
16
Tertiary Marine and Nonmarine Climatic Trends
JACK A.WOLFE and RICHARD Z.POORE
U.S. Geological Survey
INTRODUCTION
Climates of the Tertiary period have been more thoroughly studied and discussed than those of any pre-Quaternary interval. Despite this seeming wealth of data (or perhaps because of such wealth), many researchers have arrived at differing conclusions about both the intensity of warm intervals and temperature fluctuations during the Tertiary. Some of the differences are related to sampling at widely spaced stratigraphic intervals, imprecise correlations of samples, and/or the use of different techniques to arrive at paleotemperature estimates. These problems are greatly intensified in attempting to relate marine and nonmarine trends; to overcome one or more of these problems, our discussion will concentrate on two geographic regions.
PALEOGENE OF THE ATLANTIC BASIN
The Paleogene of the Mississippi embayment of southeastern North America contains an extensive sequence of land floras. These flora were, for the most part, included by Berry (1916) in his “Wilcox flora” and were thus considered by him to be approximately synchronous. Work by many other stratigraphers in recent decades has demonstrated that Berry’s “Wilcox flora” was based on material from rocks of early Paleocene through late Eocene Age.
These Mississippi embayment floras all represent lowland vegetation, and thus the altitudinal variations that can affect analysis of floras in continental interiors are negated. In turn, the data on land floras can be related to paleoclimatic models based on data from deep-sea sediments in the North Atlantic. Reliance on data from the North Atlantic and its borderlands also has the advantage that this region has had a simple and well understood plate-tectonic history during the Tertiary in comparison with that of many other regions, and thus paleolatitudes are well known.
The interpretation of climatic change during the Paleogene in the North Atlantic is based both on paleontological evidence and on isotopic data (Figure 16.1). The paleontological interpretations largely follow previously published data (Haq and Lohmann, 1976; Haq et al., 1977), which involve recognizing distinctive latitudinally zoned nannofossil assemblages and tracing the latitudinal migrations of these assemblages through time. However, an exception is our interpretation of the mid-
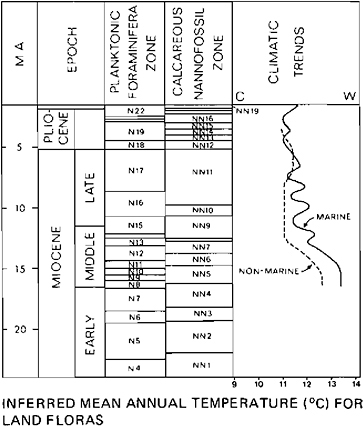
FIGURE 16.1 Comparison of temperature trends in the North Atlantic Paleogene with inferred temperatures of Paleocene and Eocene land floras from the Mississippi embayment. Extent of bars indicates possible age range of land floras in terms of planktonic time scale.
dle Eocene Discoaster assemblage (Haq et al., 1977) as representing low- rather than middle-latitude climate. Our interpretation is in accord with the isotopic record from the Deep Sea Drilling Project (DSDP) site 398 (Vergnaud-Grazzini, 1979); data on the early Paleocene climate is also based largely on somewhat meager isotopic records.
Interpretations of the land floras are based primarily on physiognomic analyses of assemblages (Wolfe, 1971, 1978). Such interpretations rely on the present observed correlation between the physical features of leaf assemblages and particular climatic regimes rather than on the identification of particular taxa in fossil assemblages and the present distribution of related taxa.
The marine and nonmarine records are, overall, strikingly similar in the timing of major temperature fluctuations during the Paleocene and Eocene. Marked warm intervals occurred in the early Paleocene, latest Paleocene-early Eocene, late middle Eocene, and latest Eocene. Note that our biostratigraphic and radiometric calibrations indicate that a major decrease in mean annual temperature occurred at about 39 million years ago (Ma); we suggest that the “terminal Eocene” cooling of some workers (e.g., Thierstein and Berger, 1978) is in fact this major cooling within the late Eocene. The subsequent latest Eocene warm intervals were also followed by a major cooling event following the Eocene-Oligocene boundary (34–35 Ma) that led to temperatures cooler than any known in the Eocene. The Mississippi embayment sequence is not known to contain land floras of early Oligocene age; however, sequences of radiometrically dated floras in western North America document this cooling event (Wolfe, 1971). The event at the Eocene-Oligocene boundary is particularly pronounced on land because the event was characterized by both a major decrease in mean annual temperature and a major increase in mean annual range of temperature (Wolfe, 1978).
The degree of warmth indicated for the Paleogene has been much discussed; although the consensus is that all the Paleocene and Eocene was warmer than present, some workers consider that the level of warmth was only moderately higher than now and that the highly equable climates were the primary factor in allowing the poleward excursions of tropical and subtropical organisms (e.g., Axelrod and Bailey, 1969). Our data indicate that, during warm intervals in the Eocene, mid-latitude temperatures were significantly higher than today. Tropical-subtropical nannoplankton assemblages in the North Atlantic that are the analogs of the Paleogene tropical-subtropical assemblages are now confined to latitudes 25–28° N. During times of the Paleocene and Eocene, tropical-subtropical species form a significant part of nannoplankton assemblages far poleward of 30° N.
The early Eocene appears to represent the warmest part of the Tertiary. The marine record in the North Atlantic, as well as elsewhere, indicates that the most poleward (50–55° N) excursion of low-latitude assemblages occurred at this time; the tropical-subtropical belt may have been double its present latitudinal extent. Data from land plants are also indicative of a major expansion of tropical climates (i.e., mean annual temperature ~25°C)—climates that are currently found within 20–25° of latitude from the equator. The Mississippi embayment floras that occur as far north as 36° latitude are tropical during the warm intervals of the early and middle Eocene. Floras from the west coast of North America were, during the late middle Eocene, tropical to north of latitude 40–45° (Wolfe, 1978); latest Eocene floras indicate paratropical climate at these latitudes. Note that extratropical latest Eocene planktonic foraminiferal assemblages are associated with paratropical land floras in the Pacific Northwest. Thus the juxtaposition of relatively warm land climates with relatively cool marine conditions characteristic of the U.S. Pacific margin today was established in the late Eocene.
On the Gulf of Alaska borderlands (60° N), land floras indicating mean annual temperatures as high as 22°C were thought to be of middle Eocene age based on provincial correlations by marine mollusks (Wolfe, 1977). In the planktonic chronology, however, these Alaskan paratropical floras and the associated mollusks are early Eocene, and other Alaskan floras referable to the late middle Eocene, although indicative of considerable warmth, represent cooler conditions than the early Eocene floras. Similarly, planktonic foraminiferal and nannoplankton assemblages from early Eocene rocks of the eastern Gulf of Alaska contain a major low-latitude component (Poore and Bukry, 1979). Early Eocene floras from Ellesmere Island certainly represent mean annual temperature in excess of 3°C, i.e., at least 22°C higher than today (L.J.Hickey, Smithsonian Institution, personal communication).
In combination with significantly higher mean annual temperatures during parts of the Eocene, mean annual range of temperature and latitudinal temperature gradients were less. Western Washington, which today has mean annual ranges of temperature of 11–17°C, had a mean annual range of temperature as low as 4–5°C during the Eocene (Wolfe, 1978). The latitudinal temperature gradient from middle to high latitudes along western North America was about half the present gradient.
Not only were Eocene temperatures at middle to high latitudes significantly different than present temperatures, precipitation patterns were different. Areas of the western United States that now experience summer drought have Eocene floras that indicate abundant precipitation throughout the year. In contrast, the Paleogene floras from the Mississippi embayment—a region that now has abundant precipitation distributed throughout the year—exhibit a trend from tropical moist to tropical dry climate.
The temperature fluctuations indicated by our analysis of the North Atlantic marine record are difficult to quantify, but the latitudinal differences between the occurrences of tropical-subtropical assemblages indicate fluctuations in mean annual temperature of at least 10–15°C at high middle latitudes; these fluctuations were, thus, at least as great as modern seasonal fluctuations. No fluctuations are observable at low latitudes, i.e., the present tropical belt was always tropical during the Paleogene. Wolfe (1978) has suggested that the land floras from middle latitudes indicate fluctuations in mean annual temperatures of about 7°C.
The elevated temperatures suggested for Paleogene warm intervals concommitant with the major temperature fluctuations are difficult to explain by purely geographic or meteorological mechanisms endemic to the Earth. That is, low latitudes were consistently tropical, and high latitudes were at the same times significantly warmer; redistribution of heat to warm high latitudes should produce cooling at low latitudes, no matter what mechanisms resulted in redistribution. Geographic factors (e.g., the circumglobal circulation at low latitudes, generally lower altitudes of the continents, and greater extent than now of epicontinental seas) could have produced the somewhat elevated global temperature of the cool intervals during the Paleocene and Eocene compared with the global temperature today, but the high levels of warmth during, for example, the early Eocene cannot be so readily explained.
Re-expansion of low-latitude planktonic assemblages during the latest Eocene indicates a renewed warning, but these assemblages did not reach poleward as far as during warm intervals earlier in the Paleogene. Land-plant data from western North America also indicate that the latest Eocene warm interval (Kummerian Stage) was not so warm as previous warm intervals (Wolfe, 1978). This renewed warming was followed by a cooling that, for much of the Oligocene, resulted in low-latitude planktonic assemblages being restricted to about their present latitudes. The land floras that followed this terminal Eocene cooling indicate that not only was mean annual temperature on land significantly lowered, mean annual range of temperature dramatically increased to values even higher than today at middle and high latitudes of western North America (Wolfe, 1978). By the end of the Oligocene, some warming had occurred, as indicated both by planktonic microfossil and isotopic data from the North Atlantic and land floras in western North America.
NEOGENE RECORD OF THE NORTHEASTERN PACIFIC
Interpretations of temperature changes in the northeastern Pacific (Figure 16.2) during the Miocene and Pliocene are based on a variety of data. These include the varying latitudinal distributions of assemblages of calcareous nannoplankton, diatoms, silicoflagellates, and planktonic foraminifers from DSDP Legs 18 and 63 (Ingle, 1973; Barron, 1981; Bukry, 1981; Poore, 1981). We have also used isotopic data on benthic foraminifers from DSDP Site 289 [Cenozoic Paleo-oceanography Research Project (CENOP), unpublished data] to aid and supplement interpretations based on marine floras and faunas. The isotopic data presumably monitor intermediate waters at this site (Ontong Java Plateau) and thus, in part, mid- to high-latitude surface-water temperatures. Our interpretations of the marine record can be compared with analyses, again largely physiognomic, of land floras from the Pacific Northwest (Wolfe, in press) and Alaska (Wolfe and Tanai, 1980).
Temperature trends for the oceans during the early Miocene are problematic, primarily because of the lack of adequate time control. Meager isotopic data from Site 289 and elsewhere indicate that the early Miocene may have been somewhat cooler than the early part of the middle Miocene. What is not known is the warmth of the early Miocene relative to the late Oligocene. On land, the early Miocene (early Seldovian Age) floras are of cooler aspect than those of the middle Miocene as well as those of the late Oligocene. Age control on these floras, however, is uncertain relative to the planktonic time scale.
The late part of the early Miocene and the early part of the middle Miocene has generally been regarded as the warmest interval of the Neogene, although not so warm as any part of the Paleocene or Eocene. This warm interval is clearly recorded in deep-sea records in the northeastern Pacific. Following planktonic foraminiferal zone N11, a cooling trend set in that continued through the end of the Neogene, although some minor warmings of short duration occurred during the late Miocene. The youngest marked late Miocene warming at 8 Ma was followed by a gradual cooling; minor warm intervals occurred in the early Pliocene and in the early Pleistocene.
Whether the late Miocene through early Pleistocene temperature fluctuations seen in the northeastern Pacific represent worldwide climatic events is difficult to ascertain. At least some of these fluctuations may relate to changing current patterns in the northeastern Pacific, including changes in the intensity of upwelling—an indication that at least some of the late Miocene fluctuations may not represent global events in Haq’s (Chapter 13) data on Miocene nannoplankton assemblages from the Atlantic. The latitudinal distribution of these assemblages indicates that apparently only one significant warming occurred during the late Miocene rather than the three that occurred in
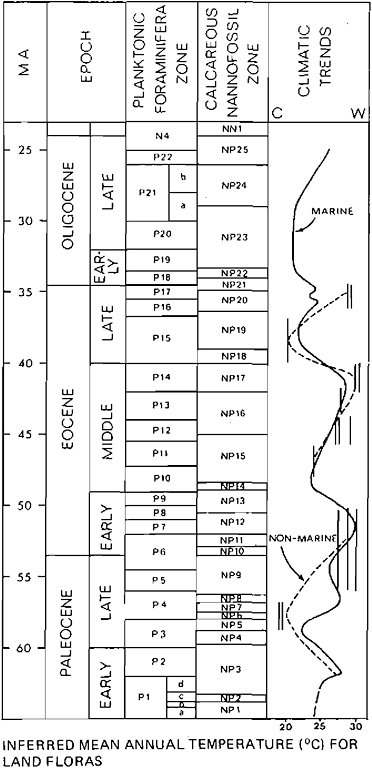
FIGURE 16.2 Comparison of temperature trends in the North-eastern Pacific middle Miocene through Pliocene with inferred temperatures of land floras.
the northeastern Pacific. Haq’s data, however, match the Pacific record in showing a latest Miocene cool event followed by a renewed warming into the early Pliocene.
In general, the land-plant record from the Neogene of the northeastern Pacific margin compares well with the deep-sea record: a temperature high in the middle Miocene (floras radiometrically dated at 17–15 Ma) followed by cooling, a slight renewed warming at about 8 Ma, and followed by a cooling at about 5 Ma. The land floras, however, also furnish important data on two other climatic factors.
First, in this region equability has clearly increased during the Oligocene and Neogene from the highly inequable temperature regime of the early Oligocene. At latitude 60° N, the increase in equability has largely resulted from a decline in summer temperatures (a decrease of about 9°C in the mean of the warm month), and at latitude 45° N summer temperature has decreased concommitant with an increase in winter temperature. The decline of summer temperature at higher latitudes must have been a major factor in the initiation of widespread northern hemisphere glaciation during the late Cenozoic.
The trends of temperature changes at various latitudes have also resulted in an increase in latitudinal temperature gradients. In turn, the effect of this increase may have been to intensify the subtropical high-pressure systems that are largely responsible for summer drought along the west coasts of the continents. The land floras from western United States definitely record a shift from a regime in which much of the precipitation was received during the summer to a regime of summer drought even in coastal areas.
The temperature fluctuations in the marine record during the late Miocene and Pliocene do not appear to be so significant as those in the Paleocene and Eocene. The Neogene fluctuations may represent changes in mean annual temperature of as much as 5°C for marine water at middle latitudes. This condition probably results from intensification of a well-defined and relatively stable eastern boundary current system—the California current—sometime in the late middle Miocene. This area of the northeastern Pacific exhibited great stability during the Quaternary, as evidenced by comparison of sea-surface temperatures at the last glacial maximum (18,000 yr ago) and today (CLIMAP Project Members, 1976, Figure 2). The land floras, however, do not indicate as much fluctuation of mean annual temperature; changes in this parameter at middle latitudes appear to be no more than 1–2°C.
SUMMARY
Paleotemperature trends interpreted from deep-sea marine organisms and from land floras are generally similar to one another, both in the timing and direction of changes. Both groups of organisms indicate that a thermal maximum occurred in the early Eocene and that major fluctuations in mean annual temperature occurred during the Paleogene. The land floras also indicate that mean annual range of temperature was low during the Paleocene and Eocene (i.e., a low seasonal contrast of temperature), although mean annual range increased to values greater than present by the Oligocene. Following a middle-Miocene warm interval, mean annual temperatures overall have decreased in extratropical areas, although this trend was interrupted by some minor reversals. The land floras indicate that, during the Neogene, the seasonal contrast of temperatures has decreased. Latitudinal temperature gradients were low during the Paleocene and Eocene and increased to their present values during the Neogene.
Precipitation patterns derived from studies of land floras indicate that an increasing drying trend occurred in southeastern North America during the Paleocene and Eocene; at the same time, western North America appears to have received abundant precipitation throughout the year. In the Oligocene and earlier half of the Miocene, western North America appears to have had abundant summer precipitation, but during the later Neogene summer precipitation decreased.
ACKNOWLEDGMENTS
John A.Barron released unpublished data on Neogene diatoms of the northeastern Pacific and extensively discussed with us their paleoclimatic significance. David Bukry furnished unpublished data on Neogene silicoflagellates. Information on Atlantic Neogene nannoplankton was furnished by Bilal Haq.
REFERENCES
Axelrod, D.I., and H.P.Bailey (1969). Paleotemperature analysis of Tertiary floras, Palaeogr. Palaeoclim. Palaeoecol. 6, 163–195.
Barron, J.A. (1981). Late Cenozoic diatom biostratigraphy and paleoceanography of the middle-latitude eastern North Pacific, DSDP Leg 63, in Initial Reports of the Deep Sea Drilling Project 63, U.S. Government Printing Office, Washington, D.C., pp. 507–538.
Berry, E.W. (1916). The lower Eocene floras of southeastern North America, U.S. Geol. Surv. Prof. Pap. 91, 481 pp.
Bukry, D. (1981). Silicoflagellate stratigraphy of offshore California and Baja California, Deep Sea Drilling Project Leg 63, in Initial Reports of the Deep Sea Drilling Project 63, U.S. Government Printing Office, Washington, D.C., pp. 539–557.
CLIMAP Project Members (1976). The surface of the ice-age Earth, Science 191, 1131–1137.
Haq, B.U., and G.P.Lohmann (1976). Early Cenozoic calcareous nannoplankton biogeography of the Atlantic Ocean, Mar. Micropaleontol. 1, 119–194.
Haq, B.U., I.Premoli-Silva, and G.P.Lohmann (1977). Calcareous plankton paleobiogeographic evidence for major climatic fluctuations in the early Cenozoic Atlantic Ocean, J. Geophys. Res. 82, 3961–3876.
Ingle, J.C., Jr. (1973). Summary comments on Neogene biostratigraphy, physical stratigraphy, and paleo-oceanography in the marginal northeastern Pacific Ocean, in Initial Reports of the Deep Sea Drilling Project 19, U.S. Government Printing Office, Washington, D.C., pp. 949–960.
Poore, R.Z. (1981). Miocene through Quaternary planktonic foraminifera from offshore southern California and Baja California, in Initial Reports of the Deep Sea Drilling Project 63, U.S. Government Printing Office, Washington, D.C., pp. 415–436.
Poore, R.Z., and D.Bukry (1979), Preliminary report on Eocene calcareous plankton from the eastern Gulf of Alaska continental slope, U.S. Geol. Surv. Circ. 804-B, pp. B141–B143.
Thierstein, H.R., and W.H.Berger (1978). Injection events in ocean history, Nature 276, 461–466.
Vergnaud-Grazzini, C. (1979). Cenozoic paleotemperatures at site 398, eastern North Atlantic: Diagenetic effects on carbon and oxygen isotopic signal, in Initial Reports of the Deep Sea Drilling Project 47, U.S. Government Printing Office, Washington, D.C., pp. 507–511.
Wolfe, J.A. (1971). Tertiary climatic fluctuations and methods of analysis of Tertiary floras, Palaeogr. Palaeoclim. Palaeoecol. 9, 27–57.
Wolfe, J.A. (1977). Paleogene floras from the Gulf of Alaska region, U.S. Geol. Surv. Prof. Pap. 997, 108 pp.
Wolfe, J.A. (1978). A paleobotanical interpretation of Tertiary climates in the northern hemisphere, Am. Sci. 66, 694–703.
Wolfe, J.A. (in press). Climatic significance of the Oligocene and Neogene floras of northwestern United States, in Evolution, Paleoecology, and the Fossil Record, K.J.Niklas, ed., Praeger, New York.
Wolfe, J.A., and T.Tanai (1980). The Miocene Seldovia Point flora from the Kenai Group, Alaska, U.S. Geol. Surv. Prof. Pap. 1105, 52 pp.





