
Lighting Up the Mechanome
MATTHEW J. LANG
Massachusetts Institute of Technology
Cambridge, Massachusetts
Genomics and general “omics” approaches, such as proteomics, interactomics, and glycomics, among others, are powerful system-level tools in biology. All of these fields are supported by advances in instrumentation and computation, such as DNA sequencers and mass spectrometers, which were critical in “sequencing” the human genome. Revolutionary advances in biology that have harnessed the machinery of polymerases to copy DNA or used RNA interference to capture and turn off genes have also been critical to the development of these tools. Researchers in biophysics and bio-mechanics have made significant advances in experimental methods, and the development of new microscopes has made higher resolution molecular- and cellular-scale measurements possible.
Despite these advances, however, no systems-level “omics” perspective—the “mechanome”—has been developed to describe the general role of force, mechanics, and machinery in biology. Like the genome, the mechanome promises to improve our understanding of biological machinery and ultimately open the way to new strategies and targets for fighting disease.
Many disease processes are mechanical in nature. Malaria, which infects 500 million people and causes 2 million deaths each year, works through a number of mechanical processes, including parasite invasion of red blood cells (RBCs), increased RBC adhesions, and overall stiffening of RBCs. As the disease pro-
gresses, it impairs the ability of infected cells to squeeze through capillaries, ruptures RBC membranes, and releases newly minted malaria parasites.1
Cancer, another leading cause of death, also progresses in many mechanical ways, such as the machinery of viral infection, which is responsible for hepatitis B, human papilloma virus, and related cancers. In addition, metastasis involves the detachment of tumor cells from the initial growth site, migration, circulation, and subsequent invasion through reattachment to new locations.
Intracellular changes in cancerous cells bring about cell growth and proliferation, cell motility, and migration. These processes are currently targeted by Taxol, a potent chemotherapy drug that impedes the progression of cancer by stabilizing microtubules in a polymerized state, thus shutting down the machinery of cell division.
The human genome is contained in approximately 3 billion base pairs of DNA. If the pieces of DNA in our cells were connected end-to-end, the chain would be roughly as long as one’s outstretched arms. Compared to other cellular filaments, DNA is quite flexible. To fit into cells, it is compacted in highly ordered structures through multiple hierarchical levels.2 In addition to the conventional Watson and Crick base pairs, DNA forces and interactions include histone protein-DNA nucleosome complexes, chromatin fibers, chromatids, and, ultimately, 46 chromosomes. In general, our biological structures have many levels of structural hierarchy and are highly organized.
MOLECULAR MACHINERY
From our DNA sequence, biological motors, including DNA and RNA polymerases and the ribosome, make RNA, and ultimately proteins, through coupled mechanical and chemical steps. This impressive machinery copies and translates with great accuracy. The ribosome, a huge motor spanning 20 nanometers (nm), which is actually composed of about 65 percent ribosomal RNA, assembles proteins from messenger RNA. Many antibiotics target the critical machinery of ribosomal translation in bacteria.
Other molecular motors, such as myosins and kinesins, run on different cytoskeletal tracks—actin filaments and microtubules, respectively.3 Myosins, which are responsible for the contraction of skeletal and cardiac muscle, contribute to cell
migration and cell-division processes, such as contractile ring closure. Similarly, kinesins, which are important in transport and cell division, are small, processive motors that run toward the plus (fast polymerizing) end of microtubules. In some cases, such as the kinesins that carry vesicles containing neurotransmitters along the sciatic nerve axon, this progress takes place over the length scale of about a meter and is a critical mechanism for cargo delivery.4
The design of biological motors can be classified by cataloging the motor’s general structural features, fuel type, stepping distance, stall force, and other mechanical parameters. Detailed measurements of the motility cycles and underlying mechanisms for motility also provide information about how these mechanisms work.5 Ultimately, “sequencing” the mechanome will lead to the discovery of the design features of biological motors in general, enabling us to catalogue them and outline the rules that govern their behavior.
Comparisons of the family trees of kinesins and myosins reveal a range of structural forms, including single-, double-, and tetramer-headed members of various lengths connecting the motor and cargo-binding domains. They also share some structural elements at the molecular level, such as the binding pocket for adenosine triphosphate (ATP) (Vale, 1996), and both are fueled by hydrolysis of ATP. Another example of a common motor design is in AAA+ motors, which are similarly structured but run on a variety of track types (e.g., microtubules, DNA, and peptides) and are involved in a number of critical cellular processes (Ogura and Wilkinson, 2001). Nature, it appears, copies and pastes design elements for motors with similar roles and physical constraints. Despite different tracks and overall purposes for motility, all of the designs of common biological machines have been optimized.
CELLULAR MACHINERY
Cellular machinery (e.g., the processes of cell division, wound healing, and migration) can also be classified by general design principles. Forces originating from many sources (e.g., biological motors, polymerization of filaments, molecular conformational transitions, and fluid pressure) can act on structures, including cytoskeletal filaments, membrane barriers, receptor ligand linkages, and
protein-complex adhesion sites. These interactions typically occur in cycles, such as migration, extension processes, adhesion, contraction, and detachment.
Highly tuned control over these forces and structures is orchestrated through mechanical and biochemical signals that initiate and terminate motility cycles. Mechanotransduction, a critical aspect of control, refers to the process whereby cells actively respond to mechanical signals through changes in morphology, signaling, and biochemical response.
Migration of the neuronal growth cone exhibits a range of interesting cell machinery. The main core structure of the growth-cone axon consists of bundled microtubules. At the tip of this bundle, actin filaments are structured in both bundled finger-like spikes (fillo-podia) and web-like flattened sheets (lamellipodia). Actin filaments can be arranged in a variety of higher level structures coordinated by a series of actin-binding proteins (ABPs) (Figure 1).
The overall structures of ABPs vary widely, but many have a common actin-binding domain. In cell division, an actin-based ring-like bundle contracts to separate the cell into progeny cells. The design elements of this process, called cytokinesis, appear to be cut and pasted. For example, in cell division of higher plants, a similar process generates a microtubule-based structure at the cell-division locus (Jurgens, 2005). Even bacteria contain tubulin and actin homologues (FtsZ and MreB, respectively) that form higher level structures and machinery that resembles their counterparts, such as microtubules (MreB) and
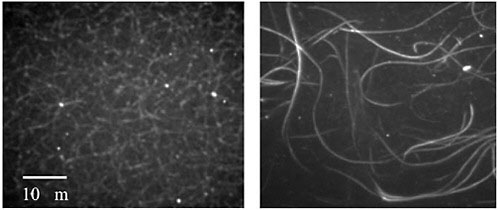
FIGURE 1 The images above show networks of actin filaments organized by different actin-binding proteins (ABPs). The ABP on the left, consisting of filamin, forms a meshwork arrangement of filaments. The ABP on the right, α-actinin, organizes actin filaments into bundles. Despite differences in network morphology, filamin and α-actinin have similar actin-binding domains. Source: Photos courtesy of Hyungsuk Lee and Jorge Ferrer, in collaboration with Prof. Roger Kamm’s laboratory, Massachusetts Institute of Technology.
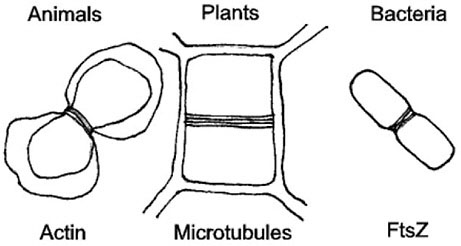
FIGURE 2 This cartoon shows similar stages of cell division in animal cells, plant cells, and bacteria. In all three, a common structural end point is achieved by ring formation through different types of filaments: actin filaments, microtubules, and FtsZ, respectively.
the formation of the cytokinetic ring in bacteria (FtsZ) (Erickson, 1998; van den Ent et al., 2001) (Figure 2).
TISSUE AND HIGHER LEVEL MACHINERY
Mechanical processes are critical, not only at the molecular level, but also at the tissue level and higher levels. In fact, the mechanics of the extracellular environment is an important factor in determining its fate. Stem cells plated on substrates of different stiffnesses differentiate into different cell types, taking on properties of tissues similar to their mechanical environments. For example, mesenchymal stem cells grown on a matrix with elasticity resembling that of tissues such as 1 kPa brain, 10 kPa muscle, and 100 kPa bone differentiate into cells resembling neurons, muscles, and osteoblasts, respectively (Engler et al., 2006).6
The external environment of a cell contains many organized mechanical structures. A typical extracellular matrix is composed of cable-like collagen molecules, rubber band-like elastin molecules, and sponge-like proteo-glycans. So, for example, tendons contain collagen organized into a bundle form with many levels of hierarchy, including mineralization. Tendons are designed to withstand
low strain and fatigue and can sustain many cycles of loading and unloading over a long period of time. Tendon failure is a common sports injury.
In contrast to a tendon, the dense collagen network of the chorioamnion membrane surrounding a fetus is designed to eventually fail. Early failure in this structure is responsible for one-third of all premature births (Oyen et al., 2005).
Mechanical forces may also affect the organization of structures. Bone requires mechanical stimulation to maintain density, and collagen stress fibers in bone align in the direction of maximal load. Higher level mechanical systems include musculoskeletal, respiratory, cardiovascular, lymphatic, and integumental systems.
MEASUREMENTS AT THE MOLECULAR LEVEL
With the advanced instrumentation and experimental methods described below, we are poised to measure the mechanome at the molecular level. For example, force microscopy, in which controlled loads are applied to structures while they are tracked with nanometer-level resolution, can be used to measure mechanical transitions in unfolding protein structures, the rupture of individual molecular interactions, and the motility of biological motors.
A huge range of forces are required to make these measurements. At the molecular level, a balance is obtained between stabilizing an interaction against thermal energy (kT) and maintaining the ability to break an interaction through transition-state barriers of a few kT. Given that kT is in the range of 4 pN-nm (or 4.1×10−21 J),7 and typical transition distances are on the length scale of nanometers, forces ranging from 1 pN to a few tens of pN are ideal for probing molecular machinery.
ATP hydrolysis, the energy source for many biological motors, provides about 20 to 25 kT of energy. Unfolding proteins requires a force of tens to hundreds of pN. Furthermore, because cell-level machinery may involve many molecular interactions, force magnitudes on the scale of nN and more are sometimes required.
A broad range of complementary force-probe methods can meet these force requirements. These include the atomic force microscope (AFM), micropipette manipulation, magnetic traps, microfabricated cantilevers, and optical traps (Suresh, 2007). Combinations of these force probes, such as combined optical trapping and magnetic-particle manipulation, can extend the force range. The optical trap, which has been a mainstay technique in single-molecule biophysics research, is an ideal tool for probing the molecular interactions of biological motors and receptor-ligand interactions.
Fluorescence microscopy is another powerful method of observing single
molecules. With advances in this technology, it can be used to track fluorophore-labeled biological motors with nanometer-level resolution (Yildiz et al., 2004). Energy-transfer methods using pairs of fluorophores (FRET) provide a fluorescence-based ruler with a 1/r6 dependence,8 on the length scale of nanometers, which is useful for monitoring conformational changes.
Recently, optical traps and FRET have been combined on a single platform (Tarsa et al., 2007). The combination marries the power of mechanical interrogation and control of structures with the ability to observe them with fluorescence. This combined method has been demonstrated using a model system consisting of a DNA-based hairpin that can be opened and closed with about 15 pN of force from the optical trap (Figure 3). A FRET pair placed at the base of the hairpin reported whether the hairpin was open or closed through high and low FRET states, respectively. Many additional advances, such as light-based methods of laser scissors, photobleach recovery, triggering, uncaging, and time-resolved measurements, can be integrated to produce powerful new tools.
Significant advances have also been made in assay technologies, molecular biology, and linkage chemistry. Molecular biology strategies, such as the encodeable green fluorescence protein (GFP), which can genetically label proteins, have advanced the field, as have new probes, such as enhanced organic dyes and quantum dots, which have enabled observations of single molecules over extended periods of time. Molecular biology strategies can also be used to create physical handles for the manipulation of molecules, thus exploiting, for example, the aptamer interactions and tethers based on the M13 phage (Khalil et al., 2007).
CONCLUSION
With the tools and advances in assays I have described, we are beginning to probe nature’s machinery. Ultimately, we hope to gain a quantitative understanding of molecular and cellular machinery and to measure the forces and strength of the mechanical steps that underlie the mechanome. Once we understand the physical rules that govern biological systems and can measure nature’s machinery, we should eventually be able to “sequence” the mechanome.
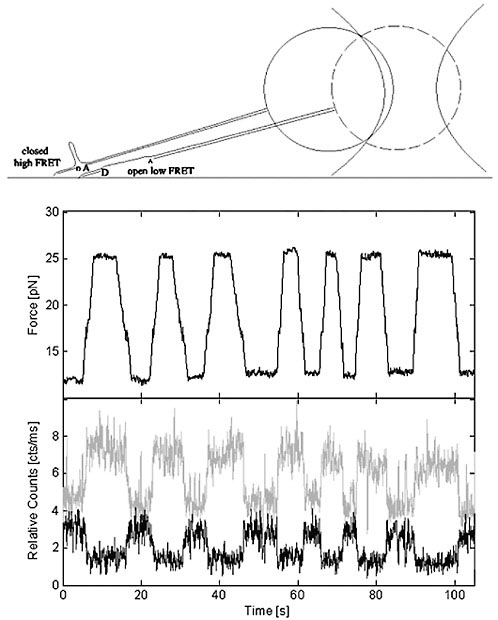
FIGURE 3 The upper frame is a cartoon showing the experimental design with open and closed states of a DNA hairpin complex controlled with force from an optically trapped bead. A donor and acceptor FRET pair is placed at the base of the hairpin to monitor whether or not it is open or closed. The middle frame shows how force on the hairpin is repeatedly transitioned from low to high with hairpin opening occurring at approximately ~18pN. The lower frame shows photon counts with donor (upper) and acceptor (lower) FRET signals changing simultaneously with the opening and closing of the hairpin.
REFERENCES
Engler, A. J., S. Sen, H. L. Sweeney, and D. E. Discher. 2006. Matrix elasticity directs stem cell lineage specification. Cell 126(4):677-689.
Erickson, H. P. 1998. Atomic structures of tubulin and FtsZ. Trends in Cell Biology 8(4):133-137.
Jurgens, G. 2005. Plant cytokinesis: Fission by fusion. Trends in Cell Biology 15(5):277-283.
Khalil, A. S., J. M. Ferrer, R. R. Brau, S. T. Kottmann, C. J. Noren, M. J. Lang, and A. M. Belcher. 2007. Single M13 bacteriophage tethering and stretching. Proceedings of the National Academy of Sciences 104(12):4892-4897.
Ogura, T., and A. J. Wilkinson. 2001. AAA(+) superfamily ATPases: Common structure-diverse function. Genes to Cells 6(7):575-597.
Oyen, M. L., R. F. Cook, T. Stylianopoulos, V. H. Barocas, S. E. Calvin, and D. V. Landers. 2005. Uniaxial and biaxial mechanical behavior of human amnion. Journal of Materials Research 20(11):2902-2909.
Suresh, S. 2007. Biomechanics and biophysics of cancer cells. Acta Biomaterialia 3(4):413-438.
Tarsa, P. B., R. R. Brau, M. Barch, J. M. Ferrer, Y. Freyzon, P. Matsudaira, and M. J. Lang. 2007. Detecting force-induced molecular transitions with fluorescence resonant energy transfer. Angewandte Chemie (International edition in English) 46(12):1999-2001.
Vale, R. D. 1996. Switches, latches, and amplifiers: Common themes of G proteins and molecular motors. Journal of Cell Biology 135(2):291-302.
van den Ent, F., L. A. Amos, and J. Lowe. 2001. Prokaryotic origin of the actin cytoskeleton. Nature 413(6851):39-44.
Yildiz, A., M. Tomishige, R. D. Vale, and P. R. Selvin. 2004. Kinesin walks hand-over-hand. Science 303(5658):676-678.










