

CHAPTER 35
ECOLOGICAL RESTORATION
Reflections on a Half-Century of Experience at the University of Wisconsin-Madison Arboretum
WILLIAM R.JORDAN III
Editor, Restoration & Management Notes, The University of Wisconsin-Madison Arboretum, Madison, Wisconsin
So far, in this volume and in thinking and discussions about the conservation of biological diversity generally, the emphasis has been on preservation of what we already have. This makes sense. Preservation obviously has a critical role to play in the conservation of diversity. At the same time, however, it is clear that by itself preservation is not an adequate strategy for conserving diversity. At best, preservation can only hold on to what already exists. In a world of change, we need more than that. Ultimately, we need a way not only of saving what we have but also of putting the pieces back together when something has been altered, damaged, or even destroyed.
Consider, for example, that
-
vast areas of both land and water have already been profoundly altered by human activities ranging from agriculture to mining and construction and to various forms of pollution;
-
barring a catastrophe on the scale of nuclear war, human-caused alterations of natural and wilderness areas will continue indefinitely;
-
certain kinds of change—notably changes in climate—are beyond human control, and they in turn will inevitably change even those areas we have succeeded in preserving;
-
existing wilderness preserves are often inadequate in size or are suboptimal in shape or design; in many cases, their value as reservoirs of biodiversity could be dramatically increased by relatively modest increases in size, which could be achieved by active reconstruction of communities around their borders;
-
numerous species are already on the brink of extinction and their habitats have been reduced to a remnant or perhaps eliminated completely, so that their only hope for long-term survival is the re-creation of their habitat by human beings; and
-
the conservation of species ex situ will have little environmental value in the long run unless we find ways of providing habitat for them, often by creating it on disturbed sites.
All these considerations push us, unwillingly it seems at times, beyond a preoccupation with preservation, either in situ or ex situ, as the single strategy for the long-term conservation of diversity and toward a recognition of the importance of an active role for our species in reversing change or repairing damage. Unless, for example, we are prepared simply to write off disturbed lands as potential contributors to diversity, we are going to have to take seriously the problem of increasing diversity on these lands. Similarly, the inevitability of further change, including changes in climate, clearly implies that in order to preserve many communities over the long haul we are going to have to learn not only how to manage them but even how to move them around (Jordan et al., in press). And this brings us to the area of environmental healing, or ecological restoration, which is the subject of this section.
PIONEERING RESTORATION AT THE UNIVERSITY OF WISCONSIN-MADISON ARBORETUM
The starting point for this discussion will be the experience of the University of Wisconsin-Madison Arboretum, where research on restoration of ecological communities native to Wisconsin and the upper Midwest has been under way since 1934. Here, under the early leadership of Aldo Leopold and John Curtis, intensive restoration has been carried out on several hundred hectares of land, most of which had been seriously degraded by farming, logging, and sporadic development during the preceding century. Gradually, 40 hectares of tallgrass prairies have been restored on degraded pasture and plowland. A small xeric prairie has been created on an artificially constructed limestone outcropping. Red and white pine forests and boreal forests have been established on old pasture sites, and two types of maple forests are being developed by underplanting existing oak forests in which the understory had been depleted by grazing. The early stages of this effort were carried out by Civilian Conservation Corps crews working out of a camp on the site between 1935 and 1941. More recent work has been carried out by University of Wisconsin-Madison researchers and by the Arboretum staff. In general, the intensity of the restoration effort declined dramatically after 1941, though work continues, and indeed the need for ongoing restoration and management is one of the fundamental lessons that has emerged from the Arboretum’s experiences.
Overall, this has been a pioneering effort, and the Arboretum’s collection of restored and partially restored communities is now the oldest and most extensive of its kind anywhere in the world. Even more to the point, however, because of the Arboretum’s experience, it is possible to make a number of observations about
the nature of restoration, about its potential and its limitations as a strategy for conserving biological diversity, and about the environmental and social conditions under which it is likely to be feasible.
TECHNICAL, ECOLOGICAL FEASIBILITY
The first lesson that one might derive from this experience is that it is indeed possible, at least under certain circumstances, to re-create reasonably authentic replicas of some native ecological communities (Blewett, 1981). For example, the Arboretum’s two restored tallgrass prairies (Curtis and Greene prairies) now include areas believed to resemble quite closely prairies native to the area—at least with respect to floristic composition. In other words, most of the appropriate vascular plants are present; they are present in more or less the right proportions and associations; and the number of inappropriate plants—that is, exotics or plants not native to the tallgrass prairies of this area—is small.
On the other hand, there are large areas on these prairies where ecological or historic authenticity is relatively low and where various exotic species are abundant. Certain of these species have proved to be extremely difficult to remove or control. Some have turned out to be capable of invading the more or less intact prairie community, often at the expense of the native plants. As a result, it is now abundantly clear that the problem of dealing with exotics is an ongoing one and that the struggle will in many instances be unending. Undisturbed natural communities are also vulnerable to invasions by exotic species but, in general, probably less so than communities in the process of being restored. Without doubt, this has turned out to be a major problem facing restorationists.
In addition, the restoration program at the Arboretum has strongly emphasized revegetation, far less attention being paid to the reintroduction of animal species. This is frequently the case in restoration and land reclamation projects, since the assumption is often made that the appropriate animals will find their way into the community once it has developed to a certain point. But this does not always happen for complex reasons that include the size of the communities, their uneven quality, and their isolation from existing animal populations. An instance of this now appears to have occurred in the Arboretum’s restored southern maple forest, where ommission of an ant species that normally aids the dispersal of the seeds of certain herbaceous plants, such as bloodroot (Sanguinaria canadensis) and wild ginger (Asarum canadense), has resulted in the development of these species into peculiar, dense patches (Woods, 1984).
A related problem with restored communities generally is their small size, which can directly influence their ecological quality. Certain animals, for example, may not inhabit restored communities simply because these communities are often too small. This is a major reason why few if any restored prairies include buffalo, for example. At present, the prairie at Fermilab in suburban Chicago is probably the largest restored tallgrass prairie in existence (Nelson, 1987). Of course, this nearly 240-hectare prairie is still very small in comparison to the millions of hectares of prairie that existed in this area at the time of European settlement, and its ability to support populations of large native animals is at best problematic.
In addition to the more conspicuous defects in the composition of restored communities, there are numerous features, such as soil structure and chemistry, composition of soil flora, populations of less conspicuous animals (including insects), and various aspects of ecosystem function, that in many instances may not be authentic. Only rarely have these been studied in any detail.
On the positive side, however, the Arboretum’s restored communities have brought back into the landscape numerous plants and animals that had become rare or had even been eliminated locally. The entire project certainly represents an enormous contribution to what might be called the native diversity of the Madison area. The Arboretum’s restored tallgrass prairies, for example, are now among the largest prairies in Wisconsin, a state that had some 4.8 million hectares of prairie and savanna at the time of European settlement (Curtis, 1959). These prairies alone include more than 300 species of native plants. Some of them, including plants such as big bluestem grass (Andropogon gerardi), compass plant (Silphium laciniatum), and yellow coneflower (Ratibida pinnata), were extremely abundant in presettlement times, often dominating whole landscapes, but were virtually eliminated from the area by the time the restoration efforts at the Arboretum began. These now flourish in the restored communities, which also provide habitat for numerous rare species. Examples from the Arboretum’s collection include such rarities as the white-fringed orchid (Habenaria leucophaea), prairie parsley (Polytaenia nuttallii), smooth phlox (Phlox glaberrima), and wild quinine (Parthenium integrifolium)—all considered threatened or endangered, at least for the state. In general, the Arboretum itself probably has more biological diversity than any other area of comparable size in the state. This is due largely to the presence of the various restored communities.
In short, the Arboretum’s experience shows that restoration of some native communities may be technically feasible under certain conditions. The ecological quality of the resulting communities may vary, but under proper conditions, it may actually be quite high, and restored communities may often resemble the historic community chosen as a model quite closely, at least in floristic composition.
SOCIAL, ECONOMIC FACTORS
At the same time, the experience of the Arboretum raises a number of questions about the cost of such projects and the social, political, and economic feasibility of carrying them out. Thus, in considering the environmental significance of the Arboretum’s restoration efforts, one should keep in mind that these efforts have been carried out under conditions that clearly limit their relevance to other situations. These conditions include first of all the fact that the Arboretum itself is part of a major university and that its work has been performed primarily for scientific and academic reasons. In other words, from the very beginning, this effort has benefited from its academic setting and has been justified as an experiment or as a way of creating communities for research, rather than as a way of coping with environmental, much less economic, problems.
The second set of conditions that have contributed to the success of the Arboretum project were those directly related to the economic and ecological con-
ditions of the 1930s, notably the Great Depression and the Dustbowl. Together, these national calamities provided conditions (specifically, cheap land, free labor in the form of the Civilian Conservation Corps, and an incentive for ecological restoration) that proved crucial to the development of the Arboretum, but that have also reduced its value as a model for carrying out restoration projects in the real world outside academia. This point carries us outside the little world of the Arboretum to the larger world, where we have to ask a crucial question: What good is restoration? Is it likely to prove merely an academic pursuit or a pastime for environmentalists who happen to be interested in an unusual form of gardening? To just what extent and under what conditions can restoration be expected to contribute in a significant way to the conservation of diversity?
These questions have not yet been dealt with systematically, as far as I am aware. But it is important that we begin to take them seriously. In general, given the interrelatedness of everything on Earth and the inevitability of change, it would seem that an ineluctable logic argues for the importance of restoration as part of any comprehensive strategy for the conservation of biological diversity. Critical as it may be as part of such a strategy, preservation has serious defects. Basically, it is a one-way strategy that offers no way of responding to change or recouping losses. By itself, any such approach is clearly inadequate because in a changing world the quality of the environment is ultimately going to depend not simply upon the amount of land we manage to set aside and to preserve but upon the equilibrium we are able to maintain between the forces of destruction—or change—on the one hand and the forces of recovery on the other. All things considered, and despite its various limitations, it seems likely that restoration will ultimately play an important role in determining the position of this equilibrium.
This being the case, the questions raised above and a whole host of corollary questions and issues take on a great deal of urgency. Can we restore ecological systems? And if so, how authentic will the results be? Which communities lend themselves to restoration, and which are likely to prove more difficult—or even impossible—to restore? To what extent can we hope to re-create communities specifically designed to provide habitat for rare and endangered species? What needs to be known in order to restore a system effectively—and efficiently? What is the state of the art for the restoration of various communities, and what currently limits the effectiveness of restoration techniques for these systems? What sorts of research need to be undertaken in order to refine these techniques?
Beyond these questions about the technical feasibility of restoration, there are the various social, economic, and political questions: How much will it cost? Who will be expected to pay for it, and why? How will the costs compare with those of preservation or with the natural recovery of disturbed systems? What incentive will society have for restoring naturally diverse communities rather than for simply reclaiming land for some other purpose such as agriculture? In general, what incentives can be found for restoring communities—incentives that will ensure that restoration is actually accomplished and that its potential for contributing to biological diversity is effectively exploited?
In fact, there are a number of such incentives, including some traditional ones such as the creation of habitat for fish and game and the use of prairies as pasture
and rangeland. There are also important aesthetic incentives in park and wilderness management and in landscape architecture.
But restored communities may well have other economic values that have not yet been fully identified or widely recognized. Examples include development of wetlands to control water distribution and quality (Holtz, 1986), of prairies to rehabilitate soils degraded by agriculture (Miller and Jastrow, 1986), and of forests as part of a program of sustained-yield timber production (Ashby, 1987). Applications such as these at least suggest ways in which restoration might eventually prove critical as a way of reintegrating native communities into the economies of developed nations, in the process returning them to the landscape on a large scale.
These questions are addressed in the four chapters that follow. The first two are devoted mainly to defining the state of the art of ecological restoration for two community types. In the first of these, Chapter 36, Joy Zedler discusses restoration of a temperate zone community, the tidal wetland. In Chapter 37, Chris Uhl addresses the much-neglected subject of tropical forest restoration. The following two chapters turn to the more socially oriented aspects of the business of restoration. In Chapter 38, John Cairns looks at disturbed lands as opportunities for increasing local and regional biodiversity through restoration. In Chapter 39, John Todd presents some ideas about creating a social, political, and economic context for restoration projects.
REFERENCES
Ashby, C. 1987. Forests. Pp. 89–108 in M.E.Gilpin, W.R.Jordan III, and J.D.Aber, eds. Restoration Ecology: A Synthetic Approach to Ecological Research. Cambridge University Press, New York.
Blewett, T.J. 1981. An Ordination Study of Plant Species Ecology in the Arboretum Prairies. Ph.D. Thesis, University of Wisconsin-Madison. 354 pp.
Curtis, J.T. 1959. The Vegetation of Wisconsin. University of Wisconsin-Madison Press. 657 pp.
Holtz, S. 1986. Bringing back a beautiful landscape—wetland restoration on the Des Plaines River, Illinois. Restoration & Management Notes 4:56–61.
Jordan, W.R. III, R.L.Peters, and E.B.Allen. In press. Ecological restoration as a strategy for conserving biological diversity. Environ. Manage.
Miller, R.M., and J.D.Jastrow. 1986. Soil studies at Fermilab support agricultural role for restored prairies. Restoration & Management Notes 4:62–63.
Nelson, H.L. 1987. Prairie restoration in the Chicago area. Restoration & Management Notes 5(2).
Woods, B. 1984. Ants disperse seed of herb species in a restored maple forest. Restoration & Management Notes 2:18.
CHAPTER 36
RESTORING DIVERSITY IN SALT MARSHES
Can We Do It?
JOY B.ZEDLER
Professor of Biology, San Diego State University, San Diego, California
Along the U.S. coastline, development has reduced the area of coastal wetlands and endangered certain wetland-dependent species. Despite the threats to biodiversity, development of wetland habitat is still permitted by regulatory agencies if project damages can be mitigated by improving degraded wetlands or creating new wetlands from uplands. For example, the California Coastal Act allows one-fourth of a degraded wetland to be destroyed if the remaining three-fourths is enhanced. The expectation is that increased habitat quality will compensate for decreased quantity.
The concept sounds reasonable, but biodiversity is continuing to decline. Why? First, the process allows a loss of habitat acreage. Second, there is no assurance that wetland ecosystems can be manipulated to fulfill restoration promises. The magnitude of the problem is well illustrated by examples from southern California, where more than 75% of the coastal wetland acreage has already been destroyed, where wetland-dependent species have become endangered with extinction, and where coastal development pressures rank highest in the nation. This chapter reviews several restoration plans and implementation projects and suggests measures needed to reverse the trend of declining diversity.
RESTORATION PLANS
Several large development projects in southern California wetlands have recently been approved by the California Coastal Commission (see Figure 36–1). Three federally endangered species are affected by such projects: the California least tern (Sterna albifrons browni), light-footed clapper rail (Rallus longirostris levipes), and salt marsh bird’s beak (Cordylanthus maritimus spp. maritimus; see Figure 36–2).
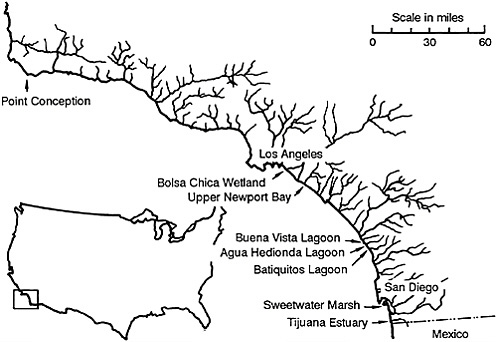
FIGURE 36–1 Sites of some coastal development projects in southern California. In all, there are 26 coastal wetlands between Point Conception and the Mexico-U.S. border.

FIGURE 36–2 The salt marsh bird’s beak grows near the upper wetland edge. As an annual plant, its seeds germinate after winter rainfall to maintain the population; as a hemiparasite, its seedlings grow roots that can attach to those of other plants, thereby increasing its supplies of water and nutrients. Photo by J.Zedler.
There would also be an impact on the Belding’s Savannah sparrow (Passerculus sandwichensis beldingi), which is listed as endangered by the state, and on several plant species of regional concern (Ferren, 1985).
Projects That Show Losses in Wetland Area
At Bolsa Chica Wetland, more than 1,200 acres (480 hectares) of lagoonal wetland will be reduced to 951 acres (366 hectares) of restored wetland (California State Coastal Conservancy, 1984). Mitigation plans are not final, but the draft concept includes cutting an ocean inlet to serve a new marina. Inland from the marina are sites for restored wetlands with controlled tidal flushing. Uplands designated as “environmentally sensitive habitat areas” that lie within the lowland area and that will be destroyed during development are to be relocated adjacent to the restored wetland in a bluff-edge (linear) park. The draft concept plan accommodates development, but does not ensure maintenance of biodiversity. The restoration activities are based on the assumption that habitat values can be created and moved about at will.
In Los Angeles Harbor, about 400 acres (160 hectares) of shallow water fisheries habitat will be filled to construct new port facilities. At this project site, there is no habitat available to be restored—all the wetlands have been filled or dredged. Thus, off-site mitigation has been approved. Batiquitos Lagoon, more than 80 miles (130 kilometers) south of Los Angeles, will be dredged to create deep-water habitat and increase tidal flushing. According to plans (California State Coastal Conservancy, 1986), the net loss of aquatic habitat in Los Angeles will be mitigated by altering (not increasing) habitat elsewhere. The dredging of Batiquitos Lagoon will remove sediments and, at least temporarily, solve the occasional problems of algal blooms (odors and fish kills after sewage spills). However, maximizing tidal flushing at Batiquitos Lagoon (to replace fisheries habitat in Los Angeles Harbor) will destroy existing salt marsh habitat (Figure 36–3) and reduce the area of shallow water and mudflat habitat. The mitigation plan contains two strikes against biodiversity—the loss of area and the loss of existing functional wetland types.
At Aqua Hedionda Lagoon, about 14 acres (5.6 hectares) of wetland were filled to build a four-lane road. The mitigation plan (U.S. Army Corps of Engineers, 1985) promised to enhance diversity and increase the functional capacity of the lagoon. Brackish-water ponds were planned for a wetland transitional area (itself a rare habitat type); a 2-acre (0.8-hectare) dredge spoil island was to be built for bird nesting; and a 7-acre (2.8-hectare) debris basin was proposed within a riparian area to reduce sedimentation into the lagoon. Flaws in the plan became clear when construction of the brackish ponds began. Pits were dug to a depth of 6 feet (1.8 meters) without encountering groundwater. Areas that were modified included transition habitat, pickleweed marsh (Figure 36–4), brackish marsh, and riparian habitat. The wetland lost both acreage and habitat quality.
All these projects show a net loss in wetland habitat area. Proponents argue that the lost areas are already degraded. However, they could be enhanced to maintain biodiversity. The fact that four wetland-dependent species have become endangered in Southern California while coastal wetlands have shrunk by 75% indicates a cause-effect relationship. There is some minimum area required to
support regional biodiversity and a limit to the number of species that can be packed into individual wetlands. Populations are dynamic; some migrate and use several wetlands, whereas others experience local declines and must reinvade from another refuge. The need to maximize area available for wetland species is indicated by population declines that have followed human disturbance and environmental catastrophes.
Several species may be lost simultaneously if a wetland experiences multiple catastrophes. At Tijuana Estuary, the combination of destabilized dune sands (following long-term trampling), the winter storms of 1983, dune washovers, and channel sedimentation led to closure of the ocean inlet in April 1984. The drought of 1984 coincided with an 8-month nontidal period. The population of endangered light-footed clapper rails dropped from about 40 pairs to 0 and did not fully recover after tidal flushing was restored (16 pairs were present in 1987). In addition, there were major declines of three salt-marsh plant species—cordgrass (Spartina foliosa), annual pickleweed (Salicornia bigelovii), and sea-blite (Suaeda esteroa)—and none has recovered to pre-1983 levels. This salt marsh has shifted from the region’s most-diverse to a species-poor wetland (Zedler and Nordby, 1986).

FIGURE 36–3 Salt flats may appear to have low habitat value, but many unusual insects, some of them threatened with extinction, are found only in these open areas. In the winter, runoff and high tides inundate the areas, and they become highly productive ecosystems. What appears to be barren in summer is heavily used by shorebirds and dabbling ducks in winter, as migrants visit the flats and feed on the abundant insects and algae. Photo by J.Zedler.

FIGURE 36–4 Pickleweed marsh may seem monotonous, but close inspection will reveal a variety of insects, invertebrates, and dozens of species of microscopic algae. Individuals and trails of the California horn snail (Cerithidea californica) are visible in the tidal pool. Photo by J.Zedler.
Maintenance of the region’s resources through years of wet and dry periods, with and without closure to tidal flushing, requires that each habitat type be maintained at several different wetlands so there will be refuges during periods of environmental extremes. Further losses in habitat area cannot be justified.
Projects That Replace Functional Wetland Habitat with Modified Wetland Habitat
Some restoration projects retain acreage but exchange one type of habitat for another. In these cases, functional wetland habitats may be destroyed in order to create some other habitat type. Following are some examples.
The City of Chula Vista’s Bayfront Development Plan calls for several developments near and in the last major salt marsh within San Diego Bay (90% of the Bay’s wetland has already been developed). The plan includes a multistory hotel and a nature center to be built on an island that is surrounded by Sweetwater Marsh and San Diego Bay. Residential and commercial buildings would surround the marsh. To provide access, three roads are to be built over the wetland. The plan will also require modification of the wetland for the construction of debris basins. Wetland restoration is planned to mitigate impacts. The U.S. Fish and Wildlife Service has concluded that portions of the project jeopardize the following
endangered species: California least tern, light-footed clapper rail, California brown pelican (Pelicanus occidentalis), and salt marsh bird’s beak.
At Los Cerritos Wetland near Los Angeles, a complicated plan (California State Coastal Conservancy, 1982) proposes development of some wetland in exchange for an equal area of wetland creation. In all, 129 acres (51.6 hectares) of wetland will be retained. Some dikes that now prevent tidal flushing will be breached; some new areas will be graded to allow tidal flow. The restored wetlands will be divided into four segments and surrounded by high-density urban uses. Buffers between the wetland and development are as narrow as 25 feet (7.5 meters) for much of the project. A main concern is that existing wetland habitat will be lost and that the artificially created replacements cannot guarantee maintenance of biodiversity.
At Upper Newport Bay, a sediment control plan within a California State Ecological Reserve has received wide political support, in part because dredging in the upper bay reduces sedimentation in the lower bay’s marina. Sedimentation in the upper bay is a long-term threat to the marsh habitat, but sudden changes in hydrology may have a negative impact on the habitat of endangered species. Upper Newport Bay has the highest density of light-footed clapper rails in the state of California and some of the region’s most robust cordgrass vegetation. Shallow-water and transitional habitats are being traded for deeper channels, and the value of the new habitats to biodiversity is uncertain.
At Buena Vista Lagoon, sediment control measures were also taken. Shallow-water areas were deepened, and dredge spoils were placed alongside them in the wetland. The dredge spoil islands became hypersaline, bricklike substrates that have not developed the desired vegetation or significantly improved the status of the least tern population.
CONCLUSIONS CONCERNING RESTORATION PLANNING
Several observations on the status of restoration plans can be made:
-
Many large projects result in a loss of wetland acreage.
-
Mitigation measures for lost habitat often involve changing one type of wetland habitat into another, rather than creating wetland from upland habitat.
-
Proposed projects are planned and reviewed individually rather than with a regional perspective. While cumulative impacts may be considered, there is no regional coordination to set priorities and guide decision making. There is no way to ensure that the wetlands with the greatest potential for maintaining clapper rails will be managed for clapper rails.
-
There is no single source of information on restoration projects, no center that keeps records on changes in biodiversity to ensure that resource agencies are aware of changes in individual wetlands, no comprehensive monitoring programs to assess changes in biodiversity (although some endangered species are censused annually), and no mechanism to require suitable and comparable methods for the few monitoring programs that have been planned.
IMPLEMENTATION OF RESTORATION PROJECTS
To enhance, restore, or create wetland habitat requires manipulation of the physical environment (especially the topography and the degree and timing of fresh- and seawater influence) as well as the biota (e.g., by introducing target species and eliminating undesirable ones). Research in this area is just beginning; most of the work has been done on a trial-and-error basis, and the evaluation criteria are not yet standard.
Assessing Success
Restoration success must be measured in time scales that relate to the species being managed and to the periodicity of extreme environmental conditions characteristic of the region. Successful creation of clapper rail habitat cannot be measured by censusing mortality of cordgrass a few weeks after transplantation. Rather, such projects need to be followed at least until clapper rails establish breeding populations. Measures of restoration success must be done within spatial scales that relate to whole ecosystems. The degree to which breaching a dike and restoring tidal flushing can enhance a lagoon must be measured beyond channel biota and water quality, because there will also be substantial impacts on intertidal marshes. Likewise, dredging to improve fish diversity cannot be considered successful if endangered birds become extinct in the process. In short, restoration success must be measured at the ecosystem level and with long-term evaluation. To date, this has not been done.
Summary of Trials
All the projects described above incorporate some element of habitat creation or restoration, for which there are no guaranteed benefits to threatened species. Many projects have been designed to restore wetlands and mitigate losses, but in no case have ecosystem functions been duplicated, nor have endangered species been rescued from the threat of extinction. Projects to reduce sedimentation have as one goal the creation of fish and benthic invertebrate habitat (sometimes to provide food for the California least tern). Marsh restoration projects have focused on vegetation used by target bird species (cordgrass for clapper rails, pickleweed for Belding’s Savannah sparrows).
While some wetland plant species can be transplanted successfully and others will invade voluntarily given suitable conditions (Zedler, 1984), there are only a few cases where the marsh ecosystem has been monitored for several years (Broome et al., 1986; Homziak et al., 1982) and no example of a threatened species that has been increased as desired. Attempts to restore wetlands in southern California have generally failed to attract target species. In a few cases, the California least tern has nested on dredge spoil islands—but not always where its use was planned. One briefly successful site was an 80-acre (132-hectare) island in south San Diego Bay, which was planned for salt marsh and fish habitat.
Conclusions Concerning Implementation
Four observations on the status of wetland habitat restoration in southern California can be made.
-
Selected plant species (e.g., cordgrass) can be transplanted successfully.
-
No plant or animal populations have been taken off the endangered list as a result of restoration projects.
-
Wetland restoration assessments have not been made for entire ecosystems but have been limited to one or a few target species.
-
No studies have been conducted to determine the minimum wetland area or configuration of multiple wetlands required to maintain regional biodiversity.
Therefore, it is premature to conclude that an artificial tidal wetland can develop and replace the functions of a natural one. Furthermore, there is no evidence that restoration of degraded wetland habitat can compensate for lost habitat area.
PROSPECTS FOR THE FUTURE
To restore biodiversity in the nation’s coastal wetlands, we must understand the factors controlling these ecosystems and develop the ability to modify them to meet desired management goals. We must make substantial advances in ecotechnology—the scientifically sound manipulation of ecosystems to maintain natural diversity and achieve specific management objectives. The field is relatively new in ecology. Only one journal, Restoration and Management Notes, and a few books focus on the topic. Although most of the work in this area concerns disturbed ecosystems, all ecosystems need some management to maintain their natural hydrology as well as air and water quality.
Ecosystems of greatest concern tend to be those whose areas have been reduced and whose species are threatened with extinction. Rare species are difficult to study, because the conditions that allowed them to thrive no longer exist. Manipulative experimentation, required to establish cause-effect relationships, cannot always be done without threatening the endangered populations even further. Bringing animals or plants (even seeds) into the laboratory may reduce field populations to levels that jeopardize population recruitment. Thus, maintenance of biodiversity must be based on an understanding of the factors that control the ecosystems in which rare species persist—the type of long-term, ecosystem-level research now funded by the National Science Foundation, the National Oceanic and Atmospheric Administration through its Sea Grant Program, and other agencies. A new research emphasis could allow major advances to be made in wetland ecotechnology. I recommend manipulative experimentation, first in replicate mesocosms (medium-size artificial ecosystems), followed by experimental restoration at the ecosystem level. This approach was adopted by the U.S. Environmental Protection Agency in their research plan for the nation’s wetlands (Zedler and Kentula, 1985).
Ecosystem-level experiments have not been incorporated into wetland restoration projects. The contention that artificial or restored wetlands can maintain biodiv-
ersity must be tested. Every restoration project can include experimentation in its design, e.g., to provide different tidal flows; to test different hydroperiods, salinities, and nutrient inputs; to use different transplantation regimes; or to vary the width of buffer zones, with treatments appropriately replicated. Detailed, long-term evaluation of the experiments will document success or failure to maintain natural diversity. In either event, we will learn whether it can be done and why it succeeds or fails. The present practice of poorly planned, unreplicated, undocumented trials leads mainly to errors whose causes cannot be identified. Only as our understanding of factors controlling wetland ecosystems improves can we ensure the restoration and maintenance of biodiversity.
ACKNOWLEDGMENTS
Research on wetland restoration was funded in part by NOAA, National Sea Grant College Program, Department of Commerce, under grant number NA80AA-D-00120, project number R/CZ-51, through the California Sea Grant College Program, and in part by the California State Resources Agency.
REFERENCES
Broome, S.W., E.D.Seneca, and W.W.Woodhouse, Jr. 1986. Long-term growth and development of transplants of the salt-marsh grass Spartina alterniflora. Estuaries 9:63–74.
California State Coastal Conservancy. 1982. Los Cerritos Wetlands: Alternative Wetland Restoration Plans Report. State of California—Resources Agency, State Coastal Conservancy, Oakland. 49 pp. +appendixes.
California State Coastal Conservancy. 1984. Staff Presentation for Public Hearing: Bolsa Chica Habitat Conservation Plan. State of California—Resources Agency, State Coastal Conservancy, Oakland. 31 pp. +exhibits.
California State Coastal Conservancy. 1986. Batiquitos Lagoon Enhancement Plan Draft. State of California—Resources Agency, State Coastal Conservancy, Oakland. 183 pp. +appendixes.
Ferren, W., Jr. 1985. Carpinteria Salt Marsh. Publication #4, Herbarium, University of California, Santa Barbara, Calif. 300 pp.
Homziak, J., M.S.Fonseca, and W.J.Kenworthy. 1982. Macrobenthic community structure in a transplanted eelgrass (Zostera marina) meadow. Mar. Ecol. Prog. Ser. 9:211–221.
U.S. Army Corps of Engineers. 1985. Public Notice of Application for Permit, Application No. 85–137-AA. U.S. Army Corps of Engineers, Los Angeles District. 13 pp.
Zedler, J.B. 1984. Salt Marsh Restoration: A Guidebook for Southern California. Report No. T-CSGCP-009. California Sea Grant, La Jolla, Calif. 46 pp.
Zedler, J.B. In press. Salt marsh restoration: Lessons from California. In J.Cairns, ed. Rehabilitating Damaged Ecosystems. CRC Press, Boca Raton, Fla.
Zedler, J.B., and M.Kentula. 1985. Wetland Research Plan. Corvallis Environmental Laboratory, U.S. Environmental Protection Agency, Corvallis, Oreg. 118 pp.
Zedler, J.B., and C.S.Nordby. 1986. The Ecology of Tijuana Estuary: An Estuarine Profile. U.S. Fish Wildl. Serv. Biol. Rep. 85(7.5). 104 pp.
CHAPTER 37
RESTORATION OF DEGRADED LANDS IN THE AMAZON BASIN
CHRISTOPHER UHL
Assistant Professor, Department of Biology, The Pennsylvania State University, University Park, Pennsylvania
The deforestation and degradation of the Amazon ecosystem have important global implications. Conservation groups have responded by working to establish national parks and biological reserves in Amazonia. Meanwhile, scientists have been attempting to document the types and frequency of both natural and human-induced disturbances in Amazonia and the capacity of its ecosystems to recover from disturbances. Central to this ecological research is the question, How much can Amazon forests be abused and still recover? As discussed herein, forest communities in the Amazon reform naturally following natural disturbances; however, forest regeneration is slow and uncertain following some of the larger-scale, more-intensive human-induced disturbances that are becoming increasingly common in the area. In such cases, humans may have to change hats and become restorers rather than exploiters.
NATURAL DISTURBANCES IN AMAZONIA
There is a tendency to believe that the Amazon rain forest has existed in a pristine, cathedral-like state for tens of thousands or even millions of years and that this forest is just now being disturbed for the first time because of the development activities of modern human beings. There is ample reason to believe, however, that disturbance has always been a common feature of Amazon forest ecology.
Winds causing forest treefalls and forest fires have probably been the most important natural disturbances during Amazon forest history. Several studies have shown that treefall disturbances are common in Amazon forests. In fact, it would
not be out of the ordinary to see or hear a tree crashing to the ground on an afternoon walk through the forest. From 4 to 6% of any Amazon forest will be studded with canopy openings (light gaps) formed by treefalls. Recently formed light gaps have a ground layer of tree and vine seedlings. As a light gap patch ages, it enters the building phase during which it develops into a densely stocked patch of pole-sized trees. The patch reaches a mature phase when it contains a mix of large trees, poles, and seedlings. With a practiced eye, it is possible to walk through the rain forest and detect these light gaps and identify the building and mature phase patches—testimonies to past disturbances and to the dynamic nature of these ecosystems. Importantly, this type of small-scale disturbance is more of a subsidy than a stress to the plant community, because the resources critical for growth—light, water, and nutrients—are more readily available in treefall gaps than in the undisturbed forest understory.
Another natural disturbance that has no doubt been an important part of past Amazon disturbances is fire. For example, in the Upper Rio Negro region of Amazonia, charcoal is widespread and abundant in the soil. Radiocarbon dating of soil charcoal samples (Sanford et al., 1985) indicates that numerous fires have occurred during the past 6,000 years in this area. The radiocarbon dates correspond well with what are believed to have been dry episodes during recent Amazon history. The presence of abundant charcoal in Rio Negro soils is not an anomoly. Amazon researchers from EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária) and INPA (Instituto Nacional de Pesquisas da Amazonia) in Brazil concur that charcoal is common in the soils of the central and eastern Amazon. Indeed, it appears to be much more difficult to find sites that do not have charcoal than to find sites that do.
Given the prevalence of fire throughout the history of the Amazon forest, how does Amazon vegetation respond to fire disturbance? Our studies of forest succession following forest cutting and burning disturbances at San Carlos de Rio Negro, Venezuela (Uhl and Jordan, 1984) provide an indication of regrowth potential following fire. We found that forest reforms quickly on burned sites. This is because many Amazonian-tree species have the ability to sprout after damage, and although fires do kill many stems outright, a pool of individuals survives burning and quickly sprouts. In addition, Amazon forests have a rich seed bank (from 500 to 1,000 seeds of successional woody species per square meter), and a portion of these seeds survive burning, germinate, and become established. The ability of some rain forest species to survive fire disturbances, e.g., when seeds are buried and thus protected and later sprout, may be the result of natural selection (i.e., fire may have been a selecting agent for these characteristics).
DISTURBANCE BY HUMANS IN AMAZONIA
Human-induced disturbances, such as slash-and-burn agriculture and the conversion of forest to pasture, are generally more intense than the natural disturbances just discussed. Nature’s disturbances are over in an instant (a treefall, a wildfire), whereas humans prolong the disturbance period in their efforts to wrest some benefit from the land.
In slash-and-burn agriculture, the felled forest land is used to grow subsistence crops such as cassava, beans, and fruits. When these farm plots are abandoned and if they are not disturbed, they eventually return to forest, but the process takes a long time. The frequent weeding typical of slash-and-burn agriculture means that succession begins and is curtailed many times before sites are finally abandoned. These repeated weedings cause striking shifts in the composition of the regrowth. Most notably, the number of woody pioneer species declines after each weeding, whereas the density of forbs and grasses increases. Forbs and grasses are able to germinate, flower, and set seed in the interval between weedings and therefore can build up high plant densities and large seed banks. In contrast, the woody pioneer species that are established from seeds surviving the burn are weeded from the site before they have had time to produce more seeds. Because the agricultural practices of cutting, burning, and weeding largely eliminate mechanisms of on-site regeneration, the only way for forest species to establish on farms is through seed dispersal. This dependence on seed dispersal slows succession, because many of the animal species that routinely disperse seeds of forest species do not frequent large forest openings. Approximately 200 years are required for abandoned farm plots to reach mature forest proportions (Saldarriaga, 1985). That forest regeneration proceeds at all on abandoned farm sites is the result of two important factors:
-
Slash-and-burn clearings are relatively small (usually =1 hectare). Hence, seed dispersal distances are short.
-
The period of farm use is relatively short (±3 years). Hence, safe germination sites, e.g., slash piles (the shaded, moist zones along the sides of decaying logs), and soil nutrients are still available to ensure the establishment of some forest trees.
Forest regeneration on intensively used cattle pasture in Amazonia is more problematic, because the disturbances there can have the following characteristics:
-
They are frequently prolonged and can therefore result in a highly compacted soil and allow time for the decomposition of slash, which normally provides important microhabitats for the establishment of seedlings.
-
They can involve repeated burning and weeding, which eventually destroy all means of on-site woody regeneration and further homogenize the site, eliminating establishment microhabitats.
-
They can be very large and, hence, make seed dispersal from distant forests extremely unlikely.
A ROLE FOR RESTORATION ECOLOGY
Regeneration to forest occurs naturally in Amazonia following natural disturbances and small-scale human-induced disturbances, but where disturbances are severe, human intervention may be necessary to ensure reforestation. Restoration efforts are most needed on abandoned mine sites and highly degraded pastures.
The most extensive work on abandoned mines is being done by Oliver Henry Knowles, a resident ecologist for the Vale do Rio Dulce Bauxite Mining Company
located near Porto Trombetas in Pará, Brazil. This company mines about 60 hectares per year. The topsoil (about 20 centimeters thick) is bulldozed into stockpiles during mining. After bauxite is removed, the mined trench is refilled with overburden and the stockpiled topsoil is respread on the site. The soil is then sliced at 1-meter intervals to a 90-centimeter depth to increase aeration and facilitate root penetration. Finally, the area is planted with nursery-raised seedlings of native forest species. Although growth is slow (most species attain about 20 centimeters in height per year), survival is good. The principal impediments to restoration are insect pests (e.g., leaf-cutter ants, grasshoppers, and caterpillars) and soil nutrient deficiencies.
Restoration of highly degraded Amazon pasturelands is also receiving attention through a study I am conducting in collaboration with Robert Buschbacher, Daniel Nepstad, and Adilson Serrao near Paragominas in northern Pará, Brazil. The vegetation of our study area is lowland evergreen rain forest. Rainfall is approximately 1,700 millimeters per year, and there is a distinct dry season from June through November. As a first step in determining how to rehabilitate degraded pastures, we are studying how natural forest and degraded pasture environments differ with respect to microclimate, soil water and nutrient availability, and physical properties of soil. This research sets the stage for our central research goal: to determine how biological and physical forces act to retard the establishment of rain forest tree species in highly degraded Amazon pastures. Our data suggest that forest trees have difficulty establishing in degraded pastures because of three factors:
-
few seeds of forest trees are being dispersed into pasture environments;
-
most seeds that do arrive are killed by seed predators; and
-
the few seeds that do manage to germinate eventually die because of harsh environmental conditions.
Seed Dispersal
The fact that few seeds of forest trees are being dispersed into pastureland is a fundamental impediment to regeneration of these areas. Our floristic survey has shown that less than 15% of the forest species have fruits with adaptations for long-distance autodispersal (e.g., for dispersal by wind). The majority of tree fruits are fleshy and appear to be dispersed by birds, bats, and both arboreal- and ground-dwelling mammals. Hence, if seeds of forest tree species are to arrive at pasture environments, they will usually have to be carried there by animals. With this in mind, we are censusing and trapping (i.e., mist netting) birds and bats in forest edge, forest second-growth, and open-degraded pasture environments. In the case of birds, we have identified more than 150 species in our area, but only a subset of them are frugivores, and of the frugivores, fewer than 10 will move out into large openings. We are now studying the movement patterns of the few species that we have identified as potential seed vectors. Our goal is to critically evaluate the role that these species play in seed movement and to begin to elucidate the determinants of movement for these species.
Postdispersal Seed Predation
Seed predators are also important roadblocks to the establishment of forest trees in degraded pastures. To study the fate of seeds that are artificially dispersed to degraded pastures, we placed seeds in groups of 8 that are 10 centimeters apart at 30 widely spaced stations. By repeatedly revisiting these stations, we are able to determine disappearance rates and, if we are lucky, the animal species that are removing the seeds. In studies on the tree Inga sp., leaf-cutter ants (Atta sexdens) were carrying off the seeds to their subterranean burrows within minutes of placement. Large seeds of other species were slowly consumed in situ, presumably by rodents, judging from teeth marks on partially consumed seeds.
In recently completed studies, we placed seeds of forest tree species simultaneously in pastures, in closed forest, and in canopy openings caused by treefalls. For all six species tested, seed removal was much more rapid in the degraded pasture than in the forest environments. All these findings suggest that seed predation is an important impediment to forest regeneration in degraded pastures.
Harsh Environmental Conditions in Pastures
Harsh environmental conditions present a final impediment to the establishment of forest trees in degraded pastures. The physical conditions of the pasture and forest contrast sharply and may significantly impede seedling survival and growth in the pasture. During the 6- to 7-month wet season usually ending in June or July, daily rains maintain pasture and forest soils near saturation. In the ensuing dry season, rain falls episodically, usually totalling less than 100 millimeters per month, and soil moisture levels decline between rain events. In the pastures, tensions of 2.0 megapascals are reached in soil 15 centimeters below the surface, whereas in forests, moisture is rarely held at tensions greater than 1.5 megapascals. This means that pasture plants rooted in the 15 centimeters of soil are subjected to intense drought (i.e., permanent wilting point conditions), whereas water availability in the surface soil of forests is adequate.
In short, the seasonal changes in moisture and energy conditions near the ground surface of degraded pastures are dramatic compared to conditions in the forest. During the wet season, the availability of soil moisture and radiation load in degraded pastures is very similar to conditions beneath closed-canopy forest. During drought episodes, soil and air moisture deficits and air temperatures in pastures greatly exceed those of the forest.
A summary of the factors that limit the establishment of forest trees in degraded pastures is provided in Figure 37–1.
RESTORATION ECOLOGY STRATEGIES
We have begun a series of field experiments to try to overcome these impediments to forest recovery on degraded lands. The seed dispersal limitation is straightforward and can be overcome, since humans can act as the seed dispersal agent. Alternatively, instead of individual seeds, humans could disperse onto degraded lands packets of forest surface soil with its component seed bank. This might allow whole
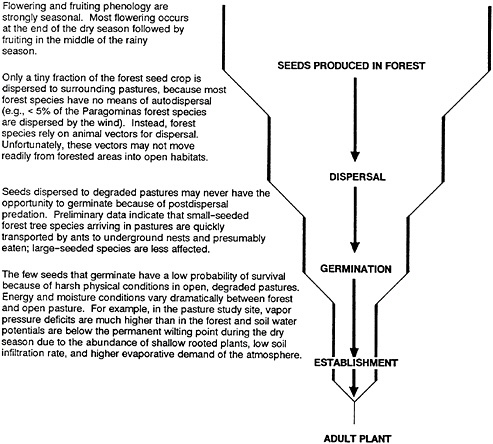
FIGURE 37–1 A summary of the factors that limit the establishment of forest trees in degraded pastures in the Eastern Amazon, Paragominas, Pará.
community patches to develop in unison. Another approach is to consider new types of seed vectors. Daniel Janzen (Chapter 14 of this volume) is studying the use of cattle as forest seed dispersal agents in old pastures in Costa Rica. In a like manner, there may be certain bird or bat species that could provide valuable seed dispersal services if introduced into a landscape mosaic of forest and abandoned pastures.
Seed predation in highly degraded sites will be difficult to overcome. Conventional approaches are to bury introduced seeds, saturate the environment with seeds, soak introduced seeds in repellent chemicals, and introduce predators or parasites to reduce the population of the seed predator. Of course, this need to protect the seeds can be bypassed entirely by introducing seedlings, which, however, can be attacked by seed predators, particularly leaf-cutter ants. Given the cost involved in these approaches, it seems wiser to choose species for introduction on the basis of their resistance to predators. Hence, we need to learn enough about seeds and their predators to be able to predict what kinds of seeds are least vulnerable to
predation. Our results to date show that tough-coated, heavy (e.g. ≥10g) seeds are the most predator resistant.
Harsh environmental conditions, particularly those related to drought and excessive radiation, are also not easily overcome. In field experiments, we have found that many forest species require shade (in the form of a shade cloth) and irrigation treatments (at least during their first dry season) to ensure establishment. Moreover, it appears that big-seeded forest species have a higher probability of establishment than small-seeded pioneer species in degraded pasture environments.
The critical first step in forest restoration is to foster the establishment of some predator-resistant, stress-tolerant tree species. Deep-rooted species that are able to extract water and nutrients from the lower soil horizons would be particularly appropriate. (In our forest studies we have found that root systems of some tree species extend beyond 10 meters in depth!) Once a few scattered trees are present in these pastures, the subsequent phases of forest development may occur naturally. This is because isolated trees attract bird and bat seed vectors by serving as perch and roosting sites and they provide favorable microsites for establishment (i.e., the shaded, moist conditions created by trees facilitate establishment of dispersed seeds).
Our studies should ultimately prove useful by providing guidelines for the reforestation of degraded pastures, should that become a land-use goal at some future time. For example, by coupling precise descriptions of pasture environments (in terms of barriers to the dispersal, germination, and establishment of tree seeds) with a knowledge of tree morphology and physiology, tree species can be selected for introduction into degraded pastures based on characteristics such as degree of tap rooting, ability to penetrate compacted soil horizons, drought tolerance, and competitive ability.
REFERENCES
Saldarriaga, J.G. 1985. Forest Succession in the Upper Rio Negro of Colombia and Venezuela. Ph.D. Thesis, University of Tennessee, Oak Ridge.
Sanford, R.L., Jr., J.Saldarriaga, K.Clark, C.Uhl, and R.Herrera. 1985. Amazon rain-forest fires. Science 227:53–55.
Uhl, C., and C.F.Jordan. 1984. Succession and nutrient dynamics following forest cutting and burning in Amazonia. Ecology 65:1476–1490.
CHAPTER 38
INCREASING DIVERSITY BY RESTORING DAMAGED ECOSYSTEMS
JOHN CAIRNS, JR.
University Center for Environmental Studies, and Department of Biology, Virginia Polytechnic Institute and State University, Blacksburg, Virginia
The large and growing number of human beings on Earth together with rapid advances in technological development ensure extensive damage to ecosystems. For example, Vitousek et al. (1986) have shown that nearly 40% of potential net primary terrestrial productivity is used directly, co-opted, or foregone because of human activities. Mabbutt (1984) noted that 35 million square kilometers of land is at least moderately desertified. Surface and subsurface mining, hazardous substances, clear-cutting, and a variety of other human-induced stresses have taken their toll on ecosystems. Damaged ecosystems in all parts of the world, even where the human population is sparse, comprise abundant material for experimentation.
Ideally, all ecosystems should be restored to their original condition; however, this is often difficult or impossible for the following reasons: detailed ecological information about the original condition is not available, techniques for recolonizing the damaged ecosystem with original species are not adequate, and there is no satisfactory source of organisms for recolonizing to the original condition. Under these circumstances, one might reasonably consider alternative ecosystems that would be ecologically superior to the damaged condition but often markedly different ecologically from the original system (Cairns, 1980, 1985). In some cases, wetlands lost elsewhere to development can be established on disturbed sites where they did not formerly exist. In others, additional habitat can be provided for rare, threatened, or endangered species (see, for example, Bruns, in press). A plan for developing these capabilities has been described by Cairns (1987 and in press).

Devastation following logging near Pellston, Michigan, around the turn of the century. Photo courtesy of Gary R.Williams of Glen Ellyn, Illinois.
EVIDENCE FOR CAUTIOUS OPTIMISM
In view of the present modest capability to predict ecosystem development following remedial measures to repair anthropogenic stress, it may seem arrogant to propose rehabilitation to increase diversity. However, it is abundantly clear that we must both arrest and repair widespread ecological damage. By doing this, we should learn much about both the structure and function of ecosystems if only because there are, unfortunately, all too many experimental sites that have been seriously damaged. One might object that the existence of an ecosystem restoration capability would provide an excuse for further damage. This seems quite unlikely once the difficulties and costs of restoring damaged ecosystems are more thoroughly documented and widely understood. This will be immediately apparent if the restoration costs are the responsibility of the organization or persons causing the damage.

This 1978 photo clearly shows the restoration of a site where comparable damage occurred near that shown on the opposite page, now part of University of Michigan Biological Station at South Fishtail Bay. Photo courtesy of Gary R.Williams of Glen Ellyn, Illinois.
Many biologists are unaware that some of the world’s leading biological field stations are located on sites that suffered severe ecological damage less than 100 years ago. For example, the University of Michigan Biological Station near Pellston occupies a site that was once extensively logged, followed by extensive burning of slash (e.g., unmarketable limbs). It is now a Biosphere Preserve under the Man and the Biosphere Program. Photographs of both the destruction and recovery of this site have been published (see, for example, Figures 38–1 and 38–2) (Cairns, 1980). Another station, the Rocky Mountain Biological Laboratory (RMBL), is located at Gothic, Colorado, a former mining town where a number of ecologically damaging mining activities, such as smelting, occurred (Vandenbusche, 1981). In fact, the Laboratory’s 205 acres (82 hectares) of land at Gothic already function as a small nature preserve (Brussard, 1982). A number of highly regarded ecologists
regularly return each summer to these field stations to conduct research that is published in respected professional journals.
Thus, the use of a formerly damaged ecosystem to preserve biological diversity is already an established practice. In both field stations just mentioned, restoration management practices appear to have been preventing further damage and letting natural processes occur. While I was writing this chapter, Boyce A.Drummond, Director of the Pikes Peak Research Station, and Mark G.Noble, Field Director of the Mountain Research Station, Institute of Arctic and Alpine Research, University of Colorado, visited RMBL. It occurred to me to learn whether the field stations where Drummond and Noble worked were situated on formerly damaged ecosystems.
The Mountain Research Station area was logged and burned around the turn of the century, and although there were no restoration management practices other than protecting the ecosystem from further damage, the lodgepole pine (Pinus contorta) once characteristic of the area has recolonized. It is an interesting western example of the same logging and burning practices that seriously affected the site of Michigan’s Biological Station. The Pikes Peak Research Station is located on a former ranching site, and some ranching is still going on nearby. On the station site itself, grazing was stopped in spring 1985. Although neither station has become restored to its original condition, both have witnessed the reestablishment of species characteristic of the original site, such as ponderosa pine (Pinus ponderosa), mountain muhly grass (Muhlenbergia), Arizona fesque (Festuca montana arizonica), and purple milk vetch (Astragalus dasyglottis).
The Pikes Peak Research Station provides an excellent example of another opportunity, namely, the use of formerly damaged ecosystems to increase regional diversity of both habitat and species while affording research opportunities for ecologists. Surely, utilization of present ecological knowledge and theory would produce better results sooner. If not, we are in deep trouble!
On July 31, 1986, a letter was sent to 61 directors and other persons affiliated with field stations from a list of more than 200 names. To summarize a detailed letter, there were four major requests for information:
-
Is the field station located on a formerly damaged ecosystem?
-
If so, were any management practices used to enhance the recovery process?
-
Is the facility formally designated as a nature preserve or a biosphere reserve?
-
Is the field station an ecological oasis surrounded by ecosystems that are substantially altered or damaged?
As of September 18, 1986, there were nine responses to this request. One response covered 26 sites in the Natural Reserve System managed by the University of California (information from C.Ronald Carroll). Of these, 11 were nature reserves or preserves on formerly damaged ecosystems. The other eight respondents were the W.K.Kellogg Biological Station, operated by Michigan State University (information from George H.Lauff); the Tyson Research Center, operated by Washington University (information from Richard W.Coles); the Sagehen Creek Field Station, operated by the University of California at Berkeley (information from Don C.Erman); the Turtle Cove Biological Research Station, operated by
Southeastern Louisiana University (information from Robert W.Hastings); Mountain Lake Biological Station, operated by the University of Virginia (information from J.J.Murray); the Notre Dame Environmental Research Center (information from Stephen R.Carpenter); Alice L.Kibbe Life Science Station, operated by Western Illinois University (information from John E.Warnock); and Milwaukee Field Station, operated by the University of Wisconsin (information from Millicent S.Ficken). All these are located on formerly damaged ecosystems, ranging from worn-out farms to sites affected by lumbering and other disturbances. Of the eight, four were formally designated as nature reserves or some other similar category. Two had no official designation, although it is clear they were serving the purpose without the formal designation. The formality of the designation was in some doubt for two, although their uses were not. In short, all were effectively being used as nature preserves, although the formality of the designation varied.
For the damaged sites in the University of California Natural Reserve System, no particular management practices were in effect other than letting nature take its course and excluding the ecologically damaging influences. For the others, there were some management practices such as planting trees, but these were relatively low-key undertakings. Efforts did not, for example, involve large-scale interventions such as the colonization of species. Probably more than half the sites could be described as ecological oases, although only in a few cases was the contrast with neighboring land startling.
Although this is a relatively small sample, it provides substantive evidence that:
-
Damaged ecosystems in temperate regions can recover rather rapidly in such a way that the biological diversity of the system is vastly improved. Much evidence suggests that tropical terrestrial ecosystems are far more fragile.
-
Simple management practices, such as benign neglect, often result in a vastly improved condition.
-
Good ecological research can be conducted on formerly damaged ecosystems, indicating that the processes in them are ecologically interesting and comparable in many respects to those in undisturbed ecosystems.
-
The quality of these ecosystems relative to most of the surrounding countryside is sufficiently superior to justify such designations as nature reserves or preserves and experimental ecological preserves.
OBSTACLES TO RESTORATION RESEARCH
The few laws regarding the restoration of damaged ecosystems tend to be so prescriptive that they often impede implementation of obvious solutions to relatively simple problems (Cairns, 1985, 1986). It is even more regrettable that laws do not encourage the experimentation so necessary to the development of both the science and the art of ecological healing.
Bradshaw (in press) espouses a systematic approach to each wasteland problem. In the first step, the qualities of the site and the needs of the region and its people must be interrelated. Bradshaw believes that the process requires both imagination and science but that the science has to be a particularly practical, constructional
form of ecology in many ways more allied to engineering than to much of modern analytical biology. The result is very rewarding. Noss (1983) cautions, however, that too much fragmentation may not produce the desired results and that a more comprehensive view is required for perpetuation of regional diversity. He recommends a regional network of preserves with sensitive habitats protected from human disturbance. This approach would enhance the alpha, beta, and gamma1 diversity levels of Whittaker (1972). But even oases may serve in the restoration of damaged ecosystems (e.g., Marsh and Luey, 1982).
All this will require a substantial adjustment in the outlook of most ecologists; however, since they will have only damaged ecosystems to study if present trends continue, it is in their enlightened self-interest to prevent further environmental degradation and reverse the trend where degradation has occurred. Fortunately, there are now newsletters (e.g., the U.S. Forest Service’s Rehabilitation News) and journals (e.g., Restoration and Management Notes, edited by William Jordan) to inform ecologists and other interested persons about this rapidly developing field.
Two zoologists and a medical doctor have explored an analogy between the practice of human medicine and the practice of stress ecology (Rapport et al., 1980, 1981). This approach may be quite useful in communicating ecosystem rehabilitation in societies now concerned with personal health.
PLANNING TO INCREASE DIVERSITY
Sites for restoration ecology may range from a few to hundreds of square kilometers. The Guanacaste National Park (GNP) in Costa Rica is 700 square kilometers. Janzen (in press) has a detailed plan to integrate this park into local and national Costa Rican society. The details of integration are discussed by Janzen in Chapter 14 of this volume. The park is large enough to maintain healthy populations of all animals, plants, and habitats known to have originally occupied the site and to contain an excess of needed habitat so that some can be intensively used by visitors and researchers. The GNP has three functions:
-
to use existing dry forest fragments as seed sources to restore about 700 square kilometers of topographically diverse land to a dry forest sufficiently large and diverse to maintain into perpetuity all animal and plant species and their habitats known to have occupied the site originally;
-
to restore and maintain a tropical wildland so that it can provide material goods such as plant and animal gene banks and stocking material; and
-
to use a tropical wildland as the stimulus and factual base for a reawakening of the intellectual and cultural offerings of the natural world to audiences with a local, national, and international philosophy that is user-friendly.
Janzen (in press) estimates a land purchase cost of $8.8 million and a start-up endowment of $3 million. As of May 12, 1987, Janzen (personal communication) states that 51% of the terrain had been purchased. These are relatively small costs for such a large project, and the world needs a large restoration pilot project with these goals. A few restoration efforts of this magnitude may be more cost-effective in increasing diversity than many smaller projects.
INTEGRATED RESOURCE MANAGEMENT
On July 8, 1986, I took my class on stressed ecosystems from RMBL to the Mexican Cut tract approximately 10 kilometers away. As of August 3, 1986, the total Mexican Cut tract owned by the Nature Conservancy and administered by RMBL was approximately 960 acres (384 hectares)—larger than the main site of the RMBL. The road through RMBL is no superhighway and was blocked east of Schofield Pass (elevation 10,707 feet, or approximately 3,200 meters) by 50 feet (15 meters) of snow from an avalanche. We hiked the remaining 4 miles (6.4 kilometers), fording an icy mountain stream and surmounting patches of snow in the process. Mexican Cut (elevation approximately 11,200 feet, or 3,360 meters) is not easily accessible by foot, yet John Harte, University of California at Berkeley, has been studying the effects of acid snow melt there for 6 years.
The students were astounded to find a stressed ecosystem in such a remote location. They carefully helped with a census of a declining population of salamanders (Ambystoma tigrinum). Who, the students asked, is supposed to prevent this from happening? A good question with an unsatisfactory answer.
Despite congressional intentions that one agency manage environmental problems, in reality that responsibility is fragmented and dispersed. This process of compartmentalization occurs within agencies as well. Turf battles occur within and between agencies. No quality control system can work effectively under these circumstances. At a meeting held in Charleston, South Carolina, in December 1985, I found that agency administrators were well aware of this problem and eager to do something constructive about it. Restructuring agency missions is not easy at the state level and is truly formidable at the federal level. Even then we are still in deep trouble, as demonstrated by discussions between Canada and the United States about acid rain.
We can make some progress in increasing diversity through restoration of damaged ecosystems without integrated resource management. However, as at Mexican Cut, deleterious materials (e.g., lead, aerosols; Elias et al., 1975) of known composition but uncertain and distant origin can affect and even destroy the best local and regional plans. Until we cope with the larger problems, the local successes will be temporary and therefore unsatisfactory.
ESTABLISHMENT OF ECOLOGICAL RESERVES
A committee of the National Research Council (NRC, 1981) recommended that ecological reserves be established throughout the country as sites for the field observations of baseline ecosystems. Its report further recommended, “Selected
ecosystems representative of geographic regions and unique or fragile systems should be studied and the results used in identifying key species, properties of populations, and ecosystem processes” (NRC, 1981, p. 78). If these ecological reserves were sufficiently large, they might also be used as a source pool of species to colonize damaged ecosystems. The studies of properties and processes useful as baseline information would be equally useful in rehabilitating damaged ecosystems, which could then serve as source pools of species for additional rehabilitation efforts, leaving the ecological reserve to serve its original purpose.
GNOTOBIOTIC ECOSYSTEMS
Taub (1969) has established gnotobiotic (species-defined) microcosms for toxicity testing and other purposes. Although these assemblages of species might never occur together in nature, they appear to function reasonably well together and have many interactions typical of natural systems. There are a number of situations where such artificial assemblages might be used on a larger scale to increase diversity. Some illustrative examples follow.
Power Line Right of Ways
Enormous areas under electric power transmission lines afford opportunities for creative ecological management. If these corridors were put together, they would probably approach the land area of one of our smaller states. Even in strips, however, they represent a considerable challenge. Vegetation is kept low by spraying or cutting. Both measures are expensive, and both may affect adjacent ecosystems. Drift from sprays may be toxic, and the corridor may be a barrier to some species. Development of an artificial assemblage of plant species that would not exceed a particular height would obviate the use of sprays or cutting and, if carefully structured, would be less of an ecological barrier than the present system.
Hazardous Waste Site Closure
There are a large number of hazardous waste sites in the United States where the highest concentrations of chemicals can be removed and transformed but where, for a variety of reasons (e.g., Novak et al., 1984, state that “acid leachate will continue to be emitted from this area for some years in the future”), lower concentrations must be left in place. Selection of species for colonizing these areas should be based on their ability to decrease the intrusion of these chemicals into groundwater supplies, to immobilize the chemicals within the site, and to accelerate degradation of the compounds into less harmful materials. Successful colonization requires not only that the organisms contribute to these benefits but also that they be tolerant of residual concentrations. Selection on this basis might well produce a species assemblage that has never occurred together in nature. Nevertheless, ecological benefits would be substantial as would benefits to diversity. Furthermore, an appropriate selection of a few organisms with only marginal tolerance to the hazardous materials would provide biological sentinels warning of unfavorable trends on the site. These assemblages could be replaced with natural assemblages as hazardous conditions diminish.
Reintroduction of Societal Wastes into the Environment
All sewage and many industrial waste-treatment plants depend heavily on biological processes, although one might not reach this conclusion from examination of an operator’s training manual. However, the ecological distance between the waste-treatment plant and natural ecosystems is too great. A gnotobiotic system designed to act as an ecological buffer between the two systems should help increase diversity in the natural system by more efficiently carrying out processes not completed by the waste-treatment system for which the natural system is not well equipped. Since most waste-treatment plants are not located in pristine ecological environments, these ecological buffers would be constructed on damaged or partly damaged ecosystems.
The selection of species for these assemblages should be based on their tolerance for the waste and on their ability to carry out the remaining transformation processes. They might or might not occur naturally together. The only contaminants one can easily process in this way are those from point sources. However, by appropriate design of run-off collection systems, nonpoint discharges, such as agricultural wastes and surface mine wastes, can be converted to point source discharges. Since most of these contaminants are waterborne, artificial wetlands are probably the best buffer ecosystems for coping with them. Such systems can tolerate heavy loadings of both organic wastes and some toxicants. Recent descriptions may be found in Brooks et al. (1985).
GROUNDWATER DETOXIFICATION
There is persuasive evidence of groundwater contamination in North America, but its extent has not been well defined. There seems to be little concern about the fate of the organisms that inhabit the aquifers, however, even though they occupy a unique habitat and are part of the global diversity. Some of the contaminated aquifers might be partly decontaminated by the introduction of engineered microorganisms to degrade the wastes. Although presumably less dangerous, the water would not yet be normal. In some cases, quality could be further improved by pumping the water to the surface and passing it through an artificial wetland before recharge through a pipe or by normal infiltration.
The many contaminated and depleted aquifers could be recharged by artificial wetlands designed for this purpose. These can be built on damaged ecosystems, as Brooks et al. (1985) have shown.
PROSPECTS FOR RESTORATION ECOLOGY
Damaged ecosystems can be changed from ecological liabilities to assets that are useful in both increasing diversity and protecting natural systems in a variety of ways. They can be developed as ecological buffers between waste-treatment systems and natural systems. Damaged ecosystems can be used to replace wetlands and other scarce habitats lost to development, even when such habitats did not originally exist on the damaged site. They can be developed as refuges for rare, en-
dangered, or threatened species. Damaged ecosystems that may act as ecological barriers (e.g., power line right of ways) can be converted to “bridges” between the ecosystems on either side. Finally, they can be used to recharge and possibly purify our groundwater aquifers.
None of these desirable events will happen unless the present prescriptive regulations on rehabitation of damaged ecosystems are made more flexible to encourage experimentation. The sizable information base needed for effective ecosystem rehabilitation will not be generated until more ecologists are willing to work on damaged ecosystems.
It is probably not an exaggeration to say that much of the planet is occupied by partially or badly damaged ecosystems. Restoring them is probably the best means of increasing diversity. If we put as many resources and as much energy into restoring this planet as we have into the space program, Curry’s (1977) vision of reinhabiting Earth might become a reality.
REFERENCES
Bradshaw, A.D. In press. Alternative end points for reclamation. In J.Cairns, Jr., ed. Rehabilitating Damaged Ecosystems. CRC Press, Boca Raton, Fla.
Brooks, R.P., D.E.Samuel, and J.B.Hill, eds. 1985. Wetlands and Water Management on Mined Lands. Proceedings of a conference, October 23–24, 1985, Pennsylvania State University. Pennsylvania State University, University Park. 393 pp.
Bruns, D. In press. Restoration and management of ecosystems for nature conservation in West Germany. In J.Cairns, Jr., ed. Rehabilitating Damaged Ecosystems. CRC Press, Boca Raton, Fla.
Brussard, P.F. 1982. The role of field stations in the preservation of biological diversity. BioScience 32(5):327–330.
Cairns, J., Jr., ed. 1980. The Recovery Process in Damaged Ecosystems. Ann Arbor Science Publishers, Ann Arbor, Mich. 167 pp.
Cairns, J., Jr. 1985. Keynote address: Facing some awkward questions concerning rehabilitation management practices on mined lands. Pp. 9–17 in R.P.Brooks, D.E.Samuel, and J.B.Hill, eds. Wetlands and Water Management on Mined Lands. Proceedings of a conference, October 23–24, 1985, Pennsylvania State University. Pennsylvania State University, University Park.
Cairns, J., Jr. 1986. Restoration, reclamation, and regeneration of degraded or destroyed habitats. Pp. 465–484 in M.Soulé, ed. Conservation Biology: The Science of Scarcity and Diversity. Sinauer Associates, Sunderland, Mass.
Cairns, J., Jr. 1987. Disturbed ecosystems as opportunities for research in restoration ecology. Pp. 307–320 in W.R.Jordan III, M.E.Gilpin, and J.D.Aber, eds. Restoration Ecology: A Synthetic Approach to Ecological Research. Cambridge University Press, New York.
Cairns, J., Jr. In press. Restoration ecology: The new research frontier. In J.Cairns, Jr., ed. Rehabilitating Damaged Ecosystems. CRC Press, Boca Raton, Fla.
Curry, R.R. 1977. Reinhabiting the earth: Life support and the future primitive. Pp. 1–23 in J. Cairns, Jr., K.L.Dickson, and E.E.Herricks, eds. Recovery and Restoration of Damaged Ecosystems. University Press of Virginia, Charlottesville.
Elias, R., Y.Hirao, and C.Patterson. 1975. Impact of present levels of aerosol Pb concentrations on both natural ecosystems and humans. Pp. 257–272 in International Conference on Heavy Metals in the Environment, October 27–31, 1975. Toronto.
Janzen, D.H. In press. Guanacaste National Park: Tropical ecological and cultural restoration. In J.Cairns, Jr., ed. Rehabilitating Damaged Ecosystems. CRC Press, Boca Raton, Fla.
Mabbutt, J.A. 1984. A new global assessment of the status and trends of desertification. Environ. Conserv. 11:103–115.
Marsh, P.C., and J.E.Luey. 1982. Oases for aquatic life within agricultural watersheds. Fisheries 7(6):16–19, 24.
NRC (National Research Council). 1981. Testing for Effects of Chemicals on Ecosystems. National Academy Press, Washington, D.C. 103 pp.
Noss, R.F. 1983. A regional landscape approach to maintain diversity. BioScience 33(11):700–706.
Novak, J.T., W.R.Knocke, M.S.Morris, G.L.Goodman, and T.Jett. 1985. Evaluation of an acidic waste site cleanup effort. Pp. 111–120 in Proceedings of the 40th Industrial Waste Conference. May 14–16, 1985, Purdue University, West Lafayette, Ind. Butterworth, Boston.
Rapport, D.J., C.Thorpe, and H.A.Regier. 1980. Ecosystem medicine. Pp. 179–182 in J.B. Calhoun, ed. Perspectives on Adaptation, Environment and Population. Praeger Scientific, New York.
Rapport, D.J., H.A.Regier, and C.Thorpe. 1981. Diagnosis, prognosis and treatment of ecosystems under stress. Pp. 269–280 in G.W.Barrett and R.Rosenberg, eds. Stress Effects on Natural Ecosystems. Wiley, Chichester, United Kingdom.
Taub, F. 1969. Gnotobiotic models of freshwater communities. Vehr. Internat. Verein. Limnol. 17:485–496.
Vandenbusche, D. 1981. The Gunnison Country. Vandenbusche, Gunnison, Colo. 472 pp.
Vitousek, P.M., P.R.Ehrlich, A.H.Ehrlich, and P.A.Mason. 1986. Human appropriation of the products of photosynthesis. BioScience 36(6):368–373.
Whittaker, R.H. 1972. Evolution and measurement of species diversity. Taxon 21:231–251.
CHAPTER 39
RESTORING DIVERSITY
The Search for a Social and Economic Context
JOHN TODD
President, Ocean Arks International, Falmouth, Massachusetts
Restoration ecology is beginning to develop into two quite distinct disciplines. Although they have a lot in common, it is the differences between the two directions that are important. The first type of restoration ecology is primarily an academic field of activity in which there is an attempt to recreate authentic ecosystems of the past, particularly those that have been destroyed or modified by human alteration or abuse. There is an emphasis on selecting the correct species mix, and there is an effort to recreate the original ecological relationships at least as far as they are known. Exotic species or organisms intrinsic to other environments are shunned. Here, whether a forest or a prairie is the focus, restoration means recreating the original state both in terms of structure and species. These newly created ecosystems have the intrinsic value of maintaining important gene pools in regions where the organisms have previously flourished. They can also teach a great deal about the dynamic processes of ecosystems and succession. The pioneering work at the University of Wisconsin-Madison Arboretum (see Jordan, Chapter 35) falls into this category.
The second approach to restoration ecology operates from a different set of assumptions. The overall objective is different, although much of the knowledge and techniques are comparable. A hypothetical example might help to illustrate what I mean. If a mixed-forest hillside is logged and clear-cut, then turned into pasture for cattle, later grazed by sheep, and finally allowed to lose its topsoil and erode into gullies that eventually can support only coarse grasses and thistles, it is apparent that human activity has destroyed a complex and diverse habitat and replaced it with a degraded one. The hill’s ability to support abundant life is reduced as well as its capability to underwrite economic activity. In this second category,
the restoration ecologist is interested in the structure or architecture of the original forest because of its ability to do things that cannot be done by degraded and eroded soils. The forest can build soils, control the capture and release of moisture, and regulate nutrient cycles. It can withstand climatic fluxes and perturbations, and critically important to the naturalist, it can support an enormous spectrum of life forms. The forest’s dimensions extend from the tree canopy to root depths often tens of meters below the soil surface.
In the hillside example, the structural, as distinct from the species-specific, restoration ecologist wants to build fertile soils, develop a water and nutrient regimen, and assemble an ecosystem that mimics the original structural integrity of the forest. The actual organisms selected to do the job may or may not be the original species. Often, the ecologist will seek out equivalent species of plants or animals that have secondary properties as well. For example, a tree species that is not adapted to the environment, but is highly valuable, provides an economic dimension to the process. Also the land restorer, like a farmer, will in most instances use a large array of biotechnologies and technological aids to orchestrate and even speed up successional and other biological processes. Sophisticated bioengineering is used to recreate the equivalent of hundreds of years of topsoil within a decade or two.
The mission of the two restoration ecologies can be quite different too. Structural ecologists are willing to tolerate wider margins of uncertainties and gaps in knowledge, thereby adopting a more applied viewpoint simply because they see the planetary environmental crisis as the backdrop against which they work. To reverse desertization and habitat destruction, it ultimately will be necessary to undertake ecological restoration on a vast, planetary scale. This means that the task cannot be guided by charity based on social conscience, since there isn’t enough of either even to finance or underwrite the required backup ecological research. It is hard to avoid the conclusion that if there is to be any meaningful change, restoration ecology will have to become quite simply a major economic activity. Just as the activity seeks to recreate the forest on the hill, it will also be expecting the hill to become a sustainable and environmentally enhancing economy. A goal will be the provision of a wide variety of marketable products as a by-product of the restoration process. This is an extremely ambitious objective for a young field of endeavor, but one that is essential to its widespread application.
Fortunately there exists, albeit widely scattered, the biological knowledge and field experience to build a science and practice of habitat recovery. This knowledge needs to have an ecological framework whose elements will be found in geology, climatology, agriculture, forestry, horticulture, aquaculture, limnology, research ecology, landscape architecture, and natural history, to name some of the more relevant fields of endeavor. There will also be a need to develop and adapt tools and machines for use in agriculture, earth moving, forestry, waste management, and process engineering.
Returning to our hypothetical example of the eroded hillside, the first step in the restoration process would be to arrest rapid rainwater runoff and to reduce erosion. Among the many approaches that can be taken are various tillage techniques, microcatchments, contouring, and land sculpturing to regulate water move-
ment. Most of the methods require sophisticated machines, careful engineering, and timed planting to be successful. Next, the degraded soils, if they are to be rebuilt quickly, will require the precise addition of minerals, fertilizers, organic matter, and vegetation to effect rapid stabilization and to increase its moisture retention. The subsequent step, intentionally recreating a forest, requires a knowledge not only of the original forest cover but also of a range of equivalent species that play an analogous role while serving as a key economic component. This ecosystem is in some respects a cross between a forest and an orchard. In our hypothetical example the wild deer of the original may be replaced by domestic animals—not necessarily cattle, however, but by the European fallow deer (Cervinae dama), which fit ecologically and are highly marketable because of the extremely low cholesterol levels in their flesh.
The goal of this ecological restoration is the production of food and fiber on a commercial scale. It is not agriculture but an ecology with agricultural elements within a broader biological framework. It does not have the environmental destructiveness of monocrop agriculture or simpler agricultural systems. Its function is to restore diversity and to be bountiful in terms directly useful to humans. Linking together nature’s restoration requirements with the economic needs of people may be the only way the terrestrial fabric of the planet can be rebuilt.
Large amounts of human labor are essential to the restoration process. For example, labor is needed to plant new trees and to provide them with adequate moisture and protection from weed competition or predation by grazing animals. To support the required labor, restoration ecology will have to attract capital. The future hillside ecosystem will have to be seen as a prudent investment, possibly providing favorable returns within years rather than decades.
The goal of much of my research and planning over the last 15 years has been to find ways of economically underwriting the restoration and diversification process. This has involved the development of a family of biotechnologies that are in essence short-cycle ecosystems with economic by-products that also have the capacity to catalyze the longer-cycle restoration processes. These biotechnologies have been proven successful and in some contexts have been shown to be cost-effective and economically feasible (Todd and Todd, 1984). We have not yet had the opportunity to integrate all the subsystems into a full-scale restoration project, but an outline of a restoration project in the Mediterranean has been prepared (Todd, 1983, 1984). In addition, projects for the west coast of Costa Rica and the Atlantic coast of Morocco are now in the planning stage.
The Costa Rican project is intended to reclaim lands badly degraded because of earlier inappropriate agriculture. The proposed Moroccan project involves creating a diverse plant environment where the desert and the sea meet. In this instance, the newly created ecosystems will provide part of the underpinnings for a new human settlement.
In all the above examples, the land is not currently fit for intensive agriculture or easy restoration. A new biotechnology, which we have named the desert-farming module, provides the short-term ecological economy needed to initiate the restoration cycle. The development of this technology began at the New Alchemy Institute under my direction in 1974. In summary, a desert-farming module is a
translucent solar-energy absorbing cylinder with a capacity up to 1,000 gallons (3,785 liters) that is filled with water and seeded with more than a dozen species of algae and a complement of microscopic organisms (Figures 39–1, 39–2, and 39–3). Within these cylinders, phytoplankton-feeding fishes and omnivorous fishes are cultured at very high densities. The species selected depend upon climate, region, and market opportunities: the range of species we have studied is broad, including African tilapia (Tilapia spp.), Chinese carps (Cirrhinus molitorella, Ctenopharyngodan idellus, Hypophthalmichthys molitrix, and Aristichthys nobilis), and North American catfish (Ictalurus spp.) and trout (Saluelinus spp.).
Dense populations, up to one actively growing fish per 2 gallons (7.6 liters), produce high levels of waste nutrients beyond the capability of the ecosystem to take up. The module recycles these nutrients in the following ways: nutrients released from the decomposed waste material are absorbed by the fish, the plankton, and the crop plants rafted on the cylinder surface. In addition, partially digested algae that floculate out and settle to the bottom are periodically discharged through a valve to fertilize and irrigate the surrounding ecosystem under restoration. The root systems of the vegetable crop plants take up the nutrients before they reach toxic levels and also capture detritus; they function as living filters by purifying the water.
These modules can yield more than 250 pounds (113.5 kilograms) of fish annually in a 25-square-foot (2.3-square-meter) area, depending on species and supplemental feeding rates. At the same time, each unit can produce 18 heads of lettuce weekly, i.e., more than 900 heads per year (Zweig, 1986). Tomatoes and cucumber crops

FIGURE 39–1 Solar aquaculture ponds. A dense population of fishes can be seen in the module in the foreground. Photo by J.Todd.

FIGURE 39–2 Solar aquaculture ponds in winter conditions on Cape Cod, Massachusetts. Photo by J.Todd.
can also be grown on the surface for even higher economic yields. An additional benefit of the modules is conservation of water. Since evaporation is almost eliminated from the surface, rates of water replenishment are based on plant evapotranspiration and the amount of water released from the module to irrigate and fertilize the adjacent area (Figures 39–3 and 39–4).
Desert-farming modules are an agroecology that require initial seed capital to construct and establish. But to a large extent, tillage, harvesting, fertilizing, and irrigation are a substitute for the heavy equipment that would otherwise have to be used for establishing and operating a farm on degraded soils. Not only are the

FIGURE 39–3 Desert-farming module in solar greenhouse with lettuce growing hydroponically on surface. Photo by R.Zweig.
modules less costly than the equipment, but it may well turn out that they are much more likely to support the restoration process technically and economically.
Within a given land restoration project, the modules could be established in rows in the most highly degraded areas. Young trees on the shaded side of these cylinders could be planted and subsequently nurtured by the periodic release of water and nutrients. On the sunny side of the modules, a variety of short-term economic crops could be established to add to the produce from the module. The labor needs for the module-based agriculture could also be used to tend the emerging ecosystems.
The approach based on the desert-farming module need not be static in the sense that the modules, having fed and watered the newly emergent vegetation including trees through their most vulnerable stages, could be shifted to new locations to repeat the process. In this way, the short-cycle biotechnology could spread its benefits to surrounding ecosystems over a larger geographic area.
For arid environments, such as the Moroccan coast, we have developed a bioshelter system to assist with diversifying an area ecologically. The bioshelter is a transparent climatic envelope or greenhouse structure that houses the fish and vegetable modules. Our prototype is a circular geodesic structure that functions as a solar still and as an incubator for the early stages of the ecological diversification process (Figure 39–5). These bioshelters can even operate where there is no fresh water. In this extreme case, the aquaculture modules are placed inside the climatic envelope and water from the sea is pumped through them. During the day, the

FIGURE 39–5 Prototype bioshelter. A geodesic dome housing fish and vegetable modules. Photo by J.Todd.
structure heats up until the temperature differential between the seawater in the tanks and the air is great enough to cause the tanks to sweat fresh water, which irrigates the ground around them. Tree seedlings are then planted in this moist zone. Drought-tolerant trees can also be established around the outer periphery of the structure. At night, the moisture-laden air cools to the desert sky, causing water droplets to form on the interior skin of the climatic envelope. With the prototype shelter, we found that drumming on the structure’s membrane in the early morning caused the droplets to fall like rain inside, thereby making it possible to plant the entire interior of the module. Inside the tanks, marine fish and crustacea such as mullet and shrimp can be cultured to form the basis of an economy. After a few years, the original cluster of climatic envelopes can be moved to a new location to repeat the cycle, leaving an established, semiarid agroecosystem behind.
These are two examples drawn from a range of biotechnological options that could help reverse environmental degradation and restore diversity and bounty to a region. These advanced technologies may well prove to be essential tools in creating sustainable environments.
In all of this, there is of course the fundamental question of land tenureship and social constraints that will ultimately drive any ecological changes. One option for countries with private land holdings and a willingness to tackle serious long-range environmental issues might be the creation of restoration corporations. In this particular model, a corporation would have the financial capability of buying large blocks of ruined land and to hire and train local farmworkers to operate the desert-farming modules, process the foods and other by-products, and tend the
emerging ecosystems. When the ecosystems are ready for agriculture, natural resource management, and conservation on a given section of land, the corporation could divest itself of the land, selling it to the formerly landless workers who would be trained land stewards and farmers. The original corporation would become a mortgage banking company for the new landholders and at the same time continue to own and operate part of the infrastructure, including the training component, and to share in processing and marketing. Its earnings would be used in turn to acquire new blocks of land and repeat the cycle. In this particular model, there is the potential theoretically to link capital, institutional structures, information, and family-held productive lands to effect land restoration. There is also an opportunity to set aside lands as wilderness, since the intensive agriculture will reduce pressure on the overall environment. Even in poorer parts of the world, there is the chance to generate enough wealth to underwrite continuing ecological research out of which new models of Earth stewardship will arise.
REFERENCES
Baum, C.M. 1981. Gardening in fertile waters. New Alchemy Q. Summer (5):3–8.
Todd, J. 1983. Planetary healing. Annals of Earth Stewardship 1(1):7–9.
Todd, J. 1984. The practise of stewardship. Pp. 152–159 in W.Jackson, W.Berry, and B.Coleman, eds. Meeting the Expectations of the Land. North Point Press, San Francisco.
Todd, N., and J.Todd. 1984. Bioshelters, Ocean Arks, City Farming: Ecology as the Basis for Design. Sierra Club Books, San Francisco. 210 pp.
Zweig, R. 1986. An integrated fish culture hydroponic vegetable production system. Aquaculture 12(3):34–40.












































