
INTRODUCTION
An ecosystem approach to fisheries management is “one that is geographically specified, adaptive, takes account of ecosystem knowledge and uncertainties, considers multiple external influences, and strives to balance diverse societal objectives” (Sissenwine and Murawski, 2004). The ecosystem approach recognizes the effects of fisheries on the ecosystem and of ecosystem state and variability on the fisheries (Crowder et al., 2008). Much has been written about what elements should be included in such an approach since the United States, together with many other countries, embraced an ecosystem approach to fisheries management (Pikitch et al., 2004; Murawski, 2007).
Section 207 of the Sustainable Fisheries Act (SFA) provided initial guidance on inclusion of ecosystem principles in the management of the nation’s fisheries.1 The decision to include this guidance was based on the perceived realization that traditional management measures were insufficient to ensure sustainable fisheries. The basic premise is that fished stocks form essential components of complex marine ecosystems that must be well understood to be effectively managed. Guidelines recognize that multiple interdependent relationships exist among stocks, their fisheries, and the ecosystem in which they reside. This section of the SFA mandated formation of the Ecosystems Advisory Panel of the National Marine Fisheries Service, which was tasked with reviewing the progress toward incorporation of ecosystem principles in Fishery Management Plans (FMPs).2 The resultant Panel report (NOAA, 1999) specified the need to better account for and minimize bycatch and discard of fish, identify and take measures to protect essential fish habitat, and determine the effects of fishing on the environment.
Although consideration of factors beyond the single-species dynamics is clearly a sound objective, the details of how to do this are still a subject of ongoing research, and a variety of approaches are being pursued. Much work is being conducted by the U.S. Fisheries Science Centers to incorporate ecosystem considerations into fisheries management (Hollowed et al., 2011). Approaches include incorporating indices of environmental and biological conditions into stock assessment projections (Hare et al., 2010), using multi-species and food-web models to assess the effects of harvesting strategies (Link et al., 2011), and investigating new forecasting methods (Deyle et al., 2013). These models could be used to support rebuilding efforts, but they will not be comprehensively reviewed in this report.
In the context of fish stock rebuilding, changes in the ecosystem can alter rebuilding rates and the target biomass level to which an overfished species should be rebuilt. The most direct ecosystem consideration for rebuilding is that “MSY stock size (BMSY) means the long-term average of the stock or stock complex…that would be achieved by fishing at FMSY,” where MSY (Maximum Sustainable Yield) is defined “under prevailing ecological, environmental conditions.”3The challenge is to define these MSY-based reference points, given background levels of variability, and to adjust them as necessary in response to ecosystem changes. This chapter is structured around the main ecosystem considerations for fisheries management, with sections on climate, habitat, and ecological interactions. It continues with a discussion of the possibility of incorporating ecosystem factors into rebuilding plans and ends with findings and conclusions.
_________________
1 Sustainable Fisheries Act, Pub. L. No. 104-297 § 207, 110 Stat. 3559, 3621 (1996).
2 Sustainable Fisheries Act, Pub. L. No. 104-297 § 207, 110 Stat. 3559, 3621 (1996).
3 50 C.F.R. 600.310 (e)(i) (2009).
ASSUMPTIONS ABOUT POPULATION STRUCTURE AND DYNAMICS
It is important to consider ecosystem effects on population dynamics that are implicit rather than explicit. Density-dependent processes are expected to reduce the population growth rate as depleted populations rebuild. Density dependence is integral to production models and is included in rebuilding projections when a stock-recruitment model is used. Individual growth rates can also vary with population density and the amount of available food (see discussion on ecological interactions, below). As an example of changing reproductive potential, the growth rate of Georges Bank haddock (Melanogrammus aeglefinus) declined following recruitment of the very large 1999 and 2003 year classes (Brodziak et al., 2008). Density-dependent growth is typically not considered in population projections, but, where it occurs, it affects the calculation of biological reference points such as BMSY and FMSY.
Rebuilding plans require projections of stock dynamics, which are sensitive to assumptions about the future state of the ecosystem. The population models commonly used to project stock rebuilding are generally single species (i.e., no interactions among species), assume continuation of historical conditions in the ecosystem (including variability) into the future (i.e., stationarity assumption), and calculate the biomass reference points under stable equilibrium assumptions. These assumptions may oversimplify the problem; recent observations suggest more complex dynamical behavior operating in fishery ecosystems (Box 5.1).
In particular, natural populations can exhibit dynamical behaviors broadly described as nonlinear, including multiple equilibria (regime shifts), limit cycles, and chaotic dynamics (May, 1973). More generally, nonlinear dynamics simply means that population behavior depends on ecosystem state. Indeed, state-dependence is how nonlinearity is measured in ecological time series (S-maps, Sugihara, 1994); it implies that ecosystem effects must be studied synergistically, not one factor at time. Nonlinear dynamics can be driven by fishing, environmental variability, or trophic interactions (Steele and Henderson, 1984; Anderson et al., 2008). There is growing evidence of nonlinear dynamics in fish populations (Dixon et al., 1999; Glaser et al., 2011), as well as growing consensus that ecosystem and multi-species effects are important. The existence of nonlinear dynamics has profound implications for the way we think about fisheries ecosystems, how we model fish populations, and ultimately our expectations for stock rebuilding.
Although addressing nonlinear dynamics and complex ecosystems can appear to be daunting, new empirically based methods that use time series data for reconstructing complex ecosystem dynamics may provide a practical simplification for understanding the role of interspecific interactions and environmental influences on population dynamics (Sugihara and May, 1990; Sugihara et al., 2012). This empirical time-series-based paradigm emphasizes dynamics rather than static statistical relationships and thereby avoids issues related to so-called “mirage correlations” that appear then disappear through time (Planque et al., 2010) (Box 5.2). This approach is currently being examined by several National Marine Fisheries Service (NMFS) science centers (Southwest Fisheries Science Center, Northeast Fisheries Science Center, Southeast Fisheries Science Center) and may provide guidance for a next-generation dynamic management paradigm based on forecasting production (Deyle et al., 2013).
CLIMATE CHANGES AND SHIFTING BASELINES
Environmental variability affects fish population dynamics on temporal scales ranging from interannual to decadal and millennial (Cushing, 1982). According to the National Standard 1 Guidelines (NS1G), “If environmental changes affect the long-term reproductive potential of the stock or stock complex, one or more components of the status determination criteria (SDC) must be respecified.”4 The NS1G require a high standard for changing SDC so that it will not undermine the statutes that mandate an end to overfishing.
As an example of changing reproductive potential, recruitment of Atlantic croaker (Micropogonias undulatus) depends on over-wintering temperature in the estuaries where juveniles rear. Hare et al. (2010) fit a temperature-dependent stock-recruitment model to data for Atlantic croaker that greatly reduces the unexplained recruitment variability (Figure 5.1). This statistical relationship allows for estimation of biological reference points such as BMSY with more precision, as well as for projections of levels of population abundance and sustainable harvest under assumed future temperature conditions. Similar models with environment-dependent stock-recruitment relationships are being formulated for species with rebuilding plans, such as winter flounder, Psuedopleuronectes americanus (NEFSC, 2011). However, these models may fail under environmental conditions other than those used to fit the model; alternate approaches may be needed (Box 5.2).
“Long-term environmental changes affect both the short-term size of the stock or stock complex and the long-term reproductive potential of the stock or stock complex.”5With climate change, environmental conditions into the future are assumed to change in a smooth progression from decade to decade. A common response of coastal and marine finfish species to climate change is a shift in their geographic distribution so that they maintain themselves in preferred temperature conditions. Fish species may shift their geographic distributions poleward or to deeper water (Nye et al., 2009). As species distributions shift, it may become necessary to change stock boundaries and definitions.
_________________
4 50 C.F.R. 600.310 (e)(2)(iii)(B) (2012).
5 50 C.F.R. 600.310(e)(2)(iii).
BOX 5.1
A Simplifying Paradigm for Complex Ecosystem Dynamics
Population models used in ecology and fisheries, developed in the mid 20th century, have stability and equilibrium as foundational concepts. Although mathematically tractable and well defined, “stability” and the implication of “equilibrium” in ecological systems began to give way in the 1980s and 1990s to growing evidence that “change” rather than “constancy” is the rule and that nonlinear instability, thresholds, and chaos can be ubiquitous in nature (Sugihara and May, 1990; Grenfell et al., 1998; Scheffer and Carpenter, 2003). Nonetheless, models with stable equilibria and MSY remain the benchmark for fisheries assessment and management (Quinn and Collie, 2005). These equilibria are the basis for calculating the biological reference points that are used to determine fish population status and the targets for rebuilding overfished stocks.
A new paradigm explicitly recognizes the growing evidence from field measurements on natural populations that nonlinear complexity and instability are ubiquitous, and that a stable equilibrium (even multiple equilibria), though simple and manageable, is likely to be the idealized exception rather than the rule. This new paradigm views fisheries as dynamic, complex systems of interdependent variables, represented by simple non-parametric models that depend on a data-driven construct, rather than on assumptions of stationary variability and stable equilibrium.
The Simplifying Approach
Complex systems are typically modeled using differential (or difference) equations that describe the transition through time between different states of the system. Each state is represented as a vector of state variables, x(t) (e.g., abundances of foxes, rabbits, and grasses, temperature, stock levels, prey levels, etc.), and the set of all states through which a dynamic system transitions forms a geometric construct known as an attractor manifold, M. The manifold describes how ecosystem state variables relate to each other through time—a dynamic version of Huchinson’s n-dimensional niche. If there are rules governing ecosystem changes (i.e., if ecosystems are not purely random), then there is an attractor manifold to be uncovered (Box 5.1, Figure 1) (Deyle and Sugihara, 2011). Attractor manifolds determine (express) relationships among variables and can be obtained simply by re-plotting the time series data. Constructing manifolds empirically from ecological time series is the basis of the approach. Box 5.1 Figure 1 illustrates the following three core ideas:
- Nonlinear State Dependence (panel a). If there is an attractor manifold M that is not flat (a hyperplane), then relationships between variables will depend on system state (e.g., Y1 and Y3 are positively correlated at some times and negatively associated at other times). Baltic Sea fisheries, for example, exhibit radically different dynamic control regimes (top-down versus bottom-up) depending on the threshold abundance of planktivores, causing the correlations between fish and zooplankton to change sign (Casini et al., 2009). Thus, if a fishery exhibits nonlinear state dependence, then fish populations, fishing pressure, and environmental effects should be considered together (Deyle et al., 2013).
- Time Series as Observation Functions (panel b). A time series {Yi} is a projection of the dynamics occurring on M. More generally, the Yi are observation functions of the dynamics on M. The Yi may be fundamental coordinates or they may be any function (e.g., rotations or linear combinations of the original Cartesian coordinates) that maps points in M to time series observations. The key insight is that ecological time series can appear complex because they are projections into one dimension of dynamics occurring in higher dimensions.
- State Space Reconstruction and Takens Theorem (panel c). If all the variables and equations governing an ecosystem were known, then we could construct the attractor manifold by direct simulation. In fact, it is possible to reconstruct the manifold empirically, if we only had time series for all the variables. This manifold would be an empirical expression of all of the dynamic relationships among variables observed in the data. However, in practice we may only have time series information about one species. A key result from dynamical system theory—the Takens embedding theorem—proves that one can reconstruct the dynamical attractor for a system from data in the form of lagged samples of just one variable, such as Yi. Thus, state space reconstruction (SSR) is a method to recover an approximation of M from time series. This is illustrated in panel c, where the shadow manifold M1’ is constructed using lags of time series {Y1}. The reconstruction captures the essential topology and dynamics of the original system. Further refinements include (1) using multivariate reconstructions that are more mechanistic (Dixon et al., 1999; Deyle and Sugihara, 2011), (2) identifying and incorporating stochastic environmental forcing, and (3) exploring environmental scenarios (Deyle et al., 2013).
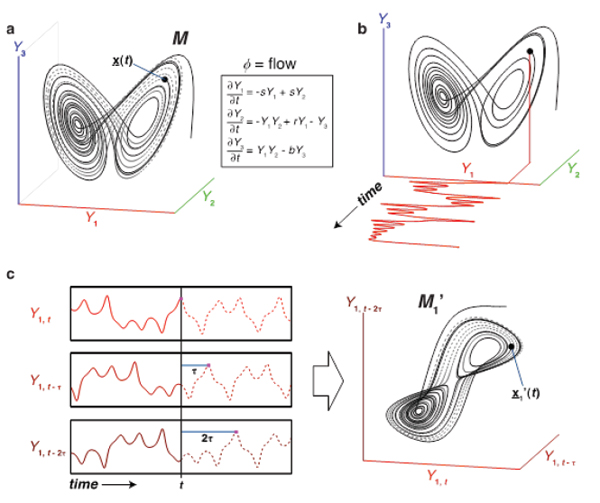
BOX 5.1, FIGURE 1 (a) The Lorenz butterfly attractor example. The attractor manifold M is the set of states through which the system progresses. x(t) is the state of the system at time t, and the dynamics are defined by the Lorenz equations. (b) A time series is simply a projection of the system states from M to a coordinate axis (Y1 is a state variable of the system). The manifold can be constructed from the component time series. (c) Following Takens theorem (Takens, 1981), lags of the time series {Y1,} can act as coordinate axes to construct a shadow manifold M1’, which maps 1:1 to the original manifold M (the visual similarity between M1’ and M is apparent). These shadow manifolds can be used for ecosystem-based prediction, identifying causal variables, and much else.
SOURCE: Reproduced from Sugihara et al., 2012; see also the supplemental animations.
This new paradigm has several important implications for fisheries science in general and for stock rebuilding in particular. The nonequilibrium nature of these models challenges the basis of calculating biological reference points, particularly biomass reference points such as unfished stock size (B0) and BMSY. Although ecosystems may have tens to thousands of interacting variables, their essential dynamics at any time may involve relatively few key variables or dimensions. Several studies indicate that the relevant ecosystem dimension for certain fish species is often relatively low (involving from three to eight dimensions), and ecosystem dimensions also demonstrate relatively high predictability (Dixon et al., 1999; Hsieh et al., 2005). Although forecasting skill may be high in the short term, the ability to make medium- and long-term forecasts is limited by unstable dynamics and forcing by a stochastic environment (Glaser et al., 2013).
BOX 5.2
Mirage Correlation the Bane of Ecosystem Science
Many are familiar with Berkeley’s 1710 dictum “correlation does not imply causation.” Less well appreciated is the fact that in nonlinear systems the converse “causation does not imply correlation” is also true. Ecosystems are particularly perverse on this issue by exhibiting mirage correlations—associations among variables that spontaneously come and go or even switch sign (Sugihara et al., 2012). This common behavior of nonlinear systems can create conceptual sand traps that distract research effort, continually causing us to rethink relationships that we thought we understood. This is particularly relevant for investigating causative environmental factors (e.g., ocean temperatures) affecting fish production.
The alternation of Pacific sardine, Sardinopssagax, and northern anchovy, Engraulismordax, in the California Current ecosystem is perhaps the most classic example of attempts to understand ecosystem effects on pelagic fish populations. Jacobson and MacCall (1995) found a significant relationship between sardine recruitment success and sea surface temperature (SST) using a generalized additive model (GAM). Based on this relationship, the Pacific Fishery Management Council modified the sardine management plan to afford extra protection when SST is unfavorable (PFMC, 1998). However, the GAM does not readily accommodate the interacting effects of explanatory variables. The state-dependence of recruitment-environment relationships suggests that methods besides static linear correlation analysis are required.
Put simply, the problem has been the use of static linear methods to investigate a nonlinear dynamical system. In trying to understand environmental factors driving nonlinear fishery ecosystems, we are interested in how variables affect each other dynamically (causally). Based on the ideas presented in Box 5.1, convergent cross mapping (CCM) is a recent tool that leverages the idea from Takens theorem that variables in a dynamic system share information about each other (Sugihara et al., 2012). Thus, if two variables are dynamically connected (influencing each other’s time series), then it is possible to predict the state of one from the other, and CCM tests for this. In the sardine-anchovy example, application of CCM showed that SST affected both sardine and anchovy, but there was no interaction between sardine and anchovy (Box 5.2, Figure 1).
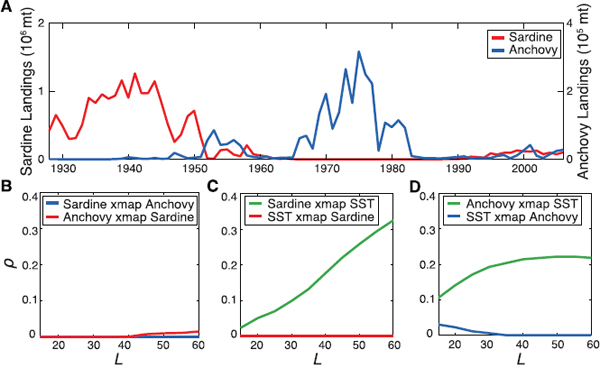
BOX 5.2, FIGURE 1 Detecting dynamic causation in a pelagic fishery.
NOTES: (A) California landings of Pacific sardine (Sardinos sagax) and northern anchovy (Engraulis mordax), (B) convergent cross mapping (CCM) of (or lack thereof) sardine versus anchovy, (C) sardine versus SST (Scripps Pier), and (D) anchovy versus SST (Newport Pier). The ability of CCM to predict the states of the forcing variable from the manifold of the forced variable is measured by the correlation coefficient between the predicted and observed states (p), which increases with time series length L, as points in the manifold become closer. This analysis shows that sardines and anchovies do not interact with each other and that both are forced by temperature. “Sardine xmap Anchovy” means cross mapping anchovy from sardine, etc.
SOURCE: Reproduced from Sugihara et al. (2012).
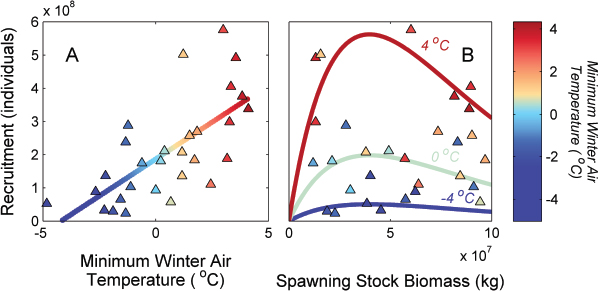
FIGURE 5.1 Relationship between Atlantic croaker (Micropogonias undulatus) recruitment, temperature, and spawning stock size.
NOTES: (A) Relationship between recruitment and minimum winter air temperature in Virginia. (B) Environment-dependent stock-recruitment relationship illustrated at three temperature levels, -4, 0, and +4°C.
SOURCE: Reproduced with permission from Hare et al., 2010.
Biological reference points such as BMSY, which are based on demographic parameters, become moving targets with changing climate. Productivity may increase, as in the Atlantic croaker example, or it may decrease as seems to be the case of winter flounder at its southern range. The effect of climate is expected in cases such as winter flounder, but the relationship between climate and productivity is not well enough understood to make the types of long-term projections that are required for rebuilding programs (NEFSC, 2011). However, with better understanding of environmental trends and coupling of the environment and fishing to stock production it is possible to evaluate management strategies under climate-change scenarios (e.g., see Ianelli et al., 2011; Punt, 2011).
A primary determinant of productivity in marine fish populations is the per capita recruitment rate (Myers et al., 1999). The ratio of recruits per spawner can therefore be used to identify persistent shifts in productivity. Acadian redfish (Sebastes faciatus) had a period of increased productivity from 1980 to 2000, which contributed to its rebuilding (Appendix C, Figure C.1).6 In contrast, five stocks from the New England and Mid-Atlantic regions experienced persistent downward shifts in recruits per spawner during the 1990s and 2000s: Mid-Atlantic black sea bass, Gulf of Maine haddock, scup, summer flounder, and Southern New England/Mid Atlantic (SNE/MA) winter flounder (Appendix C). These shifts help to explain the declining biomass of Gulf of Maine haddock (Figure 3.18) and lack of rebuilding of SNE/MA winter flounder (Appendix C, Figure C.58), despite the target fishing mortality (FACL) being reduced to as close as possible to zero since 2009, resulting in FACT=0.24 FMSY in 2012 (Figure 3.7). The classification of stock status as overfished or not, and in turn the requirement for a rebuilding plan, is uncertain when there is an appearance of a change in productivity regime, as exemplified by the recent change in status of the SNE/MA yellowtail flounder (see Chapter 4). The per capita recruitment of SNE/MA yellowtail flounder has varied around its median level, apart from the very strong 1987 year class (Appendix C, Figure C.63). The decline in recruitment of this stock was mainly due to low mature biomass, not a shift in productivity. This interpretation does not support the recent decision to lower the rebuilding target for this stock by favoring a hypothesis of regime shift (see Chapter 4).
Changes due to decadal regime shifts occur on time frames intermediate between the short term (i.e., internannual) and the longer term. MSY-based reference points and rebuilding targets can be specified for the current regime, while recognizing the possibility that the ecosystem may switch to a different regime within the time period covered by the rebuilding plan. Regime shifts are challenging because they are difficult to predict or characterize beyond recognizing that some rapid large-scale, system-wide change occurred. Alaska crab stocks were at high abundance levels during the 1960s and 1970s, and then they suffered steep declines in the early 1980s (Kruse et al., 2010). Although overharvesting contributed to these declines, several stocks have not recovered, even with low or no harvesting. Follow-
_________________
6 To view any of the time series plots (Figures C-1 to C-64), go to http://www.nap.edu/catalog.php?record_id=18488 and click on “Related Resources.”
ing the implementation of rebuilding plans in 2000, the St. Matthew blue king crab stock recovered beyond BMSY, but the Pribilof Islands blue king crab stock has not rebuilt even though all fishery-related mortality has been essentially zero for a decade (Figure 5.2). Although the mechanisms for lack of rebuilding are unclear (e.g., temperature, predation, lack of large males), the Bering Sea and the Gulf of Alaska appear to have shifted from a regime conducive to crab productivity to a regime more favorable to walleye pollock and other groundfish species.
Regime shifts in marine fish stocks appear to be quite common (Vert Pre et al., 2013), and they create a dilemma for developing and assessing the performance of rebuilding plans. Conditioning rebuilding targets on an earlier, high-abundance regime could make rebuilding unattainable under the new prevailing environmental conditions. This scenario can lead to reductions in F that are so far below FMSY that it will be difficult to reach a target that is set too high within a fixed time frame, as well as to constraints on other species in a mixed-stock fishery (see Chapter 4). Conversely, species rebuilding targets based on a recent period of low productivity could forgo larger potential harvests if stocks could, in fact, rebuild to their earlier high abundance levels. Also challenging is knowing whether hysteresis or delayed recovery may be operating. Although progress is being made in detecting regime shifts with theoretical models (Scheffer et al., 2009) and experimental lakes (Carpenter and Brock, 2006), it is extremely difficult in practice with relatively short time series of empirical data.
Harvest strategies are needed that perform reasonably well under the alternative hypotheses considered plausible. Constant harvest rate policies, in which the harvest rate corresponds to the average productivity, have been shown to perform well in some simulated cases by allowing biomass to track changes in productivity (Parma, 1990; Walters and Parma, 1996). However, such policies may increase risks in the face of persistent low-productivity regimes. The commonly used “hockey stick” control rules respond to regime shifts by reducing fishing mortality at low biomass levels (Spencer and Collie, 1997). Alternatively, the harvest rate can be adjusted dynamically in response to measured changes in stock productivity (Collie et al., 2012). Harvest control rules that account for regime shifts in recruitment have been investigated for some species that have had rebuilding plans, such as snow crab in the eastern Bering Sea (Szuwalski and Punt, 2013).
Although it is common to attribute population declines exclusively to fishing or exclusively to the environment, in most cases, the observed stock dynamics are probably a combination of fishing and the environment. Furthermore, fishing and environmental effects may interact in ways that are nonadditive (Hsieh et al., 2008; Deyle et al., 2013). Planque
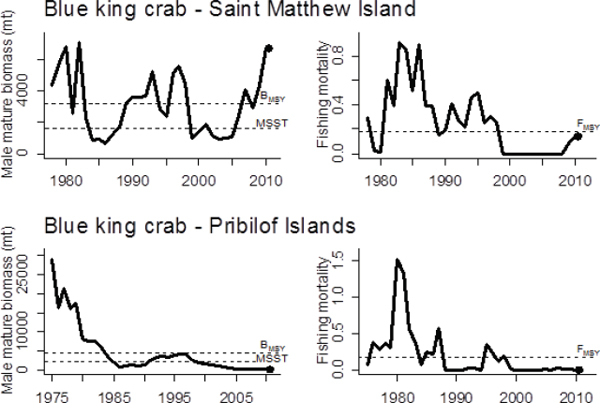
FIGURE 5.2 Biomass and fishing mortality rate of two blue king crab stocks in the Bering Sea: St. Matthew Island and the Pribilof Islands.
SOURCE: Appendix C, Figures C.10 and C.11.
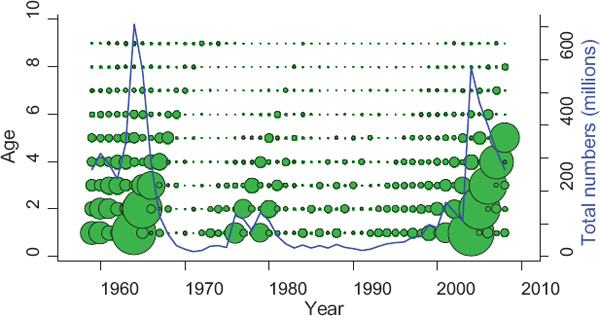
FIGURE 5.3 Age structure of the Georges Bank haddock stock shown in relation to total population size (blue solid line).
NOTES: The area of each bubble is proportional to the numbers at each age in each year from 1959 to 2008. The numbers were estimated with an age-structured stock assessment.
SOURCE: NEFSC, 2008.
et al. (2010) reviewed many ways in which fishing can alter the sensitivity of marine populations and ecosystems to climate. Among these, the alteration of demographic structure is most relevant to stock rebuilding because depleted stocks are likely to have truncated age structure and reduced genetic population structure (Olsen et al., 2004). Populations with truncated age structure are more variable because they are measurably more nonlinear (express greater dependence on ecosystem state) and are more dependent on recruiting age classes (Anderson et al., 2008).
Truncating the age structure may reduce the ability of populations to cope with sequences of poor conditions. A possible consequence of fishing-induced change in age structure may be an increased coupling between recruitment and environmental conditions. Support for this hypothesis comes from the Northeast Arctic cod, for which the correlation between recruitment and ocean temperature strengthened as the population declined and the modal age of the spawning biomass declined from age 13+ to 7 (Otterson et al., 2006). An important implication of this hypothesis for overfished stocks is that, although recruitment may seem to depend only on the chance occurrence of favorable environmental conditions, rebuilding the demographic structure of the population will increase the probability of strong recruitment. More generally, Planque et al. (2010) concluded:
If, as it is agued here, exploitation can affect the way populations respond to climatic forcing, it is likely that recovery to a given population abundance or biomass will not be sufficient to also restore the patterns of population responses to climate. This will require that population characteristics other than biomass (e.g., demographic and spatial structures) also be restored. Another consequence is that statistical climate-population relationships may display recurrent appearance/disappearance sequences, as has also been observed for a number of populations. (See Box 5.2.)
Thus, stocks needing rebuilding may become more sensitive to environmental variation, exhibiting more nonlinear state dependence and variability (Anderson et al., 2008). This sensitivity could affect the rate at which a population can recover, and increased variability may obscure the success of rebuilding. Attaining a biomass target may depend on first restoring the age structure of the stock. For example, the age structure of Georges Bank haddock became truncated following the stock collapse between 1970 and 1995 (Figure 5.3). The expansion of the age structure in the late 1990s preceded very strong year-classes in 1999 and 2004.
HABITAT LOSS AND CARRYING CAPACITY
The importance of fish habitat was recognized in the 1996 Sustainable Fisheries Act (see Chapter 2). This reauthorization of the Magnuson-Stevens Fishery Conservation and Management Act (MSFCMA) defined essential fish habitat (EFH) as “those waters and substrate necessary to fish for spawning, breeding, feeding, or growth to maturity.”7 The
_________________
7 Sustainable Fisheries Act, Pub. L. No. 104-297 § 102(3), 110 Stat. 3559, 3561 (1996).
Regional Fishery Management Councils (RFMCs) were tasked with describing and identifying EFH for each managed species and with minimizing to the extent practicable adverse effects on EFH, all by October 1998. In implementing the MSFCMA, the NS1G specified that “if manmade environmental changes are partially responsible for a stock or stock complex being in an overfished condition, in addition to controlling fishing mortality, Councils should recommend restoration of habitat and other ameliorative programs to the extent possible.”8
The habitat requirements of purely marine fish species have been catalogued through EFH amendments to fisheries management plans. For many of the most important commercially harvested species, EFH is defined by generalized habitat types, benthic substrates, and depth ranges of the different life stages, but rarely indicates specific geographic locations. At a fundamental level, carrying capacity may be limited by available habitat, but it is difficult to determine which habitat attributes are limiting for most species. For example, red snapper in the Gulf of Mexico has a strong association with vertical structures, such as oil and gas rigs; yet, the degree to which the addition of such habitat increases stock productivity or simply spatially redistributes stock biomass remains controversial (Shipp and Bortone, 2009).
Habitat considerations for certain life stages have been well defined because of known reliance of the species on specific habitats and the documented degradation of these habitats. Impacts on freshwater habitat are known to limit the recovery of anadromous species such as Pacific salmon. In the western United States, for example, dams limit upstream migration, eliminating entire watersheds from salmon production. Altered flow regimes affect the thermal habitat, while logging and channelization of streams affect spawning and rearing habitat (NRC, 2004). In the northeast United States, dam removal on the Kennebec River and the installation of fishways on the Acushnet River have restored habitat to anadromous fish (RAE, 2013). Counts of river herring (Alosa spp.) have increased in these rivers since these fish passage improvements were made.
Habitat changes can also occur on broader spatial scales and simultaneously affect multiple species. For example, the coastal salt marsh habitat of Louisiana is well known for supporting the early life stages of many of the Gulf of Mexico’s commercially important stocks (Boesch and Turner, 1984). Coastal wetlands have been lost at a rate between 60 and 100 km2/year, reducing juvenile habitat and likely causing reductions in concomitant productivity of many species (Boesch et al., 1994; Day et al., 2005). There are plans to restore the wetlands of coastal Louisiana (Boesch and Turner, 1984), which would occur over multi-decadal time frames, similar to those of the stock rebuilding plans for long-lived species, and over a broad spatial area. Successful restoration could affect the available habitat for multiple species and thus their productivity. In a study of salt marsh restoration in Delaware Bay, the intertidal fish community composition converged with that in reference marshes within the 9-year study period (Able et al., 2008).
Similarly, various benthic habitats on the continental shelves are essential for feeding and reproduction of exploited species. Changes in the structure, function, or aerial extent of these benthic habitats may affect rebuilding rates. Mobile bottom-fishing gear exerts pervasive effects on benthic habitats on the continental shelf (Collie et al., 2000). The disturbance to benthic communities caused by trawling and dredging has been quantified in numerous studies (NRC, 2002a) and is known to vary with depth, sediment type, and taxa (Kaiser et al., 2006). Bottom fishing can reduce the production of benthic communities (Jennings et al., 2001; Hermsen et al., 2003). Differences in diet composition have been observed in demersal fish species in areas with contrasting levels of bottom fishing disturbance (Smith et al., 2013). Finally, differences in growth rate of plaice, Pleuronectes platessa, were found across a gradient of fishing disturbance in the Celtic Sea (Shepherd et al., 2010). These studies suggest that benthic habitat may limit fish feeding and growth, but results have been mixed, making them difficult to scale to the population level.
Some sensitive habitats have been designated Habitat Areas of Particular Concern (HAPCs) and afforded protection from bottom fishing disturbance, for example in the case of the juveniles of demersal species such as Atlantic cod. As management tools, closed areas protect fish habitat as well as fish stocks (see Chapter 4). For example, increases in the abundance, biomass, and production of benthic epifauna occurred following large area closures on Georges Bank (Collie et al., 2005). The timescales of recovery for habitats range from years in soft sediments to decades and even centuries for hard substrates (NRC, 2002a).
Some environmental and climate effects on fish populations are likely to be mediated through habitat. Climate variability alters fish reproductive habitat (see discussion on climate changes and shifting baselines, above). For example, the cumulative spawning habitat available for sardine and anchovy was evaluated across the California Current by relating shipboard collection with remote-data series of sea surface temperature (Reiss et al., 2008). On the northeast shelf, intensity of the fall phytoplankton bloom has been hypothesized to stimulate benthic productivity and thereby the reproductive contribution and recruitment success of Georges Bank haddock (Friedland et al., 2008). This mechanism may have played a role in rebuilding this haddock stock. The ranges and reproductive habitats of many species are likely to shift with climate change, and such shifts may be amplified by fishing (Hsieh et al., 2008). These shifts include range expansions at the northern edge of species distributions (e.g., summer flounder juveniles rearing in northern estuaries) and contractions at the southern edge of species distributions (e.g., winter flounder declining in Delaware Bay).
_________________
8 50 C.F.R. 600.310 (e)(2)(iii)(C) (2009).
In summary, many species depend on particular habitats to support the growth and survival of specific life stages, suggesting that habitat loss could limit rates of rebuilding. However, in most cases, the relationships between habitat and productivity have not been quantified. If habitat recovers quickly, then it might accelerate stock recovery and increase the success of short-term rebuilding plans. If long term, then habitat recovery may be more important for gradually increasing the carrying capacity of the stock.
ECOLOGICAL INTERACTIONS
How should ecological interactions be considered when formulating rebuilding plans? The total productivity of a fish community is ultimately limited by production at lower trophic levels. Food-web models implicitly assume that consumers compete for limited food resources (Collie, 2001). When fish stocks are depleted, their prey species are consumed by other predators that may increase in abundance, thereby limiting availability of the common prey. It may then be difficult to simultaneously rebuild all overfished species to their single-species BMSY levels without reductions in other consumer species. Food-web models of the Georges Bank fish community suggest that rebuilding the principal groundfish species (e.g., cod, haddock, yellowtail flounder) to their BMSY levels would require restructuring of the fish community and repartitioning of energy within the food web (Collie et al., 2009; Link et al., 2011).
Population growth can be limited by prey abundance at critical life stages. Recent work has related population growth of Atlantic cod to trends in zooplankton abundance. In particular, two copepod taxa, Pseudocalanus spp. and Centropages typicus, which are nutritionally important for larval cod, have declined in spatially discrete areas where cod populations have not responded to stock rebuilding measures (Friedland et al., 2013). Sherwood et al. (2007) related the reproductive condition of medium-sized cod (30-69 cm) off Newfoundland and Labrador to diet. Cod off southern Newfoundland with more pelagic diets had higher somatic condition, lipid stores, and spawning potential than more northerly cod, which preyed almost entirely on shrimp (Pandalus borealis). The authors hypothesized that rebuilding Newfoundland and Labrador cod stocks will require a return to a food web in which cod feed mainly on pelagic species such as capelin (Mallotus villosus).
In a multi-species context, biological reference points should vary with changes in growth, maturity, and especially natural mortality (Collie and Gislason, 2001). The estimation of reference points, including FMSY and BMSY, depends on the demographic parameters of the rebuilding species, which in turn depend on the dynamics of other species in the food web. In particular, the reference points of harvested prey species are conditional on the abundance of their predator species. The level of predation could be a factor in the rebuilding of prey species such as butterfish in the mid-Atlantic and crab stocks in Alaska. Predation mortality can be incorporated in stock assessments implicitly with a time-varying natural mortality rate or explicitly with a dynamic multi-species model (Hollowed et al., 2000). To date rebuilding analyses have incorporated neither time-varying natural mortality nor the unstable dynamics that this could produce.
Species that are predators as adults experience predation during their early life stages. Predation by clupeid species (such as herring) is hypothesized to be a substantial source of mortality on the eggs and larvae of gadid (cod) species (Daan et al., 1985). This predator-prey feedback can lead to alternate clupeid- or gadid-dominated states. A meta-analysis of cod-herring interactions indicated negative effects of herring on cod recruitment for several stocks in the North Atlantic (Minto and Worm, 2012). When embedded in a length-based model of the Georges Bank fish community, this predation effect delayed, but did not prevent, cod rebuilding (Collie et al., 2013a). These, and similar results for the North Sea (Speirs et al., 2010), suggest that, although they may exist, such predation triangles do not necessarily result in depensation that would prevent cod stocks from rebuilding. A detailed analysis of Atlantic cod on the eastern Scotian Shelf found no evidence that rebuilding was delayed by a high biomass of forage species that could prey on cod eggs and larvae; rather the lack of rebuilding was attributed to high natural mortality at the adult stages (Swain and Mohn, 2012).
The community aspects of rebuilding have been investigated with size-based models. Rebuilding a target species can have indirect predation effects on smaller species and on the juveniles of larger species (Andersen and Rice, 2010). Overfished species rebuild at different rates, altering the predator-prey dynamics compared with the unexploited fish community (Collie et al., 2013b). In simulations, prey species released from predation rapidly increased and overshot their unexploited equilibrium level. In contrast, large predator species increased slowly, failing to recover after 25 years of simulation. Again, the delayed rebuilding of predator species was not due to depensation, but it suggests hysteresis in community rebuilding.
Although some species subject to rebuilding are currently managed as stock complexes (e.g., Caribbean grouper, Hancock Seamount groundfish complex), most rebuilding plans are for single species, and fishery management plans require species-specific annual catch limits. An ecosystem perspective might consider rebuilding aggregate species groups instead of rebuilding on a stock-by-stock basis. Aggregate production models have been used to estimate sustainable yield at the community level (Brown et al., 1976; Mueter and Megrey, 2006) and can be used to set a cap on the Total Allowable Catch (TAC) across multiple species. On the one hand, there is evidence of compensation within functional groups (Auster and Link, 2009), so functional groups based on diet similarity would be the units of management; depletion of one species could be offset by increases in another species in the same functional group. On the
other hand, there is considerable overlap among functional groups, however they are defined. Most functional groups are dominated by a few species, such that the functional-group dynamics simply reflect the dynamics of the dominant species in the functional group.
INCORPORATING ECOSYSTEM CONSIDERATIONS INTO REBUILDING PLANS
A general conclusion is that rebuilding plans should consider the structure and functioning of populations and ecosystems in a wider sense to maximize the ability of fish populations to rebuild. Rebuilding plans should ideally entertain a broader spectrum of ecosystem dynamics and possible outcomes than is typically considered in single-species rebuilding projections, particularly in light of what is currently known about the prevalence of such dynamics in nature. Reductionist approaches that try to separate the effects of fishing and the environment may overlook important interactions.
Biological reference points based on MSY (and its proxies) are moving targets because of the complex and dynamic nature of ecosystems. Reference points, or possibly other performance criteria, should be sought that are appropriate for the observed dynamics, can accommodate ecosystem changes, and have no unintended consequences for rebuilding. If a reference point or formula is adjusted for prevailing ecological conditions, then it should aim to reduce fishing mortality when productivity declines. As a result of ecosystem dynamics, including biological competition and predation, fisheries management involves tradeoffs among harvested species, even if conscious decisions are not made about the tradeoffs or if they cannot be predicted with confidence.
What is possible given the present level of scientific understanding? We know many of the mechanisms that make ecosystems dynamic, and our understanding is advancing rapidly. Environmental variables can be included in the estimation of reference levels where they are known to affect demographic parameters (e.g., weight-at-age, maturation, fecundity, recruitment) of the species. Some of the better-understood ecosystem considerations can be incorporated into Management Strategy Evaluations (MSEs) of rebuilding (Punt, 2011). Multi-species models of intermediate complexity can be statistically fit to time-series data of interacting species (Plaganyi et al., 2012). A likely consequence of applying these approaches is an increase in the uncertainty in stock rebuilding projections.
In most cases, we do not yet now know enough to predict the future state or to manipulate ecosystems to achieve desired tradeoffs among species (even if there was agreement on which tradeoffs are desirable). Therefore, needed are practical, operational, and robust management strategies for fisheries rebuilding (see Chapter 7 for additional discussion). Meta-analyses and MSEs are probably key tools to advance these methods. Analysts can embrace multiple working hypotheses and integrate performance outcomes across hypotheses, weighted by the probability of each hypothesis being true. Other poorly understood ecosystem considerations can only be accounted for in a qualitative sense.
Stock rebuilding can proceed without full understanding of ecosystem dynamics. A previous NRC committee (NRC, 1999a, p. 5) concluded, “[A] significant overall reduction in fishing mortality (F) is the most comprehensive and immediate ecosystem-based approach to rebuilding and sustaining fisheries and marine ecosystems.” Since that report, there has been a significant overall reduction in F as indicated by a reduction in proportion of stocks suffering overfishing (see Chapter 3). A better understanding of the dynamics of depleted fish stocks depends on the continuation of existing data collection programs, because many analytic methods are constrained by short time series. Process-oriented studies are needed to elucidate the interactions between fish and their environment (e.g., the dependence of fish production on habitat).
Some ecosystem considerations may imply longer rebuilding times or require lower fishing mortality rates for rebuilding. Conversely, favorable environmental conditions can reduce rebuilding times. Either way, ecosystem considerations should not be used as excuses for inaction. They do not contradict a tenet of fisheries science, that harvested stocks have finite capacity to compensate for increased mortality, but they do supplement and extend it. Most fish stocks can rebuild when fishing mortality is reduced (see Chapter 3). If population dynamics are nonlinear, as recent studies suggest, then fishing mortality may need to be reduced below a threshold level to initiate rebuilding (Collie et al., 2004).
Stocks with episodic recruitment such as rockfish off the West Coast pose a special challenge because of the difficulty of distinguishing regime shifts from delayed rebuilding. However, the appropriate management action in both cases to rebuild a depleted stock is to reduce fishing mortality below FMSY. The stock should eventually rebuild if there is episodic recruitment. If the stock has shifted to a lower productivity regime, then it can still be sustainably harvested with the lower fishing mortality rate, but it may not rebuild to biomass targets defined under past conditions. In the case of a regime shift, the lack of recovery would not be considered a management failure. This example leads to a more general conclusion that ecosystem considerations can temper our expectations about the levels to which stocks can rebuild and the time it takes to get there. However they do not alter the general prescription of reducing fishing mortality on depleted stocks.
FINDINGS
5.1: Ecosystem considerations imply a broader spectrum of population dynamics and possible outcomes than is typically considered in single-species rebuilding projections.
Stock biomass estimates and projections can vary greatly in response to alternative plausible assumptions (models) and parameter values used in simulations, because the underlying population dynamics are nonlinear.
5.2: With climate change, and because of the complex and dynamic nature of ecosystems, biological reference points (such as BMSY) based on single-species production functions are likely to change over time.
5.3: Fishing and the environment interact in ways that are nonadditive. Fishing-induced age truncation amplifies the effect of recruitment variability on population dynamics, and it may increase coupling between recruitment and environmental conditions leading to more variable recruitment. An important implication of this hypothesis for overfished stocks is that rebuilding the demographic structure of the population will increase the probability of strong recruitment.
5.4: Habitat loss and degradation may limit reproduction, feeding, and growth, but studies have been inconclusive and it is difficult to scale results to the population level. For most species, the relationship between habitat and productivity remains unquantified. Process-oriented studies can elucidate the interactions between fish and their environment (e.g., the dependence offish production on habitat).
5.5: In a multispecies context, growth, maturity, and natural mortality are influenced by the abundance of interacting species, although these effects are difficult to predict. Biological reference points for forage species may need to allow a larger proportion of the production of these species to be available to predators than for higher trophic-level species.
5.6: To address species interactions, stock assessments have been linked in a number of ways. Multi-species models can inform the natural mortality (predation) rates used in single-species assessments of prey species (e.g., herring, menhaden, walleye pollock). For species that are both predator and prey, incorporation of species interactions requires a dynamic multi-species model (e.g., age-structured multi-species models that have been developed for several ecosystems).
5.7: As a result of ecosystem dynamics, such as biological competition and predator-prey interactions, fisheries management involves tradeoffs between harvested species, even if tradeoffs are not deliberate decisions and outcomes are often unpredictable.
5.8: Scientific understanding of ecosystem dynamics is advancing rapidly. In some cases, understanding has advanced enough to model dynamics, which may be used to inform fisheries management decisions. However, in most cases, scientific understanding of ecosystem dynamics is insufficient to confidently predict the future state or to achieve desired tradeoffs among species (even if there were agreement on which tradeoffs are desirable). These cases depend on having pragmatic, operational management strategies that acknowledge this kind of uncertainty.
5.9: Ecosystem considerations, among other reasons, argue for more emphasis on rebuilding plans that maintain reduced fishing mortality for an extended period (e.g., longer than the mean generation time). This strategy rebuilds age structure and is more robust than is a focus on biomass targets, which may be more or less attainable depending on environmental conditions.












