
3
Which Nutrients Matter?
KEY POINTS IN CHAPTER 3
This chapter reviews the nutrients that exert the greatest control on eutrophication:
-
Phosphorus is the nutrient usually controlling freshwater lake eutrophication.
-
In contrast, eutrophication in most coastal marine ecosystems is primarily controlled by nitrogen.
-
Even though nitrogen usually controls eutrophication of coastal systems, it is important to manage both phosphorus and nitrogen inputs since phosphorus is important in some of systems and since managing only nitrogen without also managing phosphorus inputs can lead to a situation where phosphorus becomes the nutrient controlling eutrophication.
-
A variety of ecological and biogeochemical mechanisms lead to these differences between freshwater and coastal marine ecosystems, including the relative inputs of nutrients from adjoining systems, the preferential storage or recycling of nitrogen versus phosphorus within the ecosystem, and the extent to which nitrogen fixation can alleviate nitrogen shortages.
-
Eutrophication of coastal systems is often accompanied by decreased silica availability and increased iron availability, both of which may promote the formation of harmful algal blooms.
The major nutrients that cause eutrophication and other adverse impacts associated with nutrient over-enrichment are nitrogen and phosphorus. In this chapter, we discuss why nitrogen is of paramount importance in both causing and controlling eutrophication in coastal marine ecosystems. This is in contrast to lakes, where eutrophica-
tion is largely due to excess inputs of phosphorus. Also discussed in this chapter are other elements—particularly silicon and iron—that may be important in regulating harmful algal blooms in coastal waters and in determining some of the consequences of eutrophication.
NITROGEN AND PHOSPHORUS IN ESTUARIES AND LAKES
After extensive study in the early 1970s, a consensus developed that phosphorus was the nutrient most responsible for nutrient over-enrichment in freshwater lakes (Edmondson 1970; Vollenweider 1976; Schindler 1977), and since then better control of phosphorus loadings to lakes has gone a long way toward mitigating freshwater eutrophication (Carpenter et al. 1998). In contrast, research indicates that in numerous estuaries and coastal marine ecosystems (at least in the temperate zone) nitrogen generally is more limiting to primary production by phytoplankton, and nitrogen inputs are more likely to accelerate eutrophication (Howarth 1988; Vitousek and Howarth 1991; Nixon 1995; Paerl 1997). Note that the concept of “nutrient limitation” is often poorly defined and used rather loosely; the committee follows the definition of control of the potential rate of primary production, allowing for potential changes in the composition of the ecosystem (Howarth 1988). Thus, a nutrient is limiting if its addition to the system increases the rate of net primary production.
There are exceptions to the generality that nitrogen is limiting in coastal ecosystems. For instance, certain temperate estuaries, such as the Apalachicola on the Gulf coast of Florida and several estuaries on the coast of the Netherlands in the North Sea appear to be phosphorus limited (Myers and Iverson 1981; Postma 1985; Brockman et al. 1990). In the case of the North Sea estuaries, phosphorus limitation is probably the result of extremely high nitrogen inputs combined with fairly stringent control of phosphorus inputs (Howarth et al. 1995, 1996). In the case of the Apalachicola, phosphorus limitation results from a relatively high ratio of nitrogen to phosphorus in nutrient inputs, although in this case the high ratio may reflect the relatively small amount of human disturbance in the watershed and the relatively low nutrient inputs overall (Howarth 1988; Billen et al. 1991).
For nearshore tropical marine systems, it is commonly believed that phosphorus is more limiting of primary production (Howarth et al. 1995). This is probably true for many tropical lagoons with carbonate sands that are relatively unaffected by human activity (Smith and Atkinson 1984; Short et al. 1990). However, such lagoons may move toward nitrogen limitation as they become eutrophic (McGlathery et al. 1994; Jensen et al. 1998). Also, even oligotrophic tropical seas may be nitrogen limited away
from shore; for example, much of the Caribbean Sea away from the immediate shorelines appears to be nitrogen limited (Corredor et al. 1999).
Nutrient limitation of primary production switches seasonally between nitrogen and phosphorus in some major estuaries, such as Chesapeake Bay (Malone et al. 1996) and in portions of the Gulf of Mexico, including the “dead zone” (Rabalais et al. 1999). Even in these systems, nitrogen is probably the nutrient responsible for the major impacts of eutrophication. The production of most of the biomass that sinks into bottom waters and leads to low-oxygen events is more likely to be controlled by nitrogen than by phosphorus; when primary production is phosphorus limited in these systems, relatively little of the production tends to sink out of the water column (Gilbert et al. 1995; Malone et al. 1996; Rabalais et al. 1999).
Acceptance of the need to better control nitrogen inputs to coastal marine waters has been slower than acceptance of phosphorus control to manage eutrophication in freshwater systems. Many marine scientists recognized the nitrogen problem decades ago, yet the need for nitrogen control was hotly debated throughout the 1980s (NRC 1993a). For some locations, such as the Baltic Sea, the debate continues (Hellström 1996; Elmgren and Larsson 1997; Hecky 1998; Howarth and Marino 1998). Nonetheless, efforts to manage coastal eutrophication by controlling nitrogen inputs lag far behind the widespread success in managing lake eutrophication by controlling phosphorus inputs (NRC 1993a).
EVIDENCE FOR NITROGEN LIMITATION IN COASTAL MARINE ECOSYSTEMS
Most of those who in the 1980s disagreed with the assertion that nitrogen is the key to regulating marine eutrophication in coastal marine systems instead argued that phosphorus is the critical nutrient, as in lakes. In general, they either challenged the type of evidence used by marine scientists to infer nitrogen limitation of primary production (Smith 1984; Hecky 1998; Hecky and Kilham 1988), or doubted that there was a reason to believe that eutrophication in coastal marine systems is different in any fundamental way from eutrophication in lakes. Both arguments, however, have now largely been refuted.
Most early studies of nutrient limitation and eutrophication in coastal waters relied on fairly short-term and small-scale enrichment experiments to infer limitation by nitrogen (Ryther and Dunstan 1971; Vince and Valiela 1973) or made inferences from pure-culture studies (Smayda 1974). When applied to the problem of lake eutrophication in the 1960s and early 1970s, these approaches often led to the erroneous conclusion that nitrogen or carbon, rather than phosphorus, was limiting in lakes. Later,
whole-lake experiments clearly showed that phosphorus and not nitrogen or carbon was the key nutrient regulating eutrophication in lakes (Schindler 1977). Consequently, the scientific community that studied lake eutrophication and the lake water quality management community developed an appropriate skepticism for bioassay experiments (NRC 1993a). Other types of evidence also have been used to infer nitrogen limitation in coastal ecosystems, including relatively low ratios of dissolved inorganic nitrogen to phosphorus (Boynton et al. 1982). These approaches also can be criticized, since concentrations of dissolved inorganic nutrients do not always accurately reflect their biological availabilities (Howarth 1988; Howarth and Marino 1990).
Ecosystem-scale experiments were the galvanizing force that led to the clear conclusion that eutrophication in lakes is best managed through controlling phosphorus inputs (Schindler 1977; NRC 1993a). A decade ago, there were no comparable experiments testing the relative importance of nitrogen and phosphorus as regulators of eutrophication in coastal marine ecosystems (Howarth 1988). However, since 1990 the results of three larger-scale enrichment experiments in estuaries have been published, all clearly showing nitrogen limitation in the systems (Howarth and Marino 1998).
One of these experiments, a mesocosm experiment conducted at the Marine Ecosystem Research Laboratory (MERL) on the shores of Narragansett Bay in Rhode Island, was specifically designed to see if coastal systems respond to nutrient additions in the same manner as lakes (Oviatt et al. 1995). Large mesocosms containing water and sediment from Narragansett Bay were maintained for a period of four months; many previous studies in MERL mesocosms has demonstrated that these systems accurately mimic much of the ecological functioning of Narragansett Bay. In this experiment, mesocosms received no nutrient enrichment or were enriched with nitrogen, phosphorus, or both. The level of nitrogen and phosphorus enrichment paralleled those used in a whole-lake eutrophication experiment at the Experimental Lakes Area in Canada, an experiment where phosphorus inputs clearly led to eutrophication and where nitrogen had no effect on rates of primary production (Schindler 1977). In sharp contrast, the addition of nitrogen (either alone or with phosphorus) but not of phosphorus alone to MERL coastal mesocosms caused large increases in both rates of primary production (Figure 3-1A) and the abundance of phytoplankton (Figure 3-1B; Oviatt et al. 1995).
Another whole-ecosystem estuarine study followed the impacts of experimental alteration of nutrient releases from a sewage treatment plant into Himmerfjarden, an estuary south of Stockholm, Sweden, on the Baltic Sea. The response of the estuary to nutrient inputs from sewage (the
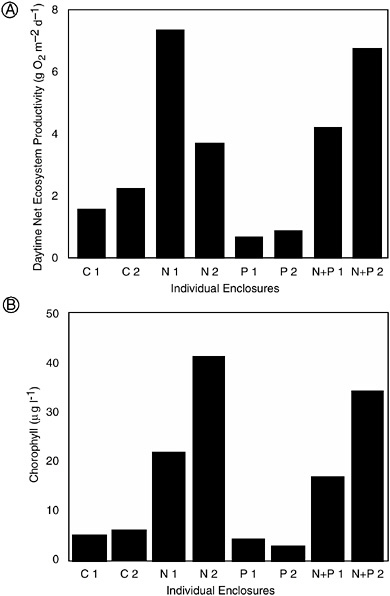
FIGURE 3-1 Response of estuarine mesocosms in Narragansett, Rhode Island, to experimental nutrient additions, clearing showing nutrient limitation by nitrogen but not phosphorus. (A) Mean daytime rate of net ecosystem production. (B) Mean chlorophyll concentration (an indicator of phytoplankton biomass). On the x-axis, C are control systems that received no nutrient additions, N are systems that were enriched with nitrogen, P are systems that were enriched with phosphorus, and N and P are systems that received both nitrogen and phosphorus. The numbers show replicate systems (two replicates for each treatment). The experiment ran for two months in the summer (modified from Oviatt et al. 1995).
primary input to this system) was studied from 1976 to 1993 (Elmgren and Larsson 1997). For the first 12 years, nitrogen loads gradually increased while phosphorus loads gradually decreased. For a one-year period beginning in the fall of 1983, phosphorus additions were greatly increased (by stopping the phosphorus removal during sewage treatment). Subsequently, phosphorus removal was again used, but nitrogen inputs were increased by 40 percent in 1985 as a result of an increase in population served by this particular sewage treatment plant. Finally, nitrogen removal technology was gradually introduced to the sewage treatment plant between 1988 and 1993, gradually reducing the nitrogen load to the value originally seen in 1976.
Throughout the 17 years of observation, the concentration of total nitrogen tended to reflect the nitrogen input from the sewage treatment plant (Elmgren and Larsson 1997), and both abundances of phytoplankton (Figure 3-2A) and water clarity (Figure 3-2B) were clearly related to the total nitrogen concentration. Total phosphorus concentrations varied independently of total nitrogen over time in Himmerfjarden, and total phosphorus was a poor predictor of phytoplankton abundances. This is strong evidence that nitrogen was the element most controlling eutrophication in this estuary. During the year that phosphorus loadings were experimentally increased, there was no effect on primary production; however, there was an unusually large bloom the following spring, probably due both to some residual high levels of phosphorus and to an unusually high input of nitrogen from spring floods (Elmgren and Larsson 1997).
A third whole-ecosystem study explored long-term changes in Laholm Bay, an estuary on the southwestern coast of Sweden (Figure 3-3). Early signs of eutrophication appeared there in the 1950s and 1960s and steadily increased over time (Rosenberg et al. 1990). The earliest reported signs of eutrophication were changes in the community composition of macroalgae species, and over time filamentous algae typical of eutrophic conditions have become more prevalent. Harmful algal blooms have become much more common, particularly in the 1980s (Rosenberg et al. 1988, 1990). During the early stages of eutrophication in Laholm Bay, inputs of both phosphorus and nitrogen to the estuary were increasing. However, from the late 1960s through the 1980s, phosphorus inputs decreased by a factor of almost two, while nitrogen inputs continued to increase (more than doubling) (Rosenberg et al. 1990). During this same period, plankton blooms continued, clearly indicating that nitrogen controlled the Laholm Bay eutrophication.
These three ecosystem-scale experiments show only that nitrogen controlled eutrophication in Narragansett Bay, Himmerfjarden, and Laholm Bay. Importantly, however, the finding in each of these three systems is
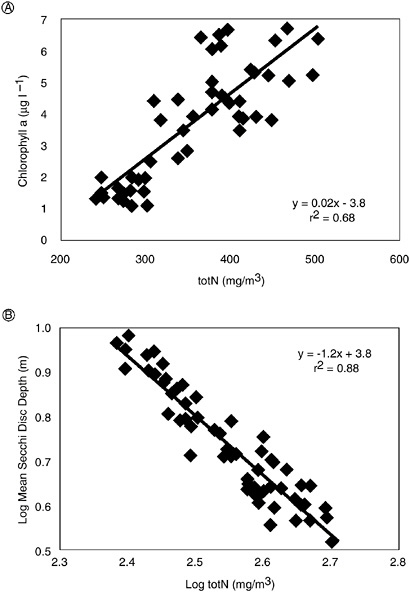
FIGURE 3-2 Long-term observations in the Himmerfjarden estuary south of Stockholm, Sweden, over a period of years in which nitrogen and phosphorus additions were experimentally altered through changes in sewage treatment. (A) The relationship between the mean concentration of total nitrogen and the chlorophyll a in the surface water layer (modified from Elmgren and Larsson 1997). (B) The relationship between the mean concentration of total nitrogen in the surface water layer and the water clarity (secchi disc depth) (modified from Elmgren and Larsson 1997). Note that the major period of eutrophication in this estuary coincided with a period in which nitrogen inputs were increasing yet phosphorus inputs were decreasing.
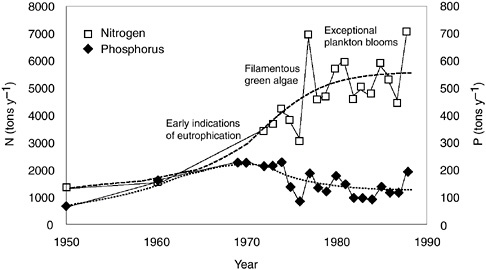
FIGURE 3-3 Transport of nutrients to Laholm Bay, Sweden. Periods of significant changes in the marine biota are also indicated (modified from Rosenberg et al. 1990).
consistent with conclusions drawn from short-term bioassay studies and from ratios of dissolved inorganic nitrogen:phosphorus in these ecosystems (Granéli et al. 1990; Oviatt et al. 1995; Elmgren and Larsson 1997; Howarth and Marino 1998). These three ecosystem experiments therefore add credence to the application of bioassay data and inorganic nutrient data in assessing whether nitrogen or phosphorus is more limiting in estuaries. The large preponderance of bioassay data in estuaries and coastal marine systems indicates nitrogen limitation (Howarth 1988), as does the generally low inorganic nitrogen:phosphorus ratio found in most estuaries at the time of peak primary production (Figure 3-4; Boynton et al. 1982). Thus, taken together the three whole-ecosystem scale and bioassay data from many sites lead to the conclusion that nitrogen availability is the primary regulator of eutrophication in most coastal systems.
MECHANISMS THAT LEAD TO NITROGEN LIMITATION IN COASTAL MARINE ECOSYSTEMS
What ecological or biogeochemical mechanisms can lead to nitrogen control of eutrophication in most coastal marine systems and to phosphorus control in so many freshwater lakes? This question was reviewed by Howarth (1988) and Vitousek and Howarth (1991). Here we summarize
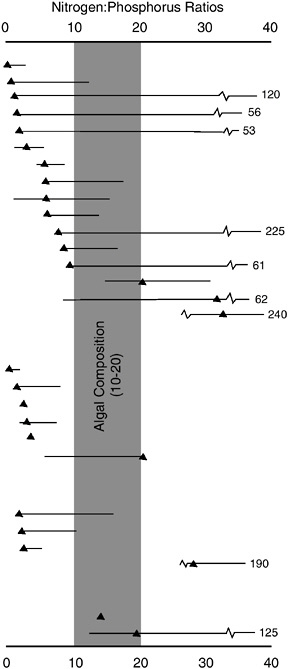
FIGURE 3-4 Summary of nitrogen:phosphorus ratios in 28 sample estuarine ecosystems. Horizontal bars indicate the annual ranges in nitrogen:phosphorus ratios; solid triangles represent the ratio at the time of maximum productivity. Vertical bands represent the typical range of algal composition ratios (modified from Boynton et al. 1982).
and update those reviews. Whether primary production by phytoplankton is nitrogen or phosphorus limited is a function of the relative availabilities of nitrogen and phosphorus in the water. Phytoplankton require approximately 16 moles of nitrogen for every mole of phosphorus they assimilate (the Redfield ratio of nitrogen:phosphorus=16:1) (Redfield 1958). If the ratio of available nitrogen to available phosphorus is less than 16:1, primary production will tend to be nitrogen limited. If the ratio is higher, production will tend to be phosphorus limited.
The relative availabilities of nitrogen and phosphorus to the phytoplankton is determined by three factors (Figure 3-5):
-
the ratio of nitrogen to phosphorus in inputs to the ecosystem;
-
preferential storage, recycling, or loss of one of these nutrients in the ecosystem; and
-
the amount of biological nitrogen fixation.
For each of these factors, there are reasons why nitrogen limitation tends to be more prevalent in coastal marine ecosystems than in lakes. For instance, lakes receive nutrient inputs from upstream terrestrial ecosystems and from the atmosphere, while estuaries and coastal marine systems receive nutrients from these sources as well as from neighboring oceanic water masses. For estuaries such as those along the northeastern coast of the United States, the ocean-water inputs of nutrients tend to have a nitrogen:phosphorus ratio well below the Redfield ratio due to denitrification on the continental shelves (Nixon et al. 1995, 1996). Thus, given similar nutrient inputs from land, estuaries are more likely to be more nitrogen limited than are lakes.
Another factor to consider is that the ratio of nitrogen to phosphorus in nutrient inputs from land will tend to reflect the extent of human activity in the landscape. As the landscape changes from one dominated by forests to one dominated by agriculture and then industry, total nutrient fluxes from land increase for both nitrogen and phosphorus, but the change is often greater for phosphorus and so the nitrogen:phosphorus ratio tends to fall (Billen et al. 1991; Howarth et al. 1996). This, too, influences why nitrogen limitation is of primary importance in estuaries (NRC 1993a). The occurrence of phosphorus limitation in the Apalachicola estuary, for instance, may be the result of the relative low level of human activity in most of the watershed. This suggests that there is a tendency for estuaries to become more nitrogen limited as they become more affected by humans and as nutrient inputs increase overall (Howarth et al. 1995).
The biogeochemical processes active in an aquatic ecosystem affect the availability of nutrients to phytoplankton in that particular system.
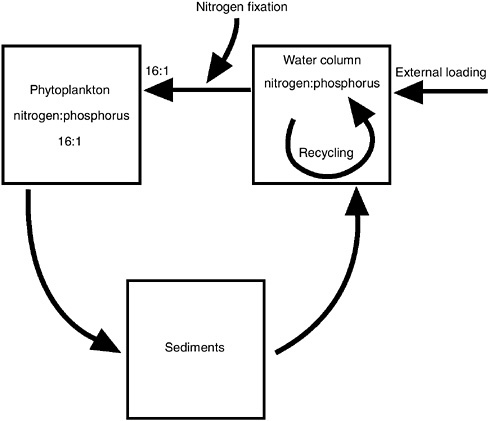
FIGURE 3-5 Factors that determine whether nitrogen or phosphorus is more limiting in aquatic ecosystems, where one of these macronutrients is limiting to net primary production. Phytoplankton use nitrogen and phosphorus in the approximate molar ratio of 16:1. The ratio of available nitrogen in the water column is affected by: 1) the ratio of nitrogen:phosphorus in external inputs to the ecosystem; 2) the relative rates of recycling of nitrogen and phosphorus in the water column, with organic phosphorus usually cycling faster than organic nitrogen; 3) differential sedimentation of nitrogen in more oligotrophic systems; 4) preferential return of nitrogen or phosphorus from sediments to the water column due to processes such as denitrification and phosphorus adsorption and precipitation; and 5) nitrogen fixation (modified from Howarth 1988; Howarth et al. 1995).
Of these processes, the sediment processes of denitrification and phosphate adsorption are the dominant forces that affect the relative importance of nitrogen or phosphorus limitation on an annual or greater time scale. Other processes, such as preferential storage of phosphorus in zooplankton (Sterner et al. 1992), act only over a relatively short period.
Denitrification is often a major sink for nitrogen in aquatic ecosystems, and it tends to drive systems toward nitrogen limitation unless counter-balanced by other processes such as phosphorus adsorption and storage (Howarth 1988; Seitzinger 1988; Nixon et al. 1996). The overall magnitude of denitrification tends to be greater in estuaries than in freshwater ecosystems, but this may simply be a result of greater nitrogen fluxes through estuaries (Seitzinger 1988). When expressed as a percentage of the nitrogen input to the system lost through denitrification, there appears to be relatively little difference between estuaries and freshwater ecosystems (Nixon et al. 1996). That is, available evidence indicates that denitrification tends to drive both coastal marine and freshwater ecosystems toward nitrogen limitation, with no greater tendency in estuaries. In fact, the tendency toward nitrogen limitation—based on this process alone—might be greater in lakes, since lakes generally have a longer water residence time, and the percent nitrogen loss through denitrification is greater in ecosystems having a longer water residence time (Howarth et al. 1996; Nixon et al. 1996).
A sediment process counteracting the influence of denitrification on nutrient limitation is phosphorus adsorption. Sediments potentially can absorb and store large quantities of phosphorus, making the phosphorus unavailable to phytoplankton and tending to drive the system toward phosphorus limitation. This process is variable among ecosystems (Howarth et al. 1995). At one extreme, little or no phosphorus is adsorbed by the sediments of Narragansett Bay, and virtually all of the phosphate produced during decomposition in the sediments is released back to the water column (Nixon et al. 1980). This, in combination with nitrogen lost through denitrification, is a major reason that Narragansett Bay is nitrogen limited (Figure 3-6; Nixon et al. 1980; Howarth 1988). Caraco et al. (1989, 1990) suggested that lake sediments have a greater tendency to adsorb and store phosphorus than do estuarine sediments; if this were true, this differential process would make phosphorus limitation more likely in lakes than in estuaries. However, the generality of a difference in phosphorus retention between lakes and coastal marine sediments has yet to be established. It is also important to note that eutrophication may lead to less denitrification since the coupled processes of nitrification and denitrification are disrupted in anoxic waters.
Among estuaries, the ability of sediments to adsorb phosphorus is variable (Howarth et al. 1995), with little or no adsorption occurring in systems such as Narragansett Bay and almost complete adsorption of inorganic phosphate in some other systems, such as those along the coast of the Netherlands (van Raaphost et al. 1988). Chesapeake Bay sediments show an intermediate behavior, with some of the inorganic phosphorus released during sediment decomposition being adsorbed and some
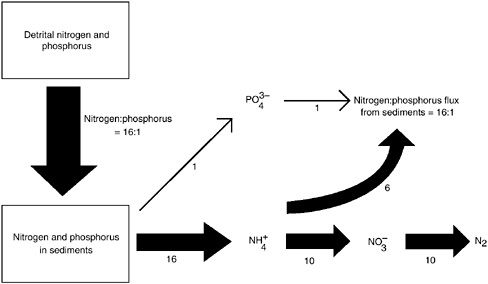
FIGURE 3-6 Schematic diagram showing nutrient regeneration from the sediments of Narragansett Bay, Rhode Island. Nitrogen and phosphorus enter the sediments as particulate matter in approximately the Redfield ratio of 16:1. Phosphorus mineralized during decomposition is released back to the water column, whereas much of the mineralized nitrogen is lost through the combined processes of nitrification and denitrification (modified from Nixon et al. 1980; Howarth 1988).
released to the overlying water (Boynton and Kemp 1985). The reasons for this difference in behavior among systems are not well understood. However, there is some indication that the ability of coastal marine sediments—both in tropical and in temperate systems—to adsorb and store phosphorus decreases as an ecosystem becomes more eutrophic, at least until they become extremely hypereutrophic, as in the case of some of the estuaries in the Netherlands (Howarth et al. 1995). For temperate systems, the lessened ability to sorb phosphate as a system becomes more eutrophic results from decreased amounts of oxidized iron and more iron sulfides in the sediments; for tropical carbonate systems, the rate of sorption of phosphate decreases as the phosphorus content of the sediment increases. These changes result in an increase in phosphorus availability in eutrophic systems, intensifying nitrogen limitation and encouraging the growth of algae and other organisms (including some heterotrophic organisms, such as the heterotrophic life stages of Pfiesteria) with high phosphorus requirements.
The process of nitrogen fixation clearly has different effects on nutrient limitation in freshwater lakes and coastal marine ecosystems. If a lake of moderate productivity is driven toward nitrogen limitation, blooms of heterocystic, nitrogen-fixing cyanobacteria (“blue-green algae”) occur, and these tend to fix enough nitrogen to alleviate the nitrogen shortage (Schindler 1977; Howarth et al. 1988a). Primary productivity of the lake remains limited by phosphorus (Schindler 1977). This was demonstrated experimentally in whole-lake experiments at the Experimental Lakes Area in northwestern Ontario, where a lake was fertilized with a constant amount of phosphorus over several years. For the first several years, the lake also received relatively high levels of nitrogen fertilizer, so that the ratio of nitrogen:phosphorus of the fertilization treatment was above the Redfield ratio of 16:1 (by moles). Under these conditions, no nitrogen fixation occurred in the lake. The regime was then altered so that the lake received the same amount of phosphorus, but the nitrogen input was decreased so that the nitrogen:phosphorus ratio of the inputs was below the Redfield ratio. Nitrogen-fixing organisms quickly appeared and made up the nitrogen deficit (Schindler 1977; Flett et al. 1980). This response is a major reason that nitrogen limitation is so prevalent in mesotrophic and eutrophic lakes (Schindler 1977; Howarth 1988).
Estuaries and eutrophic coastal waters provide a striking contrast to this behavior. With only a few exceptions anywhere in the world, nitrogen fixation by planktonic, heterocystic cyanobacteria is immeasurably low in mesotrophic and eutrophic coastal marine systems, even when they are quite nitrogen limited (Horne 1977; Doremus 1982; Fogg 1987; Howarth et al. 1988b; Paerl 1990; Howarth and Marino 1990, 1998). This major difference in the behavior between lakes and estuaries allows nitrogen limitation to continue in estuaries (Howarth 1988; Vitousek and Howarth 1991).
Much research has been directed at the question of why nitrogen fixation by planktonic organisms differs between lakes and coastal marine ecosystems, with much of this focused on single-factor controls, such as short residence times, turbulence, limitation by iron, limitation by molybdenum, or limitation by phosphorus (Howarth and Cole 1985; Paerl 1985; Howarth et al. 1988a; Howarth et al. 1999; Paerl and Zehr 2000). A growing consensus has developed, however, that nitrogen fixation in marine systems—estuaries, coastal seas, as well as oceanic waters—probably is regulated by complex interactions of chemical, biotic, and physical factors (Howarth et al. 1999; Paerl and Zehr 2000). With regard to estuaries and coastal seas, recent evidence indicates that a combination of slow growth rates caused by low availabilities of trace metals required for nitrogen fixation (iron and/or molybdenum) and grazing by zooplankton and
benthic animals combine to exclude nitrogen-fixing heterocystic cyanobacteria (Figure 3-7; Howarth et al. 1999).
Nitrogen fixation by planktonic cyanobacteria does occur in a few coastal marine ecosystems, notably the Baltic Sea and the Peel-Harvey inlet in Australia (Howarth and Marino 1998). In the Baltic, rates of nitrogen fixation are not sufficient to fully alleviate nitrogen limitation (Granéli et al. 1990; Elmgren and Larsson 1997). The reason that nitrogen fixation occurs in the Baltic but not in most other estuaries and seas remains disputed (Hellström 1998; Howarth and Marino 1998), but a model based on the interplay of trace metal availability and grazing as controls on nitrogen fixation correctly predicts that nitrogen fixation would occur in the Baltic but not in most estuaries (Howarth et al. 1999); this model result is driven by the greater availability of trace metals at the low salinity of the Baltic compared to most estuaries. The reason why nitrogen fixation occurs in the Peel-Harvey (Lindahl and Wallstrom 1985; Huber 1986), and also a similar estuary in Tasmania (Jones et al. 1994), remains unknown. One hypothesis is that this is a result of extreme eutrophication, which has driven these systems anoxic, increasing trace metal availability and lowering grazing by animals (Howarth and Marino 1998; Howarth et al. 1999). Nitrogen fixation in both estuaries has only begun in the recent past, and only as they became extremely eutrophic.
Oceanographic scientists have long believed that phosphorus is the long-term regulator of primary production in the oceans as a whole (Redfield 1958; Broecker 1974; Howarth et al. 1995). In this view, nitrogen limitation can occur in oceanic surface waters, but this is a transient effect that is made up for by nitrogen fixation over geological time scales. Recently, Tyrrell (1999) formalized this concept with a simple, 6-variable model of nutrient cycling and primary production in the world’s oceans. The basic concept is appealing, if as yet unproven, in that it explains the strong correlation of dissolved nitrogen and phosphorus compounds over depth profiles in the oceans.
Based on this conceptual view of the interaction of nitrogen and phosphorus over geological time scales, Tyrrell (1999) concluded that coastal eutrophication is largely a phosphorus problem, and that “removal of nitrates in the river supply should lead to increased nitrogen fixation, no significant effects on final nitrate concentrations, and no significant effect on eutrophication.” The Committee on Causes and Management of Coastal Eutrophication disagrees strongly. While nitrogen fixation in oceanic waters may alleviate nitrogen deficits over tens of thousands of years, nitrogen fixation simply does not occur in most estuaries and coastal seas and does not alleviate nitrogen shortages. Therefore, decreasing nitrogen inputs to estuaries will not in general lead to increased nitrogen fixation. The Tyrrell model operates on geological time scales for oceans as the
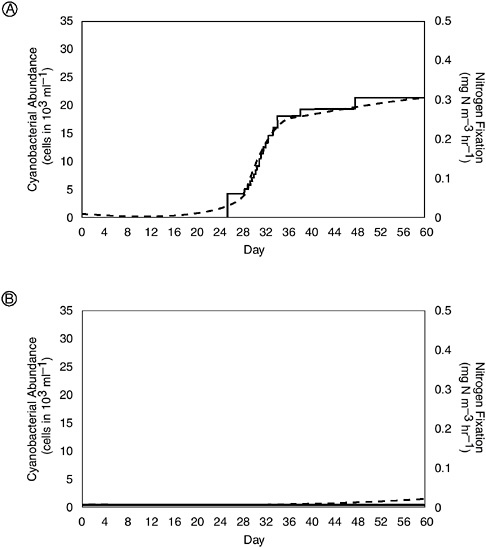
FIGURE 3-7 Results from a simple simulation model designed to show the importance of top-down and bottom-up factors as regulators of nitrogen fixation in lakes and estuaries. (A) Shown are the number of cyanobacterial cells (dashed line) and rates of nitrogen fixation (solid line) over time under typical conditions for a freshwater lake and (B) under identical conditions except for a lower availability of molybdenum as one would find in an estuary. Molybdenum is required for nitrogen fixation, and its lower abundance leads to slower growth rates by cyanobacteria. In combination with typical rates of grazing by zooplankton, this slower rate of growth in estuaries excludes nitrogen-fixing organisms that flourish in lakes (modified from Howarth et al. 1995).
whole, a time scale not applicable to estuaries, coastal seas, and continental shelves where water residence time varies from less than one day to at most a few years. Nitrogen fixation does occur in the Baltic Sea, yet even there the water residence time is on the scale of a few decades, thousands-fold shorter than the time scale of response by nitrogen fixation in Tyrrell’s model. While debate continues as to whether or not nitrogen fixation completely alleviates nitrogen shortages in the Baltic, much evidence shows that it does not and that much of the Baltic Sea remains nitrogen limited (Granéli et al. 1990; Elmgren and Larsson 1997; Hellström 1998; Howarth and Marino 1998; Savchuck and Wulff 1999).
THE IMPORTANCE OF SILICA AND IRON IN COASTAL SYSTEMS
Although nitrogen is the element primarily controlling eutrophication in estuaries and coastal seas, and phosphorus is the element primarily controlling eutrophication in lakes, other elements can have a major influence on the community structure of aquatic ecosystems and can influence the nature of the response to nutrients. A key element in this regard is silica, an element required by diatoms. The availability of silica in a waterbody has little or no influence on the overall rate of primary production, but when silica is abundant, diatoms are one of the major components of the phytoplankton. When silica is in low supply, other classes of algae dominate the phytoplankton composition.
Inputs of biologically available silica to aquatic systems come largely from weathering of soils and sediments. The major human influence on silica delivery to coastal marine systems is to decrease it, as eutrophication in upstream ecosystems tends to trap silica before it reaches the coast (Schelske 1988; Conley et al. 1993; Rabalais et al. 1996; Turner et al. 1998). Thus, the concentration of silicate in Mississippi River water entering the Gulf of Mexico decreased by 50 percent from the 1950s to the 1980s (Figure 3-8), a time during which nitrogen and phosphorus fluxes and concentrations increased (Goolsby et al. 1999). Eutrophication in a system can further decrease silica availability as it is incorporated into diatoms and stored in bottom sediments, as demonstrated in the Baltic Sea (Wulff et al. 1990). As discussed in Chapter 4, a decrease in silica availability, particularly if accompanied by increases in nitrogen, may encourage the formation of some blooms of harmful algae as competition with diatoms is decreased (NRC 1993a; Rabalais et al. 1996). As noted by Smayda (1989), for all cases where long-term data sets are available on silica availability in coastal waters, a decrease in silica availability relative to nitrogen or phosphorus has been correlated with an increase in harmful algal blooms.
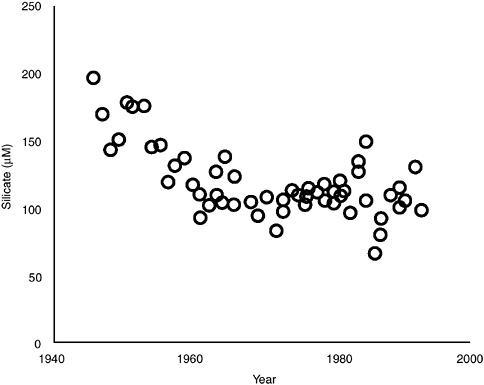
FIGURE 3-8 The concentration of dissolved silicate in waters of the Mississippi River near the Gulf of Mexico from the 1940s into the 1990s. Note the decrease during the 1950s and 1960s, probably in response to eutrophication in upstream freshwater ecosystems trapping silicate (modified from Rabalais et al. 1996).
Decreasing silica availability and the consequent lower abundances of diatoms also lowers organic matter sedimentation and thereby have a partially mitigating influence on low-oxygen events associated with eutrophication. In many coastal systems there may, however, still be sufficient silica to fuel diatom blooms during the critical spring bloom period when the majority of sedimentation often occurs (Conley et al. 1993; Turner et al. 1998). Further, eutrophication can lead to other complex shifts in trophic structure that might either increase or decrease the sedimentation of organic carbon (Turner et al. 1998).
Iron is another element that can affect the community composition of phytoplankton. As discussed in Chapter 4, greater availability of iron may encourage some harmful algal blooms. In some oceanic waters away from shore, iron availability appears to be a major control on rates of
primary production (Martin et al. 1994; Coale et al. 1996). However, there is no evidence that iron limits primary production in estuaries and coastal seas (although it may partially limit nitrogen-fixing cyanobacteria in estuaries) (Howarth and Marino 1998; Howarth et al. 1999). Although iron concentrations are lower in estuaries than in freshwater lakes, concentrations in estuaries and coastal seas are far greater than in oceanic waters (Marino et al. 1990; Schlesinger 1997). The solubility of iron in seawater and estuarine waters is low, and complexation with organic matter is critical to keeping iron in solution and maintaining its biological availability. Eutrophication tends to increase the amount of dissolved organic matter in water, and therefore may act to increase iron availability. Furthermore, hypoxia and anoxia accompanying eutrophication may enhance iron availability in the water column due to iron release from sediments as the reducing intensity increases (NRC 1993a).





















