the strongly oxidizing surface environment.Thus, LDMS may be a good technique for evaluating the degree of oxidation and stability of organic compounds that are formed in situ (martian organics) or are delivered exogenously to the surface of Mars.

FIGURE 2. Laser desorption mass spectrometry of polycyclic aromatic hydrocarbons in carbonates examined in the ALH84001 (top) and the Nakhla (bottom) martian meteorites. The PAHs detected in ALH84001 are identical to those reported by McKay et al. in 1996. The Nakhla PAHs also display some of the same PAHs as ALH84001;however, evidence for oxidation of pyrene (mass =202) is evident with the detection of mass = 218 (202 + 1 oxygen) and mass = 234 (202 + 2 oxygens). This may be evidence for the exposure of organic compounds to the “superoxide ” atmosphere detected by Viking in 1976. The recognition of these oxygenated PAHs (or oxy-PAHs) may have been impossible to detect using the pyrolysis GC-MS available on the Viking lander.
Iron Isotopes as Biomarkers
As we have learned from examining the martian meteorites, isotopic fractionation studies of the lighter elements (i.e., carbon, sulfur, etc. ) are extremely useful in determining the origin of extinct organic compounds (biogenic versus abiogenic). Recently, advances in mass spectrometric techniques have led to research into the fractionation of Fe and other transition metals (e.g., Cu, Zn, Ti, Mo). Since none of these elements are radioactive, nor are they the products of long-lived radioactive decay, the isotopic variations observed must result from massdependent fractionation. Some of these metals have been studied in the laboratory to ascertain their biological and nonbiological fractionation processes. Isotopes of iron (Fe) are of particular interest since Fe-bearing phases, including biominerals, are widespread, are resistant to alteration, and on Earth, have been linked to biologically mediated processes.
Like carbon, Fe can serve as an electron donor, providing metabolic energy to some microbes under both aerobic and anerobic conditions. Fe is also intimately associated with the oldest rocks on Earth. Coupled with its availability for carbon fixation (Fe2+) , its association with banded iron formations (BIFs) and some of the deepest organisms on the phylogenetic tree, Fe usage appears to be an early invention of life. Thus far, most of the research on Fe isotopes has involved establishing the degree of fractionation in biological and nonbiological processes. For example, it has been shown that δ56Fe in marine sediments is shifted by -1.5‰ compared to igneous rocks. However, some complications have been recognized such as the similarity in fractionations of iron
meteorites, loesses, and paleosols (only 0. 3 ‰) to igneous rocks.78 Thus, as is the case for any new tracer, more measurements are needed to establish the various fractionation patterns and to further determine the usefulness of Fe isotopes as a biomarker of life. Nevertheless, the research to date clearly shows the potential for the use of Fe isotopes in biogeochemical life studies and should be closely watched as, perhaps, a unique and novel approach in the search for life signs on Mars and beyond.
Extinct Fossil Life (Electron-beam Techniques)
Evidence of extinct fossil structures can be sought in detail at various detection levels ranging from macroscopic stromatolitic structures, to microfossils, to the intermolecular distribution of carbon isotopes and organic compounds of the associated organic matter. Depending on their depositional environment, fossilization mechanism and diagenetic history, both macro-and microscale biogenic structures, including biofilms, can be preserved with varying amounts of their original organic contents. In all cases, especially the absence of organic matter, the field “context ” of the sample site and the texture and fabric of the structures are critical in determining biogenicity.. Since the latter information also undergoes degradation over time, laboratory and field studies of both the fossilization and the subsequent diagenesis processes are needed to determine the time scales over which biogenic features are lost or preserved under various environmental conditions.
Electron-beam techniques (scanning and transmission electron microscopy) combined with optical microscopy provide powerful tools for the characterization of putative fossil structures, often in three dimensions, with respect to their location within the mineral matrix, their morphology, texture, and size. Energy-dispersive x-ray analyzer (EXDA) and electron energy-loss (EEL) attachments, as well as electron microprobes (EM), provide essential chemical and mineralogical measurements that support biogenic origin. Similarly, ion beam techniques like time-of-flight secondary ion mass spectrometry (TOF-SIMS) afford high spatial resolution imaging of organic matter and even stable isotopic measurements on specific structures or locations within structures.
None of these techniques alone, however, can adequately address the question of biogenicity. Nowhere is this more evident then in the assessment of ALH84001 where this array of methods was used to evaluate potential putative martian fossils, yet agreement amongst the scientific community that such evidence was biological could not be achieved. The absence of organic remains within potential fossil structures detected using this approach makes the problem even more difficult. In this context, studies aimed at determining what biological morphologies, fabric, and features are not produced by inorganic processes take on high priority. These considerations suggest that fossil life detection by microscopy alone remains a hope for the future rather than a promise for the present.
In Situ Techniques for Life Detection
The Grand Challenge
As part of NASA’s strategy in the search for life signs on Mars and elsewhere, the Jet Propulsion Laboratory (JPL) in Pasadena, California, has initiated the Grand Challenge Program (GCP) to develop in situ measurement techniques that are capable of detecting chemical signatures of life. The GCP will invoke interdisciplinary research goals combining science and technologies that will go beyond the current state of the art and will be implemented in the next decade for future missions in space science exploration. Funding for the program will also remain independent from the NASA R&D funding commitments, allowing participants to continue to develop technologies unimpeded by the normal fluctuations in the agency’s budget.
Among the topics of interest for Phase 1 of the GCP program are the following:
-
Identifying chemical signatures for life that are “non-Earth centric ” and independent of specific molecules or unusual life properties on Earth;
-
Developing techniques and statistical strategies for identifying signatures in a background of “non-life ” and testing them on Earth; and
-
Devising concepts for how measurements can be implemented in situ with emphasis on the development of “miniaturized ” in situ instruments.
The GCP concept so far shows great promise in the development of new approaches and instrumentation that will emphasize these topics to explore for life on Mars and elsewhere in the solar system.
Sensor Web
A promising new technique being developed at JPL is a sensor web system that consists of several spatially distributed sensor pods that can be deployed to monitor and explore a variety of environments.79 Of particular interest is the use of the sensor web to conduct long-term monitoring of biogenic gases (e. g., H2S, FeS, NH3, or CH4 oxidation) in extreme environments on Earth, as well as on Mars and bodies in the outer solar system.
The small size and minimal energy and weight requirement of the sensor web (essential for any flightcompatible life detection technique) are demonstrated by considering an analogy to Sojourner rover, which weighed about 15 kg. At a minimum communication distance of 50 m between pods, a monitoring area of 500 × 500 m 2 area is possible. This would be a true advantage over a rover like Sojouner that only moved a radial distance of 7 m on the surface of Mars. On the other hand, a sensor web deployed with a rover could provide information needed to track down appropriate samples to evaluate. Thus, sensor web represents the type of new technology that can be used for future life detection missions.
Earth Analogues and the Search for Extinct Organic Matter Beyond Mars
Europa
If liquid water exists beneath the surface ice layer on Europa, then one of the environmental requirements for life as we define it will have been met. Additional observations of a source of energy for metabolism and the recognition of the availability of key biogenic elements would be a further indication that life may have originated and could still be present on Europa. The detection of energy sources and possible biogenic elements, however, is not a trivial one. The major goal for exploration of Europa is to establish the presence of an ocean beneath the ice cap. If this is indeed the case, then the emphasis will shift to exploration at depth within the ice to address the question of europan life.
Like Mars, the ices of Europa require analytical approaches that will allow one to differentiate between abiotic and biotic sources of organic material. However, much of what may be delivered exogenously or deposited in situ (europan organic matter) in the ice layers could be destroyed by the intense radiation at the surface of Europa. On the other hand, as has been suggested by the Galileo images, layers of dark material (organics or hydrated minerals) incorporated just a few millimeters beneath the ice may remain unaffected by irradiation. Nevertheless, the severe conditions existing at the surface of Europa present a significant challenge that will require appropriate tests to enhance our opportunity to successfully detect extinct organic compounds. The development of specific instrumentation that is directly related to life present in the ice and ocean environment of Europa will require the following:
-
In situ chemical approaches capable of probing for specific cations, anions, and organic compounds essential to metabolism and life;
-
Impact penetrator-type devices for burrowing through the ice;
-
A cryobot for exploring the ice environment; and
-
A hydrobot for exploring the aqueous environment underneath the ice cap.
Such approaches are truly in their infancy and, thus, require further testing with respect to their development and reliability.
Lake Vostok
Since the exploration of Europa will necessitate an advanced technological approach, testing the equipment using an appropriate Earth analogue, such as Lake Vostok near the South Pole in Antarctica, would greatly facilitate the development of in situ instrumentation. Exploration of the ice cap and lake at Vostok will provide a unique environment for the deployment of a variety of instruments. In addition, several advanced drilling approaches have been developed to recover ice from Lake Vostok.80 Ice core studies have already revealed significant information pertaining to the nature of climate change over the past ~400, 000 years.81 Moreover, recent analyses of portions of Vostok ice cores revealed between 2 × 102 and 3 × 102 bacterial cells per milliliter and low concentrations of potential growth nutrients, suggesting that Lake Vostok may contain viable microorganisms (Figure 3).82,83 The detection of microorganisms under a thick ice cap (4000 m) in Lake Vostok supports the

FIGURE 3. The examination of accreted ice from the Lake Vostok ice core drilling project revealed between 200 and 300 bacterial cells per milliliter of meltwater and low concentrations of potential growth nutrients suggestive of viable organics living in the lake underneath 4000 m of ice. Studies of Earth analogues such as Lake Vostok may be necessary to prepare for future missions to moons like Europa, which may have a liquid ocean beneath a thick ice cap. SOURCE:D. M. Karl, D. F. Bird, K. Bjorkman, T. Houlihan, R. Shackelford, and L. Tupas, “Microorganisms in the Accreted Ice of Lake Vostok, Antarctica, ” Science 286:2144-2147, 1999.
notion that the lake could serve as a terrestrial analogue to guide the design of samplers and experiments to be used in life probe missions to Europa and beyond (e. g., Titan, Callisto, Ganymede).
Hydrothermal Vents
Another possible terrestrial analogue for testing advanced technological approaches would be extreme environments such as those around hydrothermal vents. The potential for assessing life at high temperatures can be tested by identifying the sources of energy, the quantity of energy, and the extent to which the energy can be focused. These kinds of studies would greatly facilitate our effort to establish the minimum energy requirements needed to support life beyond Earth. Current studies to quantify energy sources have focused on organisms (chemoautotrophs, organisms that gain energy from chemical reactions) that are capable of capturing energy from environments to generate organic compounds from inorganic forms (e.g., carbon, nitrogen, sulfur, phosphorus). As a complement to these empirical and laboratory efforts, in situ measurements in the hydrothermal environment would allow us to further constrain the energy requirements needed to support organisms. Efforts are under way to use submersible technology to assess the energy sources available and to further test the reliability of the instrumentation in these environments.
Conclusions
The search for life on planets like Mars and perhaps the moons of Jupiter is a testable hypothesis that is of immense interest to the public and researchers worldwide. Our future missions to Mars should begin with the examination of the latest MGS images that will aid in the assessment of the proper landing sites and will maximize the probability of detecting any record of a past martian biosphere. The instrumentation selected for the detection of extinct organic compounds must be capable of differentiating between mechanisms of formation (biogenic versus abiogenic) and the sources (martian or meteoritic) of the material. Beyond these in situ efforts, samples should be “cached ” for sample--return missions.
Future exploration efforts beyond Mars should begin with observations carried out in orbit and possibly the deployment of rovers and long-term sensor systems that will provide preliminary measurements of the environment. In addition, testing of instrumentation in the appropriate environment on Earth is essential to the development and implementation of these analytical approaches in a martian or europan environment. The information gained from such approaches can then be used in the development of a series of missions to explore for life signs.
BIOMARKERS AND BIOSIGNATURES OF FOSSIL ORGANIC COMPOUNDS
J. Michael Moldowan
Department of Geological and Environmental Sciences
Stanford University
Abstract
Biosignatures in organic lipids include specific biomarker compounds and other features (stereochemistry, nonrandom distributions) of the lipid fingerprint that cannot be explained by abiogenetic synthesis. In a general screening for organic biosignatures both lipid distributions and the occurrence of complex molecular structures requiring biosynthesis (biomarkers) would seem important. In screening for biomarkers it may be possible to recognize whether any of the three extant domains, Bacteria, Archaea, and Eukarya, were present by including hopanes, head-to-head isoprenoids, and steranes, respectively, in the search protocol. Chirality, and specifically the enatiomeric ratio, is a potentially important biosignature of some biomarkers related to their origins of enzyme assisted synthesis. Enantiomeric excess is preserved through diagenesis in lipids, such as steranes and hopanes, which have quaternary asymmetric carbon atoms in their structures. Relatively mild diagenetic conditions can result in racemization of molecules with only tertiary asymmetric carbon atoms, such as acyclic isoprenoids and amino acids. Distributions of non-biomarker compounds can amount to a biosignature. An example would be a normal alkane distribution that shows predominance of the even (or odd) carbon numbered compounds, reflecting biosynthetic pathways that homologate by two carbon atom increments. Measurements of stable carbon isotope ratios on individual compounds could suggest enzymatic intervention by offering the possibility to recognize big carbon isotopic fractionation effects in individual molecules, thus improving the legibility of biosignatures.
Introduction
In the search for what evidence could be indicative for extraterrestrial life, the analysis of lipids should be very important. Most organic lipids on Earth are derived from organic cellular components. The more complex ones found in sediments or the environment that can be unambiguously related to enzymatic synthesis have been called biological markers or biomarkers since the 1960s.84,85 Recently, the search for evidence for extraterrestrial life has catalyzed a popular extension of the use of the term “biomarker ” to cover all manner of evidence for life.. However, at a recent workshop (Biosignatures Workshop, 2000) it was agreed by consensus that “biomarker ” would be used in the classical sense to refer to molecular fossils and that any type of evidence for life, including the occurrence of the so-called molecular fossil biomarkers, would be referred to as a “biosignature.”86
Discussion
Definitions
Biomarkers or biological markers have been defined as “complex organic compounds composed of carbon, hydrogen and other elements which are found in oil, bitumen, rocks and sediments and show little or no change in structure from their parent organic molecules in living organisms. Most, but not all, biomarkers are isopentenoids, composed of isoprene sub-units. Biomarkers include pristane, phytane, steranes, triterpanes, porphyrins, and other compounds.”87 This definition references some of the compounds that are most commonly identified as biomarkers. They typically show complex structures that are not particularly stable compared to other possible organic molecules of the same size. Yet, kinetic energy barriers to their destruction are significant and they survive for billions of years if kept at low temperatures in the absence of oxygen.88 They show the result of enzymatic syntheses that assemble molecular structures frequently having a shape and size necessary to perform a specific function in the cell.89 Often this is accomplished at some energy expense for the organism, compared to assembling other less complex lipids of greater thermal stability.
In general, lipids are found as complex mixtures. We can recognize biosignatures in these mixtures when their composition is out of balance with what could be formed from abiotic synthesis alone. Some characteristics of a given biosignature may include the following:
-
Excesses of some structural isomers or homologues over others outside the realm of possible random (abiotic) synthesis;
-
Repeating structural subunits in a molecule, for example, the C5 isopentene subunits in isoprenoids; and
-
Enantiomeric or other stereochemical excesses that do not reflect relative thermal stability.
Structures and Their Significance
The most abundant individual hydrocarbons produced either by abiogenetic processes or by enzymatic synthesis are the normal alkanes. A large component of these are biosynthetically produced as functionalized compounds, such as fatty acids. Diagenetic processes convert them to n-alkanes. Fischer-Tropsch synthesis, an abiotic process, also produces these straight hydrocarbon chains, again mostly with functionality.90Figure 1

FIGURE 1. Mass chromatographs at m/z 85 monitoring for acyclic alkanes from a saturate fraction (a) produced by a FischerTropsch-type synthesis (from T. M. McCollom, G. Ritter, and B. R. T. Simoneit, “Lipid Synthesis Under Hydrothermal Conditions by Fischer-Tropsch-type Reactions, ” Orig. Life Evol. Biosph. 28:1-14, 1998) and (b) of a petroleum sample. The Fischer-Tropsch synthesis, which serves as a model for abiogenic synthesis, gives a smooth distribution of n-alkanes. Petroleum and rock extracts generally show uneven n-alkane distributions, and the biomarkers pristane and phytane are usually present. These features can be recognized as a biosignature.
contrasts the distribution of saturate hydrocarbons obtained by Fischer-Tropsch synthesis with that of a petroleum sample. The smooth distribution of n-alkanes in the Fischer-Tropsch product typifies what would be expected from abiotic synthesis. There is no outstanding excess or lack of any particular homologue. The envelope of distribution of the homologues may be controlled by conditions of the synthesis, such as temperature, pressure, and catalyses. The oil sample shows multiple expressions of biological synthesis and, therefore, a clear biosignature. Relative abundance of several n-alkanes (C25, C28, C30, C34, C38) above the envelope of n-alkane peaks indicates preferential synthesis for their precursors in one or a group of organisms. The predominant even carbon-numbered alkanes in this sample could be from sedimentary reduction of fatty acids that are almost entirely even carbon numbered. The isoprenoids pristane and phytane, classical biomarkers having structures with regularly repeating isopentenyl subunits, show prominently in the mass chromatogram. Branched hydrocarbons that occur between the n-alkanes are in greater abundance than those formed in the Fischer-Tropsch synthesis, and this may be another biological attribute. In fact, simple methyl-branched alkanes where the methyl group shows a positional preference along the carbon chain may be a key biosignature, because of the link between these lipids and primitive organisms.91
Many biomarkers carry taxon specificity in which they identify input from various kingdoms, families, and even individual species. Figure 2 gives examples showing taxon-specific biomarkers from the three domains, Bacteria, Archaea, and Eukarya. Squalene is a widely used isoprenoid intermediate in the biosynthesis of many polycyclic biomarkers, but the cyclization varies in different domains. Bacteria cyclize squalene to make hopanes, while in eukaryotes, squalene lies on the pathway that eventually leads to sterols. Archaea have a unique way of joining isoprenoids to form a “head-to-head ” linkage.
What is known about the chemistry of bacterial lipids suggests there are relatively few themes used by these organisms. As mentioned, many prokaryotes synthesize squalene and cyclize it to hopene. Hopene can be further

FIGURE 2. Syntheses of many lipids that are recognized as biomarkers incorporate isoprene as a repeating subunit. Three domains of extant life, Archea, Bacteria, and Eukarya, carry signature membrane lipids—head-to-head linked isoprenoids, hopanes, and sterols -respectively. Squalene (shown as squalane without double bonds) acts as intermediate for hopane and sterol biosynthesis. Sterols are eventually preserved as steranes in sediments.
extended by adding a five-carbon chain derived from a sugar (ribose) to give bacteriohopane.92 Thus, the hopane molecules, which have been said to comprise the most abundant large-molecule repository of carbon on Earth,93 derive almost entirely from two biomolecular types:hopene (C30) and bacteriohopane (C35). These basic compounds, degraded and rearranged by catalytic and thermal effects of diagenesis and catagenesis, form hundreds of hopanoid products that can be found in sediments.94 Postdepositional degradation alters the biochemistry, but often does not erase it. Many of the products can still be recognized as biomarkers, albeit partially altered ones. Figure 3 shows some of the diagenetically altered hopane structures that can be found among the saturate hydro

FIGURE 3. Examples of saturate biomarkers formed from bacteriohopanetetrol during diagenesis. Bacteriohopanetetrol (C35) and diplopterol (C30, not shown) and closely related compounds may account for most biomarkers of the hopane series. In addition to those shown and other known saturate hopanoids, numerous aromatic analogues are known. Altogether, more that 200 sedimentary hopanoids are known. (A) Reduction of bacteriohopanetetrol under mild diagenetic conditions results in complete preservation of the carbon structure in immature sediments. (B) The commonly encountered pseudohomologous series of saturate hopanoids likely formed by degradation of the side chain and E-ring. (C) Degraded and rearranged hopanes showing the known carbon number ranges. (D) Epimerization in hydrocarbons occurs only at ternary asymmetric carbon atoms. Stereochemical configuration is frozen at quaternary carbon atoms. For C31 hopane, stereochemically frozen quaternary carbon atoms and those tertiary carbon atoms known to epimerize are indicated by * and #, respectively. The known diastereomeric series of C31 hopanes is shown. Hopanes with 17β,21β(H), 22R stereochemistry occur in sediments with a very mild thermal history and are absent in thermally mature sediments. Hopanes with 17α,21β(H) stereochemistry predominate over 17β, 21α(H)-hopanes and 17α, 21β(H), 22S predominates over 17α, 21β(H), 22R in thermally mature sediments (e.g., see Figure 4).
carbons of ancient sediments and oil. Figure 4 shows a typical m/z 191 chromatogram that monitors for hopanes and related compounds and some other terpanes.

FIGURE 4. Partial mass chromatogram of the m/z 191 response for the saturate fraction of an oil from the North Caspian Basin, Kasakhstan, shows peaks indicating biomarkers of microbial and algal origin. Labeled peaks are as follows: Numbers refer to number of carbon atoms in the chemical structures. Ts are tricyclic terpanes (cheilanthanes) possibly related to tasmanites, a single-celled alga; G is gammacerane, attributed to bactivorous ciliates; Ns are neohopanes, hopanes with rearranged structures; D is diahopane, a rearranged hopane; Ms are moretanes; Hs are 17α-hopanes, diastereomers of biosynthetic hopanes (17β ) , with various side chains (see Figure 3) accounting for the range in carbon number 27 to 35. Hopane homologues ≥H31 show stereochemical antipodes at C-22 indicated by R or S. SOURCE: J. M. Moldowan, “Biomarkers,” in McGraw-Hill Yearbook of Science and Technology, McGraw-Hill, Boston, Mass., 2000, pp. 36-39.
In most cases, lipids that have asymmetric carbon atoms (carbon atoms with four different groups attached) are enzymatically biosynthesized as one or the other antipode. For example, amino acids are always biosynthesized as the L enantiomer rather than the D-enantiomer. Some biomarkers show multiple asymmetric centers. However, diagenesis has a tendency to invert certain asymmetric centers. Thus, ancient organic matter that still contains amino acids may tend to show a racemic mixture (50:50 mixture) of L- and D- acids. In the analysis of lipid biomarkers we are seldom concerned with simple enantiomers, because they are difficult to resolve by the available analytical technology. However, compounds with multiple asymmetric centers can be resolved as diastereomers, in cases where some but not all of the asymmetric centers have been inverted. Asymmetric centers carrying a hydrogen atom (tertiary carbon centers) tend to invert, while asymmetric quaternary carbon centers cannot. Inversion at asymmetric centers is called epimerization and does not generally result in a 50:50 mixture of the two configurations (e.g., see Figure 4) when multiple asymmetric centers are involved unless all centers are free to invert. Inversion at quaternary centers during diagenesis is not possible, and disruption of these centers results in molecular rearrangements and loss of the base structure.
Archaea may be even more primitive than bacteria, but less has been published about the structures of their lipids. The head-to-head isoprenoids appear to be Archaea specific.95 These compounds include some with rings and possibly others with shorter carbon chains. There may also be simple branched hydrocarbons that are peculiar to Archaea. Both Archaea and Bacteria are known to produce smaller isoprenoids, like phytane. Thus, acyclic isoprenoids could be useful to indicate even very primitive life.
Recent articles on the analysis of early Earth sediment extracts, revealed that a 2700-million-year old rock from Australia contained a variety of complex biomarkers.96,97 These workers showed the occurrence of phytosynthesis even at those early times, by the presence of 2-methylhopanes, biomarkers common to photosynthetic cyanobacteria. Even more striking was the discovery that a homologous series of steranes, comparable to that found in Proterozoic through Phanerozoic marine sediments, was also present. This strongly suggests the presence of Eukarya (Figure 2) , and indeed, a quicker evolution of Eukarya by as much as a billion years, than had been previously suggested based on fossil evidence.98,99 The quick advancement into cellular organelles on Earth indicated by these studies suggests that analysis for Eukarya-specific biomolecules should not be excluded in the search for extraterrestrial life.
Isotope Ratio Gas Chromatography-Mass Spectrometry
Another research tool that could add important information about biosynthetic origin of compounds would be isotope ratio gas chromatography-mass spectrometry (IRGC-MS). It provides information about carbon isotope fractionation on individual hydrocarbons. In mixtures of highly degraded or thermally altered hydrocarbons it may still be possible to detect excess carbon isotopic fractionation, compared to that which might be available from abiotic synthesis, by using IRGC-MS. Certainly, this would be an added piece of information to consider should sufficient lipid extract be available for the analysis.
ORGANIC CHEMISTRY OF METEORITES: CRITERIA FOR ABIOTIC ORIGINS
Sherwood Chang
SETI Institute
Abstract
The Murchison meteorite contains a rich inventory of organic matter. The distribution of molecular structures and C, H, and N isotopic compositions among the organic components exhibit characteristic abiotic patterns distinctly different from those of terrestrial biochemistry. Together, these patterns provide criteria that can be used with cautious confidence to distinguish between organic matter of biological and nonbiological origins in extraterrestrial samples.
Introduction
Although telescopes may eventually take the search for extraterrestrial life beyond the confines of our solar system, a capability for detecting signs of life on extrasolar planets remains outside our current reach.100 Within our grasp lie Mars and Europa, and for these bodies, sample return missions and robotic experiments loom in the foreseeable future as opportunities for seeking life beyond Earth.
Whether data are obtained at remote sites or extracted from returned samples, the credibility of any conclusion drawn concerning the presence or absence of alien life will depend critically on the ability to establish and apply criteria that distinguish unambiguously the products of biological from nonbiological processes.
A decades-long search for the earliest evidence of life on Earth has produced molecular, isotopic, and stereochemical properties that are useful for assessing the biogenic origins of extraterrestrial organic matter. The value of these traits for discerning alien life hinges on how common they are to all biochemistries;101 the worth of criteria that characterize abiogenic organic matter depends on their absence in all biochemistries.
Why the Organic Matter in Carbonaceous Meteorites?
Carbonaceous meteorites are some of the most studied objects in the solar system, principally because of the presence of condensates from our solar nebula and other stars, primitive bulk elemental compositions, mineral phases formed in liquid water (in CI and CM types), and diverse organic components. The critical connection between liquid water and life on Earth lends special significance to the earliest known occurrence of liquid water and abundant organic compounds in the solar system.102 Apparently, prebiotic evolution occurred early on some parent bodies of carbonaceous meteorites, but fell short of the origin of life.
Samples from carbonaceous meteorites offer a rich and accessible source of information for identifying model characteristics for organic matter that may have formed during prebiotic evolution on Mars, Europa, and other extraterrestrial environments. They also exhibit spectral properties that resemble those of the abundant C-type main belt asteroids.103 The proximity of the asteroid belt to Mars ensures that the planet’s surface contains carbonaceous meteoritic debris and interplanetary dust particles.104 Organic matter, whether prebiotic or biological and asteroidal or martian, would have been best preserved in sedimentary rocks. Such samples are the highestpriority targets for remote analyses or return to Earth.
Molecular Structural Characteristics of Meteoritic Organic Matter
Before the fall of the Murchison meteorite (CM type)in 1969, terrestrial contamination obscured the organic chemistry of carbonaceous meteorites. In preparing to study lunar samples returned that same year, state-of-theart analytical methods designed to minimize terrestrial contamination were developed that ushered in a new generation of meteorite analyses focusing on Murchison samples. Ensuing investigations uncovered an extensive inventory of compounds whose structures, stable isotopic compositions, and stereochemistry reveal a rich organic chemistry.105
The 2 percent total carbon in the Murchison meteorite takes on a variety of forms: The mole fraction of carbon occurring as water-and solvent-soluble organic compounds varies between 0.2 and 0.3 from sample to sample. The bulk of the carbon, ranging in mole fraction from 0.6 to 0.8, resides in a high molecular weight, solvent-and acid-insoluble component whose detailed properties remain poorly understood. This material appears to be a mixture of submicron carbonaceous grains resembling the CHON particles detected in the coma of comet Halley.106 The insoluble fraction also contains minor amounts of extrasolar condensates in the form of nanometersized diamonds, silicon carbide, and graphite grains.107 Carbonates formed during the epoch of liquid water make up the remaining 0.02 to 0.1 mole fraction of carbon.
The classes of meteoritic organic compounds that have familiar biochemical counterparts include amino acids, fatty acids, purines, pyrimidines, and sugars (G. Cooper, personal communication). In addition there are alcohols, aldehydes, amides, amines, mono-and dicarboxylic acids, aliphatic and aromatic nonpolar hydrocarbons, polar hydrocarbons as heterocyclic aromatics, hydroxy acids, ketones, phosphonic and sulfonic acids, sulfides108 and ethers.109 Concentrations of compound classes vary widely from less than 10 (amines) to tens (amino acids)to hundreds of parts per million (carboxylic acids).110 Chromatographic analyses of virtually all classes of acyclic compounds reveal complex molecular assemblages containing homologous series of compounds up to C12 in some cases (carboxylic acids).
Three distinctive patterns of structural variation with molecular weight emerge from studies of these mixtures, no one of which is exhibited by the classes of compounds used in living systems. Data for amino acids and hydroxy acids are chosen to illustrate these patterns, the former for their centrality to terrestrial biochemistry, and both for their wide range of possible structural variations and the availability of relevant meteoritic data. Undoubtedly, future analysts of extraterrestrial organic matter will target amino acids because an alien biochemistry is likely to evolve catalytic polymers consisting of similar multifunctional building blocks.
-
Molecular abundances decrease with increasing carbon number. As illustrated by the α-methyl branchedand straight-chain series of α-amino acids, plots of their concentrations (log nanomoles per gram)versus carbon numbers yield linear correlations with declining slopes of about -7.111 These trends suggest growth of amino acid carbon skeletons by single carbon additions.
-
Abundances of branched-chain isomers exceed those of the straight chain. For example, for compounds containing the same number of carbon atoms, the concentration of the α-methyl-and the β-methyl-branched isomers each surpasses that of the straight-chain isomer.
-
Complete structural diversity prevails at the lower carbon number. This last pattern is illustrated by Table 1, which compares the numbers of known isomers of the acyclic monoamino monocarboxylic acids and the acyclic monohydroxy monocarboxylic acids with the numbers of theoretically possible structures. Overall, 57 amino acids occur among the 159 possible C2 to C7 isomers. Analytical sensitivity and diminishing abundances at higher molecular weights limit further identifications. Remarkably, almost all structural isomers from C2 to C7 occur in the α-amino and α-hydroxy acids; from C2 to C5 nearly all isomers of β -and γ-substituted acids appear. Note that Murchison also contains assemblages of structurally diverse acyclic unsubstituted, and monoamino-and monohydroxy-substituted dicarboxylic acids.112
In contrast, life on Earth employs only 20 common protein α-amino acids and all of these have an α-H. Biochemical counterparts account for less than one-third of the 33 listed in the second column of Table 1. Life on Earth uses a ubiquitous but restricted number of isomers taken predominantly from a few classes of organic compounds to fulfill its requirements for structure and function. Biosynthetic pathways manifest themselves in patterns of structural variation in these classes,113 just as abiotic pathways are reflected in the patterns exhibited by meteoritic compounds (see below).
Structural diversity also occurs among cyclic meteoritic compounds. A plethora of polycyclic aromatic hydrocarbons up to 750 daltons have been found in Murchison samples, along with a multitude of C 15 to C 30 branched alkyl-substituted mono-, di-, and tricyclic alkanes.114 Becker et al. also found an extensive suite of high molecular weight C72 to C270 fullerenes.115 Polycyclic compounds together with the alkyl phosphonic and sulfonic acids comprise the most thermally stable species in the Murchison inventory. They and the acid-insoluble material are the likeliest meteoritic components to survive delivery to and diagenesis on Mars.
TABLE 1. Comparison of Numbers of Acyclic Monoamino Monocarboxylic Acids and Monohydroxy (shown in bold) Monocarboxylic Acids of the Murchison Meteorite with the Number of Possible Theoretical Isomers (shown in bold italic) (compounds with other substituent groups are omitted). (After J. R. Cronin and S. Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite,” in The Chemistry of Life’s Origins, J. M. Greenberg, C. X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209-258.)
|
C Atoms |
α |
β |
γ |
δ |
ε |
ζ |
Unknown |
|
2 |
1, 1, 1 |
— |
— |
— |
— |
— |
— |
|
3 |
1, 1, 1 |
1, 1, 1 |
— |
— |
— |
— |
— |
|
4 |
2, 2, 2 |
2, 2, 2 |
1, 1, 1 |
— |
— |
— |
— |
|
5 |
3, 3, 3 |
6, 6, 3 |
3, 3, 3 |
1, 1, 0 |
— |
— |
1 |
|
6 |
8, 8, 8 |
12, 3, 1 |
11, 4, 0 |
4, 2, 0 |
1, 1, 0 |
2 |
|
|
7 |
18, 18, 12 |
29, 0, 0 |
29, 0, 0 |
20, 0, 0 |
5, 0, 0 |
1, 0, 0 |
2 |
 |
|||||||
The observed patterns of variation in molecular structure and abundance with increasing carbon number suggest synthesis routes entailing small free-radical initiators and intermediates.116 Such pathways tend to produce all possible structural isomers at lower carbon numbers by more or less random synthesis. Primary reactions in interstellar clouds produced mixtures of nitriles and other compounds. When exposed to liquid water on the parent body, the nitriles were converted to various substituted and unsubstituted carboxylic acids, including the amino acids mentioned earlier. Other classes of compounds were likely to have formed in secondary reactions. For example, the Strecker synthesis of cyanohydrins and aminonitriles from HCN, aldehydes or ketones, and ammonia or amines, followed by hydrolysis, could account for synthesis of some of the α-amino-and α-hydroxy acids. Amino acids substituted at more distant positions from the carboxyl group require other synthetic pathways.
Chirality of Amino Acids in the Murchison Meteorite
Syntheses of organic compounds do not produce chiral products in the absence of a chiral agent, whether it be an energy source (e.g., circularly polarized light), a surface substrate (e.g., quartz crystals), or a reactant. A recent evaluation of abiotic mechanisms proposed for the origin of chiral molecules on the primitive Earth concluded that they are “inapplicable and implausible.”117 The absence of chirality in products of prebiotic evolution experiments strengthened the presumption that natural abiotic synthesis invariably produces racemic compounds. Indeed early analysts of Murchison amino acids declared them racemic.118 Under the looming specter of contamination, later reports of L-enantiomeric excesses in several meteoritic counterparts of biological amino acids generated a swirl of controversy.119-121 In 1997, Cronin and Pizzarello resolved the issue in favor of modest L-enantiomeric excesses of 2 to 9 percent.122 They avoided the pitfalls of contamination by making measurements on 2-amino2,3-dimethylpentanoic acid, α-methylnorvaline, and isovaline. All three compounds are α-methyl substituted; the first two have no known biological counterparts, and the third has a restricted distribution in fungal antibiotics. Engel and Macko have found evidence for nonracemic Murchison amino acids using an isotopic approach.123
Bailey et al. suggest that the observed enantiomeric excesses could have been induced by circularly polarized light arising from dust scattering in regions of high-mass star formation.124 These sources occur more widely than do the supernova remnants or pulsars that were first proposed by Rubenstein et al. as sources of circularly polarized synchrotron radiation.125 Regardless of production mechanism, the nonracemic amino acids testify to the reality of a naturally occurring abiotic chiral process. Future researchers will need to be cautious in using chirality to assign biotic origins to extraterrestrial compounds.
Isotopic Compositions of Carbon, Hydrogen, and Nitrogen in Murchison Organic Matter
Although bulk analyses of the isotopic compositions of C, H, and N of the Murchison meteorite hold no surprises, detailed examination of organic components reveals anomalies that call for unusual production mechanisms and environments of origin. The organic matter generally contains deuterium enrichments that start above the upper limits of the terrestrial range (δD +100 ‰) and extend to +2500 ‰, approaching those of interstellar molecules.126 Values of δ13C range from −18 to +40. Results are displayed in Figure 1 for organic components on which simultaneous measurements of δ13C and δD are available.
The heavy isotope enrichments in C and H vary widely from component to component and extend well beyond the limits of terrestrial experience. This is also true for δ13N values, which range from +18 to more than +98 ‰. 127 Note that the strongest heavy-isotope enrichments occur in polar organic compounds. Variations are large even within compound classes. Such variability suggests different reaction pathways or different formation

FIGURE 1. C and H isotopic composition of organic components in Murchison meteorite. SOURCE: J.R. Cronin and S. Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite,” in The Chemistry of Life’s Origins, J.M. Greenberg, C.X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209-258, and G.W. Cooper, M.H. Thiemens, T.L. Jackson, and S. Chang, “Sulfur and Hydrogen Isotope Anomalies in Meteorite Sulfonic Acids,” Science 277:1072-1074, 1997.
temperatures, sampling of isotopically dissimilar reactant reservoirs, or some combination of these and other factors. Temperature effects accord with the view that the deuterium enrichments arise from gas phase (ionmolecule) and grain surface (atomic or free radical) reactions occurring at low temperatures in interstellar clouds.128 Sampling of isotopic reservoirs are consistent with the presence in Murchison of originally interstellar molecules and stellar condensates containing distinctive nucleosynthetic isotopic compositions.129
In sharp contrast, biochemical processes typically do not yield such wide isotopic variability within or among compound classes, nor do the products of laboratory prebiotic syntheses.130,131 The isotopic record of fossil life preserved in sediments of increasing age generally shows a relatively narrow range of carbon isotopic compositions for both extractable132 and insoluble organic fractions.133,134 The 20-35‰ difference ((∆δ ) between the organic carbon and the isotopically heavier coeval carbonate (representing the source reservoir) is generally taken as an isotopic biomarker. Murchison organics preserved in ancient sediments on Mars, however, might well yield a similarly narrow range of compositions, as diagenesis destroys the minor polar components leaving the thermally stable insoluble carbon and hydrocarbons as relics. The δ13C of carbonate in Murchison is about +42‰; therefore, the ∆δ could range from +20 to +60 ‰, overlapping the terrestrial biomarker values. As with chirality, isotopic criteria must be used cautiously for distinguishing biological from nonbiological materials, particularly in an Earth-like environment with a single dominant isotopic reservoir for each of C, H, and N.
Compounds extracted from returned martian samples can be subjected to intermolecular and intramolecular isotopic analyses.Measurements on individual Murchison compounds show each carboxylic acid containing ~16 ‰ more 13C than the light hydrocarbon of corresponding carbon number..In both homologous series, δ13C values decline in parallel fashion as carbon number increases.135 Intramolecular measurements on acetic acid indicate that the carboxyl group is relatively enriched in 13C by 15 to 30 ‰. These results suggest synthesis of carboxylic acids by stepwise addition of the elements of CO2 to hydrocarbons. Cooper et al. also carried out intramolecular carbon, hydrogen, and sulfur isotopic measurements on individual Murchison sulfonic acids.136 Measurements at similar levels of molecular detail may prove invaluable in establishing sources and production mechanisms; they should certainly be included in the isotopic arsenal for future studies of extraterrestrial organic matter.
Organic Survey Analysis of the Murchison Meteorite
Survey analyses for volatile and organic components will be among the key preliminary measurements carried out on rocks returned from Mars.One such technique was used successfully in critical experiments on the 1976 Viking lander. Martian soil samples were progressively heated under vacuum to ~500°C followed by product analysis with a combined gas chromatograph-mass spectrometer (GC-MS).137 The measured ion abundances set a 4-part-per-billion upper limit on organic matter in martian soil, which argued persuasively for the absence of detectable signs of life at the Viking landing sites.
In a similar experiment, the data shown in Figure 2 were produced from mass spectra recorded every 17 seconds as a sample of a Murchison-like meteorite was heated under vacuum from 25 to 1400°C. The mass pyrograms show gas evolution in the temperature ranges over which water-bearing clays, carbonates, and sulfates release their H2O (250 to 600°C) , CO2 (500-55°C) , and SO2 (400-800°C). Dehydration, decarboxylation, and oxidative desulfurization of organic compounds also yield these same species over the range <200 to 600°C. Hydrocarbons and N-and O-containing organic compounds evolve from 100 to ~700°C due to pyrolysis of parent organic matter of differing thermal stabilities. The CO2, CO, and N2 released above ~600°C represent refractory carbonaceous material that has already lost its alkyl and functional groups and is undergoing oxidation by inorganic silicates and oxides. While this hardy material would have been the most likely organic component to have survived on the martian surface, it would not have been detected by the Viking pyrolysis GC-MS with its 500°C temperature limitation.
Survey analyses of this sort, while incapable alone of addressing the origin of organic compounds, would provide means for detecting organic matter and identifying sedimentary rock samples either in terrestrial laboratories or on planetary surfaces. Such screening would be invaluable for identifying samples for return to Earth or for determining the relative abundances of rock types at landing sites. The analytical methodology is well established,

FIGURE 2. High-resolution mass pyrograms for the Jodzie (CM) inclusion. SOURCE: T.E. Bunch, S.Chang, U. Frick, J.M. Neil, and G. Moreland, “Carbonaceous Chondrites: I—Characterization and Significance of Carbonaceous Chondrite (CM) Xenoliths in the Jodzie Howardite,” Geochim. Cosmochim. Acta 43:1727-1742, 1979, Figure 6.
and it could be readily adapted to modern instrumentation for robotic exploration. A pyrolysis-GC-isotope ratio however, could have potential for life detection.
Conclusions
Analyses of Murchison organic matter reveal characteristic patterns of variation in molecular composition. These include the following:
-
Complete structural diversity within compound classes among isomers of the lower carbon homologues;
-
Molecular abundances in a class declining with increasing carbon number;
-
Branched-chain isomers predominating at a given carbon number; and
-
C, H, and N isotopic compositions varying widely among and within compound classes.
Since these attributes may not be held in common by all abiotic reservoirs of extraterrestrial organic matter, it is unwarranted to view the observed traits as universal abiotic indicators. Molecular assemblages of biological origin, however, will rarely if ever exhibit these patterns. Therefore, these criteria may be used with cautious confidence to distinguish nonbiological from biological organic matter. The discovery of L enantiomeric excesses of abiotic origin in some Murchison amino acids means that chirality is not a foolproof molecular indicator of biology.
ISOTOPES OF IRON: BIOMARKER PROSPECTS
Ariel D. Anbar
Department of Earth and Environmental Sciences
University of Rochester
Abstract
The isotopes of iron are fractionated during microbially mediated reduction of ferrihydrite. The isotopic composition of iron in modern and ancient ferromanganese sediments displays a range of values similar to those produced biologically, which could reflect biological production of iron in nature. Because of the ubiquity of ironbearing phases in nature and the diverse uses of iron in microbial metabolism, these observations demonstrate the potential of iron isotope research for the detection of ancient life. Nonbiological chemical processes can similarly fractionate Fe isotopes, but the importance of nonbiological fractionation in nature has yet to be determined.
Introduction
Isotopic indicators are extremely useful in the detection of life in the geologic record. Compared to both molecular fossils and microfossils, isotopic indicators can be more robust against alteration processes. A notable example is the observation of 12C-enriched organic residues in strongly metamorphosed sediments deposited >3.85 billion years ago.138 This constitutes the earliest evidence of life on Earth.
Study of isotope fractionation of elements lighter than S has been ongoing for nearly 50 years. In contrast, until recently there was virtually no research into the fractionation of Fe and other transition metal isotopes— biogenic or otherwise. Such research was hampered by difficulties in making precise measurements of the isotopic compositions of these elements.
Recent advances in mass spectrometric tools and techniques have greatly simplified such measurements, leading to rapid progress. Natural variations in the isotopic compositions of Fe (masses 54, 56, 57, 58), Cu (63, 65), Zn (64, 66, 67, 68, 70), Tl (203, 205), and Mo (92, 94, 95, 96, 97, 98, 100) have been reported.139–145 Because these isotopes are neither radioactive nor the products of long-lived radioactive decay, these variations must result from mass-dependent fractionation of metal isotopes. For some of these metals, biological and nonbiological fractionation processes have been studied in the laboratory.146–150
Isotopes of Fe are of particular interest because Fe-bearing phases, including well-known biominerals, are widespread in nature and are more robust against alteration than many C-bearing materials, and because biological fractionation of Fe isotopes has been reported by Beard et al.151 Hence, biogenic Fe isotope signatures could prove useful for detection of life in the geological records of Earth or Mars.
Research on the Fe isotope system is advancing rapidly. This paper reviews the basis for interest in Fe isotopes and the status of Fe isotopes as biomarkers.
Why Iron Isotopes?
When considering the utility of Fe (or any other metal) for isotopic biomarker studies, four key questions must be answered:
-
Is the isotopic mass difference large enough to potentially produce a detectable fractionation?
-
Does the biological use of Fe justify attention?
-
Do biogenic fractionations exist?
-
Are biogenic fractionations larger than those resulting from natural nonbiological processes?
The first two questions, which should be addressed prior to isotopic study, are discussed below. The latter two, the subjects of ongoing research, are addressed in the following section.
Fractionation Potential
To answer the first question, it is useful to understand why mass-dependent fractionation occurs at all. Briefly, isotope fractionation results from differences in the zero-point energies (ZPEs) between chemical bonds that are identical except for isotopic substitution. In the simple case of a diatomic molecule, A-B, ZPE =1/2 hν, where ν is the vibrational frequency of the bond and h is Planck’s constant. Approximating the bond as a harmonic oscillator, ν =1/2π (k/μ), where k is the force constant and μ is the reduced mass of the bond (μ = mAmB/(mA + mB)). Hence, bond strengths are mass dependent. Because bond strength is inversely related to ZPE, bond strength increases upon substitution of heavier isotopes. Thus, if we consider two isotopes of A, A1 and A2, where mA1 >mA2 ,bond A1-B is stronger than A2-B.
This mass dependence leads to differences in reaction rate constants: A1-B will react more slowly than A2-B. This is the origin of “kinetic” isotope effects in the case of unidirectional or incomplete reactions. An example is the fractionation of 12C from 13C during photosynthesis, where 12CO2 is processed faster than 13CO2. Kinetic isotope effects favor reaction of the lighter isotope.
Mass dependence also leads to differences in equilibrium constants and, hence,“equilibrium” isotope effects. Such effects favor partitioning of the heavier isotope into the stronger bonding environment. Although complete equilibrium is often not achieved, especially in biological systems, such free-energy differences can drive isotope fractionation in nature. An example is the fractionation of C isotopes between CO2 gas and the carbonate ion (CO32–).
A rough guide in assessing the relative potential for fractionation of different elements can be made by considering the simplified case in which mB >>mA. Here, ZPEA1-B/ZPEA 2 -B ~ (mA 2 /mA1). The relative values of (mA2 /mA1) for a variety of biologically important elements are shown in Table 1. It is immediately apparent that while the values for transition metals are generally smaller, they are of comparable magnitude to values for other elements. Hence, it is reasonable to expect that transition metals, including Fe, could show natural variations in isotopic composition similar to those of lighter isotopes of order 1 to 10 parts per thousand (1 to 10 ‰).
Table 1
Some Elements of Biological Interest
|
Stable Isotopesa |
ν(mA2 /mA1 ) |
|
|
C |
12C, 13C |
1.041 |
|
N |
14N, 15N |
1.035 |
|
S |
32S, 34S |
1.031 |
|
Ca |
40Ca, 44Ca |
1.049 |
|
Se |
76Se 78Se, 80Se, 82Se |
1.039 |
|
Fe |
54Fe, 56Fe, 57Fe |
1.027 |
|
Ni |
58Ni, 60Ni, 61Ni, 62Ni |
1.034 |
|
Cu |
63Cu, 65Cu |
1.016 |
|
Zn |
64Zn, 66Zn, 67Zn, 68Zn |
1.031 |
|
Mo |
92Mo, 94Mo 98Mo, 100Mo |
1.043 |
|
a Isotopes with average abundance >1 %. A2 is the heaviest and A1 the lightest isotope. |
||
However, it must be emphasized that for each element, the extent of isotope fractionation depends on the chemical reactions in which the element is involved. For example, while C and Ca compare favorably in Table 1, the natural variability of 40Ca/44Ca is about one-tenth that of 12C/13C. This reflects differences in the environmental biogeochemistry of these elements. Therefore, to assess the relative utility of Fe for biomarker studies, its use in biology must be considered.
Iron in Biology: Present and Past
From a strictly biological perspective, Fe is of interest because it has the most diverse usage of any metal in biochemistry. Briefly, Fe is found in cofactors and enzymes such as porphyrins, which are ubiquitous in electrontransfer cytochromes; in enzymes that bind CO, O2, and NO; and in various oxidases; Fe-S proteins, such as ferredoxin, a component in the electron transfer chain of Photosystem I and the Fe4-S4 cofactors in nitrogenase; methylreductase; and Fe-based superoxide dismutase, present in prokaryotes, chloroplasts, and mitochondria.
Most interesting is the fact that Fe, like Mn, can be used as an oxygen substitute in respiration.152 Like C, it can also serve as an electron donor, providing metabolic energy to Fe-oxidizing microbes under both aerobic and anaerobic conditions. Because there is only one stable isotope of Mn, Fe is the only metal involved in this manner in microbial respiration that also has the potential for detectable fractionation effects. Hence, as with C, there is potential for kinetic fractionation of Fe isotopes during enzyme-catalyzed unidirectional reactions in Fe metabolism. The signature of such processes could be found in the isotopic composition of Fe minerals formed from biologically processed iron.
In addition, the uptake of Fe by microorganisms involves conversion from inorganic complexes to extremely strong organic complexes with Fe-specific biogenic ligands (“siderophores”). Hence, there is potential for both kinetic and equilibrium fractionation of Fe isotopes during Fe uptake. Such uptake can be extremely efficient and, because of mass balance considerations, leads to the possibility that biogenic Fe isotope signatures might also be found in residual Fe not taken up by organisms.
Fe also stands out compared to other metals because, from an evolutionary perspective, it was probably particularly important for early life. For example, in the anoxic environment of the Archaean Earth, the availability of Fe may have led to the widespread use of Fe2+ as an energy source for carbon fixation. Both anaerobic Fe oxidation and Fe reduction may have been important in the formation of banded iron formations (BIF).153 The deepest organisms on the phylogenetic tree contain various Fe-bearing proteins. Hence, it is all but certain that the uptake and use of Fe was an early invention of life on Earth. It is likely that biological Fe utilization would also have been an early invention of life on Mars, if life existed there.
Therefore, it is reasonable to expect that microbial processing of Fe generates isotopic signatures that will prove useful for detection of ancient life. This expectation has motivated Fe isotope research.
Recent Research Findings
To test this expectation, three complementary research approaches have been pursued by several research groups:
-
Determination of the extent of Fe isotope variability in nature;
-
Study of Fe isotope fractionation processes in laboratory model systems; and
-
Theoretical modeling of the magnitude of Fe isotope fractionation.
The results of this research are summarized below and in Figure 1. In this discussion, variations in the isotopic composition of Fe are presented using the “δ” notation:
δ56 Fesample = [(56Fe/54Fe)sample/(56Fe/54Fe)standard – 1 ] × 1000‰.
Natural Variations
Variations in the isotopic composition of Fe have been determined in a range of terrestrial and lunar igneous rocks, iron meteorites, and loesses, as well as in ancient and recent sediments.154,155These measurements provide unequivocal evidence that the isotopic composition of Fe varies in nature.
Beard and Johnson first observed that δ56Fe in marine sediments is more variable than in igneous rocks.156 Fe isolated from Pacific and Atlantic ferromanganese nodules is shifted by ~ –1.5 ‰ compared to the range for

FIGURE 1. Summary of all δ56Fe data in refereed and unrefereed literature as of April 2000. See text for details and sources.
igneous rocks, while Fe isolated from the light-colored, Fe-poor layer of a Proterozoic BIF is shifted by ~ +1 ‰ compared to the darker, Fe-rich layers. By comparison, Fe in igneous rocks varies over a range of <± 0.3 ‰. This contrast strongly suggests that Fe isotopes are fractionated during (bio)geochemical processing of Fe in the environment. Because microbial activity is believed to play an important role in such processing, this observation is at least consistent with the existence of biogenic fractionation effects.
Zhu et al. carried out a detailed examination of the isotopic composition of Fe in a well-characterized Atlantic ferromanganese crust.157 They found that δ56Fe varies over a range of ~1‰, with an average value of ~ –1‰ compared to a laboratory Fe standard, iron meteorites, and loess (note that Zhu et al. report their data as ε57 Fesample =[(57Fe/54Fe)sample/(57Fe/54Fe)standard – 1 ] × 104 . These values have been translated to δ56Fe for comparison here. Assuming mass-dependent fractionation of Fe isotopes, δ56Fe ~0.067 ×ε 57Fe).
Direct comparison of these two studies is slightly problematic because different standard materials were used, and there has been, as yet, no interlaboratory calibration. However, the total range of values reported by Zhu et al. for iron meteorites, loesses, and paleosols is only ~0.3 ‰.158 This is similar to the range for igneous rocks reported by Beard and Johnson.159 Hence, both data sets reveal that the greatest variability of δ56Fe is in ferromanganese sediments and that δ56Fe in recent marine sediments is shifted by ~ –1‰ compared to all other Fe ((with the exception of Proterozoic BIF).
The isotopic composition of Pb (206Pb/204Pb and 208Pb/204Pb) in the ferromanganese sediments is also reported by Zhu et al.160 The Pb isotopic system differs from the Fe system in that the 206Pb and 208Pb are the products of
U radioactive decay. Nevertheless, correlated, secular trends are observed in δ56Fe and the Pb ratios. Because mass-dependent fractionation of Pb isotopes is expected to be minuscule, this correlation argues against in situ Fe isotope fractionation in the sediments. Instead, the secular variations in δ56Fe apparently reflect correlated changes in the sources of Fe and Pb to the sediments. However, it must be stressed that this conclusion in no way invalidates the observation that Fe in these sediments is isotopically fractionated from other Fe and provides no direct information about fractionation processes.
Laboratory Studies—Biological
Substantial laboratory effort has been devoted to biological fractionation processes. The most extensive published biological studies, performed by Beard et al.,161 focused on Fe isotope fractionation during microbially mediated reduction of ferrihydrite (an Fe(III) oxyhydroxide precipitate) by Shewanella algae. S. algae is a mesophilic, dissimilatory Fe-reducing organism that uses Fe(III) as an electron acceptor during respiration. Hence, this organism is a good candidate for Fe isotope fractionation research.
In a series of experiments conducted under carefully controlled conditions, it was clearly demonstrated that the initial δ56Fe of the dissolved Fe(II) produced by S. algae is shifted ~ –1.3‰ compared to the initial isotopic composition of the ferrihydrite substrate. These data provide evidence that Fe isotopes are fractionated during microbially mediated reduction of ferrihydrite.
The fractionation mechanism is not yet clear. However, the observation that the Fe(II) product is isotopically lighter than the ferrihydrite reactant strongly suggests preferential uptake and/or reduction of 54Fe versus 56Fe, consistent with a kinetic isotope effect. Such a kinetic effect could occur during extraction of Fe(III) from the ferrihydrite or during enzyme-catalyzed reduction.
Consistent results have been reported for other Fe-reducing bacteria grown on ferrihydrite substrate.162 Brantley et al. have presented evidence that Fe leached from the mineral hornblende in the presence of organic ligands (including siderophores) is fractionated by ~ –1‰ compared to the mineral-bound Fe.163 This may provide insight into the mechanism of biogenic fractionation, suggesting that fractionation occurs during uptake of Fe rather than during reduction.
In contrast to these studies, Mandernack et al. found no evidence of fractionation of similar magnitude in intracellular magnetite produced by magnetotactic bacteria.164 It is not yet known whether this reflects basic differences in Fe metabolism or the sensitivity of fractionation to growth conditions. Regardless, this finding reveals that biogenic Fe is not necessarily fractionated with respect to Fe sources.
Laboratory Studies—Nonbiological
Fractionation of Fe isotopes by chemical processes in the absence of biology was demonstrated by Anbar et al.165 during elution of Fe in HCl media from small (~4 cm) columns packed with anion-exchange resin. The range of values observed in the elution fractions (~6‰)was larger than seen in biological experiments to date. Integrated over all fractions (which accounted for 100 percent of the loaded Fe), δ56Fe = 0, as expected from mass balance. This study provides unequivocal evidence that Fe isotopes can be fractionated by nonbiological chemical processes.
It was proposed that this fractionation results from equilibrium isotope exchange between dissolved FeCl4–, which binds to the resin,and cationic and neutral Fe-Cl complexes (FeCl3 and FeCl2 +). The former is a tetrahedral complex, while the latter are octahedral,providing a difference in bonding environment that could drive such fractionation. This is not a unique interpretation; it is possible that kinetic effects could also explain the data.
Because this particular ion exchange system is not representative of chemical processes in the environment, experiments with other nonbiological systems are urgently needed. As yet,there are no such studies in the peerreviewed literature.
Bullen et al.have presented data from controlled experiments in which dissolved Fe(II) is slowly oxidized by addition of O2 at constant pH and precipitated as ferrihydrite.166 In these experiments, δ56Fe of the initial ferrihydrite was fractionated ~1‰ compared to the dissolved Fe. As expected from mass balance, as the fraction
of Fe precipitated approaches unity, δ56Fe of the accumulated precipitate approaches 0‰, while the residual dissolved Fe approaches ~ –1‰.
Theoretical Studies
The magnitude of equilibrium fractionation between simple complexes can be predicted from experimentally determined vibrational frequencies. Such data are not always available for transition-metal compounds of interest in biomarker applications (e.g., common inorganic and organic complexes), and application to minerals is less straightforward. However, Schauble et al.167 have predicted equilibrium fractionation of 1 to 10‰ between dissolved Fe complexes. Using a novel approach based on Mossbauer data, Polyakov and Mineev predict equilibrium fractionation of 1 to 10‰ between common Fe-bearing minerals.168However, these predictions have not yet been tested against measurements.
As yet there have been no theoretical studies of kinetic effects applied to Fe isotopes.
Synthesis
The evidence accumulated to date (Figure 1) demonstrates that the isotopic composition of Fe in ancient and modern Fe-rich sediments varies over a range of ~2.5‰. This variation is nearly 10 times larger than that of all other natural materials analyzed. Laboratory studies indicate that fractionations of per-mil magnitude can be produced by dissimilatory Fe-reducing bacteria and by leaching of Fe from minerals by biogenic ligands.
These observations demonstrate the potential of Fe isotope measurements for the detection of ancient biological activity. As an example, Beard et al. suggest that the value δ56Fe ~ +1‰ in the light-colored, Fe-poor layer of a Proterozoic BIF reflects biological conversion of insoluble Fe(III) oxyhydroxides to soluble Fe(II) by Fereducing bacteria.169 Such a process would be expected to preferentially remove 54Fe, producing a residue with δ56Fe >0.
While such applications of the Fe isotope system are credible, they should be approached with some caution. First, as pointed out by Zhu et al.,170 fractionation in sediments does not necessarily reflect in situ processes. This is an important caveat that may invalidate some interpretations of sedimentary δ56Fe variations. However, it must be recognized that this point has no direct bearing on whether natural δ56Fe variations are of biological or nonbiological origin. For example, given the results of Brantley et al.,171 it is easy to envision biogenic Fe isotope fractionation during chemical weathering or during dissolution of aeolian material in seawater, which would deliver already-fractionated Fe to the sediments.
Second, and more importantly, it has not yet been demonstrated that nonbiological processes are incapable of producing comparable fractionations in nature. Clearly,chemical fractionation of Fe isotopes in the absence of biology is possible; the entire range of observed natural variations has been readily produced in a contrived laboratory system.172 The preliminary data of Bullen et al. suggest that nonbiological processes could be particularly important in ferromanganese sediments.173 However, many more such studies, covering a wider range of phases and reactions, are obviously needed.
The ultimate impact of such studies on biomarker applications is unclear. However, regardless of the outcome, it is very likely that the Fe isotope system will prove to be useful in biogeochemical research. Similar debates animated the development of both the C and the S isotope systems. In the case of C isotopes, it is now widely accepted that nonbiological processes typically occurring at Earth’s surface cannot easily reproduce the magnitude of fractionation produced by photosynthesis. Hence, C isotopes are readily employed for detection of ancient life. On the other hand, in the case of S isotopes, such a simple distinction between biological and nonbiological fractionation magnitudes is not possible. Nevertheless, S isotope studies have been profoundly useful in biogeochemical research,in both modern and ancient environments. Research into Fe and other metal isotope systems is likely to be at least as useful as S isotope research in the study of ancient environments and life, and may yet prove as useful as C.
DEVELOPMENT OF IN SITU MEASUREMENT TECHNIQUES FOR DETECTING THE CHEMICAL SIGNATURES OF LIFE
Wesley T. Huntress, Jr.
Geophysical Laboratory
Carnegie Institution of Washington
Abstract
A “Grand Challenge” Research Program has been instituted at Caltech’s Jet Propulsion Laboratory (JPL) to develop in situ measurement techniques that are capable of detecting the chemical signatures of life. The JPLCaltech concept of a Grand Challenge, the events leading to adoption of the current theme, and the participating investigators and their research goals are described.
A Short History of JPL Grand Challenges
A Grand Challenge is a Caltech-JPL institutional investment in a long-term research and development program to enable exciting new missions in space science and exploration. They are generally intended to develop the science and technologies that will enable accomplishments in a new realm of exploration well beyond the current state of the art. Grand Challenges set long-term interdisciplinary research goals, combining science and technology objectives, to try and accomplish something never done before. They should be doable in 10 to 15 years, progress must be measurable, and it must be clear when the program is finished. Grand Challenges are long-term Caltech investments in major technological innovations that will lead to the establishment of a new NASA program with a major impact on the future of space exploration. These new NASA programs would replace or extend current investments in the JPL mission in space.
The Grand Challenge concept is also a means to provide stable funding at the investigator level independent of the annual and often large fluctuations in NASA R&D funding commitments. NASA R&D funding in any one field and at any one center is subject to fierce competition among a large number of constituencies including other NASA centers, universities, NASA Headquarters, the Administration, and Congress—which leads to a long history of large fluctuations that seriously hinder progress toward any particular long-term goal.
There have been two previous JPL Grand Challenges. The first was “Detection of Planets Around Stars” (1984 to 1996). This program was triggered by an image of the dust disk around β Pictoris taken in 1981. The program tackled this difficult problem by investing in the development of near-infrared optical interferometery. A 100-meter interferometer using 1-meter optics was first built at Palomar to develop the optical component technology, followed by investment to scale the technology up to the Keck twin 10-meter telescopes. These investments have enabled the future Space Interferometer Mission (SIM), which has been funded by NASA, and a larger Terrestrial Planet Finder mission scheduled after SIM. The research conducted in this Grand Challenge was key to enabling the NASA Origins Program, and the JPL program was terminated with NASA establishment of the Origins Program.
The second JPL Grand Challenge to be established was “In-Situ Planetary Exploration and Robotics” ((1990). This was triggered after every planet in the solar system (except Pluto) had been visited by at least one spacecraft. The event was the Voyager flyby of Neptune at the edge of the solar system in 1989. At that time, there also were three orbiters in development—Magellan to Venus, Galileo to Jupiter, and Cassini to Saturn—so that by 2005, orbiters would have explored the planets from Venus to Saturn. The next stage after flybys and orbiters was exploration of planetary surfaces, so this Grand Challenge was instituted to develop the technologies and science instruments for surface exploration. This Grand Challenge enabled the Sojourner rover for Mars Pathfinder and is still under way in JPL’s Center for In-Situ Instrumentation.
Selection and Management of the New JPL Grand Challenge
Three potential topics for the third Grand Challenge emerged from an extensive consensus-building process at JPL and the Caltech campus:
-
In situ detection of extraterrestrial life. This topic was triggered by discoveries in biology, planetary exploration, and astronomy that have led to the impression that life might be more commonplace in the universe than previously expected. The topic is also perfectly complementary to the ongoing Grand Challenge in situ planetary exploration and robotics.
-
Interstellar travel and exploration. This topic was triggered by discoveries of extrasolar planets and the prospect for learning more about them. It suffered a real disadvantage because of the immense technological challenge posed to propulsion and spacecraft reliability.
-
Robotic colonies in space This topic is also synergistic with the current new phase of planetary surface exploration. It envisioned a new paradigm for robotic planetary exploration, carried out by a semi-intelligent, selftasking, and self-servicing robotic infrastructure on a planetary surface operating only on high-order goals from humans on Earth. These robotic colonies were seen as a preface to eventual human arrival and the establishment of a joint human-robotic outpost.
After selection of topic 1 by the JPL director in January 1999, an external expert review was called to assess the prospects for this challenge, and recommendations were forwarded to JPL’s director in April of 1999. Caltech and JPL established an institutional funding line at about $1 million per year, with a funding schedule in two phases. The first 18-month phase was for concept development, followed by a second phase of about 4 years for development of a consensus measurement strategy and instruments. Phase I investigations were solicited in May from the Caltech-JPL community and reviewed by an External Review Panel composed of the first four members of what became the External Review Board:
|
Wesley Huntress (chair) |
Carnegie Institution of Washington |
|
James Ferris |
Rensselaer Polytechnic Institute |
|
Harold Morowitz |
George Mason University |
|
Steven Squyres |
Cornell University |
|
Kathy Olsen |
NASA Headquarters, Chief Scientist |
|
William Schopf |
University of California, Los Angeles |
|
Norman Pace |
University of Colorado |
Recommendations for selection and funding were made to JPL’s chief scientist and director in September. It is expected that after results are examined from the first phase, the second phase will call for new proposals and could result in program restructuring. Funding was received by the investigators in November 1999.
The Phase I solicitation required that the principal investigator be from JPL, but also required teaming with outside investigators at other academic institutions and strongly encouraged Caltech campus participation. The solicitation also asked that each team have the capability to carry out research on three objectives:
-
Identifying the chemical signatures of life. Define the most likely non-Earth-centric signatures; universal properties for living systems, not dependent on specific molecules or peculiar properties of life on Earth. Define a strategy for employing them in a systematic search for extraterrestrial life.
-
Developing measurement techniques to detect these signatures. Develop measurement techniques and statistical strategies for identifying these signatures in the background of nonlife. Test these methods on Earth samples with prepared controls.
-
Conceptualizing the possibilities for miniaturizing in situ instrumentation. Devise concepts for how the identified measurement techniques can be implemented in-situ on a remote planetary surface. This task should lead to proposals for the development of miniaturized in situ instruments.
The response, however, did not meet this objective. The proposals received generally fell into four categories:
-
Category I—addressed all three goals: proposed to define chemical signatures, identify required measurements, and conceptualize potential miniaturization schemes;
-
Category II—adopted a particular chemical property or process as a signature of life and proposed a series of predefined measurements to identify the property or process;
-
Category III—submitted by instrument scientists offering their technique as a de facto signature and proposing to develop their instrument for measuring in situ samples; and
-
Category IV—unresponsive.
There was only one proposal in Category I; there were three in Category II, twelve in Category III, and four in Category IV. This distribution should have been expected from a science community at JPL heavy in instrument scientists. The selected proposals are the following:
-
Category I: Signature Definition Studies
Conrad Biological Signature Definition
-
Category II: Specific Signatures with Defined Multiple Measurements
Beauchamp Chirality—mass spectroscopy/chromatography/electrophoresis/chirooptical
Mojarradi Metabolism—e-transport/porphyrins/heat production/DNA
Grunthaner Diagenesis—XPS/mass spectroscopy/microfluidics
-
Category III: Instruments Proposed as Individual Signature Detectors
Webster Laser spectroscopy (and mass spectroscopy) isotope ratios
Kanik Ion mobility spectrometer
Bearman Chemical imaging
Sinha Fe isotope mass spectroscopy
George X-ray fluorescence.
JPL has uncharacteristically chosen a nonresident scientist to manage this Grand Challenge Program,Wesley Huntress of the Geophysical Laboratory, Carnegie Institution of Washington; the local project scientist is Kenneth Nealson of the Jet Propulsion Laboratory. The role of the project manager is to bring the Grand Challenge Research Program to a successful conclusion, where success is defined as a credible, consensus in situ measurement strategy for the detection of life. He will also conduct external scientific and technical reviews during Phase I on 6-month centers. The role of the project scientist is to provide on-site scientific guidance, to ensure progress toward the common goal for in situ detection of life, and to instill a sense of common scientific purpose among the investigators.
This new research program has only just begun and is not yet 6 months old. There are no firm results yet to report. The work done so far does show that even should the ultimate goal prove elusive, there will be a great deal of excellent science accomplished. Some confidence that the program will reach its goal is found in the strategy this program has adopted for life detection. The approach is to look for structures in samples that appear to be nonrepresentative of the common—particularly structures that seem out of equilibrium with the remainder of the sample. One methodology is to look for these structures through elemental analysis, which has no preconceived notion of the particular structure or composition used by life other than that the residue of life must have a distinguishable structure and a chemistry out of equilibrium with the environment. This methodology, applied to terrestrial samples, would seek out structures in rock samples with enrichment of carbon, nitrogen, sulfur, and phosphorus; depletion of aluminum, silicon, iron, and other metals; and association of C, N, O, S, and P both amongst themselves and with Fe, Ni, Co, and other enzymatic metals.
MINIATURIZED IN SITU INSTRUMENTS FOR AMINO ACID DETECTION ON SOLAR SYSTEM BODIES
Jeffrey L. Bada
Scripps Institution of Oceanography
University of California, San Diego
Abstract
The Mars Organic Detector (MOD) is a simple, compact instrument designed for the sensitive in situ detection of organic compounds derived from either prebiotic organic chemistry or extinct/extant organisms. MOD, which was recently selected for the NASA HEDS (Human Exploration and Development of Space) 2005 Mars Explorer Program mission, uses sublimation-based extraction coupled with fluorescence detection to carry out highly sensitive analyses for amino acids or amines and polycyclic aromatic hydrocarbons (PAHs) directly on the martian surface. MOD also has a tunable diode-laser spectrometer that measures the amount of evolved water and carbon dioxide in order to better understand the mineralogy associated with any detected organic compounds, as well as provide a better evaluation of the inventory of water and carbonates in the martian regolith. The next generation of MOD will include a microfabricated capillary electrophoresis system capable of determining the enantiomeric composition (D/L ratios) of any detected amino acids in order to ascertain whether they were of biotic or abiotic origin. MOD-type instruments could also be used to carry out similar in situ analyses on other solar system bodies.
Introduction
As we begin this new millennium, one of the major scientific questions that confronts humanity is whether life exists beyond Earth. There are reasons for optimism. Many of the organic compounds thought to be necessary for the origin of life appear to be widespread throughout the universe. Extrasolar planets seem to be omnipresent companions of main sequence stars. If the conditions that resulted in the origin of life on Earth are common on these extrasolar worlds, then it is reasonable to assume that life could be prevalent in the universe. Within our own solar system, whether life originated on Mars, and perhaps even still exists there today, is an area of intense debate. Although we have sent spacecraft to Mars to search for life and analyzed martian meteorites for possible evidence of life, these efforts have left us with no definitive answers. With increasing evidence of a large subsurface water ocean on the jovian moon Europa, there is the possibility that this body could have a primordial soup similar to that which gave rise to the origin of life on Earth or could potentially even harbor living organisms.
Because of technological limitations, the direct search for extraterrestrial life is presently (and will be for a long time) confined to bodies in our solar system, mainly Mars and Europa. During the next decade the search for evidence of extinct or extant life on Mars will be a central focus of both NASA and the European Space Agency (ESA) as a flotilla of spacecraft explore the martian surface and return martian samples back to Earth for comprehensive state-of-the-art analyses. The challenges are daunting. Missions to Mars are costly and risky, as evidenced by the recent losses of the Mars Climate Orbiter and Mars Polar Lander spacecraft. Collecting and returning samples to Earth, while appealing because of the direct hands-on analytical advantages they provide, are constrained by the amount of material than can be returned and containment issues related to potential biological hazards associated with possible extant martian organisms being transported back to Earth. Spacecraft-based robotic instruments designed to carry out direct analyses for evidence of prebiotic chemistry or life are constrained by mass and power limitations, as well as challenges associated with obtaining samples suitable for analysis.
A major potential impediment to the exploration of Mars for traces of life is the forward contamination of the planet with either terrestrial organisms or biomolecules. This problem makes it essential that organic analyses be carried out as early as possible in the Mars exploration program in order to provide a useful baseline data set for comparison with future analyses.
Searching for Organic Compounds on Mars
In searching for evidence of extinct or extant life on Mars and elsewhere, the detection of organic compounds is considered to be of fundamental importance. Carbon-based polymers such as nucleic acids and proteins make up the core molecules required to carry out the central biological functions of replication and catalysis. Without these functions, life as we know it could not exist.
One of the primary objectives of the 1976 Viking missions was to determine whether organic compounds were present in martian surface soils. Using a pyrolysis procedure, in combination with a highly sensitive gas chromatograph-mass spectrometer (GC-MS), Viking did not detect any organic compounds above a level of a few parts per billion in surface samples at two different landing sites.174 Despite the negative results, it is nevertheless possible that organic compounds, or even living organisms,175 are present on the martian surface at levels below the detection limit of the Viking GC-MS. For example, it is now apparent that the Viking pyrolysis GC-MS instruments would not have detected the presence of nearly a million bacterial cells in 1 gram of soil.176 In addition, oxidation reactions involving organic compounds, whether they are biotic or abiotic in origin, on the martian surface would likely produce nonvolatile products that would also not have been detected by the Viking GC-MS.177
Meteorites from Mars have also been investigated to assess whether they contain organic compounds possibly derived from life. Unfortunately, contamination of martian meteorites by terrestrial-derived organic compounds greatly compromise these investigations.178 This contamination problem underscores the importance of doing in situ organic compound analyses on Mars before samples are returned to Earth, where even under the best of circumstances they will be exposed to some level of terrestrial contamination.
Because amino acids are the building blocks of proteins in terrestrial organisms, they are excellent target compounds in the search for life elsewhere. In addition,over 70 different amino acids have been identified in carbonaceous meteorites,179 which have almost certainly been delivered to Mars throughout its history.180 While it is not certain that martian biology would use the exact same set of amino acids as life on Earth, their ubiquity as constituents of organic material in the solar system suggests that amino acids would have been available for incorporation into living entities on Mars just as they were on Earth. Amino acids derived from either extinct or extant life, and from the infall of meteorites and cosmic dust, could be present on the surface of Mars.
Another class of organic compounds of interest are polycyclic aromatic hydrocarbons. Although PAHs have no known role in biochemistry on Earth, they can be produced from the long-term (tens of millions of years or more) decomposition of biologically derived organic compounds.181 PAHs have been identified in the interstellar medium182 and in carbonaceous meteorites.183 They may be the most abundant single class of organic compounds in the universe, perhaps making up as much as 10 to 20 percent of the total carbon.184 Given the infall of meteorites and cosmic dust throughout the history of Mars, PAHs would be expected to be one of the organic components of the martian surface, especially if samples contained fragments of carbonaceous chondrites.
The MOD Concept
To evaluate whether organic compounds are present on Mars, in-situ-based analytical techniques with sensitivities several orders of magnitude greater than those of the Viking GC-MS are required. In addition, given the limitations of available resources (especially cost), instruments must be compact and able to detect the targeted compounds without requiring extensive sample processing.
MOD is an in situ instrument that has been developed to search for traces of the key organic compounds, amino acids or amines and PAHs, directly on the martian surface. MOD is based on the following concepts:
-
Amino acids and PAHs can be directly sublimed from natural samples by heating to 450°C under partial vacuum, thus eliminating the need for the aqueous and organic solvents used in laboratory analyses; and
-
The sublimed amino acids are condensed on a cold finger coated with a reagent specific for amino acids and detected at very high sensitivities using ultraviolet fluorescence. Sublimed PAHs can be directly detected on the cold finger because they are naturally fluorescent when exposed to ultraviolet light.
Laboratory experiments have demonstrated the feasibility of these concepts. A mixture of dry amino acids, or a crushed sample of a fossil mollusk shell, was placed into a sublimation apparatus that was then evacuated to 5 to 6 torr to approximate martian atmospheric pressure.185 A cold finger was cooled to –195°C with liquid nitrogen, and the apparatus was heated to 450°C for various time periods ranging from 30 seconds to several minutes. The material that sublimed onto the cold finger was then analyzed. No decomposition into amines, which are produced by amino acid decarboxylation, was observed with the pure amino acid mixtures. The behavior of amino acids in a fossil mollusk shell during sublimation was found to be more complex than pure amino acid mixtures. Virtually all of the amino acids were decomposed into amines. However,the amines can be readily detected with the same methodologies used for amino acid detection. Even when amino acid decomposition to amines is significant, the presence of amino acids in the original sample can still be inferred.
Primary amines were found to react with the reagent fluorescamine coated on a cold-finger surface to yield intensely fluorescent derivatives. Experiments indicate that the fluorescamine reaction proceeds in the dry state; no solvent is necessary. Amino acid or amine detection limits with this method are in the 10–13 –10–14 mole range. Thus, even if amino acids were present in a martian sample at a level of a few parts per trillion, they would be detectable by the fluorescamine-based method.
PAHs also readily sublime under the same conditions used for amino acids. The detection of sublimed PAHs can be carried out directly on the cold finger without the need for derivatization reagents because these compounds are extremely fluorescent when irradiated with near-ultraviolet light. Detection limits with this approach are in the subfemtomole (10–15 mole) range.
The 2005 MOD Instrument
Because of its advanced development and ability to address key scientific issues, MOD was recently selected to be part of the HEDS lander instrument package for the 2005 Mars Explorer Program mission. The 2005 version of MOD carries out two simultaneous complementary experiments: (1) simple sublimation-based extraction, fluorescence detection, and quantitation of amino acids, amines, and PAHs at subpicogram (<10–12 gram) levels (i.e., ~1000 times more sensitive than Viking) and (2) the detection and quantitation of evolved water and carbon dioxide using a tunable diode laser (TDL) in order to assess the water and carbonate inventory of the martian surface.
The MOD instrument consists of the following components (Figure 1):
-
A rock crusher that pulverizes samples;
-
The organic detector, consisting of a sublimation cell, a chemical detector, and a fluorescence analyzer; and
-
The TDL spectrometer (not shown in Figure 1), which consists of a Herriott cell, a dual laser system, and a miniature capacitance manometer and Pirani gauge to measure absolute pressure.
MOD operates by the stepwise heating of a crushed sample at martian ambient pressure to a temperature of 950°C. During the heating process, the target organic compounds will sublime from the sample. At the same time, bound water, along with carbon dioxide from the decomposition of various carbonate minerals, will be released. The sublimed material is collected on a cold finger for analysis. The detector plate on the cold finger is divided into two zones: one zone is coated with fluorescamine for amino acid detection; the other zone is uncoated and is used to directly detect PAHs, which do not require a reagent in order to produce an intense ultraviolet fluorescent signal. The quantities of water and carbon dioxide, and their isotopic compositions, evolved during the stepheating cycle are determined using the TDL spectrometer. Even if no organic compounds are detected, the TDL spectrometer provides information on the amounts of bound water and carbonate minerals contained in the Mars regolith. With minor modifications, the TDL spectrometer can also be used by measure the water and carbon dioxide content, and isotopic composition, of the martian atmosphere.

FIGURE 1. The MOD 2005 instrument. A sample collected by a drill or a scoop is dropped into the rock crusher, which pulverizes the sample and then drops it into the oven. After closing the oven at Mars ambient pressure, the crushed sample will be heated stepwise to 950°C. Amino acids and PAHs in the sample will be sublimed and collected on the sample wheel (right end of the oven), which is cooled to Mars nighttime temperatures (around –100°C). The condensed compounds are detected by fluorescence using laser-based sensors. A TDL spectrometer (not shown), which is on the far side of this view, measures the released amounts and isotopic composition of water as well as carbon dioxide produced from the decomposition of various carbonate minerals. The entire instrument weighs ~2 kg and fits in the palm of your hand.
MOD—The Next Generation
A central problem in amino acid analyses of martian samples is not only identifying and quantifying which compounds are present, but also distinguishing those produced abiotically from those synthesized by extinct or extant life.186 Amino acid homochirality (enantiopurity) provides an unambiguous way of distinguishing between abiotic and biotic origins. Proteins made up of both D- and L-amino acids would not likely have been efficient catalysts in early organisms because they could not fold into bioactive configurations such as the α-helix. However, enzymes made up of all D-amino acids function just as well as those made up of only L-amino acids, but the two enzymes use the opposite stereoisomeric substrates. There are no biochemical reasons why L-acids would be favored over D-amino acids. On Earth, the use of only L-amino acids by life is probably simply a matter of chance. We assume that if proteins and enzymes were a component of extinct or extant life on Mars, then amino acid homochirality would have been a requirement. However, the possibility that martian life was (or is) based on D-amino acids would be equal to that based on L-amino acids.
The detection of a nonracemic mixture of amino acids in a martian sample would be strong evidence for the presence of an extinct or extant biota on Mars. The finding of an excess of D-amino acids would provide irrefutable evidence of unique martian life that could not have been derived from seeding the planet with terrestrial life (or the seeding of Earth with martian life). In contrast, the presence of racemic amino acids, along with abiotic amino acids such as α-aminoisobutyric acid, could be indicative of an abiotic origin or, alternatively, the racemization of biotically produced amino acids.187
A potential impediment to the search for life on Mars is the forward contamination of the planet with either terrestrial organisms or, more likely, terrestrial biomolecules. This problem would be of great importance in assessments of whether there are any amino acids indigenous to Mars. Because of the distinctive L-enantiomer signature of amino acids associated with terrestrial life, chiral amino acid analyses can be used to monitor the level of forward contamination of Mars that occurs during the course of planetary exploration. This requires that amino acid analysis data be acquired as early as possible in the Mars exploration program in order to provide a useful baseline data set for comparison with future analyses. A long-range monitoring program would be critical in assessing forward contamination during the eventual human exploration of Mars.
A relatively new technology that shows promise for spacecraft-based amino acid enantiomeric analysis is microchip-based capillary electrophoresis (μCE). With μCE, both the identity and the enantiomeric composition of amino acids can be determined at sub-part-per-billion levels. The μCE-based analyses are about an order of magnitude faster than analytical methods such as conventional capillary electrophoresis (CE) and high-performance liquid chromatography (HPLC). Such short analysis times are an inherent advantage for robotic in situ measurements carried out from a spacecraft. In addition, μCE has a detection limit more than three orders of magnitude better than conventional HPLC. Thus, proportionally smaller samples (~100 pl or 10–10 l) can be analyzed, another important advantage for in situ spacecraft-based instruments.
A μCE chip system has been used to explore the feasibility of using such devices to analyze for amino acid enantiomers in extraterrestrial samples.188 The test system consisted of a folded electrophoresis channel (19.0 cm long × 150 μm wide × 20 μm deep) that was photolithographically fabricated in a 10-cm-diameter glass wafer sandwich, coupled to a laser-excited confocal fluorescence detection apparatus providing subattomole (<10–18 mole) sensitivity. The μCE analysis system consists of a stack of wafer-scale components that individually provide the liquid flow channels, the capillary separation zones, the electrophoretic controls, the fluid logic, and the detection system.This μCE system is more than an order of magnitude smaller in size than conventional laboratory bench top amino acid analytical instruments.
A critical aspect is that enantiomeric ratios can be rapidly and accurately determined using the microfabricated μCE chip instrument. Using a sodium dodecyl sulfate/γ-cyclodextrin pH 10.0 carbonate electrophoresis buffer and a separation voltage of 550 V/cm at 10°C, baseline resolution is observed for the enantiomers of valine, alanine, glutamic acid, and aspartic acid in only 4 minutes (Figure 2). Enantiomeric ratios of amino acids extracted from the Murchison meteorite using this μCE chip system closely matched values determined by HPLC.
The reduced time, resources, and sample requirements for microfabricated μCE translate into a significant reduction in mass, power, and volume. With an estimated mass of ~1 kg, a volume of ~1000 cm3, and a power requirement of ~2 W, the μCE chip system provides a compact, low-mass instrument suitable for a wide variety of in situ exobiology applications. For spacecraft-based analyses, a microfluidics-based sample processing system will be needed in order to deliver an amino acid extract suitable for analysis by a μCE system. The design of such a system is presently under way.
Conclusions
This discussion has focused on amino acid detection systems tailored for missions to Mars. However, other solar system bodies, such as Europa, Saturn’s moon Titan, asteroids, and comets, likely hold information about natural abiotic synthetic processes and the conditions necessary for synthesis of the organic compounds needed for the origin of life. In the case of Europa, compounds derived from living entities could possibly be present. In situ analyses carried out on these solar system bodies could thus potentially provide information about the suite of organic compounds that may have been present on prebiotic Earth and how the organic compounds used by

FIGURE 2. Baseline resolution of several amino acid enantiomers using the μCE chip system. SOURCE: L.D. Hutt, D.P. Glavin, J.L. Bada, and R.A. Mathies, “Microfabricated Capillary Electrophoresis Amino Acid Chirality Analyzer for Extraterrestrial Exploration,” Anal. Chem. 71:4000-4006, 1999.
extraterrestrial life compare with those used by terrestrial organisms. The MOD instrument concept described here for investigations on Mars could easily be tailored for use on other bodies of interest in the solar system.
Acknowledgments
The development of MOD has been funded by the Planetary Instrument Definition and Development Program (PIDDP) and the Mars Instrument Development Program (MIDP) of NASA and the NASA Specialized Center of Research and Training in Exobiology at the Scripps Institution of Oceanography. The following members of the “MOD squad ” contributed to this work:: C. LaBaw, C. Mahoney, G. McDonald, O. Serviss, C.R. Webster, and F. Grunthaner (NASA-JPL); R. Mathies and L. Hutt (UC Berkeley); and G. Kminek, O. Botta, and D. Glavin (UCSD).
REFERENCES FOR PAPERS IN SESSION 4
1. J.W. Schopf, “Fossils and Pseudofossils: Lessons from the Hunt for Early Life on Earth,” in Size Limits of Very Small Micro-organisms: Proceedings of a Workshop, National Academy Press, Washington, D.C., 1999, pp. 88-93.
2. J.M. Schopf, “Modes of Fossil Preservation,” Review of Palaeobotany and Palynology 20:27-53, 1975.
3. J.D. Farmer and D.J. Des Marais, “Exploring for a Record of Ancient Martian Life,” Journal of Geophysical Research 104(E11):2697726995, 1999.
4. Space Studies Board, National Research Council, Size Limits of Very Small Microorganisms: Proceedings of a Workshop, National Academy Press, Washington, D.C., 1999, p. 85.
5. See, for example, R.E. Summons and M.R. Walter, “Molecular Fossils and Microfossils of Prokaryotes and Protists from Proterozoic Sediments,” American Journal of Science 290-A:212-244, 1990.
6. See, for example, M. Schidlowski, J.M. Hayes, and I.R. Kaplan,“Isotopic Inferences of Ancient Biochemistries: Carbon, Sulfur, Hydrogen, and Nitrogen,” in Earth’s Earliest Biosphere: Its Origin and Evolution, J.W. Schopf (ed.), Princeton University Press, Princeton, New Jersey, 1983, pp. 147-186.
7. See, for example, T.J. Beveridge, M.N. Hughes, H. Lee, K.T. Leung, R.K. Poole, I. Savvaidis, S. Silver, and J.T. Trevors, “MetalMicrobe Interactions: Contemporary Approaches,” Advances in Microbial Physiology 38:178-243, 1997.
8. See, for example, J.W. Schopf and M.R. Walter, “Archean Microfossils: New Evidence of Ancient Microbes,” in Earth’s Earliest Biosphere: Its Origin and Evolution, J.W. Schopf (ed.), Princeton University Press, Princeton, New Jersey, 1983, pp. 214-239.
9. See, for example, R. Buick, “Microfossil Recognition in Archean Rocks:An Appraisal of Spheroids and Filaments from a 3500 M.Y. Old Chert-Barite Unit at North Pole, Western Australia,” Palaios 5:441-459, 1990.
10. See, for example, J.W. Schopf, “Fossils and Pseudofossils: Lessons from the Hunt for Early Life on Earth,” in Size Limits of Very Small Microorganisms: Proceedings of a Workshop, National Academy Press, Washington, D.C., 1999, pp. 88-93.
11. See, for example, J.W. Schopf and M.R. Walter, “Archean Microfossils:New Evidence of Ancient Microbes,” in Earth’s Earliest Biosphere: Its Origin and Evolution, J.W. Schopf (ed.), Princeton University Press, Princeton, New Jersey, 1983, pp. 214-239.
12. See, for example, H.J. Hofmann and J.W. Schopf, “Early Proterozoic Microfossils,” in Earth’s Earliest Biosphere: Its Origin and Evolution, J.W. Schopf (ed.), Princeton University Press, Princeton, New Jersey, 1983, pp. 321-360.
13. R. Buick, “Microfossil Recognition in Archean Rocks: An Appraisal of Spheroids and Filaments from a 3500 M.Y. Old Chert-barite Unit at North Pole, Western Australia,” Palaios 5: 441-459, 1990.
14. A.H. Knoll, “Exceptional Preservation of Photosynthetic Organisms in Silicified Carbonates and Silicified Peats,” Philosophical Transactions of the Royal Society of London B 311:111-122, 1985.
15. See, for example, L.P. Knauth, “Petrogenesis of Chert,” in Silica: Physical Behavior, Geochemistry, and Materials Applications, P.J. Heaney, C.T. Prewitt, and G.V. Gibbs (eds.), Reviews in Mineralogy, Volume 29, Mineralogical Society of America, Washington, D.C., 1994, pp. 233-258.
16. M.R. Walter and D.J. Des Marais, “Preservation of Biological Information in Thermal Spring Deposits:Developing a Strategy for the Search for a Fossil Record on Mars,” Icarus 101:129-143, 1993.
17. S.L. Cady and J.D. Farmer, “Fossilization Processes in Siliceous Thermal Springs: Trends in Preservation Along Thermal Gradients,” in Evolution of Hydrothermal Ecosystems on Earth (and Mars?), Ciba Foundation Symposium No.202, G.R. Bock and J.A. Goode (eds.), John Wiley and Sons, Chichester, U.K., 1996, pp. 150-173.
18. J.D. Farmer, “Thermophiles, Early Biosphere Evolution, and the Origin of Life on Earth: Implications for the Exobiological Exploration of Mars,” Journal of Geophysical Research 103:28457-28461, 1998.
19. See, for example, K.O. Stetter, “Hyperthermophiles in the History of Life,” in Evolution of Hydrothermal Ecosystems on Earth (and Mars?), Ciba Foundation Symposium No.202, G.R. Bock and J.A. Goode (eds.), John Wiley and Sons, Chichester, U.K., 1996, pp. 1-18.
20. See, for example, D.M. Ward, R. Weller, J. Shiea, R.W. Castenholtz, and Y. Cohen, “Hot Spring Microbial Mats: Anoxygenic and Oxygenic Mats of Possible Evolutionary Significance,” in Microbial Mats, Physiological Ecology of Benthic Microbial Communities, Y. Cohen and E. Rosenberg (eds.), American Society for Microbiology, Washington, D.C., 1989, pp. 3-15.
21. See, for example, J.A. Baross and S.E. Hoffman, “Submarine Hydrothermal Vents and Associated Gradient Environments as Sites for the Origin and Evolution of Life,” Origins of Life 15:327-345, 1985.
22. S.L. Cady and J.D. Farmer, “Fossilization Processes in Siliceous Thermal Springs:Trends in Preservation Along Thermal Gradients,” in Evolution of Hydrothermal Ecosystems on Earth (and Mars?), Ciba Foundation Symposium No. 202, G.R. Bock and J.A. Goode (eds.), John Wiley and Sons, Chichester, U.K., 1996, pp. 150-173.
23. M.R. Walter and D.J. Des Marais, “Preservation of Biological Information in Thermal Spring Deposits: Developing a Strategy for the Search for a Fossil Record on Mars,” Icarus 101:129-143, 1993.
24. S.L. Cady, M.R. Walter, D.J. DesMarais, and C.E. Blank, “Exopaleontological Search Strategy for Mars Exploration: A Case for Siliceous Epithermal Deposits,” in Lunar and Planetary Science Conference XXIII, Lunar and Planetary Institute, Houston, Texas, 1997.
25. See, for example, R. Hesse, “Silica Diagenesis: Origin of Inorganic and Replacement Cherts,” Earth-Science Reviews 26:253-284, 1989.
26. J.D. Farmer and D.J. Des Marais, “Exploring for a Record of Ancient Martian Life,” Journal of Geophysical Research 104(E11):2697726995, 1999.
27. D.S. McKay, E.K.Gibson, K.L.Thomas-Keprta, H.Vali, C. Romanek, S.J. Clemett, X.D.F. Chiller, C.R. Maechling, and R.N. Zare, “Search for Past Life on Mars:Possible Relic Biogenic Activity in Martian Meteorite ALH84001,” Science 273:924-930, 1996.
28. F. Westall, “The Nature of Fossil Bacteria: A Guide to the Search for Extraterrestrial Life,” Journal of Geophysical Research 104:16437-16451, 1999.
29. F. Westall, “The Influence of Cell Wall Composition on the Fossilization of Bacteria and the Implications for the Search for Early Life Forms,” in Astronomical and Biochemical Origins and the Search for Life in the Universe, C. Cosmovici, S. Bowyer, and D. Werthimer (eds.), Editori Compositrici, Bologna, 1997, pp. 491-504.
30. See, for example, T.J. Beveridge, M.N. Hughes, H. Lee, K.T. Leung, R.K. Poole, I. Savvaidis, S. Silver, and J.T. Trevors, “MetalMicrobe Interactions: Contemporary Approaches,” Advances in Microbial Physiology 38:178-243, 1997.
31. See, for example, F.G. Ferris, “Formation of Authigenic Minerals by Bacteria.” in Biological--Interactions, J.M. McIntosh and L.A. Groat (eds.), Mineralogical Association of Canada, Ottawa, 1997, pp. 187-208.
32. D. Fortin, F.G. Ferris, and T.J. Beveridge, “Surface-mediated Mineral Development by Bacteria,” in Geomicrobiology: Interactions Between Microbes and Minerals, J. Banfield and K.H. Nealson (eds.), Reviews in Mineralogy, Volume 35, Mineralogical Society of America, Washington, D.C., 1997, pp. 161-180.
33. See, for example, K.H. Nealson, “The Limits of Life on Earth and Searching for Life on Mars,” Journal of Geophysical Research 102:23675-23686, 1997.
34. See, for example, J.D. Farmer and D.J. Des Marais, “Exploring for a Record of Ancient Martian Life,” Journal of Geophysical Research 104(E11):26977-26995, 1999.
35. See, for example, J. Wingender, T.R. Neu, and H.-C. Flemming, Microbial Extracellular Polymeric Substances: Characterization, Structure and Function, Springer-Verlag, Berlin, 1999.
36. See, for example, P. Stoodley, D. deBeer, J.D. Boyle, and H.M. Lappin-Scott, “Evolving Perspectives of Biofilm Structure,” Biofouling 14:75-94, 1999.
37. See, for example, S.L. Cady and J.D. Farmer, “Fossilization Processes in Siliceous Thermal Springs: Trends in Preservation Along Thermal Gradients,” in Evolution of Hydrothermal Ecosystems on Earth (and Mars?), Ciba Foundation Symposium No.202, G.R. Bock and J.A. Goode (eds.), John Wiley and Sons, Chichester, UK, 1996, pp. 150-173.
38. W.B. Whitman, “Prokaryotes:The Unseen Majority,” Proc. Natl. Acad. Sci. USA 95:6578-6583, 1998.
39. See, for example, K. Pedersen, “Exploration of Deep Intraterrestrial Microbial Life:Current Perspectives,” FEMS Microbiology Letters 185:9-16, 2000.
40. D.S. McKay, E.K. Gibson, K.L. Thomas-Keprta, H. Vali, C. Romanek, S.J. Clemett, X.D.F. Chiller, C.R. Maechling, and R.N. Zare, “Search for Past Life on Mars: Possible Relic Biogenic Activity in Martian Meteorite ALH84001,” Science 273:924-930, 1996.
41. M.C. Malin and K.S. Edgett, “Evidence for Recent Groundwater See page and Surface Runoff on Mars,” Science 288:2330-2335, 2000.
42. See, for example, F.G.Ferris, “Formation of Authigenic Minerals by Bacteria,” in Biological--Interactions, J.M. McIntosh and L.A. Groat (eds.), Mineralogical Association of Canada, Ottawa, 1997, pp. 187-208.
43. See, for example, A.H. Knoll, “Exceptional Preservation of Photosynthetic Organisms in Silicified Carbonates and Silicified Peats,” Philosophical Transactions of the Royal Society of London B 311:111-122, 1985.
44. L.M. Gerasimenko and I.N. Krylov, “Postmortem Alterations of Cyanobacteria in the Algal-bacterial Films in the Hot Springs of Kamchatka,” Dokl. Akad. Nauk 272:215-218, 1983.
45. J.K. Bartley, “Actualistic Taphonomy of Cyanobacteria:Implications for the Precambrian Fossil Record,” Palaios 11:571-586, 1996.
46. See, for example, S. Schultz-Lam, F.G. Ferris, K.O. Konhauser, and R.G. Wiese, “In Situ Silicification of an Icelandic Hot Spring Microbial Mat:Implications for Microfossil Formation,” Canadian Journal of Earth Sciences 32:2021-2026, 1995.
47. See, for example, K.O. Konhauser and F.G. Ferris, “Diversity of Iron and Silica Precipitation by Microbial Mats in Hydrothermal Waters, Iceland:Implications for Precambrian Iron Formations,” Geology 24:323-326, 1996.
48. E.W. Tegelaar, J.W. De Leeuw, S. Derenne, and C. Largeau, “A Reappraisal of Kerogen Formation,” Geochimica et Cosmochimica Acta 53:3103-3106, 1989.
49. G.A. Logan, M.J. Collins, and G. Eglinton, “Preservation of Organic Biomolecules,” in Taphonomy; Releasing the Data Locked in the Fossil Record, P.A. Allison and D.E.G. Briggs (eds.), Plenum Press, New York, 1991, pp. 1-24.
50. P.A. Allison and D.E.G. Briggs, “Taphonomy of Non-Mineralized Tissues,” in Taphonomy: Releasing the Data Locked in the Fossil Record, P.A. Allison and D.E.G. Briggs (eds.), Plenum Press, New York, 1991, pp. 25-70.
51. See, for example, J.W. De Leeuw and C. Largeau, “A Review of Macromolecular Organic Compounds That Comprise Living Organisms and Their Role in Kerogen, Coal, and Petroleum Formation,” in Organic Geochemistry, Principles and Applications, M.H. Engel and S.A. Macko (eds.), Plenum Press, New York, 1993, pp.23-72.
52. R.E. Summons, “Biogeochemical Cycles:A Review of Fundamental Aspects of Organic Matter Formation, Preservation,and Composition,” in Organic Geochemistry, Principles and Applications, M.H. Engel and S.A. Macko (eds.), Plenum Press, New York, 1993, pp.3-21.
53. See, for example, S.L. Cady, K. Downing, and H.-R. Wenk, “HRTEM of Microcrystalline Opal in Chert and Porcelanite from the Monterey Formation,California,” American Mineralogist 81:1380-1395, 1996.
54. See, for example, S.L. Cady, H.R.Wenk, and M. Sintuban, “Microfibrous Quartz Varieties:Characterization by Quantitative X-Ray Texture Analysis and Transmission Electron Microscopy,” Contributions to Mineralogy and Petrology 130:320-335, 1998.
55. R. Buick, “Microfossil Recognition in Archean Rocks: An Appraisal of Spheroids and Filaments from a 3500 M.Y. Old Chert-Barite Unit at North Pole,Western Australia,” Palaios 5:441-459, 1990.
56. G. Heiken and D. McKay, “Petrology of a Sequence of Pyroclastic Rocks from the Valley of Taurus-Littrow,” in Ninth Lunar and Planetary Science Conference, Houston, Texas, March 13-17, 1978, Proceedings, Volume 2, Pergamon Press, New York, 1978, pp. 19331943.
57. W. C. Phinney, D. S. McKay, C. H. Simonds, and J. L. Warner, “Lithification of Vitric-and Clastic-matrix Breccias:SEM Petrography,” in Seventh Lunar Science Conference, Houston, Texas, March 15-19, 1976, Proceedings, Volume 2, Pergamon Press, New York, 1976, pp. 2469-2492.
58. F. Westall, “The Nature of Fossil Bacteria:A Guide to the Search for Extraterrestrial Life,” Journal of Geophysical Research 104:16437-16452, 1999.
59. F. Westall, “The Nature of Fossil Bacteria:A Guide to the Search for Extraterrestrial Life,” Journal of Geophysical Research 104:16437-16452, 1999.
60. G. Heiken and D. McKay, “Petrology of a Sequence of Pyroclastic Rocks from the Valley of Taurus-Littrow,” in Ninth Lunar and Planetary Science Conference, Houston, Texas, March 13-17, 1978, Proceedings, Volume 2, Pergamon Press, New York, 1978, pp. 19331943.
61. W. C. Phinney, D. S. McKay, C. H. Simonds, and J. L. Warner, “Lithification of Vitric-and Clastic-matrix Breccias:SEM Petrography,” in Seventh Lunar Science Conference, Houston, Texas, March 15-19, 1976, Proceedings, Volume 2, Pergamon Press, New York, 1976, pp. 2469-2492.
62. K. Thomas-Keprta, D. S. McKay, and S. J. Wentworth, “Bacterial Mineralization Patterns in Basaltic Aquifers:Implications for Possible Life in Martian Meteorite ALH84001,” Geology 26:1031-1034, 1998.
63. K. Biemann, J. Oro, P. Toulmin III, L. E. Orgel, A. O. Nier, D. M. Anderson, P. G. Simmonds, D. Flory, A. V. Diaz, D. R. Rushneck, J. E. Biller, and A. L. Lafleur, “The Search for Organic Substances and Inorganic Volatile Compounds in the Surface of Mars,” Journal of Geophysical Research 82:4641-4658, 1977.
64. S. A. Benner, K. G. Devine, L. N. Matveeva, and D. H. Powell, “The Missing Organic Molecules on Mars,” Proc. Natl. Acad. Sci. USA 97:2425-2430, 2000.
65. D. S. McKay, E. K. Gibson, K. L. Thomas-Krepta, H. Vali, C. S. Romanek, S. J. Clemett, X. D. F. Chillier, C. R. Maechling, and R. N. Zare, “Search for Past Life on Mars:Possible Relic Biogenic Activity in Martian Meteorite ALH84001,” Science 273:924-930, 1996.
66. L. Becker, B. Popp, T. Rust, and J. L. Bada, “The Origin of Organic Matter in the Martian Meteorite ALH84001,” Earth Planet. Sci. Lett. 167:71, 1999.
67. A. J. T. Jull, C. Courtney, D. A. Jeffrey, and J. W. Beck, “Isotopic Evidence for a Terrestrial Source of Organic Compounds Found in Martian Meteorites Allan Hill 84001 and Elephant Moraine 79001”, Science 279:366, 1999.
68. A. J. T. Jull, J. W. Beck, and G. S. Burr, “Isotopic Evidence for Extraterrestrial Organic Material in the Martian Meteorite Nakhla”, Geochim. Cosmochim. Acta 64:3763-3772, 2000, in press.
69. J. L. Bada, “Extraterrestrial Handedness,” Science 275:942, 1997.
70. K. Brinton and G. D. McDonald, “Amino Acid Racimezation Studies in Siberian Permafrost,” Abstract, Astrobiology Workshop, NASA Ames Research Center, April 2000.
71. W. H. Nelson, R. Manoharan, and J. F. Sperry, “UV Resonance Raman Studies of Bacteria,” Applied Spectroscopy Reviews 27(1):67124, 1992.
72. J. Urmos, S. K. Sharma, and F. T. Mackenzie, “Characterization of Some Biogenic Carbonates with Raman Spectroscopy,” American Mineralogist 76:641, 1991.
73. H. G. M. Edwards, D. W. Farwell, M. M. Grady, D. D. Wynn-Williams, and I. P. Wright, “Comparative Raman Microscopy of a Martian Meteorite and Antarctic Lithic Anologues,” Planet. Space Sci. 47:353, 1999.
74. D. S. McKay, E. K. Gibson, K. L. Thomas-Krepta, H. Vali, C. S. Romanek, S. J. Clemett, X. D. F. Chillier, C. R. Maechling, and R. N. Zare, “Search for Past Life on Mars:Possible Relic Biogenic Activity in Martian Meteorite ALH84001,” Science 273:924-930, 1996.
75. L. Becker, B. Popp, T. Rust, and J. L. Bada, “The Origin of Organic Matter in the Martian Meteorite ALH84001,” Earth Planet. Sci. Lett. 167:71, 1999.
76. See paper by L. Becker, Session 4, this appendix.
77. K. Biemann, J. Oro, P. Toulmin III, L. E. Orgel, A. O. Nier, D. M. Anderson, P. G. Simmonds, D. Flory, A. V. Diaz, D. R. Rushneck, J. E. Biller, and A. L. Lafleur, “The Search for Organic Substances and Inorganic Volatile Compounds in the Surface of Mars,” Journal of Geophysical Research 82:4641-4658, 1977.
78. See paper by A. Anbar, Session 4, this appendix.
79. K. A. Delin and S. P. Jackson, “Sensor Web for In Situ Exploration of Gaseous Biosignatures,” IEEE Aerospace Conf. Proceedings, Big Sky, Montana, 2000.
80. R. Bell and D. M. Karl (conveners. ), Lake Vostok Workshop Final Report, National Science Foundation, Arlington, Va., November 78, 1998.
81. J. C. Petit, J. Jouzel, D. Raynaud, N. I. Barkov, J. M. Barnola, I. Basile, M. Bencer, J. Chappellaz, D. Davis, G. Delaygue, M. Delmotte, V. M. Kotlyyakov, M. Legrand, V. Y. Lipenkov, C. Lorius, L. Pepin, C. Ritz, R. E. Saltzman, and M. Stievenard, “Climate and Atmospheric History of the Past 420, 000 Years from the Vostok, Antarctica,” Nature 399:429, 1999.
82. D. M. Karl, D. F. Bird, K. Bjorkman, T. Houlihan, R. Shackelford, and L. Tupas, “Microorganisms in the Accreted Ice of Lake Vostok, Antarctica,” Science 286:2144-2147, 1999.
83. J. C. Priscu, E. Adams, W. B. Lyons, M. A. Voytek, D. W. Mogk, R. L. Brown, C. P. McKay, C. D. Takacs, K. A. Welch, C. F. Wolf, J. D. Kirshtein, and R. Avci, “Geomicrobiology of Subglacial Ice Above Lake Vostok, Antarctica, ”Science 286:2141-2144, 1999.
84. G. Eglinton, P. M. Scott, T. Besky, A. L. Burlingame, and M. Calvin, “Hydrocarbons of Biological Origin from a One-Billion-Year-Old Sediment, ”Science 145:263-264, 1964.
85. G. Eglinton and M. Calvin, “Chemical Fossils, ”Scientific American 261:32-43, 1967.
86. Proceedings of the Biosignature Workshop, University of California, San Diego, March 3-5, 2000.
87. K. E. Peters and J. M. Moldowan, The Biomarker Guide: Interpreting Molecular Fossils in Petroleum and Ancient Sediments, Prentice Hall, Englewood Cliffs, New Jersey, 1993.
88. See, for example, R. E. Summons, L. L Jahnke, J. M. Hope, and G. A. Logan, “2-Methylhopanoids as Biomarkers for Cyanobacterial Oxygenic Photosynthesis, ”Nature 400:554-557, 1999.
89. M. Rohmer, P. Bouvier, and G. Ourisson, “Molecular Evolution of Biomembranes:Structural Equivalents and Phylogenetic Precursors of Sterols, ”Proc. Natl. Acad. Sci. USA 76:847-851, 1979.
90. T. M. McCollom, G. Ritter, and B. R. T. Simoneit, “Lipid Synthesis Under Hydrothermal Conditions by Fischer-Tropsch-type Reactions, ”Orig. Life Evol. Biosph. 28:1-14, 1998.
91. F. Kenig, J. S. Sinninghe Damsté, A. C. Kock-van Dalen, W. I. C. Rijpstra, A. Y. Huc, and J. W. de Leeuw, “Occurrence and Origins of Mono, Di, and Trimethylalkanes in Modern and Holocene Cyanobacterial Mats from Abu Dhabi, United Arab Emirates, ”Geochim. Cosmochim. Acta 59:2999-3015, 1995.
92. G. Flesch and M. Rohmer, “Prokaryotic Hopanoids:The Biosynthesis of the Bacteriohopane Skeleton. Formation of Isoprenic Units from Two Distinct Acetate Pools and a Novel Type of Carbon/Carbon Linkage Between a Triterpene and D-Ribose, ”Eur. Journal Biochem. 175:405-411, 1988.
93. G. Ourisson and P. Albrecht, “Hopanoids I. Geohopanoids:The Most Abundant Natural Products of Earth, ”Chem. Res. 25:398-402, 1992.
94. G. Ourisson, P. Albrecht, and M. Rohmer, “The Microbial Origin of Fossil Fuels, ”Scientific American 251:44-51, 1984.
95. B. Chappe, P. Albrecht, and W. Michaelis, “Polar Lipids of Archaebacteria in Sediments and Petroleums, ”Science 217:65-66, 1982.
96. See, for example, R. E. Summons, L. L Jahnke, J. M. Hope, and G. A. Logan, “2-Methylhopanoids as Biomarkers for Cyanobacterial Oxygenic Photosynthesis, ”Nature 400:554-557, 1999.
97. J. J. Brocks, G. A. Logan, R. Buick, and R. E. Summons, “Archaean Molecular Fossils and the Early Rise of Eukaryotes, ”Science 285:1033-1036, 1999.
98. T. -M. Han and B. Runnegar, “Megascopic Eukaryotic Algae from the 2. 1-billion-year-old Negaunee Iron-formation, Michigan, ” Science 257:232-235, 1992.
99. A. H. Knoll, “The Early Evolution of Eukaryotes:A Geological Perspective, ”Science 256:622-627, 1992.
100. N. Woolf, “Finding/Studying the Pale Blue Dot, ” in Pale Blue Dot 2 Workshop: Habitable and Inhabited Worlds Beyond Our Own Solar System, L. I. Caroff and D. J. Des Marais (eds.) , NASA/CP 2000-209595, Ames Research Center, Moffett Field, California, 2000, pp. 33-46.
101. See paper by J. M. Moldowan, Session 4, in this appendix.
102. J. D. MacDougall, G. W. Lugmair, and J. F. Kerridge, “Early Solar System Aqueous Activity:Sr Isotopic Evidence from the Orgueil CI Meteorite, ”Nature 307:249-251, 1984.
103. B. Zellner and E. Bowell, “Asteroid Compositional Types and their Distributions, ” in Comets--Meteorites, A. H. Delsemme (ed.) , University of Toledo Press, Toledo, 1977, pp. 185-195.
104. G. Flynn, “The Delivery of Organic Matter from Asteroids and Comets in the Early Surface of Mars, ”Earth, Moon, and Planets 72:469-474, 1996.
105. J. R. Cronin and S. Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite, ” in The Chemistry of Life’s Origins, J. M. Greenberg, C. X. Mendoza-Gómez, and V. Pirronello (eds.) , Kluwer Academic Press, Boston, 1993, pp. 209258.
106. M. N. Fomenkova, S. Chang, and L. M. Mukhin, “Carbonaceous Components in Comet Halley Dust, ”Geochim. Cosmochim. Acta 58:4503-4512, 1994.
107. E. Anders and E. Zinner, “Interstellar Grains in Primitive Meteorites:Diamond, Silicon Carbide, and Graphite, ”Meteoritics 28:490514, 1993.
108. S. Pizzarello, personal communication.
109. G. Cooper, personal communication.
110. J. R. Cronin, S. Pizzarello, and D. P. Cruikshank, “Organic Matter in Carbonaceous Chondrites, Planetary Satellites, Asteroids and Comets, ” in Meteorites and the Early Solar System, J.. F. Kerridge and M. S. Matthews (eds.) , University of Arizona Press, Tucson, 1988, pp. 819-857.
111. J. R. Cronin, S. Pizzarello, and D. P. Cruikshank, “Organic Matter in Carbonaceous Chondrites, Planetary Satellites, Asteroids and Comets, ” in Meteorites and the Early Solar System, J.. F. Kerridge and M. S. Matthews (eds.) , University of Arizona Press, Tucson, 1988, pp. 819-857.
112. J. R. Cronin and S. Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite, ” in The Chemistry of Life’s Origins, J. M. Greenberg, C. X. Mendoza-Gómez, and V. Pirronello (eds.) , Kluwer Academic Press, Boston, 1993, pp. 209258.
113. See paper by J. M. Moldowan, Session 4, in this appendix.
114. J. R. Cronin and S. Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite, ” in The Chemistry of Life’s Origins,
J.M. Greenberg, C.X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209258.
115. L. Becker, R.J. Poreda, and T.E. Bunch, “Fullerenes: An Extraterrestrial Carbon Carrier Phase for Noble Gases,” Proc. Natl. Acad. Sci. USA 97:2979-2983, 2000. 5
116. J.R. Cronin and S.Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite,” in The Chemistry of Life’s Origins, J.M. Greenberg, C.X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209258.
117. W.A. Bonner, “Homochirality and Life,” in D--Acids in Sequences of Secreted Peptides of Multicellular Organisms, P. Jollès (ed.), EXS Volume 85, Birkhäuser Verlag AG, Switzerland, 1998, pp. 159-188.
118. See, for example, K. Kvenvolden, J. Lawless, K. Pering, E. Peterson, J. Flores, C. Ponnamperuma, I.R. Kaplan, and C. Moore, “Evidence for Extraterrestrial Amino Acids and Hydrocarbons in the Murchison Meteorite,” Nature 228:623-626, 1970.
119. M.H. Engel and B. Nagy, “Distribution of Enantiomeric Composition of Amino Acids in the Murchison Meteorite,” Nature 296:837840, 1982.
120. J.L. Bada, J.R. Cronin, M. -S. Ho, K.A. Kvenvolden, J.G. Lawless, S.L. Miller, J. Oro, and S. Steinberg, “On the Reported Optical Activity of Amino Acids in the Murchison Meteorite,” Nature 301:494-497, 1983.
121. M.H. Engel, S.A. Macko, and J.A. Silfer, “Carbon Isotope Composition of Individual Amino Acids in the Murchison Meteorite,” Nature 348:47-49, 1990.
122. J.R. Cronin and S. Pizzarello, “Enantiomeric Excesses in Meteoritic Amino Acids,” Science 275:951-955, 1997.
123. M.H. Engel and S.A. Macko, “Isotopic Evidence for Extraterrestrial Nonracemic Amino Acids in the Murchison Meteorite,” Nature 389:265-268, 1997.
124. J.A. Bailey, A. Chrysostomou, J.H. Hough, T.M. Gledhill, A. McCall, F. Menard, and M. Tamura, “Circular Polarization in Starformation Regions:Implications for Biomolecular Homochirality,” Science 28l:672-674, 1998.
125. E. Rubenstein, W.A. Bonner, H.P. Noyes, and G.S. Brown, “Supernovae and Life,” Nature 300:118, 1983.
126. W.D. Langer, E.F. van Dishoeck, E.A. Bergin, G.A. Blake, A.G.G.M. Tielens, T. Velusamy, and D.C.B. Whittet, “Chemical Evolution of Protostellar Matter,” in Protostars and Planets IV, V. Mannings, A.P. Boss, and S.S. Russell (eds.), University of Arizona Press, Tucson, 2000, pp. 29-57.
127. J.R. Cronin and S. Chang, “Organic Matter in Meteorites:Molecular and Isotopic Analyses of the Murchison Meteorite,” in The Chemistry of Life’s Origins, J.M. Greenberg, C.X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209258.
128. E. Zinner, “Interstellar Cloud Material in Meteorites,” in Meteorites and the Early Solar System, J.F. Kerridge and M.S. Matthews (eds.), University of Arizona Press, Tucson, 1988, pp. 956-983.
129. E. Anders and E. Zinner, “Interstellar Grains in Primitive Meteorites: Diamond, Silicon Carbide, and Graphite,” Meteoritics 28:490514, 1993.
130. S. Chang, D. Des Marais, R. Mack, S.L. Miller, and G.E. Strathearn, “Prebiotic Organic Synthesis and the Origin of Life,” in Earth’s Earliest Biosphere: Its Origin and Evolution, J.W. Schopf (ed.), Princeton University Press, Princeton, New Jersey, 1983, pp. 53-92.
131. C.C. Kung, R.Hayatsu, M.H. Studier, and R.N. Clayton, “Nitrogen Isotope Fractionations in the Fisher-Tropsch Synthesis and in the Miller-Urey Reaction,” Earth Planet. Sci. Lett. 46:144-146, 1979.
132. R. Summons, “Abundance and Composition of Extractable Organic Matter,” in The Proterozoic Biosphere, J.W. Schopf and C. Klein (eds.), Cambridge University Press, New York, 1992, pp. 101-115.
133. J.M. Hayes, I.R. Kaplan, and K.W. Wedeking, “Precambrian Organic Geochemistry:Preservation of the Record, ” in Earth’s Earliest Biosphere: Its Origin and Evolution, J.W. Schopf (ed.), Princeton University Press, Princeton, New Jersey, 1983, pp. 93-134.
134. H. Strauss, D.J. Des Marais, J.M. Hayes, and R.E. Summons, “The Carbon Isotopic Record,” in The Proterozoic Biosphere, J.W. Schopf and C. Klein (eds.), Cambridge University Press, New York, 1992, pp. 117-127.
135. G. Yuen, N. Blair, D.J. Des Marais, and S. Chang, “Carbon Isotopic Composition of Low Molecular Weight Hydrocarbons and Monocarboxylic Acids from Murchison Meteorite,” Nature 307:252-254, 1984.
136. G.W. Cooper, M.H. Thiemens, T.L. Jackson, and S. Chang, “Sulfur and Hydrogen Isotope Anomalies in Meteorite Sulfonic Acids,” Science 277:1072-1074, 1997.
137. K. Biemann, J. Oro, P. Toulmin III, L.E. Orgel, A.O. Nier, D.M. Anderson, P.G. Simmonds, D. Flory, A.V. Diaz, D.R. Rusneck, J.E. Biller, and A.L. Lafluer, “The Search for Organic Substances and Inorganic Volatile Compounds in the Surface of Mars,” Journal of Geophysical Research 82:4641-4658, 1979.
138. S.J. Mojzsis, G. Arrhenius, and K.D. McKeegan, “Evidence for Life on Earth Before 3, 800 Million Years Ago,” Nature 384:55-59, 1996.
139. B.L. Beard and C.M. Johnson, “High-precision Iron Isotope Measurements of Terrestrial and Lunar Materials,” Geochim. Cosmochim. Acta 63:1653-1660, 1999.
140. C.N. Marechal, P. Telouk, and F. Albarede, “Precise Analysis of Copper and Zinc Isotopic Composition by Plasma-source Mass Spectrometry,” Chem. Geol. 156:251-273, 1999.
141. J. Barling, G.L. Arnold, and A.D. Anbar, “Natural Mass-Dependent Variations in the Isotopic Composition of Molybdenum,” Earth Planet. Sci. Lett. 193:447-457, 2001.
142. X.K. Zhu, R.K. O’Nions, and Y. Guo, “Secular Variation of Iron Isotopes in North Atlantic Deep Water,” Science 287:2000-2002, 2000.
143. X.K. Zhu, R.K. O ’Nions, Y. Guo, N.S. Belshaw, and D. Rickard, “Determination of Natural Cu-Isotope Variation by Plasma-source Mass Spectrometry:Implications for Use as Geochemical Tracers,” Chem. Geol. 163:139-149, 2000.
144. M. Rehkamper, “The Precise Measurement of Tl Isotopic Compositions by MC-ICPMS:Application to the Analysis of Geological Materials and Meteorites,” Geochim. Cosmochim. Acta 63:935-944, 1999.
145. T.D. Bullen, A.F. White, C.W. Childs, D.V. Vivit, and M.S. Schulz, “A Demonstration of Significant Abiotic Iron Isotope Fractionation in Nature,” Geology 194:39-51, 2001.
146. A.D. Anbar, J.E. Roe, and J.Barling, “Nonbiological Fractionation of Iron Isotopes,” Science 288:126-128, 2000.
147. B.L. Beard, C.M. Johnson, and L.Cox, “Iron Isotope Biosignatures,” Science 285:1889-1892, 1999.
148. S.L. Brantley, L.Liermann, and T.D. Bullen, “Fractionation of Fe Isotopes by Soil Microbes and Organic Acids,” Geology 29:535538, 2001.
149. T.D. Bullen, A.F. White, C.W. Childs, D.V. Vivit, and M.S. Schulz, “A Demonstration of Significant Abiotic Iron Isotope Fractionation in Nature,” Geology 194:39-51, 2001.
150. K.W. Mandernack, D.A. Bazylinski, and W.C. Shanks III, “Oxygen and Iron Isotope Studies of Magnetite Produced by Magnetotactic Bacteria,” Science 285:1892-1896, 1999.
151. B.L. Beard, C.M. Johnson, and L. Cox, “Iron Isotope Biosignatures,” Science 285:1889-1892, 1999.
152. K.H. Nealson and D. Saffarini, “Iron and Manganese in Anaerobic Respiration: Environmental Significance, Physiology, and Regulation,” Annu. Rev. Microbiol. 48:311-343, 1994.
153. K.H. Nealson and C. Myers, “Iron Reduction by Bacteria: A Potential Role in the Genesis of Banded Iron Formations,” Amer. Journal Sci. 290A:35-45, 1990.
154. B.L. Beard and C.M. Johnson, “High-precision Iron Isotope Measurements of Terrestrial and Lunar Materials,” Geochim. Cosmochim. Acta 63:1653-1660, 1999.
155. X.K. Zhu, R.K. O’Nions, and Y. Guo, “Secular Variation of Iron Isotopes in North Atlantic Deep Water,” Science 287:2000-2002, 2000.
156. B.L. Beard and C.M. Johnson, “High-Precision Iron Isotope Measurements of Terrestrial and Lunar Materials,” Geochim. Cosmochim. Acta 63:1653-1660, 1999.
157. X.K. Zhu, R.K. O’Nions, and Y. Guo, “Secular Variation of Iron Isotopes in North Atlantic Deep Water,” Science 287:2000-2002, 2000.
158. X.K. Zhu, R.K. O’Nions, and Y. Guo, “Secular Variation of Iron Isotopes in North Atlantic Deep Water,” Science 287:2000-2002, 2000.
159. B.L. Beard and C.M. Johnson, “High-Precision Iron Isotope Measurements of Terrestrial and Lunar Materials,” Geochim. Cosmochim. Acta 63:1653-1660, 1999.
160. X.K. Zhu, R.K. O’Nions, and Y. Guo, “Secular Variation of Iron Isotopes in North Atlantic Deep Water,” Science 287:2000-2002, 2000.
161. B.L. Beard, C.M. Johnson, and L. Cox, “Iron Isotope Biosignatures,” Science 285:1889-1892, 1999.
162. T.D. Bullen, A.F. White, C.W. Childs, D.V. Vivit, and M.S. Schulz, “A Demonstration of Significant Abiotic Iron Isotope Fractionation in Nature,” Geology 194:39-51, 2001.
163. S.L. Brantley, L. Liermann, and T.D. Bullen, “Fractionation of Fe Isotopes by Soil Microbes and Organic Acids,” Geology 29:535538, 2001.
164. K.W. Mandernack, D.A. Bazylinski, and W.C. Shanks III, “Oxygen and Iron Isotope Studies of Magnetite Produced by Magnetotactic Bacteria,” Science 285:1892-1896, 1999.
165. A.D. Anbar, J.E. Roe, and J. Barling, “Nonbiological Fractionation of Iron Isotopes,” Science 288:126-128, 2000.
166. T.D. Bullen, A.F. White, C.W. Childs, D.V. Vivit, and M.S. Schulz, “A Demonstration of Significant Abiotic Iron Isotope Fractionation in Nature,” Geology 194:39-51, 2001.
167. E.A. Schauble, G.R. Rossman, and H.P. Taylor, “Theoretical Estimates of Equilibrium Fe-Isotope Fractionations from Vibrational Spectroscopy,” Geochim. Cosmochim. Acta 31:2487-2497, 2001.
168. V.B. Polyakov and S.D. Mineev, “The Use of Mössbauer Spectroscopy in Stable Isotope Geochemistry,” Geochim. Cosmochim. Acta 64:849-865, 2000.
169. B.L. Beard, C.M. Johnson, and L. Cox, “Iron Isotope Biosignatures,” Science 285:1889-1892, 1999.
170. X.K. Zhu, R.K. O’Nions, and Y. Guo, “Secular Variation of Iron Isotopes in North Atlantic Deep Water,” Science 287:2000-2002, 2000.
171. S.L. Brantley, L. Liermann, and T.D. Bullen, “Fractionation of Fe Isotopes by Soil Microbes and Organic Acids,” Geology 29:535538, 2001.
172. A.D. Anbar, J.E. Roe, and J. Barling, “Nonbiological Fractionation of Iron Isotopes,” Science 288:126-128, 2000.
173. T.D. Bullen, A.F. White, C.W. Childs, D.V. Vivit, and M.S. Schulz, “A Demonstration of Significant Abiotic Iron Isotope Fractionation in Nature,” Geology 194:39-51, 2001.
174. K. Biemann, J. Oro, P. Toulmin III, L.E. Orgel, A.O. Nier, D.M. Anderson, P.G. Simmonds, D. Flory, A.V. Diaz, D.R. Rushneck, J.E. Biller, and A.L. Lafleur, “The Search for Organic Substances and Inorganic Volatile Compounds in the Surface of Mars,” Journal of Geophysical Research 82:4641-4658, 1977.
175. B.P. Weiss, Y.L. Yung, and K.H. Nealson, “Atmospheric Energy for Subsurface Life on Mars?” Proc. Natl. Acad. Sci. USA 97:1395-1399, 2000.
176. D.P. Glavin, M. Schubert, O. Botta, G. Kminek, and J.L. Bada, “Detecting Pyrolysis Products from Bacteria on Mars,” Earth and Planetary Science Letters 185(1-2) :1-5, 2000.
177. S.A. Benner, K.G. Devine, L.N. Matveeva, and D.H. Powell, “The Missing Organic Molecules on Mars,” Proc. Natl. Acad. Sci. USA 97:2425-2430, 2000.
178. See, for example, D.P. Glavin, J.L. Bada, K.L.F. Brinton, and G.D. McDonald, “Amino Acids in the Martian Meteorite Nakhla,” Proc. Natl. Acad. Sci. USA 96:8835-8838, 1999.
179. J.R. Cronin and S.Chang, “Organic Matter in Meteorites: Molecular and Isotopic Analyses of the Murchison Meteorite,” in The Chemistry of Life’s Origins, J.M. Greenberg, C.X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209258.
180. P.A. Bland and T.B. Smith, “Meteorite Accumulations on Mars,” Icarus 144:21-26, 2000.
181. A.S. MacKenzie, S.C. Brassell, G. Eglinton, and J.R. Maxwell, “Chemical Fossils: The Geological Fate of Steroids,” Science 217:491-504, 1982.
182. L.J. Allamandola, G.G.M. Tielens, and J.R. Barker, “Interstellar Polycyclic Aromatic Hydrocarbons: The Infrared Emission Bands, the Excitation/Emission Mechanism, and the Astrophysical Implications,” Astrophys. Journal 71:733-775, 1989.
183. J.R. Cronin and S. Chang, “Organic Matter in Meteorites: Molecular and Isotopic Analyses of the Murchison Meteorite,” in The Chemistry of Life’s Origins, J.M. Greenberg, C.X. Mendoza-Gómez, and V. Pirronello (eds.), Kluwer Academic Press, Boston, 1993, pp. 209258.
184. L.J. Allamandola, G.G.M. Tielens, and J.R. Barker, “Interstellar Polycyclic Aromatic Hydrocarbons: The Infrared Emission Bands, the Excitation/Emission Mechanism, and the Astrophysical Implications,” Astrophys. Journal 71:733-775, 1989.
185. D.P. Glavin and J.L. Bada, “Isolation of Amino Acids from Natural Samples Using Sublimation,” Anal. Chem. 70:3119-3122, 1998.
186. J.L. Bada and G.D. McDonald, “Detecting Amino Acids on Mars,” Anal. Chem. 68:668A-673A, 1996.
187. J.L. Bada and G.D. McDonald, “Amino Acid Racemization on Mars: Implications for the Preservation of Biomolecules from an Extinct Martian Biota,” Icarus 114:139-143, 1995.
188. L.D. Hutt, D.P. Glavin, J.L. Bada, and R.A. Mathies, “Microfabricated Capillary Electrophoresis Amino Acid Chirality Analyzer for Extraterrestrial Exploration,” Anal. Chem. 71:4000-4006, 1999.
































































