
11
Radiation Hazards
Introduction
The long-range plans of NASA include construction and occupation of a space station in low Earth orbit, and possible human exploratory missions to the Moon and Mars within the next 25 years. The biological effects of exposure to radiation in space pose potentially serious health risks that have to be controlled or mitigated before such relatively long-term missions beyond low Earth orbit can be initiated. 1 The levels of radiation in space are high enough and the missions are long enough that adequate shielding is necessary to minimize carcinogenic, cataractogenic, and possible neurologic effects on crew members. The question to be answered is, What will provide the necessary protection, for the extent of a mission, against the biological effects of high-energy galactic cosmic-ray particles ranging from energetic protons with low mean linear energy transfer (LET) to nuclei of high atomic number and very high energies (HZE) with high LET and against the effects of transient radiation in solar particle events?*
This chapter summarizes a recent National Research Council (NRC) report, Radiation Hazards to Crews of Interplanetary Missions: Biological Issues and Research Strategies,2 on current knowledge of the types and levels of radiation to which crews will be exposed in space, and also discusses the range of possible human health effects that must be protected against. It suggests steps to be taken and the types of experiments needed to reduce significantly the level of uncertainty regarding health risks to human crews in space, and it recommends priorities for research from which NASA can obtain the information required to evaluate the biological risks faced by humans exposed to radiation in space and to mitigate such risks. The chapter outlines, in general terms, the commitment of resources that NASA
should make to carrying out these experiments in order to accomplish an effective shielding design in time for a possible mission launch to Mars.
It is recommended that, if the goal of safe interplanetary missions with human crews is sought, NASA explore various possibilities, including the construction of new facilities, to increase the research time available for experiments with HZE particles.
A recent comprehensive report to NASA3 focuses on the extent to which animal experiments are required to assess the health risks to humans exposed to the radiation in space. It examines carefully the molecular, cellular, and animal background information that will be needed to extrapolate these radiation effects to humans and concludes that animal experimentation should be pursued selectively, but decisively. This conclusion is strong support for the priority research recommendations and strategies given in this chapter.
The characteristics of the radiation field are more complicated in low Earth orbit (LEO) than in interplanetary space because the number of particles depends on altitude and on the magnitude and direction of Earth's magnetic field.4 Since the orientation of the space station will be fixed and its orbit will be at an inclination of 51.6°, the radiation dose rate depends on orbit position and also on location within the station. The latter asymmetry is about 2-fold under identical shielding. The three principal radiation sources of concern are galactic cosmic rays, protons trapped by Earth's field, and solar particle events (SPEs), whose frequencies are a function of solar activity, being highest during solar maxima. "Currently there are not techniques available for forecasting either the occurrence or the magnitude of such events."5 The peak of the next solar cycle is expected in spring of the year 2000. During an SPE the proton flux may increase 10-fold in less than 10 hours. A radiobiological problem in LEO is that the mix of high- and low-LET particles changes with altitude (more protons at higher altitudes) and with solar cycle (more high-LET particles during solar minimum).6 Clearly, real-time monitoring of dose rates and the spectra of particles contributing to the dose are important because the does equivalent, in sieverts (Sv), depends on the relative biological effectiveness (RBE), which varies with LET and, for end points of interest, is currently not well known for HZE particles. Thus, the research strategies outlined below for space missions are also necessary for understanding the radiation effects in LEO.
Determination of the frequencies of chromosomal aberrations in lymphocytes irradiated in vitro versus gamma ray dose has been used to estimate the dose equivalent for chromosomal aberrations in the lymphocytes of two astronauts on the Mir-18 115-day spaceflight.7 The differences in postflight minus preflight frequencies are calibrated to give the dose equivalent from the results of acute exposures of preflight lymphocyte samples to gamma rays. The average dose equivalent was 0.15 Sv. The technique looks promising for retrospective dosimetry, but additional data are required if the method is to be applied to chronic and nonuniform body exposures.8 If the RBEs for cancer induction, for example, differ from those for chromosomal aberrations, aberrations would not be a useful biodosimeter for cancer without knowledge of appropriate correction factors.
Statement Of The Problem
The knowledge needed to design adequate shielding has both physical and biological components: (1) the distribution and energies of radiation particles present behind a given shielding material as a result of the shield being struck by a given type and level of incident radiation and (2) the effects of a given dose on relevant biological systems for different radiation types. Each component involves significant uncertainty that must be reduced to permit effective design of shielding, given that the level of uncertainty governs the amount of shielding. It is only prudent to design shielding that will protect space crew members from the predicted, but uncertain, high levels of biological effects resulting from
their exposure to radiation. At the same time, excess shielding, based on current cost estimates, would impose an excess expenditure on the order of tens of billions of dollars.9
HZE particles impinging on shielding, or on human tissue, result in very dense ionization tracks (high LET) with numerous fragments that produce a spectrum of other energetic nuclei, protons, neutrons, and heavy fragments. The numbers of these other nuclei depend on the nature of the shielding and its mass per unit area. The energy loss of the individual particles depends on their types and energies. Thus, each particle contributes to the radiation dose and biological response, which are dependent on the number of particles, their types, and their energies. The theoretical calculations of doses per particle type obtained thus far for relevant shielding materials must be verified by ground-based experiments, because the radiation field rate in space is too complex for sufficient experimental analysis. Ionizing radiation either affects cellular macromolecules directly or reacts with water to produce free radicals that affect these macromolecules by so-called indirect effects. These effects are mitigated somewhat by the presence of free-radical scavengers in the surrounding medium. The scavengers are useful in significantly reducing the effects of low-LET radiation but, with some exceptions,10 do not seem to result in any such decreases in the damage caused by high-LET radiation.
The biological effects of fast, charged particles depend on the nature of the particle (its charge and velocity) and on the specific biological end point under observation (e.g., cell killing, mutation at a specific genetic locus, chromosomal alterations, cell transformation in vitro, and tumor induction). The RBE is taken as the ratio of the dose of gamma rays required to produce a specific effect to the dose of particle radiation required to produce the same level of effect. The RBE depends on the type of particle and the biological effect under consideration and may vary with the magnitude of the biological effect. More importantly, RBE varies greatly with the LET of the particle. For example, high-energy protons may have an RBE value approaching 1.0, whereas high-energy iron nuclei may have an RBE approaching 40. For tumor induction in animals exposed at lower doses, the relationship between RBE and LET is known for only one tumor site—the Harderian gland in mice. There are no equivalent data for tumor induction in humans. Hence, there is a great uncertainty in extrapolating from cell and scanty mouse data to evaluate human risk. Moreover, there is also great uncertainty in the extrapolation from the analysis of cancer induction in Japanese individuals exposed to acute doses of radiation resulting from the atomic bombs.11 These doses are not known precisely. Since this radiation was primarily low LET, in order to estimate risks to humans in spaceflight conditions one must extrapolate from the RBE-versus-LET data for cells in culture and from small mammals to humans. In addition, one must extrapolate from the risks due to acute exposures of humans to the low-dose-rate chronic exposures involved in space missions (except for the relatively acute exposure from solar particle events). As a general rule, as the dose rate decreases, the biological effect from a given dose also decreases. This dose reduction, in going from acute to chronic exposure, also depends on the biological system and may range from a factor of 2 to 10.12 The dose rate reduction factor for HZE particles is not well known but is probably closer to 1.13 Two other factors that must be considered, but whose impacts are currently unknown, are the effects of biochemical or cellular repair reactions following exposure to HZE particles and the effects of microgravity on such reactions. Thus, in estimating the risks to humans exposed to radiation in space, the uncertain factors are the radiation fields behind the shielding and the extrapolation, via cell culture and animal experiments, from the uncertain risks posed by acute low-LET exposure to risks posed by chronic high-LET exposure.
To quote Curtis et al.,14 "Uncertainties in these numbers are diffcult to estimate but a rough analysis leads to a 10-15% uncertainty in the initial charged particle spectra, a 50% uncertainty in the radiation transport calculation, a factor of 2-3 uncertainty in the risk coefficients for low-LET radiation (most of which is due to uncertainty in the dose and dose rate effectiveness factor) and perhaps a factor
of 2-5 uncertainty in the risk cross sections at high-LET. Thus, and overall uncertainty in the risk of radiation-induced cancer of a factor of 4-15 for a space crew in the galactic cosmic ray environment appears to exist at the present state of our knowledge." Obviously, these uncertainties have, themselves, large uncertainties.
In 1989, the National Council on Radiation Protection and Measurements (NCRP) issued its report Guidance on Radiation Received in Space Activities, which had been requested by NASA.15 That report introduced different career limits depending on gender and age of onset of exposure to radiation in space. The career equivalent dose limits of 1 to 4 Sv that were recommended were based on a lifetime excess risk of cancer mortality of 3 × 10-2 per gray (Gy) of low-LET, acute radiation. The report's scope was limited to low-Earth-orbit missions, although it considered the radiation environment in deep space and the biological effects of HZE particles. Recommendations for protection against deterministic effects were also made in 1989. The career limit of 12 Sv recommended in 1979 by the National Research Council for skin was reduced by the NCRP in 1989 to 6 Sv; the limit for the lens of the eye was reduced from 6 to 4 Sv; and the limit for blood-forming organs was set at 1 to 4 Sv, depending on age and gender. Since 1989, estimates of cancer risk based on studies of atomic bomb survivors have been increased significantly, and the NCRP will issue new recommendations in the near future.
Current Understanding Of Biological Effects Of Radiation
Types of Effects
For the purpose of settling radiation protection standards, it has been useful to divide the biological effects that are important for human health into stochastic and deterministic effects.
Stochastic effects are considered to be those due to radiation-induced changes randomly distributed in the DNA of single cells that may lead to cancer or genetically transmissible effects, depending on the target cells. Cancer occurs after a long latency period: after 2 or more years in the case of leukemia and from 2 years to decades in the case of solid cancers. It is assumed that the frequency of such effects increases with dose without a threshold and that the severity of the effect is independent of dose. Stochastic effects are the most important consideration in setting protection limits for human populations exposed to radiation at low doses. It is important to note that radiation weighting factors or quality factors apply only to stochastic effects in the dose range pertinent to radiation protection. Based on studies of atomic bomb survivors of Hiroshima who were exposed to acute levels mainly of gamma rays but also of fission neutrons at very high dose rates, estimates for the risk of contracting leukemia have been refined,16 and there are also data on mortality and the incidence of solid cancers at more than 20 sites in the human body.17 18 (The precise contribution of the neutrons from the fission reactions to the total dose at Hiroshima is poorly known but is not considered a major contributor to the risk of cancer in those exposed at Hiroshima.) In 1991, the International Commission on Radiological Protection (ICRP)19 included leukemia and eight specific sites of solid cancers in its estimates of the probability of an individual contracting a fatal cancer after whole-body exposure to low-LET radiation at 1 Gy and at a high dose rate. The estimated probabilities were 7.12 × 10-2 per person based on a multiplicative projection model and 4.16 × 10-2 per person using an additive model. 20 A similar number, 3.7 × 10-2 per adult worker, using an additive model, was given in 1997 by the NCRP.21
Deterministic effects, previously termed nonstochastic effects, occur only after exposure to relatively high doses and affect cell populations to the detriment of specific organs or whole organisms. These effects can range from acute radiation sickness to hair loss or nausea. In contrast to stochastic effects, deterministic effects are dose dependent in both frequency and severity. Deterministic effects
may occur early (in a matter of hours or days) or late (after many months or even longer). Radiation protection standards are set to prevent deterministic effects, whereas standards to protect against stochastic effects are selected to limit these effects to an "acceptable" level.
Effects Induced by Protons and Heavy Ions
In deep space, the radiation environment consists mainly of galactic cosmic radiation (GCR) at a low fluence rate. In the energy range from 100 MeV (100 × 106 electron volts [eV]) per nucleon to 10 GeV (10 × 109 eV) per nucleon, the GCR consists of 87 percent protons, 12 percent helium ions, and 1 percent heavier ions.22 Protons are also the major component of solar particle events (SPEs), with a smaller contribution by helium and heavier ions emitted from the Sun. A major difference between SPE radiation and GCR is the much greater transient fluence in the former, which in very large solar particle events can be 1010 protons/cm2 with energies greater than 10 MeV.23
No data are available for most of the deterministic effects induced in humans by exposure to protons, and very limited data are available from studies of animals. The latter24 indicate that the biological effects of the higher-energy protons (> 138 MeV) are similar to those caused by 2-MeV x rays and 60Co gamma rays. The RBE for acute mortality was about 1.0 to 1.1. In the case of 160-MeV protons, Urano et al. found RBE values ranging between 0.8 and 1.3 for killing of jejunal crypt cells, skin damage, and effects on the lens of the eye in exposed mice.25 An exception to these RBE values of about 1 was the indirect finding by Storer et al. of higher RBE values, namely 2.4 and 4.9 for 30-day lethality and testicular atrophy, respectively, in mice. 26 In the treatment of human cancer by proton irradiation, an RBE of 1.1 has been used for planning purposes, and this value does not appear to underestimate the effectiveness of the protons.
In the case of stochastic effects, there are no data for cancer induction in humans exposed to protons. In a study of the induction of tumors in mice exposed to 60-MeV protons, Clapp et al. found no RBE values greater than about 1.0.27 burns et al. reported an RBE of about 3 for the induction of skin tumors in rats exposed to 10-MeV protons compared to electrons.28 The studies discussed above were carried out with single high-dose-rate exposures that are very different from the exposures occurring in space. Burns et al. noted a reduction in tumorogenic effect with the increasing fractionation of 10-MeV proton irradiation, an indication of recovery,29 and the curvilinear response to single doses was similar to the response to low-LET radiation, indicating that the protons have attributes both of low-LET radiation and, because the RBE is about 3, of high-LET radiation. Obviously, more animal data, using protons of several energy ranges, are needed to estimate human cancer risks from galactic protons.
The deterministic effects of exposure to heavy ions have been studied in experimental animals. The RBE of various ions was determined for effects that result from cell killing in the gut, testes, and bone marrow, and in vitro systems.30 As Figure 11.1 indicates, the RBE increases with increasing LET, reaches a maximum between 100 and 200 keV/µm, and decreases rapidly at higher LET values. The maximum RBE values for effects involving cell killing, such as shown in Figure 11.1, are in the range of 2.0 to 3.0.31 However, for cataract induction in rats and mice, the RBE values obtained by Merriam are in the range of 40 to 50 at low doses.32 Although the work of Lett and co-workers and of Worgul and co-workers suggests that it may be possible, with further data, to extrapolate across species to obtain RBE values for cataract induction, current data do not allow reliable estimation of the risk of cataract induction occurring in humans as a result of exposure to radiation in deep space.33 34 Observations on radiotherapy patients indicate that very high doses of low-LET radiation give rise to deterministic-type damage. HZE particles produce high-dose ionization tracks and kill the cells they traverse. The concern about such microlesions in the central nervous system resulting from traversal of cells by heavy charged
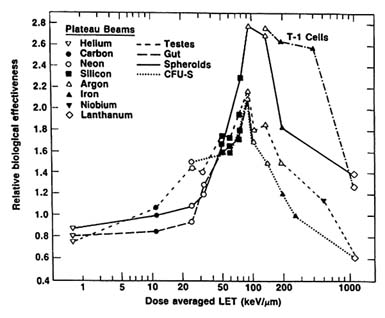
FIGURE 11.1 RBE-LET relationship for inactivation of different cells in vitro (CFU-S, intestinal crypt clonogenic cells, cells in spheroids, human T-1 cells) and loss of testis weight.
SOURCE: National Council on Radiation Protection and Measurements (NCRP). 1989. Guidance on Radiation Received in Space Activities. Recommendations of the National Council on Radiation Protection and Measurements. NCRP Report No. 98. National Council on Radiation Protection and Measurements, Bethesda, Md.
particles, such as iron, has not been eliminated, nor has any evidence been produced to show that the concern is justified.
The main concern about stochastic effects is the risk of cancer induction. It is agreed that the RBE for carcinogenesis increases with the increasing LET of the radiation. The evidence comes largely from animal experiments with fission neutrons but also from data on induction of lung tumors in humans exposed to alpha particles from radon.35 There has been only one systematic study of the relationship between the LET of heavy ions and the RBE values of the ions for tumor induction, which was carried out on the Harderian gland of mice.36 Although this gland is a suitable epithelial system, it is the only tumor model that has been examined over the range of LET values encountered in space. Hence, it is not possible to generalize on the basis of these data about RBE values for induction of cancer in important sites of the human body such as the breast, lung, and bone marrow. The data on Harderian gland tumors show a rise in RBE with increasing LET, reaching a maximum and a plateau of about 30 at approximately 100 to 200 keV/µm. 37 However, unlike the case of the RBE-LET relationship for cell killing and mutation (as in Figure 11.1),38 there is no evidence of a rapid decrease in RBE at higher LET values. Data on induction of skin tumors by argon ions39 support the expectation of a high RBE for induction of tumors by heavy ions but do not allow any more precise determination of what value should be used to estimate the risk of cancer induction in humans. Because of the importance of establishing the precise RBE values for cancer induction, more experimental studies are required.
First, a pragmatic set of studies is needed to provide data necessary for the determination of appropriate factors that should be used in making risk calculations. These should be systematic studies of RBE as a function of particle type and energy for a select number of heavy ions and for protons, using
well-defined animal models for tumorigenesis. In addition, information on dose rate and fractionation effects for protons is also needed. Improvements in risk estimates beyond those attainable with these data require a more complete understanding of the mechanisms of tumor induction and of principles that will aid in using data from experimental systems subjected to relatively high radiation doses, to estimate effects on humans exposed to low, protracted doses, and to estimate risks across populations. These kinds of studies will require the development and exploration of new model systems and the application of developing technologies in cell and molecular biology.
Priority Research Recommendations And Strategies
An estimate of the risk of adverse biological effects due to irradiation during a space mission corresponds to measurements of the relevant deterministic and stochastic biological consequences of exposure to radiation as a function of (1) dose, (2) dose rate, and (3) radiation quality. Such measurements should also be made as a function of radiation type and the shielding thickness for each type of spacecraft material. The risk estimates should include their uncertainties.
Given in the two subsections that follow, and summarized from the 1996 NRC report Radiation Hazards to Crews of Interplanetary Missions ,40 are the priority research recommendations and the issues that must be addressed in any endeavor to significantly reduce the risk and uncertainty of radiation hazards to the crews of interplanetary missions.* The research strategy presented to address each recommendation is narrowly focused on the recommendation and describes the minimum research model likely to provide the necessary data. The development of such a narrow set of strategies should not necessarily be taken as a recommendation to limit the scope of studies to those outlined below. If funds become available, many of these studies could be usefully expanded to provide additional relevant information.
In accordance with current understanding of the risks and uncertainties, the research recommendations are separated into higher- and lower-priority groups. As more data become available, some questions may shift in priority. Some strategies can be carried out independently, whereas others will be influenced by the outcome of research on the other recommended strategies and should be scheduled accordingly. The reasoning that forms the basis of these recommendations is discussed in greater detail in the 1996 report.41
Higher-Priority Research Recommendations
The higher-priority research recommendations, and strategies to answer them, follow. Research recommendations deemed important but of a lower priority are given in the next subsection.
|
1. |
Determine the carcinogenic risks following irradiation by protons and HZE particles. |
Strategy
This key recommendation requires that two related questions be addressed: (1) Can the risk due to irradiation by protons in the energy range of the space environment be predicted on the basis of the risk posed by exposure to low-LET radiation, such as gamma rays, and is there evidence for the repair of
damage in cells following fractionated exposure to protons and HZE particles? (2) What are the appropriate RBEs for HZE particles? The answers to these questions are fundamental to understanding the risk of contracting cancer as a result of travel in deep space. These important questions can be addressed using solely ground-based studies if appropriate funding and additional radiation resources such as accelerator time are made available.
Initial studies should focus on cellular effects that are relevant to cancer. Research with cells would provide a more rapid resolution than would tumor induction studies with animals of whether effects of exposure to high-energy protons are similar to those arising from low-LET radiation. Theoretical models of radiation effects, as well as currently available data for cellular and tumorigenic effects of exposure to protons (mostly at energies lower than those encountered in space), would argue that risks due to proton irradiation are similar to those from low-LET irradiation. Although considerable data are already available for protons in this energy range, these data are not satisfactory to answer questions related to high-energy protons in the 1-GeV energy range. To verify such a prediction for higher-energy protons, a series of studies should be conducted in several cellular systems, including human fibroblasts and lymphocytes, to examine the effects of protons in the 1-GeV energy range on cell killing, induction of chromosomal aberrations, and induction of gene mutations and transformation. Chromosomal aberrations could also be studied in lymphocytes from animals irradiated in vivo. By conducting such studies with both acute and fractionated exposure regimens, it would be possible to determine whether fractionation effects (sparing of radiation response by allowing for DNA repair between fractions) similar to those for low-LET radiation exist. Animal carcinogenesis experiments with protons should be conducted only if the results of cellular studies indicate discrepancies from the predictions. If tumorigenesis studies are warranted, the same animal models recommended for the study of tumorigenesis following exposure to HZE particles (described below) should be employed.
To obtain more reliable risk coefficients for HZE particles, a systematic series of studies of RBE-LET relationships for a select number of heavy ions—with emphasis on iron particles—should be conducted using well-defined animal models for tumorigenesis. Adequately defining these relationships requires that the dose-response relationship be determined for these particles in the dose range lower than 20 to 30 cGy, because at higher doses of high-LET radiation, the response appears to reach a maximum followed by a decrease. The model systems chosen should be those for which substantial dose-response data for other high- and low-LET radiation are already available. Model systems that are particularly amenable to concomitant cellular and molecular studies should receive priority. Given the fact that mice have been used more extensively than other mammalian species in studies of carcinogenesis,42 the use of the murine models for radiation-induced myelogenous leukemia and mammary gland cancer is appropriate. The conduct of these studies would require considerable commitment of time (beam time) at an HZE accelerator in an appropriate facility. Under the assumption that 3 months of beam time would be available per year, it is estimated that these studies would take approximately 6 years to complete. This estimate assumes that the irradiation of sufficient numbers of animals would require 2 years. Following irradiation, the completion of animal studies can be expected to require approximately 4 years. Under current conditions, which provide only 2 weeks of beam time each year, it would be almost impossible to complete a meaningful series of animal studies, because the period of time between the first set of animal irradiations and the last would probably be on the order of 6 years (assuming that half of the beam time were devoted to animal irradiation). This long temporal separation of experimental groups makes comparisons more difficult even with well-defined systems. Under these conditions, completion of the carcinogenesis studies would require a minimum of 10 years after the first irradiation.
Improvements in risk estimates beyond those attained with such data would require a more complete
understanding of mechanisms and of principles that will aid in the direct extrapolation of results from experimental systems to astronauts. These kinds of studies will likely require the development and exploration of new model systems and the application of developing technologies in cell and molecular biology.
|
2. |
Determine how cell killing and induction of chromosomal aberrations vary as a function of the thickness and composition of shielding. |
Strategy
The data obtained from studies conducted in response to recommendation 1 are necessary background for determining the biological effects of the specific radiation qualities and fluences in a spacecraft. The quality and dosimetry of radiation produced by HZE particles traversing shielding of different thicknesses and composition would be assessed from studies that address recommendation 6. Cellular studies should not be initiated for any particular energy (of HZE particle) and shielding until physical characterization of that radiation is completed. At a minimum, studies using cell-killing chromosomal aberrations and mutations and/or transformation as end points should be conducted using radiation qualities defined in dosimetry studies. Only the effects from acute exposures should be measured. Ground-based HZE particle sources, used with appropriate shielding to simulate in-flight conditions, should be suitable for such experiments. In-flight studies would be prohibitively difficult to conduct, and little gain in information would be realized.
It would be appropriate to conduct the initial studies in vitro using the same cell lines employed to address recommendation 1 (i.e., both rodent and human cell lines). Subsequent cytogenetic studies would have to be conducted in vivo to develop a more appropriate database for use in risk assessment calculations. It is recommended that bone marrow cells and peripheral lymphocytes, which are easily analyzed cytogenetically, be examined for chromosomal aberrations. Based on the information obtained in these cytogenetic studies, it would then be feasible to design a study to assess the induction of leukemia and breast cancer in mice exposed, behind shielding, to acute doses of HZE-particle radiation incident on the shielding (if cellular studies indicate this is necessary). Irradiation for in vitro studies could be accomplished in a relatively short time (i.e., about 2 days for each radiation type and energy). Typically, about 6 months would be required for analysis. If six radiation types were examined in consecutive order, as might be expected for a single research team, then such a study would require on the order of 3 years. Similarly, in vivo cytogenetic studies also require about 2 days of irradiation time for each radiation quality. If the in vivo and in vitro studies were performed in parallel, then both might reasonably be completed during the 3-year period.
The in vitro studies would allow the comparison of animal and human sensitivities to changes in shielding parameters. The in vivo experiments would provide data for in vitro and in vivo comparisons of cytogenic responses for the mouse. This parallelogram approach would provide estimates of the chromosomal aberration frequencies induced in humans in vivo (for bone marrow cells and lymphocytes). The mouse cancer studies (leukemia and breast cancer) can then be extrapolated in terms of human tumors by assuming that chromosomal aberration sensitivity factors apply. This approach would seem to be particularly reasonable for leukemias since chromosomal alterations are involved in the etiology of this cancer.
|
3. |
Determine whether there are studies that can be conducted to increase the confidence of extrapolation |
- from rodents to humans of radiation-induced genetic alterations that in turn could enhance similar extrapolations for cancer.
Strategy
The studies proposed for addressing recommendation 2 would yield relative sensitivity factors for mutations, chromosomal aberrations, and cell killing in rodent and human cells in vitro, and the in vivo cytogenetic studies would allow comparison of in vitro and in vivo responses in a single species, most likely the mouse. The sensitivity factors and other comparative data could then be used to provide an estimate of the responses mentioned above in humans by using cancer induction data obtained in rodents. However, the reliability of a relative sensitivity factor for this use must first be established. Chromosomal aberration and mutation frequencies induced by exposure to radiation are influenced to a great extent by the kinetics and fidelity of DNA repair processes. The relative values of these parameters for mice and humans are not well known. Therefore, a secondary measure of relative sensitivity pertinent to cancer risk assessment would be a comparison of the features of DNA repair in human and rodent cells in vitro following acute exposure to protons and HZE particles. Techniques based on pulsed-field gel electrophoresis have been developed that can measure DNA strand breaks at very low exposure levels (<10 cGy). If these experiments had to compete with other high-priority items for beam time within the total of ˜100 hours currently available per year, they would probably extend over 3 years.
|
4. |
Determine if exposure to heavy ions at the level that would occur during deep-space missions of long duration poses a risk to the integrity and function of the central nervous system. |
Strategy
A multifaceted research approach is required to address recommendation 4 so as to relate molecular changes to alterations in function. Some of the necessary experiments could take a long time, and since a few definitive answers must be obtained before final decisions about shielding and mission planning can be made, it is essential to ensure coordination of the strategy for this field of research. The studies range from the induction of DNA damage, repair, and maintenance of the fidelity of DNA into old age, to studies of the heavy-ion-induced morphological and functional changes in the central nervous system. The time necessary would vary from about 2 years for the DNA studies to perhaps 10 or more years for studies of functional changes, depending on the species required for a definitive assessment.
The scope of this research should be agreed upon by representatives of the disciplines involved, including both experimental and clinical neurologists. One essential study that could be started now is confirmation of the findings of Lett et al. on retinal cells—that late breakdown of DNA exposed to heavy ions occurs and that age at exposure is important. 43 New sensitive techniques for assessing DNA damage can be applied to the problem and also to the determination of dose-response relationships and the influence of LET. The studies described for this question could not be performed at all with only 2 weeks of beam time per year. If 3 months were available per year, then experience with similar studies44 45 46 47 suggests a rough time estimate for the performance of all the required studies of from 5 to 7 years because of the long time interval required to observe late effects. This estimate is based on the assumption that sufficient animal facilities and staff will be available at the beam site and that rodents will be used as the animal model. If rodents are not used, then another way to complete these studies in the time frame proposed might be to repeat, confirm, and extend the work of Lett et al. A minimum of three ions with a spread of LET values would have to be examined.
|
5. |
Determine if better error analyses can be performed of all factors contributing to the estimation of risk by a particular method, and determine the types and magnitude of uncertainty associated with each method. What alternate methods for calculation of risk can be used for comparison with conventional predictions in order to assess absolute uncertainties? How can these methods be used to better determine how the uncertainties in the methods affect estimates of risk to humans and estimates of mission costs? |
Strategy
The relative significance of uncertainties in risk assessments must be adequately established, and the impact of reductions in the level of these various uncertainties must be determined. The conventional approach for the assessment of risks is initially to calculate a dose, defined as the equivalent dose for the radiation field of interest corresponding to the dose of low-LET radiation that produces the same level of risk. The simplest method for obtaining the equivalent dose is to multiply the physical dose by a quality factor for the radiation field, but there are several other approaches, including models for normal-tissue responses, microdosimetric methods, and fluence-based techniques. In all of these cases, there is uncertainty associated with the method itself and additional uncertainty associated with each of the input quantities used to calculate the risk. In the former case, each of the quantities (e.g., physical dose or quality factors) required as input to establish the risk has a level of uncertainty associated with it. Reductions in the uncertainties in the values of the specific input quantities have differing effects on the magnitude of the uncertainty of the total risk, depending on the method chosen. Currently, the lack of knowledge concerning the uncertainties in the values of the quantities needed to assess risks is a major limitation in establishing realistic design requirements for a planetary mission.
In addition to uncertainties in the values of the input quantities, there is an intrinsic uncertainty associated with the method used. Since the use of only one method with a possible large uncertainty is at best questionable, it is recommended that risk estimates be determined by different, independent methods as a means of determining the overall uncertainty from input quantities and methods. The results of an error analysis (i.e., an analysis of the relative and absolute uncertainties) should be used to evaluate which methods will most effectively reduce the uncertainties in risk estimates and, therefore, uncertainties in the cost of shielding.
An analysis of the uncertainties in risk based on present data and methods could be achieved within about 1 year with proper support. Such analyses, however, should be updated routinely as part of a continuing effort throughout the entire project, and all investigators should be required to provide error analyses of their results. Lack of knowledge of cross sections for producing secondary nuclear particles in the materials used to construct a spacecraft represents a source of uncertainty that might be reduced, with a consequent potential for cost savings. However, variability in the types and energies of incident particles resulting from variation in the number and quality of solar events is not representative of an error in the input data used to calculate risk.
|
6. |
Determine how the selection and design of the space vehicle affect the radiation environment in which the crew has to exist. |
Strategy
The capability for determining effects of space vehicle selection and design is based in part on having accurate knowledge of the incident radiation field, the reaction cross sections for the incident particles reacting with vehicle materials, and the fragmentation or recoil products that such reactions produce. Current knowledge of the fragmentation products of HZE particles is limited to only a few
particles in a few materials. For knowledgeable shielding design, the initial radiation fields, reaction probabilities, and secondary particles produced as a function of angle must be determined through physical measurements, at an HZE-particle accelerator, of the particle types and energies resulting behind different compositions and thicknesses of shielding.
Based on the predictions of current transport codes, hydrogen-containing materials are preferred for shielding because they offer better shielding than other materials on a per-unit-mass basis. To properly assess the accuracy of these predictions, the transport codes used to calculate shielding efficiency have to be benchmarked against measured data for elemental (aluminum, iron, etc.) and composite shields.
Complementary measurements should be made with a microdosimetric detector of the type currently being flown in space. The absorbed dose as a function of depth should likewise be measured along the axis of the beam at selected positions along the axial plane. The measured data should be compared with predictions by the Langley Research Center transport code and/or a Monte Carlo transport code. Similar measurements as a function of depth should be made for the simplest possible geometry in space, and these results should be compared with calculations of the dose, radiation quality, and particle spectra.
Engineering of the storage for a spacecraft's supplies so as to form an enhanced "storm shelter" against transient high levels of radiation would be subject to the same verification of the accuracy of data and calculations. At the current level of availability of heavy-ion particle accelerator time, it is estimated that more than 10 years will be required to collect the necessary data.48 With increased availability of accelerator time and other resources, data collection and analysis could be compressed into a time frame of about 4 years.
|
7. |
Determine if solar particle events can be predicted with sufficient advance warning to allow crew members to return to the safety of a shielded storm shelter. |
Strategy
The ability to predict the time of occurrence and/or the magnitude of solar particle events is currently an inexact science at best. Protecting a mission crew from SPE radiation requires improving the capability to accurately predict solar events. This effort requires that information on the status of the total solar surface be continually available. One mechanism for accomplishing this would be a series of space platform monitoring stations. Given the necessary information on the status of the Sun's surface, the science and models that interpret these data must be enhanced, with the goal of achieving accurate forecasts 8 hours in advance of a spacecraft encountering an SPE. Prediction of the resources and time required to achieve this capability is beyond the expertise of the authors of this report. However, the ability to predict solar events 8 hours in advance of their occurrence is thought to be an operational requirement for a safe interplanetary mission.
Lower-Priority Research Recommendations
|
1. |
Estimate the risks of reduced fertility and sterility as a result of exposure to radiation on missions of long duration in deep space. |
Strategy
- Female: Studies of women receiving pelvic and abdominal radiotherapy in which there is good dosimetry could provide useful information on the effects of radiation on ovarian function. It is
- probable that prospective studies of women treated with cytotoxic drugs at young enough ages, in whom ovarian function is compromised, could provide valuable information when combined with modeling. Complementary studies of both normal and radiation-induced loss of ovarian follicles, preferably in a nonhuman primate, will be required.
- Male: An assessment of the effect of dose rate and protraction of radiation on spermatogenesis is essential. The study should be carried out on a primate, but studies done previously on other mammals could be extended to include low dose rate or fractionated proton exposures. Sperm counts are an easy and economical assay of the effect of exposure. However, histological studies of the testes are required, especially in cases of azoospermia (total loss of sperm). The stem cells may not be the most sensitive target, because loss of the ability of the supporting tissue to enable differentiation in the spermatogenic process may determine the probability of sterility. Paracrine mechanisms, which release locally acting substances from cells directly into intracellular space, are involved in the differentiation process, but little is known about either their role or the effects of radiation on them. Studies of men receiving cyclophosphamide (a cell-killing chemical) could provide some help in comparing the relative effects of acute and chronic administration of radiation doses on sperm production.
To improve understanding of the effects of radiation on fertility, pragmatic studies of the loss of ova or sperm and studies of the basic aspects of ovarian function or spermatogenesis should be carried out hand in hand. As much clinical data as can be obtained and are relevant should be collected, and priority should be given to animal experiments designed to answer the questions that cannot be answered from clinical data. Ideally, the studies should include the effects of repeated exposure to protons and heavy ions at low fluences. However, protracted exposure to gamma rays may be the most practical approach, and gamma rays should be an adequate surrogate for protons. Since there are several sources available for both gamma rays and protons, beam time is probably not a limiting factor for conducting this study. If the group conducting the study were co-located with the source, and the appropriate support staff and animal care facilities were available, such a study might be completed in as little as 4 years. Currently, however, such a resource does not exist.
|
2. |
Estimate the risks of clinically significant cataracts being induced by exposure to radiation at the levels that will occur on extended spaceflights. |
Strategy
A considerable body of data provides information about the induction of cataracts in different species by different types of radiation. There is, however, no consensus on how to collate the data and use them to estimate risk to humans. This objective, however, appears to be within reach and should be pursued. Another approach is to determine experimentally the relationship of RBE for cataractogenesis to LET and to apply the RBE value to the data for cataract induction in humans by low-LET radiation. A better understanding is needed of the effects of protracted exposure at low dose rates for both low- and high-LET radiation, because the data currently available for humans are for high-dose-rate, low-LET radiation.
Since research efforts on atomic bomb survivors (who were exposed to low-LET radiation) are already under way, the results of which could readily be applied to address recommendation 2, the most cost- and time-effective approach to this issue would be to ensure that current work on the survivors receives continued support.49 The cataractogenic effects of protons, the most prevalent particles in galactic cosmic rays, can be estimated directly with reasonable confidence from data on the effects of
low-LET radiation. Moreover, estimating the risk from exposure to HZE particles by any of the methods suggested so far in this report depends on the use of data obtained from humans exposed to low-LET radiation, and the major source of such data is atomic bomb survivors. Under these circumstances a sufficient answer to the question of the magnitude of the risk for cataractogenesis owing to long-duration spaceflight might be obtained in a time frame of 4 or 5 years.
|
3. |
Determine whether drugs can be used to protect against the acute or carcinogenic effects of exposure to radiation in space. |
Strategy
A program to develop drugs capable of protecting humans against the acute toxic effects of radiation has been conducted for many years under the auspices of the Department of Defense. These efforts have yielded a number of drugs that are moderately protective against the effects of low-LET radiation because they are free-radical scavengers. Such scavengers are relatively less effective against high-LET radiation because ionizations are produced more frequently as a result of direct effects rather than indirectly through the products of water radiolysis. At present, the effectiveness of such agents against acute high-dose exposure to protons, such as might be experienced during an SPE, is not known. Studies should be conducted in animal models to determine the efficacy of single doses of such drugs in protecting against the damaging effects of protons, similar to those associated with an SPE, on blood-forming cells.
More recently, studies have suggested that agents related to the compound WR-2721 may be efficacious at relatively low doses in protecting against the mutagenic and carcinogenic effects of radiation through a mechanism independent of the drug's activity as a free-radical scavenger. It is recommended that studies be pursued to determine whether such protective effects can be obtained after exposure to HZE particles. Such studies could concentrate on radiation-induced somatic cell mutagenesis, since these effects are likely to be reasonably predictive of protective effects observed for carcinogenesis in animals. Additional mechanistic studies would allow the possibility of the development of more effective and less toxic agents that might be useful for protection against late effects associated with doses resulting from SPEs. It is unlikely that a strategy of daily doses of such agents is warranted as a means of modifying the risks from daily exposure to cosmic radiation, given the relatively low risks associated with exposure at these levels, although such a strategy might be useful at the time of an SPE.
These studies would require access to appropriate facilities for irradiation with HZE particles. Under current conditions (2 weeks' available beam time per year), it is estimated that such studies would require approximately 4 years to complete, assuming that cells for these studies could be "piggybacked" with those of other cellular studies. Under more ideal conditions, with 3 months of beam time available each year, these studies would require approximately 2 years to complete.
|
4. |
Determine if there is an assay that can provide information on an individual's sensitivity to radiation-induced mutagenicity and that also can be predictive of a predisposition for susceptibility to cancer. |
Strategy
For at least 10 years, Sanford and colleagues have reported on the use of a G2 chromatid aberration assay for detecting individuals with a predisposition for cancer.50 51 52 In this assay, human lymphocytes
or cultured skin fibroblasts are irradiated with x rays, and metaphase cells arrested with the chemical colcemid between 0.5 and 1.5 hours after exposure represent cells irradiated in the G2 phase of the cell cycle. The analysis of chromatid aberrations gives a measure of chromosomal damage induced in G2; comparison of this aberration frequency with that for metaphase cells collected in the first 0.5 hour after exposure provides an estimate of DNA repair capacity. It has been reported that individuals designated as cancer prone, irrespective of the tumor type, show an enhanced frequency of aberrations and a reduced repair capacity. Attempts to duplicate the assay in other laboratories have proved unsuccessful.53 Scott et al., for example, found no difference in sensitivity between controls and lymphocytes from individuals who were homozygous or heterozygous for xeroderma pigmentosum (a DNA-repair-deficiency disease), or who had the cancer-predisposing disease familial adenomatous polyposis, or who had the cancer-predisposing syndromes Li-Fraumeni, basal cell nevus, Down's, or Fanconi's.54 They were able to show an enhancement with ataxia telangiectasia homozygotes, a very predictable result. It remains to be determined if this modified assay can detect all individuals with a cancer predisposition or at least a predisposition in specific cases. To validate the assay, it is also of considerable importance that it be conducted in several different laboratories and that an extensive sample from the general population be assessed to obtain an estimate of the range of sensitivities. Since the assay can be validated with low-LET x rays or gamma rays, no beam time is required. The analysis of at least 100 individuals in the general population and at least 10 cancer-prone families (in several laboratories simultaneously) would take approximately 2 years to complete.
|
5. |
Determine if there are differences in biological response arising from exposure to particles with similar LET, but with different atomic numbers and energies. |
Strategy
There is experimental evidence to suggest that differences in both the energy and the track structure of particles may lead to differing biological effects of exposure to radiation that are independent of LET.55 56 57 The differences in observed RBE values generally have been in the range of 2 to 3. However, the available data are derived from various sources that utilize different models and experimental conditions, thus making comparison among them difficult. Carefully designed experiments should be carried out under controlled dosimetric conditions, such that the effect of factors such as atomic number, track structure, and energy can be specifically compared in the same system. It would seem reasonable to employ well-defined experimental systems such as those proposed to address the higher-priority recommendations 1 and 2, for which substantial data are already available for various types of radiation. These would include cellular systems to examine effects on cell killing, mutagenesis, and chromosomal aberrations. If the differences observed are restricted to a factor of 2 to 3 or less, as predicted from currently available data, conducting additional experiments in animal models for tumorigenesis would not be warranted.
Based on the assumptions that the appropriate heavy ions are available, that a dedicated facility is used to minimize tuning time, and that 2 weeks of beam time per year are set aside for this strategy, the in vitro experiments could be completed in 2 years, particularly if they were carried out in parallel with those intended to address the higher-priority recommendation 2. This estimate is based on the use of three biological end points and three different LET ranges, with three particles in each range. Since it is doubtful, however, that the necessary level of resources would be reserved for lower-priority projects such as this, an estimate of 3 years is probably more realistic at current levels of availability for HZE-particle accelerator time.
If the annual available beam time were increased to 3 months, then it should still be possible to carry out this strategy in 2 years.
Time Scale Of Research
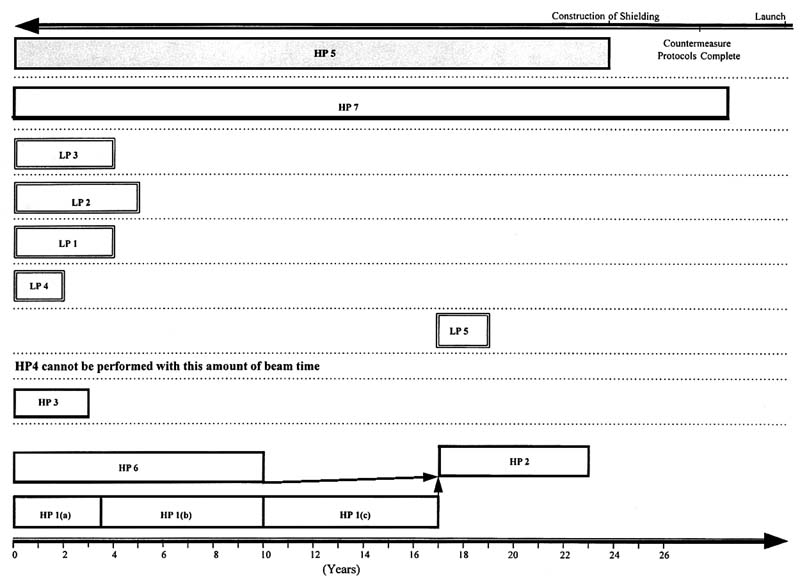
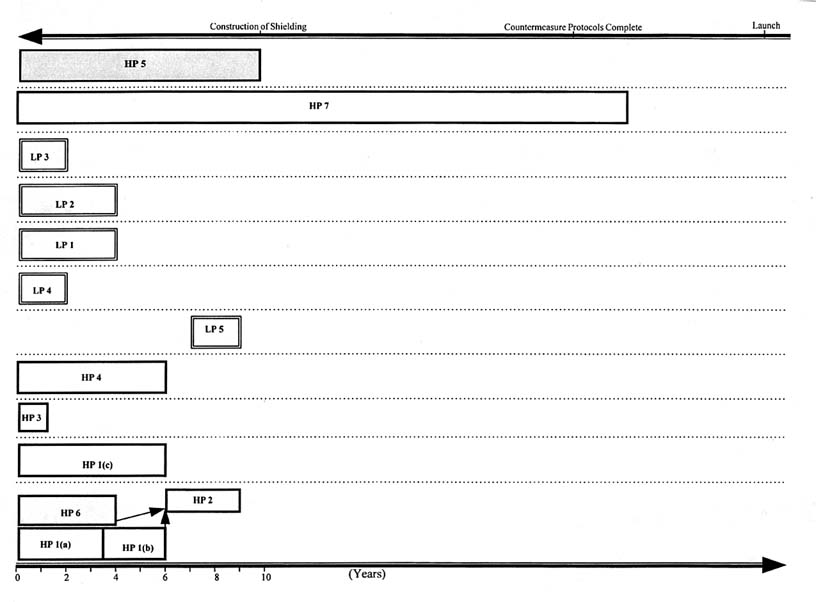
To carry out the research necessary to reduce the physical and biological uncertainties inherent in estimating risk and to design shielding to protect against a credible maximum risk, approximately 3,000 hours of beam time are required for experiments with HZE particles and energetic protons. At the present utilization rate of approximately 100 hours per year at the Brookhaven Alternating Gradient Synchrotron, the research could take more than 20 years—an unacceptably long time.
Figures 11.2 and 11.3 show potential research time lines based on the assumption that the currently available beam time at a heavy-ion accelerator of about 2 weeks per year (Figure 11.2) remains unchanged or that 3 months become available per year (Figure 11.3).
It is recommended that, if the goal of safe interplanetary missions with human crews is sought, NASA explore various possibilities, including the construction of new facilities, to increase the research time available for experiments with HZE particles.
Need For Animal Use
There are no estimates for the risk of cancer induction in humans exposed to protons, the major component of galactic cosmic radiation and solar radiation, or to heavy ions such as iron. Therefore, risk estimates currently must be based on either (1) information about the risks incurred by exposure of animals to low-LET radiation modified to allow for the different RBE values for the different types of radiation involved or (2) data from animal experiments used in conjunction with some method of extrapolating the risk estimates to humans. Both approaches are hampered by insufficient experimental data. For example, it is essential to have adequate data on the induction of cancer by radiation at a sufficient range of LET values to obtain the RBE values to estimate the risk from exposure to GCR in deep space. Obtaining such data involves the use of animals and, to a lesser extent, in vitro studies on human chromosomes and cells. Specific deterministic effects, such as reduction in fertility, cataractogenesis, and damage to the central nervous system, are important in assessing the total risk posed by prolonged sojourns in the radiation environments in space. The effects of heavy ions on the central nervous system are of particular importance. Although no information about such effects on humans is available that is suitable for setting radiation limits, it is essential that the possibility of effects on the central nervous system be adequately assessed. Because the ideal of obtaining data from primates exposed to heavy ions is unlikely to be realized, critical animal experiments must be carefully crafted and executed.
Experimental Techniques And New Data Required
This section touches on new techniques being used for the qualitative assessment of mutations and chromosomal aberrations and for the characterization of molecular events involved in tumor development. It is assumed that significant progress in the next few years will be made in these broad areas.
Estimates of cancer risk posed by low-LET radiation are quite well founded and are based on fairly extensive animal, but limited human, studies (mostly those of atomic bomb survivors). Testing the reliability of the extrapolation of results from rodent studies to humans would require a better understanding
of the mechanism of formation of specific tumor types, both background and x ray induced, for both human and animal models (with the same tumor type). Although little information is available on the genetic alterations associated with radiation-induced tumors, the methods exist to determine such alterations, and candidate genes such as the tumor suppressor gene p53 have been proposed.58 What remains to be developed are sufficiently sensitive assays for detecting mutations in nonselectable genes that could be markers of early stages in tumor development. Although specific polymerase chain reaction (PCR) methods are becoming more sensitive, they are still 1 or 2 orders of magnitude away from being able to detect induced mutations at the frequencies of occurrence required, typically mutation frequencies of 1 in 107 cells.
Limited data are available on cancer induction in rodents exposed to high-LET radiation; information on other biological effects is also sparse. It will be necessary to conduct additional cancer studies in rodents exposed to different types of high-LET radiation and to characterize the resulting tumors at the molecular level. In fact, for high-LET radiation, the conversion of DNA lesions into mutations is not well understood. To better simulate conditions of exposure during spaceflight, it is necessary to consider the effectiveness of induction of mutations by low-dose-rate exposure to both high- and low-LET radiation. The use of fluorescence in situ hybridization allows reciprocal translocations to be assessed following protracted exposure. A translocation is a significant chromosomal end point when considering genomic alterations that are associated with adverse health effects. Assays are also under development for detecting low-frequency aberrations in genes above background. Although currently available only for selectable genes such as that for hypoxanthine phosphoribosyl transferase, for which mutants have a growth advantage (i.e., they are selected for their ability to grow faster than nonmutants), it is anticipated that new assays will be available for nonselectable tumor genes and genes such as p53 and other tumor suppressor genes in the future.
The identification of populations that are genetically susceptible to cancer development is also of considerable importance. Uncovering the mechanisms involved in tumor formation is critical for this purpose but, despite considerable progress, is still a distant goal. A more attainable goal may be the development of surrogate assays for predicting increased sensitivity for tumor induction. The G2 chromosomal aberration assay described by Jones et al. is promising.59 It appears to be able to identify individuals who have at least increased radiosensitivity of lymphocytes; in one instance, this increase was quite marked in about 40 percent of breast cancer patients. More work and probably a number of modifications to the technique are in order before it can be used as a predictor of radiation sensitivity.
Ground- Versus Space-Based Research
The influence of microgravity on the effects of low-LET radiation has been reviewed by Horneck60 and was discussed by Nelson.61 Most experiments showed negligible or small effects of microgravity on radiation-induced changes. Typical changes observed had to do with increased chromosomal alterations in fruitflies and in Tradescantia (the spiderwort plant) following irradiation before liftoff. Horneck suggests that changes in chromosomal structure or position in microgravity could have prevented effective rejoining of chromosomes. On the other hand, there was no control in such experiments for vibration or acceleration during liftoff or return of the satellites. In another example cited in these reviews—an experiment measuring viability in yeast—survival was lower for microorganisms irradiated before liftoff compared to ground-based controls treated in the same way. It was noted that the difference in survival did not seem to be dose dependent. These results were interpreted as indicating that DNA repair was less efficient in microgravity. No experiments were carried out in space using a 1-g centrifuge for controls.
Recent ground-based experiments, summarized by Kronenberg, on radiation-induced DNA fragmentation, neoplastic transformation of cells plated 24 hours after irradiation, and the effects of a chemical radioprotector on mutation induction showed that DNA repair and cell recovery take place readily after low-LET irradiation, but not after exposure to HZE particles.62 Since the only reported significant effect of microgravity may be on DNA repair and cell recovery following low-LET exposure, and there seems to be no DNA repair or cell recovery following high-LET exposure, microgravity should not be important for HZE particle effects.
The above considerations indicate that HZE particles are a very important factor in the damage resulting from long space missions and that the effects of microgravity probably will not alter the cellular response to HZE particles but might actually increase the effect of low-LET radiation.
Hence, it is concluded that the majority of the useful information on radiation effects and risks will come from ground-based experiments and that radiation experiments in space, with all their logistical difficulties, will not be rewarding and may not be worth the effort.
References
1. Space Studies Board, National Research Council. 1993. Scientific Prerequisites for the Human Exploration of Space. National Academy Press, Washington, D.C.
2. Space Studies Board, National Research Council. 1996. Radiation Hazards to Crews of Interplanetary Missions: Biological Issues and Research Strategies. National Academy Press, Washington, D.C.
3. Lawrence Berkeley Laboratory. 1997. Modeling Human Risk: Cell and Molecular Biology in Context. LBNL Report 40278. U.S. Department of Energy, Washington, D.C.
4. Badhwar, G.D. 1997. The radiation environment in low-Earth-orbit. Radiat. Res. 148: S3-S10.
5. Badhwar, G.D. 1997. The radiation environment in low-Earth-orbit. Radiat. Res. 148: S3-S10.
6. Badhwar, G.D. 1997. The radiation environment in low-Earth-orbit. Radiat. Res. 148: S3-S10.
7. Yang, T.C., George, K., Johnson, A.S., Durante, M., and Federenko, B.S. 1997. Biodosimetry results from spaceflight Mir-18. Radiat. Res. 148: S17-S23.
8. Straume, T., and Bender, M.A. Issues in biological dosimetry: Emphasis on radiation environments in space. Radiat. Res. 148: S60-S70.
9. Wilson, J.W., Cucinotta, F.A., Shinn, J.L., Kim, M.H., and Badavi, F.F. 1997. Shielding strategies for human space exploration: Introduction. Chapter 1 in Shielding Strategies for Human Space Exploration: A Workshop (J.W. Wilson, J. Miller, and A. Konradi, eds.). National Aeronautics and Space Administration.
10. Malgorzata, A.W., Adelstein, S.J., and Kassis, A.I. 1998. Indirect mechanisms contribute to the biological effects produced by decay of DNA-incorporated iodine-125 in mammalian cells in vitro: Double-strand breaks. Radiat. Res. 149:134-141.
11. Board on Radiation Effects Research, National Research Council. 1990. Health Effects of Exposure to Low Levels of Ionizing Radiation: BIER V. National Academy Press, Washington, D.C.
12. Board on Radiation Effects Research, National Research Council. 1990. Health Effects of Exposure to Low Levels of Ionizing Radiation: BIER V. National Academy Press, Washington, D.C.
13. Blakely, E.A., Ngo, F.Q.H., Curtis, S.B., and Tobias, C.A. 1984. Heavy-ion radiobiology: Cellular studies. Adv. Radiat. Biol. 11: 295-389.
14. Curtis, S.B., Nealy, J.E., and Wilson, J.W. 1995. Risk cross sections and their application to risk estimation in the galactic cosmic-ray environment. Radiat. Res. 141: 57-65.
15. National Council on Radiation Protection and Measurements (NCRP). 1989. Guidance on Radiation Received in Space Activities. Recommendations of the National Council on Radiation Protection and Measurements. NCRP Report No. 98. National Council on Radiation Protection and Measurements, Bethesda, Md.
16. Preston, D.L., Kusumi, S., Tomonaga, M., Izumi, S., Ron, E., Kuramato, A., Kamada, N., Dohy, H., Matsuo, T., Nonaka, H., Thompson, D.E., Soda, M., and Mabuchi, K. 1994. Cancer incidence in atomic bomb survivors. Part III. Leukemia, lymphoma and multiple myeloma, 1950-1987. Radiat. Res. 137: S68-S97.
17. Pierce, D.A., Shimizu, Y., Preston, D.L., Vaeth, M., and Mabuchi,K. 1996. Studies of the mortality of atomic bomb survivors. Report 12, Part I. Cancer: 1950-1990. Radiat. Res. 146: 1-27.
18. Thompson, D.E., Mabuchi, K., Ron, E., Soda, M., Tokunaga, M., Ochikubo, S., Ikeda, T., Terasaki, M., Izumi, S., and Preston, D.L. 1994. Cancer incidence in atomic bomb survivors. Part II: Solid tumors, 1958-1987. Radiat. Res. 137: S17-S67.
19. International Commission on Radiological Protection (ICRP). 1991. 1990 Recommendations of the International Commission on Radiological Protection. ICRP Publication 60. Annals of the ICRP 21. Pergamon Press, Elmsford, N.Y.
20. International Commission on Radiological Protection (ICRP), 1991. 1990 Recommendations of the International Commission on Radiological Protection. ICRC Publication 60. Annals of the ICRP 21. Pergamon Press, Elmsford, N.Y.
21. National Council on Radiation Protection and Measurement (NCRP). 1997. Uncertainties in the Fatal Cancer Risk Estimates Used in Radiation Protection. Recommendations of the National Council on Radiation Protection and Measurements. NCRP Report No. 126. NCRP, Bethesda, Md., p. 73.
22. Simpson, J.A. 1983. Introduction to the Galactic Cosmic Radiation. Composition and Origin of Cosmic Rays (M.M. Shapiro, ed.). Reidel Publishing. Dordrecht, The Netherlands.
23. Simpson, J.A. 1983. Introduction to the Galactic Cosmic Radiation. Composition and Origin of Cosmic Rays (M.M. Shapiro, ed.). Reidel Publishing, Dordrecht, The Netherlands.
24. Dalrymple, G.V., Lindsay, J.R., Mitchell, J.C., and Hardy, K.A. 1991. A review of the USAF/NASA proton bioeffects project: Rationale and acute effects. Radiat. Res. 126: 117-119.
25. Urano, M., Verkey, L.J., Guitein, M., Lepper, J.E., Suit, H.D., Mendrondo, O., Gragoudos, E., and Koehler, A. 1984. Relative biological effectiveness of modulated proton beams in various murine tissues. Int. J. Oncol. Biol. Phys. 10: 509-514.
26. Storer, J.B., Harris, P.S., Furchner, J.E., and Langham, W.H. 1957. The relative biological effectiveness of various ionizing radiations in mammalian systems. Radiat. Res. 6: 189-288.
27. Clapp, N.K., Darden, D.B., Jr., and Jernigan, M.C. 1974. Relative effects of whole-body sublethal doses of 60-MeV protons and 300-kVp x rays on disease incidence in RF mice. Radiat. Res. 57: 158-186.
28. Burns, F.J., Hosselet, S., and Garte, S.J. 1989. Extrapolations of rat skin tumor incidence: Dose, fractionation and linear energy transfer. Pp. 571-582 in Low Dose Radiation: Biological Bases of Risk Assessment (K.F. Baverstock and J.W. Stather, eds.). Taylor and Francis, London.
29. Burns, F.J., Albert, R.E., Vanderlaan, M., and Strickland, P. 1975. The dose-response curve for tumor induction with single and split doses of 10 MeV protons. Radiat. Res. 62: 598 (abstract).
30. National Council on Radiation Protection and Measurements (NCRP). 1989. Guidance on Radiation Received in Space Activities. Recommendations of the National Council on Radiation Protection and Measurements. NCRP Report No. 98. National Council on Radiation Protection and Measurements, Bethesda, Md.
31. Ainsworth, E.J. 1986. Early and late mammalian responses to heavy charged particles. Adv. Space Res. 6: 153-165.
32. Merriam, G.R., Jr., Worgul, B.V., Medvedovsky, C., Zaider, M., and Rossi, H.H. 1984. Accelerated heavy particles and the lens 1 cataractogenic potential. Radiat. Res. 98: 129-140.
33. Lett, J.T., Lee, A.C., and Cox, A.B. 1991. Late cataractogenesis in rhesus monkeys irradiated with protons and radiogenic cataract in other species. Radiat. Res. 126: 147-156.
34. Worgul, B.V., Medvedovsky, C., Huang, Y., Marino, S.A., Randers-Pehrson, G., and Brenner, D.J. 1996. Quantitative assessment of the cataractogenic potential of very low doses of neutrons. Radiat. Res. 145: 343-349.
35. Committee on the Biological Effects of Ionizing Radiation, National Research Council. 1988. Health Effects of Radon and Other Internally Deposited Alpha-Emitters: BEIR IV. National Academy Press, Washington, D.C.
36. Alpen, E.L., Power-Risius, P., Curtis, S.B., DeGuzman, R., and Fry, R.J.M. 1994. Fluence-based relative biological effectiveness for charged particle carcinogenesis in mouse harderian gland. Adv. Space Res. 14: 573-581.
37. Alpen, E.L., Power-Risius, P., Curtis, S.B., DeGuzman, R., and Fry, R.J.M. 1994. Fluence-based relative biological effectiveness for charged particle carcinogenesis in mouse harderian gland. Adv. Space Res. 14: 573-581.
38. National Council on Radiation Protection and Measurements (NCRP). 1989. Guidance on Radiation Received in Space Activities. Recommendations of the National Council on Radiation Protection and Measurements. NCRP Report No. 98. National Council on Radiation Protection and Measurements, Bethesda, Md.
39. Burns, F.J., Hosselet, S., and Garte, S.J. 1989. Extrapolations of rat skin tumor incidence: Dose, fractionation and linear energy transfer. Pp. 571-582 in Low Dose Radiation: Biological Bases of Risk Assessment (K.F. Baverstock and J.W. Stather, eds.). Taylor and Francis, London.
40. Space Studies Board, National Research Council. 1996. Radiation Hazards to Crews of Interplanetary Missions: Biological Issues and Research Strategies. National Academy Press, Washington, D.C.
41. Space Studies board, National Research Council. 1996. Radiation Hazards to Crews of Interplanetary Missions: Biological Issues and Research Strategies. National Academy Press, Washington, D.C.
42. Fry, R.J.M. 1981. Experimental radiation carcinogenesis: What have we learned? Radiat. Res. 87: 224-239.
43. Lett, J.T., Keng, P.C., Bergtold, D.S., and Howard, J. 1987. Effects of heavy ions on rabbit tissues: Induction of DNA strand breaks in retinal photoreceptor cells by high doses of radiation. Radiat. Environ. Biophys. 26: 23-36.
44. Lett, J.T., Cox, A.B., Keng, P.C., Lee, A.C., Su, C.M., and Bergtold, D.S. 1980. Late degeneration in rabbit tissues after irradiation by heavy ions. Pp. 131-142 in Life Sciences and Space Research, Vol. XVIII (R. Holmquist, ed.). Pergamon Press, Oxford, England.
45. Lett, J.T., Keng, P.C., Bergtold, D.S., and Howard J. 1987. Effectsof heavy ions on rabbit tissues: Induction of DNA strand breaks in retinal photoreceptor cells by high doses of radiation . Radiat. Environ. Biophys. 26: 23-36.
46. Space Studies Board, National Research Council. 1996. Radiation Hazards to Crews of Interplanetary Missions: Biological Issues and Research Strategies. National Academy Press, Washington, D.C.
47. Williams, G.R., and Lett, J.T. 1995. Damage to the photoreceptor cells of the rabbit retina from 56Fe ions: Effect of age at exposure. Adv. Space Res. 18: 55-58.
48. Space Studies Board, National Research Council. 1996. Radiation Hazards to Crews of Interplanetary Missions: Biological Issues and Research Strategies. National Academy Press, Washington, D.C.
49. Wu, B., Medvedovsky, C., and Worgul, B.V. 1994. Non-subjective cataract analysis and its application in space radiation risk assessment. Adv. Space Res. 14: 493-500.
50. Parshad, R., Sanford, K.K., and Jones, G.M. 1983. Chromatid damage after G2 phase X-irradiation of cells from cancerprone individuals implicates deficiency in DNA repair. Proc. Natl. Acad. Sci. U.S.A. 80: 5612-5616.
51. Sanford, K.K., Parshad, R., Gantt, R., Tarone, R.E., Jones, G.M., and Price, F.M. 1989. Factors affecting and significance of G2 chromatin radiosensitivity in predisposition to cancer. Int. J. Radiat. Biol. 55: 963-981.
52. Parshad, R., Price, F.M., Pirollo, K.F., Chang, E.H., and Sanford, K.K. 1993. Cytogenetic response to G2-phase X-irradiation in relation to DNA repair and radiosensitivity in a cancer-prone family with Li-Fraumeni syndrome. Radiat. Res. 136: 236-240.
53. Bender, M.A., Viola, M.V., Riore, J., Thompson, M.H., and Leonard, R.C. 1988. Normal G2 chromosomal radiosensitivity and cell survival in the cancer family syndrome. Cancer Res. 48: 2579-2584.
54. Scott, D., Spreadborough, A.R., Jones, L.A., Robert, S.A., and Moor, C.J. 1996. Chromosomal radiosensitivity in G2-phase lymphocytes as an indicator of cancer predisposition. Radiat. Res. 145: 3-16.
55. Kranert, T., Schneider, E., and Kiefer, J. 1990. Mutation induction in V79 Chinese hamster cells by very heavy ions. Int. J. Radiat. Biol. 58: 975-987.
56. Belli, M., Cera, F., Cherubini, R., Haque, A.M., Ianzini, F., Moschini, G., Sapora, O., Simone, G., Tabocchini, M.A., and Tiveron, P. 1993. Inactivation and mutation induction in V79 cells by low energy protons: Re-evaluation of the results at the LNL facility. Int. J. Radiat. Biol. 63: 331-337.
57. Stoll, U., Schmidt, A., Schneider, E., and Kiefer, J. 1995, Killing and mutation of Chinese hamster V79 cells exposed to accelerated oxygen and neon ions. Radiat. Res. 142: 288-294.
58. Culotta, E. and Koshland, R.D.E. 1993. p53 sweeps through cancer research. Science 262: 1958-1961.
59. Jones, L.A., Scott, D., Cowan, R., and Roberts, S.A. 1995. Abnormalradiosensitivity of lymphocyte from breast cancer patients with excessive normal tissue damage after radiotherapy: Chromosome aberrations after low-dose-rate irradiation . Int. J. Radiat. Biol. 67: 519-528.
60. Horneck, G. 1992. Radiobiological experiments in space: A review. Int. J. Radiat. Appl. Instrum. 20: 82-205.
61. Nelson, G., Space-based radiation biology, presentation to the Task Group on the Biological Effects of Space Radiation, November 13, 1995, Washington, D.C.
62. Kronenberg, A., NASA space radiation health program: Ground-based radiobiology research program, presentation to the Task Group on the Biological Effects of Space Radiation, Washington, D.C.























