
Using CRISPR to Combat Human Disease Vectors
OMAR S. AKBARI
University of California, San Diego
The annual incidence of vector-borne disease exceeds 1 billion globally—roughly half of the world’s population is at risk of infection.1 Mosquito-borne diseases account for the majority of cases (WHO 2014), but there are no vaccines for most of them, so prevention, mainly through inefficient vector control of limited effectiveness, is the primary method to reduce disease burden. Furthermore, treatments for most mosquito-borne pathogens are also limited, and those that are effective are under threat from increasing pathogen drug resistance.
The severity of the problem is best exemplified by the repeated development of antimalarial resistance in Southeast Asia. In the 1990s parasite resistance to first- and second-line malaria drugs necessitated the development of combination therapies for treatment (Nosten et al. 1987, 1994). However, high resistance to these combination drugs and their later derivatives resulted in an increase in malaria-related deaths in this region (Dondorp et al. 2009; Ménard et al. 2016; Phyo et al. 2016). Therefore, in most cases, vector control is the best approach for reducing the burden of vector-borne diseases.
VECTOR CONTROL TOOLS
Chemical insecticides have historically been an important tool for mosquito control, but they have limitations, most notably their limited efficacy due to increasing vector insecticide resistance and their limited species specificity and duration. While insecticidedriven approaches have been successful in some dis-
___________________
1 World Health Organization, “Vectorborne diseases,” October 31, 2017 (https://www.who.int/en/newsroom/factsheets/detail/vector-bornediseases).
ease prevention programs (Pluess et al. 2010), for a myriad of reasons they have mixed results overall (Esu et al. 2010; George et al. 2015; Maciel-de-Freitas et al. 2014). Even in areas where sustained vector control has been achieved in the past, insecticide resistance has greatly reduced or eliminated the impact of vector control on disease transmission (Hemingway et al. 2002; Liu 2015; Macielde-Freitas et al. 2014).
Given the widespread use of insecticides and limited number of insecticide families available for vector control programs, insecticide resistance will continue to be a barrier to insecticidebased vector control. New control techniques are therefore being evaluated to complement vector control programs.
STERILE INSECT TECHNIQUE FOR INSECT CONTROL
Sterile insect technique (SIT) is the gold standard for geneticsbased insect population control. In classic SIT, insects are treated with ionizing radiation to induce male sterility and then released in high frequency to mate with wild females, resulting in nonviable progeny. Over time, repeated mass releases of sterile males suppress and can even eliminate target populations. This approach was used to eradicate the screwworm fly (Cochliomyia hominivorax; Krafsur et al. 1986), the Mexican fruit fly (Anastrepha ludens), and the Mediterranean fruit fly (Ceratitis capitata) from regions of North America (Hendrichs et al. 2002).
But in mosquitoes irradiation-based SIT causes high male mortality and exceedingly high fitness costs. For example, field studies show that the release of irradiated, sterile male Aedes albopictus led to very limited population reduction (Bellini et al. 2013) likely for these reasons.
So although irradiation-based SIT presents an environmentally friendly method of local population suppression, it is not feasible or scalable in its current form for large-scale control of mosquito populations.
NOVEL VECTOR CONTROL METHODS
In recent years innovative genetic vector control methods, such as the release of insects carrying a dominant lethal (RIDL) (Thomas et al. 2000), have demonstrated large reductions in wild vector populations (Carvalho et al. 2015; Harris et al. 2012). Other novel disease or vector control methods, such as Dengue and Zika virus transmissionblocking Wolbachia-infected Aedes aegypti and the Wolbachia incompatible insect technique (IIT), respectively, are being evaluated in the field (Schmidt et al. 2017). While effective, these methods require large numbers of mosquitoes to be raised, manually sexed, and released as adults in the field near target sites.
Building mosquito mass rearing factories in local disease endemic areas is costly and labor intensive and current procedures are error prone (Gilles et al. 2014; Papathanos et al. 2009). Female release, even in small numbers, is particu-
larly problematic to the Wolbachia IIT technology as the release will immunize the target population to the incompatible Wolbachia strain and ultimately lead to the failure of the approach. Some studies even indicate that in some contexts, Wolbachia actually enhances pathogen infection (Dodson et al. 2014; Hughes et al. 2014) or can have large vector fitness costs, which can be problematic (Joshi et al. 2014).
Additionally, the antibiotic drugs required during rearing of RIDL mosquitoes have high male fitness costs (about 5 percent that of wild-type male fitness) based on RIDL field trials in the Cayman Islands (Harris et al. 2011) and Brazil (Carvalho et al. 2015), due to the loss or alteration of gut microbiome or symbiotic bacteria as well as toxicity to mitochondrial cell functions (Chatzispyrou et al. 2015; Moullan et al. 2015). Therefore, there is still an urgent need for new vector control technologies for the suppression of wild vector populations.
USING CRISPR
The advent of CRISPR2 technology has excited the potential to engineer new game-changing technologies and innovative systems that can be used to control wild populations of mosquitoes. Two developments of particular interest are a self-limiting system termed precision-guided sterile insect technique (pgSIT) (Kandul et al. 2019) and a homing-based gene drive (HGD) (Champer et al. 2016; Esvelt et al. 2014). The unique features of these systems can make them valuable in the future to control mosquitoes, as elaborated below.
pgSIT
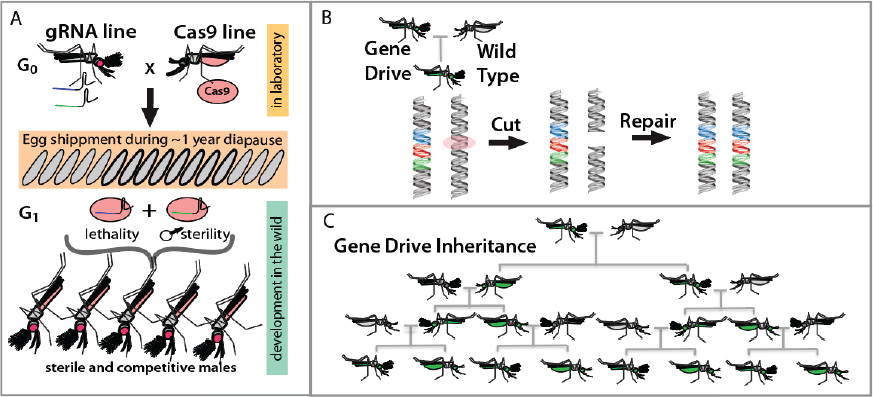
The novel CRISPR-based pgSIT mechanistically relies on a dominant genetic technology that enables simultaneous sexing and sterilization, facilitating the release of eggs into the environment and ensuring that only sterile adult males emerge. Importantly, for field applications, the release of eggs will eliminate burdens of manually sexing and sterilizing males, reducing the time and effort involved and increasing scalability. Moreover, the release of eggs should reduce the need to build factories near release sites as eggs could be shipped to release locations from a centralized facility and hatched directly in the environment.
This system was recently systematically engineered in an insect fly model system and was shown to be extremely efficient at generating 100 percent sterile males that could suppress populations. The system functions by mass producing two strains, one expressing the CRISPR-associated protein 9 (Cas9) endonuclease and the other expressing two guide RNAs (gRNAs), one targeting a gene important for female viability and the other a gene important for male fertility. When
___________________
2 CRISPR (clustered regularly interspaced short palindromic repeats) is a family of DNA sequences in the genomes of prokaryotic organisms such as bacteria.
the two separate strains are crossed the only surviving progeny are sterile males, which can be directly deployed (figure 1A).
Efforts are underway to transfer this technology to mosquitoes, and in the coming years it may be deployed in the field.
Homing-Based Gene Drives
Replacement of wild insect populations with genetically modified individuals unable to transmit disease provides an environmentally friendly, sustainable, and self-perpetuating method of disease prevention. However, transgenes that mediate disease resistance to treatment (refractoriness) may inadvertently compromise the fitness of insects that carry them. Furthermore, wild populations are large, partially reproductively isolated, and dispersed over wide areas.
Population replacement therefore requires a gene drive mechanism to spread linked genes that mediate disease refractoriness through wild populations at greater than Mendelian frequencies. In an effort to achieve this, CRISPR methods have been used to accelerate the development of HGDs in model systems in addition to mosquitoes and even mammals (Champer et al. 2017, 2018; DiCarlo et al. 2015; Gantz and Bier 2015; Gantz et al. 2015; Grunwald et al. 2019; Hammond et al. 2016, 2018; KaramiNejadRanjbar et al. 2018; Kyrou et al. 2018; Li et al. 2019; Windbichler et al. 2011; Yan and Finnigan 2018).
HGDs function by encoding the Cas9 endonuclease and an independently expressed gRNA responsible for mediating DNA base pairing directing Cas9-mediated cleavage at a predetermined site (Champer et al. 2016; Esvelt et al. 2014; Gantz and Bier 2016; Marshall and Akbari 2018). When the HGD is positioned in its target site in a heterozygote, doublestranded DNA breakage of the opposite chromosome can cause the drive allele to be used as a template (i.e., donor chromosome) for DNA repair mediated by homologous recombination. This can result in copying, or “homing,” of the HGD into the broken (receiver) chromosome, thereby converting heterozygotes to homozygotes in the germline, which can bias Mendelian inheritance ratios and lead to an increase in HGD frequency in a population (figure 1B,C).
Given recent progress toward developing HGDs in pest species such as mosquitoes (Gantz et al. 2015; Hammond et al. 2016, 2018; Kyrou et al. 2018; Li et al. 2019), there is significant enthusiasm for their potential use to control wild populations. For example, release of HGDs linked with effector genes that inhibit mosquito pathogen transmission (Buchman et al. 2019a,b; Isaacs et al. 2011; Jupatanakul et al. 2017) may lead to replacement of diseasesusceptible mosquitoes with diseaseresistant counterparts, thereby reducing pathogen transmission (i.e., population modification drive). Alternatively, HGDs targeting genes that affect the fitness of female mosquitoes could also lead to gradual population declines and potentially even elimination (i.e., population suppression drive) (Kyrou et al. 2018; Windbichler et al. 2008, 2011).
CONCLUSION
Both genetic SIT systems and modification and suppression drives have the potential to transform mosquito population control measures (Burt 2003; Champer et al. 2016; Esvelt et al. 2014), and therefore have excited discussions about their potential use, regulation, safety, ethics, and governance (Adelman et al. 2017; Akbari et al. 2015; NASEM 2016; Oye et al. 2014). Field testing of these systems over the next 5 to 10 years will help illuminate the efficacy and safety concerns of these systems.
REFERENCES
Adelman Z, Akbari O, Bauer J, Bier E, Bloss C, Carter SR, Callender C, Costero-Saint Denis A, Cowhey P, Dass B, and 23 others. 2017. Rules of the road for insect gene drive research and testing. Nature Biotechnology 35(8):716–18.
Akbari OS, Bellen HJ, Bier E, Bullock SL, Burt A, Church GM, Cook KR, Duchek P, Edwards OR, Esvelt KM, and 17 others. 2015. Safeguarding gene drive experiments in the laboratory. Science 349(6251):927–29.
Bellini R, Medici A, Puggioli A, Balestrino F, Carrieri M. 2013. Pilot field trials with Aedes albopictus irradiated sterile males in Italian urban areas. Journal of Medical Entomology 50(2):317–25.
Buchman A, Gamez S, Li M, Antoshechkin I, Lee S-H, Wang S-W, Chen C-H, Klein MJ, Duchemin JB, Crowe JE Jr, and 2 others. 2019a. Broad dengue neutralization in mosquitoes expressing an engineered antibody. SSRN Electronic Journal. https://doi.org/10.2139/ssrn.3398490.
Buchman A, Gamez S, Li M, Antoshechkin I, Li H-H, Wang H-W, Chen C-H, Klein, MJ, Duchemin J-B, Paradkar PN, Akbari OS. 2019b. Engineered resistance to Zika virus in transgenic Aedes aegypti expressing a polycistronic cluster of synthetic small RNAs. Proceedings, National Academy of Sciences 116(9):3656–61.
Burt A. 2003. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proceedings, Royal Society B: Biological Sciences 270(1518):921–28.
Carvalho DO, McKemey AR, Garziera L, Lacroix R, Donnelly CA, Alphey L, Malavasi A, Capurro ML. 2015. Suppression of a field population of Aedes aegypti in Brazil by sustained release of transgenic male mosquitoes. PLoS Neglected Tropical Diseases 9(7):e0003864.
Champer J, Buchman A, Akbari OS. 2016. Cheating evolution: Engineering gene drives to manipulate the fate of wild populations. Nature Reviews Genetics 17(3):146–59.
Champer J, Reeves R, Oh SY, Liu C, Liu J, Clark AG, Messer PW. 2017. Novel CRISPR/Cas9 gene drive constructs reveal insights into mechanisms of resistance allele formation and drive efficiency in genetically diverse populations. PLoS Genetics 13(7):e1006796.
Champer J, Liu J, Oh SY, Reeves R, Luthra A, Oakes N, Clark AG, Messer PW. 2018. Reducing resistance allele formation in CRISPR gene drive. Proceedings, National Academy of Sciences 115(21):5522–27.
Chatzispyrou IA, Held NM, Mouchiroud L, Auwerx J, Houtkooper RH. 2015. Tetracycline antibiotics impair mitochondrial function and its experimental use confounds research. Cancer Research 75(21):4446–49.
DiCarlo JE, Chavez A, Dietz SL, Esvelt KM, Church GM. 2015. Safeguarding CRISPR-Cas9 gene drives in yeast. Nature Biotechnology 33(12):1250–55.
Dodson BL, Hughes GL, Paul O, Matacchiero AC, Kramer LD, Rasgon JL. 2014. Wolbachia enhances West Nile Virus (WNV) infection in the mosquito Culex tarsalis. PLoS Neglected Tropical Diseases 8(7):e2965.
Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, Lwin KM, Ariey F, Hanpithakpong W, Lee SJ, and 12 others. 2009. Artemisinin resistance in Plasmodium falciparum malaria. New England Journal of Medicine 361(5):455–67.
Esu E, Lenhart A, Smith L, Horstick O. 2010. Effectiveness of peridomestic space spraying with insecticide on dengue transmission: Systematic review. Tropical Medicine & International Health 15(5):619–31.
Esvelt KM, Smidler AL, Catteruccia F, Church GM. 2014. Emerging technology: Concerning RNA-guided gene drives for the alteration of wild populations. eLife 3:e03401.
Gantz VM, Bier E. 2015. The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science 348(6233):442–44.
Gantz VM, Bier E. 2016. The dawn of active genetics. BioEssays 38(1):50–63.
Gantz VM, Jasinskiene N, Tatarenkova O, Fazekas A, Macias VM, Bier E, James AA. 2015. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proceedings, National Academy of Sciences 112(49):E6736–43.
George L, Lenhart A, Toledo J, Lazaro A, Han WW, Velayudhan R, Ranzinger SR, Horstick O. 2015. Community-effectiveness of temephos for dengue vector control: A systematic literature review. PLoS Neglected Tropical Diseases 9(9):e0004006.
Gilles JRL, Schetelig MF, Scolari F, Marec F, Capurro ML, Franz G, Bourtzis K. 2014. Towards mosquito sterile insect technique programmes: Exploring genetic, molecular, mechanical and behavioural methods of sex separation in mosquitoes. Acta Tropica 132 (Suppl):S178–87.
Grunwald HA, Gantz VM, Poplawski G, Xu X-RS, Bier E, Cooper KL. 2019. Super-Mendelian inheritance mediated by CRISPR-Cas9 in the female mouse germline. Nature 566(7742):105–109.
Hammond A, Galizi R, Kyrou K, Simoni A, Siniscalchi C, Katsanos D, Gribble M, Baker D, Marois E, Russell S, and 4 others. 2016. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nature Biotechnology 34(1):78–83.
Hammond AM, Kyrou K, Gribble M, Karlsson X, Morianou I, Galizi R, Beaghton A, Crisanti A, Nolan T. 2018. Improved CRISPR-based suppression gene drives mitigate resistance and impose a large reproductive load on laboratorycontained mosquito populations. bioRxiv. https://doi.org/10.1101/360339.
Harris AF, Nimmo D, McKemey AR, Kelly N, Scaife S, Donnelly CA, Beech C, Petrie WD, Alphey L. 2011. Field performance of engineered male mosquitoes. Nature Biotechnology 29(11):1034–37.
Harris AF, McKemey AR, Nimmo D, Curtis Z, Black I, Morgan SA, Oviedo MN, Lacroix R, Naish N, Morrison NI, and 13 others. 2012. Successful suppression of a field mosquito population by sustained release of engineered male mosquitoes. Nature Biotechnology 30:828–30.
Hemingway J, Field L, Vontas J. 2002. An overview of insecticide resistance. Science 298(5591):96–97.
Hendrichs J, Robinson AS, Cayol JP, Enkerlin W. 2002. Medfly areawide sterile insect technique programmes for prevention, suppression or eradication: The importance of mating behavior studies. Florida Entomologist 85(1):1–13.
Hughes GL, Rivero A, Rasgon JL. 2014. Wolbachia can enhance plasmodium infection in mosquitoes: Implications for malaria control? PLoS Pathogens 10(9):e1004182.
Isaacs AT, Li F, Jasinskiene N, Chen X, Nirmala X, Marinotti O, Vinetz JM, James AA. 2011. Engineered resistance to plasmodium falciparum development in transgenic Anopheles stephensi. PLoS Pathogens 7(4):e1002017.
Joshi D, McFadden MJ, Bevins D, Zhang F, Xi Z. 2014. Wolbachia strain wAlbB confers both fitness costs and benefit on Anopheles stephensi. Parasites and Vectors 7:336.
Jupatanakul N, Sim S, Angleró-Rodríguez YI, Souza-Neto J, Das S, Poti KE, Rossi SL, Bergren N, Vasilakis N, Dimopoulos G. 2017. Engineered Aedes aegypti JAK/STAT pathway-mediated immunity to dengue virus. PLoS Neglected Tropical Diseases 11(1):e0005187.
Kandul NP, Liu J, Sánchez Castellanos HM, Wu SL, Marshall JM, Akbari OS. 2019. Transforming insect population control with precision guided sterile males with demonstration in flies. Nature Communications 10(1):84.
KaramiNejadRanjbar M, Eckermann KN, Ahmed HMM, Sánchez Castellanos HM, Dippel S, Marshall JM, Wimmer EA. 2018. Consequences of resistance evolution in a Cas9based sex conversion-suppression gene drive for insect pest management. Proceedings, National Academy of Sciences 115(24):6189–94.
Krafsur ES, Townson H, Davidson G, Curtis CF. 1986. Screwworm eradication is what it seems. Nature 323(6088):495–96.
Kyrou K, Hammond AM, Galizi R, Kranjc N, Burt A, Beaghton AK, Nolan T, Crisanti A. 2018. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nature Biotechnology 36:1062–66.
Li M, Yang T, Kandul NP, Bui M, Gamez S, Raban R, Bennett J, Sanchez C HM, Lanzaro GC, Schmidt H, and 3 others. 2019. Development of a confinable gene-drive system in the human disease vector, Aedes aegypti. bioRxiv. https://doi.org/10.1101/645440.
Liu N. 2015. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annual Review of Entomology 60(1):537–59.
Maciel-de-Freitas R, Avendanho FC, Santos R, Sylvestre G, Araújo SC, Lima JBP, Martins AJ, Coelho GE, Valle D. 2014. Undesirable consequences of insecticide resistance following Aedes aegypti control activities due to a dengue outbreak. PloS One 9(3):e92424.
Marshall JM, Akbari OS. 2018. Can CRISPR-based gene drive be confined in the wild? A question for molecular and population biology. ACS Chemical Biology 13(2):424–30.
Ménard D, Khim N, Beghain J, Adegnika AA, Shafiul-Alam M, Amodu O, Rahim-Awab G, Barnadas C, Berry A, Boum Y, and 80 others. 2016. A worldwide map of Plasmodium falciparum K13-propeller polymorphisms. New England Journal of Medicine 374(25):2453–64.
Moullan N, Mouchiroud L, Wang X, Ryu D, Williams EG, Mottis A, Jovaisaite V, Frochaux MV, Quiros PM, Deplancke B, and 2 others. 2015. Tetracyclines disturb mitochondrial function across eukaryotic models: A call for caution in biomedical research. Cell Reports 10(10):1681–91.
NASEM [National Academies of Sciences, Engineering, and Medicine]. 2016. Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values. Washington: National Academies Press.
Nosten F, Imvithaya S, Vincenti M, Delmas G, Lebihan G, Hausler B, White N. 1987. Malaria on the Thai-Burmese border: Treatment of 5192 patients with mefloquine-sulfadoxine-pyrimethamine. Bulletin of the World Health Organization 65(6):891–96.
Nosten F, Luxemburger C, ter Kuile FO, Woodrow C, Eh JP, Chongsuphajaisiddhi T, White NJ. 1994. Treatment of multidrug-resistant Plasmodium falciparum malaria with 3-day artesunate-mefloquine combination. Journal of Infectious Diseases 170(4):971–77.
Oye KA, Esvelt K, Appleton E, Catteruccia F, Church G, Kuiken T, Lightfoot SBY, McNamara J, Smidler A, Collins JP. 2014. Regulating gene drives. Science 345(6197):626–28.
Papathanos PA, Bossin HC, Benedict MQ, Catteruccia F, Malcolm CA, Alphey L, Crisanti A. 2009. Sex separation strategies: Past experience and new approaches. Malaria Journal 8(Suppl 2):S5.
Phyo AP, Ashley EA, Anderson, TJC, Bozdech, Z, Carrara VI, Sriprawat K, Nair S, White MM, Dziekan J, Ling C, and 10 others. 2016. Declining efficacy of artemisinin combination therapy against P. falciparum malaria on the Thai-Myanmar border (20032013): The role of parasite genetic factors. Clinical Infectious Diseases 63(6):784–91.
Pluess B, Tanser FC, Lengeler C, Sharp BL. 2010. Indoor residual spraying for preventing malaria. Cochrane Database of Systematic Reviews 4:CD006657.
Schmidt TL, Barton NH, Rašić G, Turley AP, Montgomery BL, Iturbe-Ormaetxe I, Cook PE, Ryan PA, Ritchie SA, Hoffmann AA, and 2 others. 2017. Local introduction and heterogeneous spatial spread of dengue-suppressing Wolbachia through an urban population of Aedes aegypti. PLoS Biology 15(5):e2001894.
Thomas DD, Donnelly CA, Wood RJ, Alphey LS. 2000. Insect population control using a dominant, repressible, lethal genetic system. Science 287(5462):2474–76.
WHO [World Health Organization]. 2014. A Global Brief on Vector-Borne Diseases. Geneva.
Windbichler N, Papathanos PA, Crisanti A. 2008. Targeting the X chromosome during spermatogenesis induces Y chromosome transmission ratio distortion and early dominant embryo lethality in Anopheles gambiae. PLoS Genetics 4(12):e1000291.
Windbichler N, Menichelli M, Papathanos PA, Thyme SB, Li H, Ulge UY, Hovde BT, Baker D, Monnat RJ, Burt A, and 1 other. 2011. A synthetic homing endonuclease-based gene drive system in the human malaria mosquito. Nature 473(7346):212–15.
Yan Y, Finnigan GC. 2018. Development of a multi-locus CRISPR gene drive system in budding yeast. Scientific Reports 8(1):17277.
This page intentionally left blank.










