
4
Deciphering Past Climates—Reconciling Models and Observations
Forecasts of climate change for the next century are based on general circulation models (GCMs) that have been developed and tuned primarily using twentieth century records, but also with some input based on the understanding of the climate system from the more recent geological past (e.g., the Last Glacial Maximum and the Holocene). In part, this reflects the high levels of radiometric calibration and temporal resolution (subannual to submillennial) offered by near-time paleoclimate archives, which are capable of identifying the typically nonlinear components of the climate system—characterized by rapid response times—that are relevant to human society. A critical prerequisite for accurate forecasts of future regional and global climate changes based on GCMs, however, is the requirement that these models use parameters that are relevant to the future we seek to better understand. In this context, the recent climate archive captures only a small part of the known range of climate phenomena, since it has been derived from a time dominated by ice dynamics at both poles. Furthermore, the magnitude of forcing that the planet is now experiencing exceeds any that has occurred during the past ~30 million years. As GCMs are transformed into Earth System Models for the Intergovernmental Panel on Climate Change (IPCC) Fourth Assessment Report, they will encompass vastly more system physics, and deep-time climate studies will offer modelers the only real-world scenarios for testing the full climate response to the large increases in greenhouse gas levels that are projected.
As the climate system departs from the conditions captured by these well-studied near-time climate analogues, it is necessary for the scientific
community’s efforts to expand to capture the full range of variability and climate-forcing feedbacks of the global climate system, in particular for the past “extreme climate events” and warmer Earth intervals that may serve as analogues for future climate. Full testing of climate models for these time periods will require evaluation of feedback processes within models, enhanced spatial resolution, and longer simulations to better characterize climate model variability. All of these requirements, especially those for resolution and variability, will require significant computational resources.
For deep-time climate systems, the representation of paleogeographic boundary conditions can be a much greater source of uncertainty than it is for simulations based on modern geography. Furthermore, discrepancies between model outputs and paleoclimate observations may indicate the existence of additional processes, feedbacks, and/or sensitivities that are not present in the model or expressed in the modern climate system. For example, the exceptionally warm high latitudes during all past warm periods—whether transient or long term—cannot be reproduced by models without invoking unreasonable CO2 levels, revealing the inability of current models to fully capture the processes and feedbacks governing heat transport and retention or the processes that might generate heat in the polar regions under elevated atmospheric greenhouse gas levels (Covey and Barron, 1988; Rind and Chandler, 1991; Covey, 1991; Sloan and Pollard, 1998; Bice et al., 2006; Huber, 2008; Kump and Pollard, 2008; Spicer et al., 2008; Zachos et al., 2008). Thus, model development, which is based on improving specific processes and climate feedbacks and, in turn, evaluating the impact of these improvements on model simulations, depends on the availability of spatially resolved, robust, deep-time paleoclimate reconstructions of appropriate geochronological resolution and constraint. In addition, the utility of paleoclimate proxies for climate reconstruction and data-model comparisons relies on the proxies being sufficiently well preserved and the existence of an adequate understanding of the underlying processes, sensitivities, and uncertainties captured by these proxies.
Recent paleoclimate studies of deep-time successions have documented the potential of the older part of the geological record to reveal long-duration archives of forcings, responses, and long-term (centuries to tens of millennia) feedbacks that are of magnitudes and/or durations not captured by Pleistocene and Holocene paleoclimate records. Constraining the nature (e.g., rates, phasing between proxies) and origin (forcings) of climatic shifts, particularly rapid and/or transient events across climate thresholds from the deep-time record, will be greatly enhanced where orbital-scale cycles can be identified and resolved in the rock record (Box 4.1, Figures 4.1 and 4.2). Indeed, millennial to seasonal signals—at times calibrated to the orbital timescale—have been extracted from the sedimentary record spanning hundreds of millions of years (e.g., Feldman
BOX 4.1 Determining Time in the Geological Record
One of the biggest challenges in using the deep-time record for understanding Earth systems is determining the rates of processes and dating when specific events occurred. Determining rates requires very precise time control, particularly if the processes being studied occur at an ecological timescale (1 year to several centuries). One method for such precise time control is by using annually layered sediments in ancient anoxic or hyper-saline basins, as long as a few age control points are present. Examples include laminated sediments from the Pleistocene Santa Barbara Basin (Figure 4.1a), Eocene sediments of the North Sea (Figure 4.1b) (Schiøler et al., 2007), black shale sequences in the Cretaceous North Atlantic (Figure 4.1c), and the Permian Castile Formation of West Texas (Anderson, 1982). Quite highly resolved relative timescales can also be achieved using cyclic sequences, with resolutions of a few centuries to several tens of millennia, based on the identification of distinct orbital periods (Figure 4.1d; Figure 4.2). Both annual layers and orbitally tuned records can reveal ecological dynamics, snapshots of natural variability at different parts of Earth history, and the duration of threshold shifts in ecosystems.
Orbital cycles can sometimes be tied to astronomically tuned timescales during the past 40 million years to provide excellent absolute timescales. One example is the tuning of orbital cycles in glacial events during the Oligocene using combined geochemical and sediment property cycles tied to an astronomically calibrated timescale (Pälike et al., 2006a,b; see Figure 4.2). Orbital cycles have been recognized far back in the Phanerozoic sedimentary record and, together with high-resolution U-Pb dating, offer the potential to reconstruct Earth system dynamics in great detail (Erwin 2006; Davydov et al., 2010).
et al., 1993; Olsen and Kent, 1996; Eriksson and Simpson, 2000; Loope et al., 2001, 2004; Ivany et al., 2004; Wagner et al., 2004; Elrick and Hinnov, 2007; Jahren and Sternberg, 2008; Kennedy et al., 2009). The ability to precisely and accurately quantify geological time has improved dramatically with recent advances in radiometric dating and interlaboratory cross-calibration (e.g., the EARTHTIME initiative) permitting unprecedented temporal resolution (e.g., ID-TIMS [isotope dilution-thermal ionization mass spectrometry] uranium-lead [U-Pb] ages with analytical uncertainties of ≤0.01 percent; Ramezani et al., 2007). Some recent radiometric calibrations of the sedimentary record integrate astrochronology, providing Milankovitch-scale resolution through long intervals of time

FIGURE 4.1 Sediment cores can be detailed recorders of time: (a) Core sample (Core MV1012-TC-3) from the Holocene of Santa Barbara Basin, California, displaying well-developed millimeter-scale laminations related to annual cycles in biological productivity and sediment runoff. (b) Eocene laminated sediments from the North Sea (Nini-3 well), potentially offering very high resolution records of climate and ecosystem variability. (c) Similar laminations in a drill core through Cretaceous black shales from the tropical North Atlantic (from ODP Site 1259). (d) Orbital cycles in sediment color paced by the precession cycle (~21 kyr) from a drill core in the Paleocene of the western North Atlantic (from ODP Site 1051; Norris and Röhl, 1999).
SOURCE: ODP core images courtesy Integrated Ocean Drilling Program Science Services.
(e.g., Kuiper et al., 2008; Davydov et al., 2010). Furthermore, integration of orbital-stratigraphic approaches with bio-, magneto-, cyclo-, and/or chemostratigraphy has successfully placed high-resolution temporal constraints on past events (e.g., Olsen and Kent, 1996; Sageman et al., 2006; Westerhold et al., 2008; Adams et al., 2010; Galeotti et al., 2010). Several epochs and stages of the Phanerozoic have been fully orbitally tuned, presenting the possibility of geochronological resolution at 104- to 105-year scales (e.g., Hinnov and Ogg, 2007). Many of these records, however, await radiometric calibration to the absolute timescale.
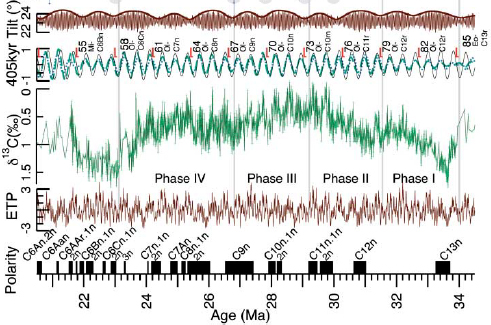
FIGURE 4.2 Astronomically tuned climate record from the Oligocene of the Central Pacific. Top two panels show the astronomical calculation of the 41,000-year cycle in the tilt of Earth’s spin axis (known as obliquity), and the 405,000-year cycle in eccentricity (thin black line). In the middle panel is the carbon isotope record of the deep Pacific which displays a well developed ~400,000-year cycle that closely matches the calculated astronomical cycle (dashed green line). ETP is the calculated cycle expected if the sediment record integrated the combined eccentricity, tilt and precession cycles. The geomagnetic polarity timescale is shown on the bottom, here calibrated to the astronomical cycles.
SOURCE: Modified from Pälike et al. (2006b).
The potential of the deep-time paleoclimate record to provide unique insight into scientific understanding of the climate system’s response to greenhouse gas forcing, however, is underdeveloped. To maximize this potential, the community is presented with three primary challenges:
• To determine precise chronologies for existing and new geological archives of paleoclimate interest—where feasible at the temporal resolutions that are possible in the Pleistocene and Holocene—through continued improvements in the precision and accuracy of geochronological techniques applicable to the sedimentary record (Ar-Ar [argon-argon] and U-Pb), and the development of novel radiometric approaches such as U-Pb dating of carbonates (Rasbury and Cole, 2009) and rhenium-osmium
(Re-Os) dating of black shales (Ravizza and Turekian, 1989; Selby and Creaser, 2005; McArthur et al., 2008a).
• To develop within such geochronological and/or orbitally tuned frameworks, marine and terrestrial—ultimately linked—time series of high temporal resolution and spatial density and distribution. For terrestrial records that are notoriously geographically fragmented and poorly resolved, this will rely in large part on systematically acquired, targeted continental drilling of continuous and highly resolved records.
• To obtain proxies of a broad range of surface and atmospheric conditions (paleotemperatures, pCO2, pO2, contents of other greenhouse gases, paleoprecipitation, seasonality, relative humidity)—of higher precision and accuracy than currently available—through some combination of proxy refinement, proxy development, and multiproxy studies.
CLIMATE MODEL CAPABILITIES AND LIMITATIONS
Current paleoclimate model capabilities (Box 4.2) include the application of fully coupled three-dimensional models to past climates. These are the same models that are used to study the present climate state and future changes to Earth’s climate, and they are the models that provide the modeling foundation for the IPCC periodic assessments. For many years, these models included only the physical aspects of the atmosphere, dynamic ocean, land, and sea ice components of the climate system. More recently, however, these models have begun to include coupling to a dynamic ice sheet model and prognostic components for biogeochemistry, atmospheric chemistry, dynamic vegetation, and ecology. Many models can even provide calculations of the isotopic composition of precipitation, making direct comparisons with δ18O marine and terrestrial proxies possible (Roche et al., 2006; Poulsen et al., 2007a, 2010; Zhou et al., 2008). Thus, global climate models have evolved from physical climate system models to more comprehensive Earth system models that permit more realistic coupling between the physical climate system and the biosphere (e.g., Slingo et al., 2009; Cadule et al., 2010).
In terms of the mean state, climate system models are able to realistically capture many characteristics of the current climate, such as observed equator-to-pole thermal gradients, large-scale spatial distribution of precipitation patterns, and various aspects of regional climate variability (e.g., El Niño-Southern Oscillation, Pacific Decadal Oscillation). Although the more comprehensive Earth system models offer many advantages, many aspects of regional-scale climates are still not captured accurately (although see Sewall and Sloan [2006], Thrasher and Sloan [2009], for exceptions). Simulating regional-scale phenomena requires the existence of high-resolution paleoclimate boundary data, which may not exist for
BOX 4.2 Climate Models
Climate models are numerical representations of the climate system that provide a means to study the processes that determine Earth’s climate state. Over the past 50 years, climate models have evolved to include a hierarchy of approaches to representing the climate system (Figure 4.3).
Geochemical box models are used to study the temporal evolution of quantities such as atmospheric CO2 and oxygen, ocean stable isotopes, and other geochemical variables. These models are based on theoretical expressions of the sources and sinks of a range of geochemical properties, providing global mean information on timescales of tens of thousands to millions of years.
Earth system models of intermediate complexity (EMICs) extend the box model concept to include spatial resolution and are useful tools to study Earth processes for timescales exceeding 10,000 years. Typically, these models include a detailed two- or three-dimensional ocean model coupled to simplified one- or two-dimensional atmospheric models. The energy balance atmospheric model predicts the geographic distribution of surface temperature and other energetic atmospheric quantities. The ocean model includes a marine biogeochemical component that simulates the chemical state of the ocean. The horizontal spatial resolution of these models is ~500 km. These models include detailed physical and biogeochemical processes that are often missing in the more complex models. However, their major limitation is that the atmospheric components are highly tuned to the present-day world, and they cannot incorporate realistic mechanical and thermodynamic forcing of the atmosphere on the ocean. Overall, these models are of value to look at transient climate change, such as the long-term fate of pCO2 over a few hundred thousand years.
Global climate models (GCMs) are the most comprehensive models for studying the climate system (IPCC, Fourth Assessment Report, Chapter 8, 2007). These models are usually composed of three-dimensional representations of the atmosphere, ocean, sea ice, and land systems. These systems are dynamically coupled and allow for feedbacks among the various components. Recently, these fully coupled Earth System Models have begun to include other processes such as atmospheric chemistry, terrestrial and marine biogeochemistry, and ecological models. With recent increases in computational power, atmospheric GCMs are now simulating the climate system on spatial scales of 50 to 75 km (Kim et al., 2008). Lower resolution versions of fully coupled GCMs (spatial resolutions of ~100 to 400 km) can be run for thousands of years (Liu et al., 2010), and this is especially important for deep-time climate research since the equilibrium time for the oceans is ~3,000 years (e.g., Kiehl and Shields, 2005). With continued
advances in computing as well as increases in the availability of massively parallel supercomputers, it is likely that multimillennial simulations using coarser-resolution GCMs (~400-km grid spacing) will soon be possible.
Such a hierarchy of climate models is essential for studying climate change on a wide range of timescales from decades to millions of years. Information from the more computationally expensive GCMs can be used as input for the EMICs, which can then be run for hundreds of thousands of years. Information from the GCMs or EMICs can also be used to better represent climate feedbacks in geochemical box models.
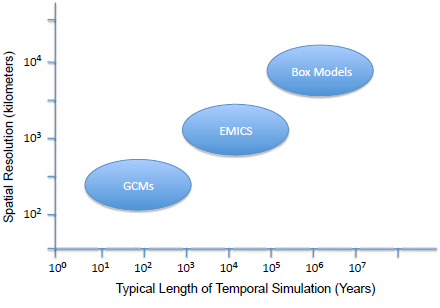
FIGURE 4.3 Graphical representation of the ranges of spatial resolution and simulation run times for the major categories of climate model.
such spatial resolutions (e.g., ~50 km). Additional challenges exist for the modeling of ancient climates, which are characterized by different paleogeography, paleotopography, atmospheric pCO2, solar luminosity levels, and other boundary conditions. Yet, the community’s confidence in the ability of GCMs to forecast future regional and global climate changes may be unfounded if these models cannot simulate past climates that dif-
fer as much from the present as the future is likely to differ from current conditions.
At this time, climate models of past periods use certain parameterizations defined for the present-day global climate system, necessarily requiring questionable assumptions about the relevance of present-day conditions to the older parts of the geological record. For example, assumptions concerning plant physiology, biome composition, and surface distribution based on present-day vegetation are not likely to have been applicable to times prior to the evolution of angiosperms (~130 million years ago [Ma]) and the expansion of grasses (~34 Ma). Similarly, the emission of precursor gases that form atmospheric aerosols is linked to current understanding of atmospheric chemistry because proxies for paleoemissions of gases that could create aerosols, in particular for those that can affect cloud properties such as biologically mediated dimethyl sulfide (Henriksson et al., 2000; Kump and Pollard, 2008), do not exist at this time. Thus, the definition of boundary conditions inferred from the geological proxy record and the parameterization of physical processes in warmer world models intrinsically come with significant uncertainty.
Key boundary conditions that must be prescribed for deep-time or warm world models include the paleogeography of land areas, past vegetation distributions, paleotopography, and ice sheet extent. Coupling to ocean models additionally requires knowledge of paleoeustasy in order to specify the distribution of shallow seas and the paleobathymetry for the deep oceans:
• Global paleogeography is an important boundary condition for constraining elevation models of continental topography and oceanic bathymetry, the geography of oceanic gateways and shallow continental (epeiric) seas that influence oceanic circulation, ocean heat transport, and climatic conditions (e.g., Crowley and Burke, 1998). The relative positions of the continents are well known back to ~200 to 180 Ma, when the oldest extant ocean floor was formed, but the uncertainty of deeper time paleogeographic reconstructions increases dramatically going farther back in time because of the absence of a seafloor record (Ziegler et al., 1983; Scotese, 2004; Blakey, 2008). Paleolatitudes can often be resolved to within about ±5°, which is somewhat coarser than the geographic resolution of the global climate models used for recent IPCC simulations of future climate. However, a greater concern is that the lack of geological record of intervening ocean basins, particularly prior to the Cretaceous, means that longitudinal control is not possible.
• Paleotopography is an important boundary condition for predicting stationary wave patterns in the atmosphere, convective effects associated with uplifted regions, and their impact on the distribution of
precipitation. Promising new approaches to reconstructing paleoaltimetry have been developed in recent years. The application of the fossil leaf stomata index to paleoaltimetry is based on the established relationship between leaf stomata frequency and ambient pCO2 and the predictable, globally conserved decrease in pCO2 with altitude (McElwain, 2004). The uncertainty, which can reach levels as large as the potential height of the surface (Peppe et al., 2010), is determined by the uncertainty in the CO2 concentration in air as a function of time. The oxygen isotope compositions of pedogenic minerals (Rowley and Currie, 2006; Forest, 2007; Sahagian and Proussevitch, 2007) and the hydrogen isotope composition of n-alkanes from epicuticular plant waxes preserved in lacustrine deposits (Polissar et al., 2009) may be sensitive proxies of surface water and precipitation compositions and, in turn, paleoelevation through isotope-altitude relationships. As this proxy is based on systematic trends in the distribution and isotopic composition of modern precipitation with climate and topography, the uncertainty in estimates is dependent on the degree to which the isotopic composition of paleoprecipitation is faithfully recorded by the authigenic (formed in situ) minerals (Blisniuk and Stern, 2005). The recently developed “clumped-isotope” carbonate paleothermometer (see discussion below) shows good potential for paleoelevation reconstruction, using assumed temperature lapse rates with elevation (Ghosh et al., 2006; Quade et al., 2007). Recent studies have demonstrated that surface uplift influences the regional climate and isotope distribution and thereby severely complicates paleoaltimetry interpretations (e.g., Ehlers and Poulsen, 2009; Poulsen et al., 2010). Therefore, ultimately, the accurate reconstruction of paleotopography requires some degree of iteration between modeling and proxy methods since topographic relief strongly affects regional climate patterns, influencing all of the proxy-based estimates (Ehlers and Poulsen, 2009).
• Paleobathymetry has been reconstructed back to the late Jurassic through well-known age-depth relationships for oceanic crust (Parsons and Sclater, 1977; Stein and Stein, 1992), but paleobathymetry for older parts of the record is far more challenging to constrain because of the loss of seafloor through subduction. The development of oceanic plateaus and oceanic swells, and uncertainty in how the rate of ocean floor production has varied over time (Rowley, 2002; Stein and Stein, 1997), further challenges reconstructions of paleobathymetry and hence sea level change (Kominz, 1984). Besides eustasy, uncertainties in estimating changes in the rate of ocean floor production also impact estimates of mantle outgassing that have been used to drive carbon cycle models and delineate the evolution of atmospheric pCO2, pO2, and CH4 through the Phanerozoic (Berner, 2006, 2009; Beerling et al., 2009). This uncertainty is significant considering that for the time intervals for which seafloor is preserved (≤180 Ma), the rate of change of seafloor production remains a debated issue (Rowley, 2002).
• The Earth’s vegetation contributes to, and is affected by, a variety of important climatic feedbacks and is thus an important boundary condition for paleoclimate modeling. Changes in vegetative land cover directly influence albedo and Earth surface radiation (Betts, 2000; Chase et al., 2000; Pielke et al., 2002; Marland et al., 2003; Horton et al., 2010). Changes in terrestrial vegetation also lead to changes in evapotranspiration and the hydrological cycle (Shukla and Mintz, 1982; Rind et al., 1990; Baldocchi et al., 2000; Alpert et al., 2006), with attendant indirect effects on radiative fluxes and atmospheric chemistry as a function of changes in cloud cover and water vapor mass in the atmosphere (Elliott and Angell, 1997; Hennessy et al., 1997). In addition, vegetative land cover both influences and is influenced by soil moisture content, and changes in soil moisture content can have significant climatic effects through shifts in the relative influences of latent heat flux versus sensible heating (Alpert et al., 2006; Niyogi and Xue, 2006). Vegetation-climate feedbacks have not yet been fully incorporated into GCMs because of the difficulty of parameterizing the complex, nonlinear interactions that range from cellular scale in physiological approaches to regional or global scale in biome and physical approaches to plant definition (Alpert et al., 2006). GCMs incorporating vegetation-climate feedbacks, however, generally yield amplified climate responses such as higher climate sensitivities (up to 5.5°C per CO2 doubling; Cox et al., 2000), larger amplitudes of paleoglacioeustasy (Horton et al., 2010), and greater high-latitude amplification of warming (DeConto et al., 1999) relative to models lacking such feedbacks. The deep-time geological record offers several large-scale “natural experiments” (see Chapter 2) from which insights regarding the operation and scaling of these vegetation-climate feedbacks can be gleaned (Peteet, 2000; Parmesan and Yohe, 2003; Horton et al., 2010). The knowledge of the composition and spatial distribution of vegetation on a global scale, prior to the evolution of angiosperms (Early Cretaceous) and grasslands (Cenozoic), however, must be further developed. A far more coordinated effort is needed to expand scientific understanding of fossil plant physiological mechanisms and to synthesize disparate paleobotanical data into more comprehensive and temporally constrained compilations that can be used to refine dynamic vegetation models for climate modeling.
Further improvements in scientific knowledge of these physical and ecological boundary conditions will require more detailed analysis of paleomagnetic, paleoclimatic, paleotectonic, and paleontologic data (Van der Voo, 1993; Parrish, 1998; Kiessling et al., 1999), as well as development of more sophisticated geodynamic models. For example, there is growing evidence for a systematic shallow bias (5-10°) in paleomagnetic data from the sedimentary record (Tauxe and Kent, 2004), increasing the paleolatitudinal
uncertainty of reconstructions developed using these data. Recent mantle flow simulations suggest that estimates of long-term (10 to 100 million years) eustatic sea level changes and the extent of continental flooding based on seismic and backstripping stratigraphic analysis of continental margin successions may reveal substantial mantle flow-induced dynamic topography on passive margins (Moucha et al., 2008). Furthermore, high-precision geochronological data from depositional and igneous systems worldwide are critical to constrain the ages of key paleogeographic events.
In addition to the need to better constrain the physical boundary conditions for global climate models applied to deep-time climates, the concentration of greenhouse gases (in particular CO2) in ancient atmospheres and the solar and spectral irradiance must be determined given their fundamental contribution to radiative forcing of the climate system. The current range of uncertainty in atmospheric CO2 and the near-complete lack of knowledge of other greenhouse gases (e.g., atmospheric methane) for many geological time periods lead to significant uncertainty in the radiative forcing of the climate system for these time periods. Improved proxies for atmospheric greenhouse gases are needed to narrow the uncertainty in the radiative forcing of the climate system.
The evolution of the solar irradiance is well constrained by solar theory over timescales of millions of years. However, variations of total solar irradiance are uncertain on timescales ranging from multidecades to multimillennia. The evolution of spectral irradiance, however, is not well constrained over geological time. Changes in spectral radiance affect the vertical deposition of shortwave energy in the atmosphere and the chemical composition of the atmosphere through photolysis processes (e.g., the formation and destruction of ozone). At present, there is no method to determine how the spectral distribution of solar irradiance has changed in the past. Finally, the latitudinal distribution of solar radiation is determined by sun-Earth geometry, and detailed celestial mechanical calculations for the temporal change of obliquity, eccentricity, and precession are limited. Clear signatures of Milankovitch cycles appear in deep-time records, but the chaotic behavior of the solar system means that Earth’s orbital variations can be computed with precision for only about the past 40 million years (Laskar et al., 2004; Pälike et al., 2004).
INDICATORS OF CLIMATE SENSITIVITY THROUGH TIME—PROXIES FOR CO2 AND MARINE TEMPERATURES
Atmospheric CO2 Proxies
Although the physical radiative forcings from greenhouse gases are well known, anticipating the consequential rise in temperature is a much
more difficult challenge because of the temporal hierarchy and complexity of feedbacks (Figure 4.4) that are triggered by CO2 perturbation (Hansen et al., 2008). Although climate sensitivities incorporate a number of major feedbacks, reconstructions of global warming during past periods of CO2 release indicate that there are additional short- and long-term feedbacks influencing temperature increases during CO2-forced climate change (e.g., Zeebe et al., 2009). Significant uncertainty accompanies future climate sensitivities (~1.5 to 6°C per doubling of CO2), with the possibility that current projections underestimate future temperatures. Calibration of climate sensitivity at “deep-time” temporal scales, which account for climate conditions that ultimately are anticipated to result from anthropogenic CO2 release and for longer-term feedbacks that apply in warmer worlds, will be highly applicable for improved climate projections for the latter half of the twenty-first century and beyond.
Climate sensitivities on deep timescales have been estimated based on paired data for sea surface temperatures (SSTs) as well as model and proxy-based CO2 reconstructions (e.g., Pagani et al., 2006; Royer et al., 2007; Zachos et al., 2008). For example, climate sensitivity for the transient Paleocene-Eocene Thermal Maximum (PETM) warming has been constrained using paleo-CO2 and paleotemperature estimates inferred from stable isotopic values of marine carbonates and biomarkers—these place the climate sensitivity during the greenhouse gas-forced event between a lower bound of 2.4-3.0°C and as high as ~4°C (Pagani et al., 2006; Panchuk et al., 2008).
Estimates of paleoatmospheric pCO2 beyond the range of ice cores are based on geochemical carbon models and an evolving toolbox of fossil organic and mineral matter proxies. Each of these methods is characterized by unique strengths, yet is limited by uncertainties and sensitivities reflecting the assumptions underlying their approach (summarized in
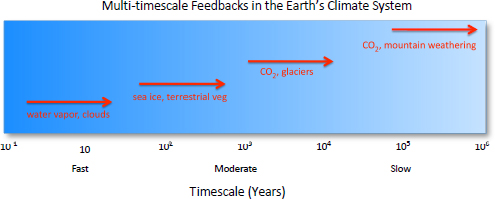
FIGURE 4.4 Schematic representation of the range of feedback scales that act on Earth’s climate system.
Royer et al., 2001a; also see Box 4.3 and Table 4.1). Geochemical mass balance models (e.g., GEOCARB and GEOCARBSULF; see Berner and Kothavala, 2001; Berner, 2006) that quantify carbon transfer rates within the long-term carbon cycle have long provided a first-order assessment of how paleo-pCO2 (and O2) evolved throughout the Phanerozoic; many intervals have been tested and refined by comparison to proxy-based CO2 estimates of higher temporal resolution (e.g., Ekart et al., 1999; Pagani et al., 2005; Montañez et al., 2007; Barclay et al., 2010).
The highest-precision paleoatmospheric pCO2 estimates, in particular for the Cenozoic and Cretaceous, are based on proxy methods that utilize fossil marine and terrestrial photosynthetic flora:
• The alkenone paleobarometer, which uses the δ13C of lipid biomarkers derived from haptophyte algae in marine sediments (Freeman and Hayes, 1992; Pagani et al., 1999), and
• The stomatal-index method to estimate paleo-pCO2 from fossil leaf stomata (Woodward, 1987; McElwain and Chaloner, 1995).
Both of these proxy methods yield pCO2 estimates with low uncertainties (tens to <100 ppmv), although marine proxy-based estimates tend to be toward the lower end of the range, while estimates inferred using terrestrial plants are generally higher (see Box 4.3). The sensitivity of these proxy methods, however, decreases dramatically above ~1,000 ppmv (±200 ppmv) due to CO2 saturation (Bidigare et al., 1997; McElwain and Howarth, 2009).
These fossil proxy approaches, however, are based on calibrations using extant taxa or their nearest living relatives, and require the assumption that modern organisms can be used to represent biological responses of extinct organisms to ancient environments—an assumption that has been challenged. For the alkenone paleobarometer, interpretation of the photosynthetic carbon isotope effect assumes constancy in the size of phytoplankton haptophyte cells, which may have varied on millennial timescales (Henderiks and Pagani, 2007). In the case of fossil vascular plants in greenhouse periods, it is unclear how taxonomic differences and greater stomatal conductance induced by elevated temperatures (Helliker and Richter, 2008) would impact their regulation of gas exchange (Franks and Beerling, 2009). Most recently, it has been suggested that bryophytes, which lack a carbon-concentrating mechanism, may be a more reliable pCO2 proxy (Fletcher et al., 2005, 2008), with applications to the Pleistocene (White et al., 1994) and much deeper time (Fletcher et al., 2008), yielding CO2 estimates that are in accord with other proxy estimates of pCO2.
Independent, mineral-based pCO2 values, to which organic proxy and geochemical model estimates can be compared (Box 4.3), have been
BOX 4.3 Miocene Climate Change—CO2 or Ocean Trigger?
The Miocene was a pivotal time in the current icehouse, marking the transition between fluctuating glacial conditions and rapid expansion of the Antarctic ice sheet under a cooler climate (Zachos et al., 2001a; Shevenell et al., 2004). Variations in oceanic heat transport and atmospheric vapor transport, governed by changes in deep-ocean circulation and oceanic gateways as well as decreasing pCO2, have been implicated as drivers of this late Cenozoic global cooling (Raymo, 1994; Flower and Kennett, 1995; Holbourn et al., 2007). Recent coupled ice sheet-climate model simulations suggest that the Miocene climate transition was largely CO2induced, involving the crossing of a threshold pCO2 of 400 parts per million by volume (ppmv) (Langebroek et al., 2008). Overall, estimates of pCO2 during the Miocene range from 140 to more than 700 ppmv (Table 4.1), illustrating the critical need for improved CO2 proxy methods (Tong et al., 2009). Moreover, contrasting estimates of atmospheric CO2 exist for the transient period of mid-Miocene warmth (~17 Ma; compare marine-based estimates of Pagani et al. [2005], and vascular plant-based estimates of Kürschner et al. [2008]) confound the precise relationship between temperature and CO2.
TABLE 4.1 Estimates of Atmospheric CO2 Levels for the Middle Miocene
| Reference | CO2 (ppmv) | Method | Uncertainties |
| Pearson and Palmer (2000) | 140 to 300 | Marine δ11B | ~20% at low CO2, very high at >500 ppmv |
| Pagani et al. (1999) | 180 to 290 | Marine δ13C (alkenone carbonate); marine δ18O | ~20% near modern ocean conditions, infinite at CO2 ≥ 2,000 |
| Royer et al. (2001b) | 300 to 450 | Leaf stomatal indices | ppmv ~20% at low CO2, very high at >500 ppmv |
| Kürschner et al. (2008) | 300 to 600 | Stomatal frequency data from tree species | ~20% at low CO2, very high at >500 ppmv |
| Cerling (1991) | <700 | Paleosol carbonate δ13C | 50-100% |
SOURCE: Modified from Tong et al. (2009); uncertainty estimates based on Hansen et al. (2008); Pagani et al. (2005); Royer et al. (2001b); and Cerling (1991).
calculated for much of the past 400 million years. For the Cenozoic, boron isotopes of foraminiferal calcite (assumed to mirror that of δ11B of borate in seawater) have been used as a proxy for seawater pH and pCO2 (e.g., Sanyal et al., 1997; Spivack and You, 1997; Foster, 2008; Pearson et al., 2009). Where the δ11B proxy has been tested against alkenone-based pCO2, there is a good agreement (e.g., Seki et al., 2010). In contrast, estimates of Mesozoic and Paleozoic pCO2 are based largely on soil carbonate and goethite paleo-CO2 proxies. The carbonate-based paleo-pCO2 barometer (Cerling, 1991, 1992), however, is particularly sensitive to soil CO2, a parameter that is challenging to constrain in fossil soils (Ekart et al., 1999; Breecker et al., 2009) and varies with soil moisture and productivity. Soil-formed iron oxides (goethite, gibbsite) provide a complementary mineral paleobarometer, because they form in soils that do not accumulate carbonate and the carbon isotope composition and content in their ferric carbonate component (Fe(CO3)OH) are typically diagenetically robust (Yapp and Poths, 1992; Schroeder and Melear, 1999;Yapp, 2004; Tabor and Yapp, 2005). Despite the large uncertainties associated with mineral-based pCO2 estimates, comparison of mineral- and plant-based estimates are important because of their complementary differences in sensitivity—plant-based proxies lose sensitivity above 800 to 1,000 ppmv, whereas mineral-based proxies are more sensitive above 1,000 ppmv (Royer et al., 2001a)—and the lack of extant plant calibrations for stomatal index-based estimates in pre-Cretaceous intervals.
Marine Temperatures
A major challenge in determining ancient climate sensitivity is the need for robust estimates of ocean temperatures and latitudinal ocean temperature gradients. This need is particularly great for climate reconstructions of the pre-Cretaceous, for which we presently have only binary “icehouse or greenhouse” reconstructions (Figure 1.2) and highly interpretive and largely unpublished climate syntheses (e.g., PALEOMAP Project1). Several proxies have been employed to reconstruct ancient sea surface and deep-water temperatures, each with its strengths and limitations. Measuring multiple proxies not only provides refined SST reconstructions but also offers constraints on taxonomic (vital), environmental, and diagenetic influences that might affect each proxy. For several decades, the gold standard for reconstructing sea surface and bottom temperatures for the post-Jurassic has been from δ18O values of foraminifera (e.g., Shackleton, 1987; Zachos et al., 2001a), whereas for older geological intervals, it has been the δ18O values of metazoan skeletal calcites (Veizer
________________________
et al., 1999; Grossman et al., 2008) and conodont apatites (Joachimski et al., 2006; Buggisch et al., 2008). There is, however, growing appreciation for the susceptibility of fossil carbonate to diagenesis and thus overprinting of the precipitation (seawater) temperature signal in its δ18O values (Box 4.4) (Schrag et al., 1995; Pearson et al., 2001, 2007; Came et al., 2007; Grossman et al., 2008; Kozdon et al., 2009; Cochran et al., 2010). Furthermore, carbonate δ18O values are influenced by both temperature and seawater δ18O, which in turn has varied through time due to varying ice sheet δ18O composition, their effects of waxing and waning ice sheets on seawater δ18O, and surface net evaporation balance, in particular in the broad, shallow epicontinental seas of the pre-Cenozoic (Holmden et al., 1998; Veizer et al., 1999; Panchuk et al., 2006). The compound influence of changing seawater δ18O and temperature on carbonate δ18O values for periods that were not ice-free and the possible effects of seawater alkalinity on carbonate δ18O (Spero et al., 1997; Zeebe, 1999) have led to increased focus on independent temperature proxy methods.
The Mg/Ca ratios of planktonic foraminiferal calcite have been shown to be sensitive to temperature, increasing exponentially with increasing SSTs, providing an SST paleothermometer (e.g., Elderfield and Ganssen, 2000; Lear et al., 2000). Application of the Mg/Ca temperature proxy to deep-sea sediments has challenged climate change paradigms by documenting dynamic glacial-interglacial temperature variation in Pleistocene tropical oceans and linkages to extratropical climates, and refining phasing between changes in atmospheric CO2, surface and deep-ocean temperature changes, and Antarctic glaciation (Lea et al., 2000; Zachos et al., 2008). Mg/Ca paleothermometry, however, requires species-specific calibration to account for environmental and vital (taxonomic) effects and may be limited by salinity, pH, and/or carbonate ion effects on magnesium partitioning in foraminiferal calcite (Dekens et al., 2002, 2008; Lear et al., 2002, 2008; Russell et al., 2004; Coxall et al., 2005; Ferguson et al., 2008; Hoogakker et al., 2009). Importantly, recent multiproxy reconstructions of deep-sea sediments document good agreement between alkenone-based and Mg/Ca temperature estimates (Bard, 2001; Dekens et al., 2008). In addition to deconvolving the temperature and ice volume signals in the isotopic record, integrating foraminiferal δ18O values and Mg/Ca ratios has permitted the reconstruction of oceanic surface salinity distributions and their effect on oceanic heat transport (e.g., Schmidt et al., 2004). Efforts to apply the Mg/Ca temperature proxy to pre-Cenozoic biotic calcites (e.g., mollusks) have been limited by the influence of vital effects in these calcifying organisms (Immenhauser et al., 2005), variation in seawater alkalinity and [CO32–], and the need to account for likely secular variation in seawater Mg/Ca and ocean saturation state.
BOX 4.4 Proxies and the “Tropical Climate Paradox”
Proxy evidence for mid- to high-latitude warmth in both the marine (Crowley and Zachos, 2000; Zachos et al., 2001a) and the continental (Greenwood and Wing, 1995; Wolfe, 1995; Markwick, 1998) realms of the Early Eocene (57-50 Ma) are consistent with global warmth and a high-CO2 atmosphere (Pearson and Palmer, 2000; Pagani et al., 2005). However, marine isotope data from low paleolatitudes initially indicated little or no ocean surface warming, and perhaps even cooling, in the Eocene tropics (Shackleton and Boersma, 1981; Barron, 1987), leading to what was termed the “cool tropics paradox” (Barron, 1987). Subsequently, scientists have revisited the low-latitude planktonic foraminiferal record (Pearson et al., 2001, 2007; Norris et al., 2002; Kozdon et al., 2009), arguing that diagenesis on the cold deep seafloor has imparted a cool overprint signal to the oxygen isotopic composition of warm-water planktonic foraminifera, biasing the record in the tropics toward cold temperatures.
The tropical climate paradox may be fully resolved with new estimates of tropical SST for the Eocene derived from pristine foraminifera and a new organic molecule proxy, TEX86,a that indicate temperatures 5-10°C warmer than previous reconstructions (Pearson et al., 2007) (Figure 4.5). Such paleotemperature reconstructions further challenge the tropical thermostatic regulation hypothesis presented in Chapter 2 and imply that the cooling trend of the Eocene was primarily a high-latitude phenomenon with little effect on the tropics, where climate remained warm and stable (Pearson et al., 2007).
________________________
aTetraether index of 86 carbon atoms; paleothermometer based on the composition of membrane lipids of marine picoplankton.
Several additional proxy methods show promise for paleothermometry but are still in the development and testing phases:
• The calcium isotope (δ44Ca) composition of well-preserved foraminifera may provide an independent paleotemperature proxy and a test of the reliability of the Mg/Ca proxy (e.g., Nägler et al., 2000), although the complex calcium isotope fractionation behavior, ancient seawater δ44Ca, and possible vital effects during biocalcification are not yet fully understood (Gussone et al., 2009). Limited δ44Ca data for Cretaceous rudist
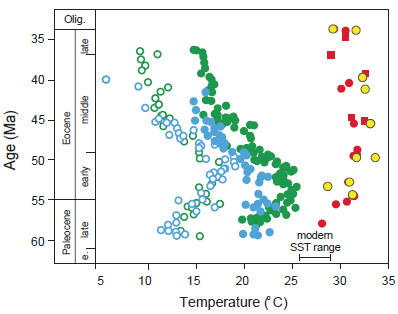
FIGURE 4.5 Tropical paleotemperature estimates based on δ18O data from unaltered benthic and planktonic foraminifera (red circles and squares, respectively) and from TEX86 data (yellow circles) from onshore drill core samples from Tanzania. These are plotted for comparison with benthic (open circles) and planktonic (green circles) foraminifera δ18O data from ODP cores collected from the tropical Pacific Ocean, inferred to have been diagenetically altered and thereby indicating anomalously low paleotemperature estimates.
SOURCE: Pearson et al. (2007).
calcite demonstrate the potential for extending this temperature proxy to pre-Cenozoic calcitic macrofauna (Immenhauser et al., 2005). Similarly, the magnesium isotope (δ26Mg) composition of aragonite corals shows promise as a seawater paleothermometer (Wang et al., 2008).
• Recent studies have documented that the clumping (ordering) of carbon and oxygen isotopes into bonds in biogenic and abiotic carbonates is temperature dependent and an independent record of the fluid δ18O in which the carbonate precipitated or stabilized during diagenesis (“clumped isotope method”; Came et al., 2007; Eiler, 2007; Tripati et al.,
2010). The carbonate clumped isotope proxy holds considerable promise for constraining marine and continental (e.g., speleothems, vertebrate bioapatite) paleotemperatures, although kinetic and diagenetic effects must be better understood (Affek et al., 2008; Came et al., 2008; Eagle et al., 2010).
• Biomarker proxies permit reconstruction of paleo-SSTs that are independent of mineral-based proxy estimates, and include UK37’ (Brassell et al., 1986), which is based on the relative abundances of C37 alkenones photosynthesized by marine green algae, and the more novel biomarker, TEX86, which is based on the relative abundances of C86 tetraether lipids that form in the water column by archaeal microbiota (Schouten et al., 2002; Eglinton and Eglinton, 2008). The low diagenetic susceptibility of these biomarkers and the preservation of tetraether lipids in sediments as old as the Cretaceous have provided new insight into the climate dynamics of the recent icehouse (Haug et al., 2004; Kienast et al., 2006; Martrat et al., 2007; Dekens et al., 2008) through Cretaceous and Eocene greenhouse periods, including contributing to the resolution of the Cretaceous cool tropics paradox (see Box 4.4) (Pearson et al., 2007; Schouten et al., 2007). Integration of multiple proxies greatly increases the range of paleotemperature sensitivity and probably also increases the precision of estimates because of the variable sensitivity of different proxies. For example, the Mg/Ca proxy is least sensitive at low temperatures, whereas the Uk’37’ method is least sensitive at high temperatures.
INDICATORS OF REGIONAL CLIMATES
Climate models and paleoclimate archives indicate that one of the larger impacts of global warming is likely to be regional changes in continental temperatures and precipitation. It is thus imperative to constrain past temporal and geographic variability in continental climate—in particular for periods of abrupt and/or major climate transitions and for climates that were warmer than the present day—in order to better understand how regional climates may change in the future. For continental settings, deep-time paleoclimate reconstructions require a multiproxy approach involving comparable proxies.
Estimating Continental Temperatures
There are numerous and reasonably well-developed proxies for estimating continental paleotemperatures from lacustrine, coastal, and terrestrial deposits. Fossil plant leaves and pollen have been the major source of continental paleotemperature estimates, because the composition and physiological properties of plant communities change rapidly
with temperature (and relative humidity) changes. For flowering plants (angiosperms), the shapes (style of leaf margin) and sizes (physiognomy) of fossil leaves have been shown to vary with mean annual temperature (MAT) (Wolfe, 1993; Wilf, 1997; Kowalski and Dilcher, 2003; Royer et al., 2005) and have been applied throughout the Cenozoic, including for the PETM (Wing et al., 2005). Plant-based continental temperature estimates for periods that predate the evolution of angiosperms (pre-Cretaceous), however, are missing. Estimates of paleocontinental temperatures also have been interpreted from pollen distributions in lake sediments, through comparison with the temperature sensitivity of modern plant communities (Overpeck et al., 1985). The pollen distribution approach, however, becomes significantly less certain on longer timescales due to increased differences among ancient and modern plant species and communities.
Mineral-based isotopic paleothermometry offers an independent set of proxies that are not restricted stratigraphically to the post-Jurassic, and for which interpretations are not limited by lack of calibration to extant plants. For example, the δ18O values of pedogenic carbonates and δ18O and δD values of hydroxylated clay minerals (kaolinite and smectite) and iron oxides (goethite and hematite) from fossil soils are being used to estimate paleotemperatures in ancient soils hundreds of millions of years old (Dworkin et al., 2005; Tabor and Montañez, 2005), although this approach requires assumptions regarding the stable isotope composition of meteoric water. This limitation can be overcome for soil carbonates by application of the carbonate clumped isotope thermometer to paleosol carbonates (Passey et al., 2010) and through oxygen isotope–mineral pair thermometry between coexisting pedogenic clays and iron oxides (Tabor, 2007). Both methods allow for paleosoil temperature estimates that are independent of the soil water δ18O in which the minerals formed. Conventional δ18O and clumped isotope analysis of vertebrate bioapatites (and δ18O analysis of body scales of some freshwater fish) offer yet another independent proxy of continental MAT and have been used to reconstruct MAT geographic patterns for deep-time warm periods (e.g., Koch et al., 2003; Fricke and Wing, 2004) and to constrain body temperatures of extant and extinct vertebrates (e.g., Barrick and Kohn, 2001; Eagle et al., 2010).
In addition to isotopic paleothermometry, the major element compositions of paleosols have been used for MAT reconstructions as far back as the Paleozoic, based on transfer functions calibrated using modern soils and associated mean annual temperatures (Sheldon et al., 2002; Retallack, 2005). Applications of this proxy yield estimates of Cenozoic paleo-MAT that are consistent with fossil leaf morphology-based paleotemperature estimates (Sheldon, 2009). For all of these mineral-based proxies, the accuracy and uncertainty of paleotemperature estimates are largely dependent on using appropriate fossil material that has been minimally altered by diagenesis.
Ultimately, reconstructions of regional variation in continental temperatures will require further calibration studies of existing proxy methods along with the development of new higher-resolution proxies and continued development of spatially highly resolved multiproxy datasets. One promising new direction is the use of biomarkers preserved in lacustrine and marginal marine sediments that appear to be diagenetically relatively robust. The presence of picoplankton members of the Archaea in ancient lake deposits opens up the possibility of using the relative abundances of their membrane lipids (TEX86) as a paleothermometer of surface water (Eglinton and Eglinton, 2008). Microbial lipid patterns in modern soils also show potential as a biomarker paleotemperature proxy, but the influence of other soil parameters on their abundances (e.g., pH) requires further calibration studies.
Estimating Regional Hydroclimates
The understanding of regional patterns for continental paleoclimate changes in the younger part of the record comes primarily from a wealth of paleolacustrine records and a rapidly increasing speleothem paleoclimate database. Lacustrine records offer the continuity and temporal resolution potential to provide key high-resolution sedimentological, geochemical, and paleontologic time series for reconstructing paleocontinental regional climate change. The paleo-water balance of some ancient lakes, such as those in the western United States, also provides strong signals of regional changes in effective moisture (e.g., Benson et al., 2003). Speleothem physical and geochemical proxies are proving to be powerful recorders of changes in regional air temperature and effective moisture (e.g., Oster et al., 2009; Wagner et al., 2010). Notably, precisely dated (U-series) speleothem records are revealing rapid—century to subdecadal—climate anomalies, often involving inter- and intrahemispheric atmospheric teleconnections (Wang et al., 2001, 2005; Yuan et al., 2004). However, the bulk of these continental records are from the late Cenozoic, and thus have not been used to reconstruct regional hydroclimate patterns during warm Earth climates.
Reconstructing regional patterns in relative humidity and precipitation is far more challenging in deep-time records because of the overall lower levels of temporal and spatial resolution, stratigraphic continuity, and geochemical susceptibility to diagenesis, although several new approaches are being evaluated. For much of the pre-Neogene, scientific understanding of climate regimes is based on low spatial and temporal resolution global syntheses of published databases (Ziegler et al., 2003; Boucot et al., 2004). The morphological characteristics of ancient soils and their bulk geochemical composition have climatic significance because the intensity of pedogenesis is dominantly related to precipita-
tion patterns and surface temperature. Quantitative proxies for estimating mean annual precipitation have been developed that use the iron content in pedogenic Fe-Mn nodules, the depth to the pedogenic carbonate horizon, or the chemical composition of particular paleosol horizons (known as the Chemical Index of Alteration, CIA), all of which are based on empirical relationships derived from modern soils (Stiles et al., 2001; Sheldon et al., 2002; Retallack, 2005; Sheldon and Tabor, 2009). These approaches have been applied to a wide age range of Phanerozoic paleosols (e.g., Driese et al., 2005; Prochnow et al., 2006), including to the PETM where these soil proxy-based precipitation estimates indicate a transient drying in western North America associated with transient global warming (Kraus and Riggins, 2007). Where multiproxy records permit, CIA-based estimates of mean annual precipitation are consistent with independent paleobotanical estimates. The measured δ18O compositions of ancient soil-formed minerals (phyllosilicates, carbonates, iron oxides, and sphaerosiderites) have been shown to be reliable proxies of soil-water δ18O and, in turn, δ18O precipitation at a given paleolatitude after consideration of evaporative or nonequilibrium effects (Stern et al., 1997; Yapp, 2000; Vitali et al., 2002; Ufnar et al., 2004; Tabor and Montañez, 2005). The fact that these minerals form in equilibrium with ambient hydrological conditions means that they provide a sensitive record, where formed, of shifts in seasonality and precipitation rates, allowing them to be used to evaluate the hydrological cycle in past greenhouse worlds and periods of icehouse-to-greenhouse transition.
In the same way that marine and lacustrine biomarkers have been used as quantitative paleothermometers, the hydrogen isotope ratios (δD) of individual lipids in fossil plant tissues show great potential for reconstructing paleocontinental hydrological conditions. Leaf wax n-alkanes are some of the most abundant lipid molecules biosynthesized by terrestrial plants (Eglinton and Hamilton, 1967), containing C-bound hydrogen that is geologically stable (Schimmelmann et al., 1999, 2006). Plant n-alkane δD values have been shown to correlate with local meteoric water δD (Sternberg, 1988), further modified by isotope enrichment in leaf water via transpiration and soil water evaporation (Sachse et al., 2006). Given the control of relative humidity on these processes, fossil leaf wax n-alkanes are being explored as a paleoaridity proxy (e.g., Liu and Huang, 2005; Pagani et al., 2006; Smith and Freeman, 2006). Scientific understanding of the role of climate and plant physiology on compound-specific δD systematics is still evolving (Chikaraishi and Naraoka, 2003; Pedentchouk et al., 2008; Diefendorf et al., 2010), requiring further empirical study of plant-water-deuterium systematics before this proxy can be applied straightforwardly to ancient continental systems. Further development and refinement of emerging proxies such as the aforementioned mineral
stable isotope and biomarker approaches, along with improved spatial and temporal resolution of terrestrial proxy records, are fundamental to successful calibration and testing of climate models for a future warmer Earth using deep-time analogues.
INDICATORS OF OCEANIC PH AND REDOX
Recent advances in instrumentation (e.g., multicollector inductively coupled plasma mass spectrometry, nanoscale secondary ion mass spectrometry) coupled with development and modern calibration of geochemical and isotopic proxies that span the periodic table have greatly expanded the range of paleoceanographic proxies of oceanic redox, alkalinity, and pH. Studies over the past decade have documented the linear or exponential relationships between trace element ratios (Mg/Ca, Cd/ Ca, Zn/Ca, U/Ca) and stable (O, C, B) isotopic compositions of carbonate-bearing fossil fauna and changes in seawater carbonate content [CO32–] and pH (Lea et al., 1999; Marchitto et al., 2000; Russell et al., 2004). For example, the U/Ca ratios of certain planktonic foraminifera genera have been used to determine variations in seawater carbonate ion content over the last glacial cycle that track atmospheric pCO2 variations archived in polar ice cores (Russell et al., 1996, 2004).
The timing and geographic extent of past events of oceanic hypoxia and anoxia can be resolved through the integration of sulfur isotopes of reduced (pyrite) and oxidized minerals (carbonate-associated sulfate in carbonates) and fossil organic matter, and the abundance and stable isotopic composition of heavy metals (e.g., Fe, U, and Mo isotopes). The abundance of redox-sensitive transition elements (V, Mo, Fe, Cr) and their partitioning between various mineral phases in organic-rich deposits have provided much insight into the origin of O2-deficient waters in pre-Cenozoic marine basins—in particular in past greenhouse worlds (e.g., Sageman et al., 2003; Meyers et al., 2005; McArthur et al., 2008b; Lyons et al., 2009; Algeo et al., 2010). Carbonate-associated sulfate data coupled with pyrite sulfur isotope data from Precambrian oceanic deposits have refined scientific understanding of the oxygenation of Earth’s early atmosphere (Kah et al., 2004). Recently, the utility of molybdenum (δ97Mo), uranium (δ238U), and iron (δ56Fe) isotopes of various components in organic-rich black shales has been demonstrated as a sensitive proxy of oceanic O2 levels, revealing the protracted oxygenation of Earth’s early ocean in the Proterozoic (e.g., Anbar and Knoll, 2002; Arnold et al., 2004), and elucidating the global expansion of oceanic anoxia during past warm periods (e.g., Jenkyns et al., 2007; Gordon et al., 2009; Duan et al., 2010; Montoya-Pino et al., 2010).
Synergy of Observations and Models
A synergistic approach combining observations and modeling provides an optimal strategy for answering critical questions regarding how Earth’s climate system has responded to varying levels of greenhouse gases and other forcing factors. Model simulations provide a global picture of the state of the climate system and also a window into how various processes operate to maintain a given climate state. Disparities between simulated climate variables (e.g., surface temperatures, precipitation, ocean circulation) and proxy observations of these state variables pose questions regarding how much of the disparity is due to model biases or deficiencies and how much is due to observational bias. An example of data bias is related to simulated tropical and subtropical SSTs during warm Paleogene climates, which for years were high compared to proxy data. Recent recognition and correction of problems with the proxy data (see Box 4.4) have not brought models and data into greater agreement. An example of model bias is related to simulated high-latitude surface temperatures in warm climate regimes, which have been too low compared to proxy data. Here, continued improvement and development of new innovative observational techniques have strengthened the conclusion that models are challenged to simulate such high polar surface temperatures. This disparity has led to active model exploration of feedback processes that may operate in warm greenhouse climates but are not revealed by data-model studies of Earth’s more recent glacial state. Thus, it is to the benefit of both observational and modeling communities to work in close collaboration through real-time data-model comparison studies. Disparities between models and observations represent synergistic research opportunities.

























