
12
Hierarchy and Connectedness as Determinants of Health and Longevity in Social Insects
Brian Johnson and James R. Carey
INTRODUCTION
It is widely known that health and longevity in humans, the most social of all vertebrate species, have strong sociological determinants ranging from marriage (Waite and Lehrer, 2003) and family (Waite and Das, 2010) to friendship networks (Couzin, 2009) and societal rank (Marmot, 2004). These determinants are the outcome of co-evolved behavioral, demographic, and ecological traits that both animate and link a society, and that are both complementary and mutually affecting. It follows that understanding these traits and the interplay between society, the individual, and individual-level health for any social species, but particularly for understanding these relationships in humans, requires deep knowledge of the roles of social networks and hierarchies (Barry and Yuill, 2012). Inasmuch as it is not possible to manipulate any human society or its individual members in any type of large-scale, controlled study, not only do many questions about the role of social factors in health go unanswered, but also much of the conventional wisdom derived from human studies remains unsubstantiated.
Because of the paucity of information on how social factors, including networks and hierarchies, affect health and longevity in social species in general and humans in particular, our broad goal of this paper is to describe a general research framework for answering questions concerned with sociality and health based on the use of social insects in general and the honey bee (Apis melifera) in particular. We begin with a section containing overviews of social insect longevity, evolution of eusociality in
wasps, general principles of social insect biodemography, and a synopsis of the biology of honey bees—the species we focus on throughout much of the remainder of the paper. This is followed with the main section involving mechanisms involved and principles determining aging and longevity in the honey bee. We end with a short section on recommendations for research and hypothesis testing.
SOCIAL INSECTS: CONCEPTS, EVOLUTION AND BIODEMOGRAPHY
Although social insects constitute only 2 percent of the more than 1 million species of insects on earth, they may constitute up to 80 percent of the insect biomass and, in total, outweigh vertebrates by 7-to-1 (Wilson, 1985; Charbonneau et al., 2013). Due to their evolved ability to exploit the concept of division of labor, they are the most ecologically successful groups in nature. They display sophisticated problem solving that emerges from simple individual behaviors that often resemble a voting procedure. Individual workers are adaptively allocated to different tasks (e.g., caring for eggs, larvae, and pupae; building; foraging; defense) in a way that is robust to changes in both task demand and to individual failure. Since the structure and dynamics of the network of interactions will affect overall colony functioning, many of the processes are regulated by interactions between individuals. Thus, the health and longevity of the colony and of individuals is mutually-affecting and complementary. Social insects are excellent models for studying all aspects of sociality as they are an evolved system in which the interacting parts can be individually tracked and manipulated (see Figure 12-1).
Sociality Concepts
Although sociality occurs to some degree in virtually all insect species in the form of the social activities of courting and mating, most biologists consider a species of insect as social only if they exhibit specific types and levels of cooperation including (1) continued care of young; (2) cooperative brood care; (3) reproductive division of labor; and (4) colonies with at least two adult generations (Carey, 2001). The various levels of social organization are thus classified according to combinations of these cooperative behaviors including subsocial, colonial, communal, cooperatively breeding, and eusocial. Although some level of social behavior is found in 12 different groups of insects and acari, eusociality is found primarily in the Hymenoptera (bees, wasps, ants) and Isoptera (termites) with a few “non-traditional” eusocial species also contained in the Homoptera, Coleoptera, and Thysanoptera.

FIGURE 12-1 Social insects—ants and honey bees.
NOTES:
(a) Individually marked ants, Temnothorax rugatulus. © Alex Wild, used by permission.
(b) Purple grains of sand built by ants to protect the colony. © Ann Dornhaus, used by permission.
(c) Queen honeybee retinue. Older egg laying queens are more attractive to workers than newly mated queens and virgin queens are the least attractive. The workers groom the queen (i.e. retinue behavior) by touching it with their mouthparts, antennae and forelegs. The workers from the queen retinue feed the queen by trophallaxis. During grooming of the queen pheromones are transferred from the queen to the workers attending it and later to other workers in the colony. Whereas worker bees typically live only around 6 weeks, queens are capable of living 6 years or more. © Kathy Keatley Garvey, used by permission.
(d) Honeybee comb showing brood chamber with eggs (not visible but in open cells in middle), larvae (see cells to right), sealed brood (closed cells), queen (marked individual in center) and nurse (worker) bees (all other adults). The honey bee hive consists of a single queen, multiple drones and from 10,000 to 40,000 workers. © Kathy Keatley Garvey, used by permission.
Lifespans in Social Insects
A summary of the range of lifespans in the four main groups of eusocial insects is given in Figure 12-2. Several aspects of this schematic diagram merit comment (Carey, 2001). First, lifespans of eusocial insect adults differ by 800-fold from the 10-14 days lifespan in some worker ants to the 30-year lifespans of ant and termite queens. As Hölldobler and Wilson note (1990), the longevity of Camponotus, Formica, and Lasius queens, ranging from 18 to 29 years, makes these ants the most long-lived insects ever recorded. Clearly, longevity is a highly selected attribute of the life history of the different species. Second, the extraordinary range in lifespans occurs not simply between the different species, but also within species. For example, queen honey bees are capable of living 3-5 years but honey bee workers only live 6-8 weeks. This is remarkable because the queens do not differ genetically from workers—they are produced through workers constructing special “queen cells” (Seeley, 1995) and feeding the developing larvae
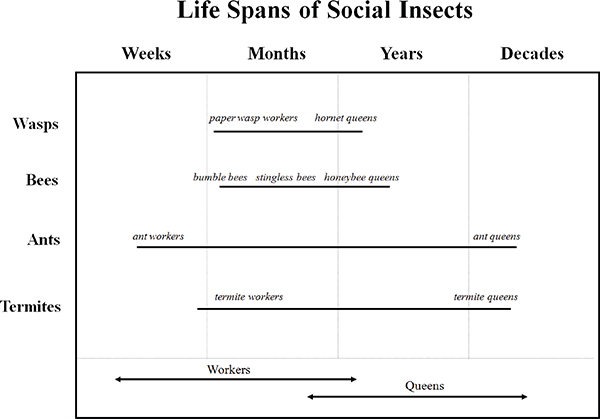
FIGURE 12-2 Lifespan ranges of species within the four main groups of eusocial insects—Wasps, bees, ants, and termites.
NOTE: Arrows in bottom panel indicate range (and overlap) of workers and queens in each of the groups.
SOURCE: Carey (2001).
“royal jelly”—a complex mixture of diet and hormones (Finch, 1990). Third, relatively speaking, males (all workers in the social hymenoptera are female, and males, often called drones, have radically different life histories) have a shorter adult lifespan than either queens or workers (Wilson, 1971; Hölldobler, 1990), mother queens live much longer than workers in all eusocial insects (Keller and Genoud, 1997; Keller, 1998), and ant and termite queens (that are subterranean) experience much greater lifespans than wasp and bee queens (that are terrestrial). Fourth, no correlation is apparent between the degree of sociality of a species (colony size, morphological divergence of queens and workers) and both the absolute and relative longevity of its queens (Hölldobler, 1990). This suggests that the ecology and demography (e.g., foraging patterns, food type) of a social insect species, rather than colonial complexity, is an important determinant of queen longevity relative to that for workers.
Evolution of Sociality in Wasps
Emergence of Wasp Bauplan
Since each successive step in the development is complex, and the success of each is dependent upon the ones that precede, it is not surprising that during the course of evolution there should be a high degree of conservatism in each step (Bonner, 1988), resulting in what is known as the bauplan (Purves et al., 2004)—an assemblage of features (typically morphological) shared by a taxonomic group.
The bauplan for wasps is based on relatively short-lived sawflies, a precursor group of hymenopterans (i.e., bees and wasps) that uses its ovipositor as a precision tool for laying eggs in highly nutritious parts of plants. Short-lived parasitoids evolved from sawfly evolutionary progenitors that used this ovipositor as a “sting” for inserting their eggs into hosts. Parasitoids then evolved into mass provisioning (i.e., lays egg on or in entire host) wasps (Edwards, 1980), their mandibular (chewing) mouth parts preadapted them for nest-building (Jeanne, 1975). Moreover, their wasp waist (i.e., petiole-like) that evolved to enhance their host-stinging ability (i.e., highly maneuverable for precision stinging) restricted wasps to essentially liquid nourishment since only liquids could pass through to the abdominal digestive tract. That, along with progressive provisioning (as-needed feeding using masticated food), then preadapted them for social evolution (and thus long life). Liquid exchange between colony members became a mechanism for within-colony communication (Hunt, 1991).
Co-evolution of Wasp Sociality and Lifespan
A schematic for the progressive evolution of sociality and longevity in wasps can be developed based on the sequence of grades that, according to Hölldobler and Wilson (Hölldobler, 1990), can be logically envisioned to have occurred during behavioral evolution including nest building and guarding, brood care, and provisioning of young. Carey (2001) used the grades of social evolution in wasps outlined in the paper by Evans (1958) as a framework for examining the co-evolution of longevity and sociality in insects (also see Hunt and Amdam, 2005). Several evolutionary grades are grouped into one of three general stages (see schematic examples illustrated in Figure 12-3).
Stage I: Emergence of nest as locus (Figure 12-3a and b). The nest is important in the co-evolution of sociality and longevity because it provides a place to which the female repeatedly returns and thus becomes the locus of activity (Starr, 1991). The nest originated as a site of brood care and development or nursery and thus provides a protected microenvironment for the developing brood, the original female, and the food provisions. As females spend longer periods sitting in the protected nest, different selective factors become important. Mortality due to accident and predation is likely to be reduced, there is selection to reduce rates of senescence, lifespan increases, and mothers are more likely to be alive when some of their offspring reach adulthood.
Stage II: Emergence of parental care (Figure 12-3c). The female brings more prey to the nest, as needed, for the developing larva. She may remain in the nest with her offspring, adding some protection for the young. Later, females may begin to feed macerated food directly to the larvae. The evolution of adult provisioning by social insects enabled pre-adults to grow rapidly towards adult competence under the protection of parental care. This provisioning not only prolongs pre-adult (larval) dependence and accelerates growth, but also it detaches the larvae from the environment—short-term food shortages that would be fatal to larvae not under parental care are ironed out as the mother continues to provision by expanding her foraging activities (Queller, 1989).
Stage III: Emergence of queen/colony concept (Figure 12-3d). The female’s life is prolonged and overlaps that of the progeny that remain with her in the nest to form an extended family. Females at this phase lay eggs into an empty cell, a trait that is thought to be important in permitting the evolution of the extended brood care of social wasps. The young females may construct more cells, lay eggs, and care for their own larvae. However, some
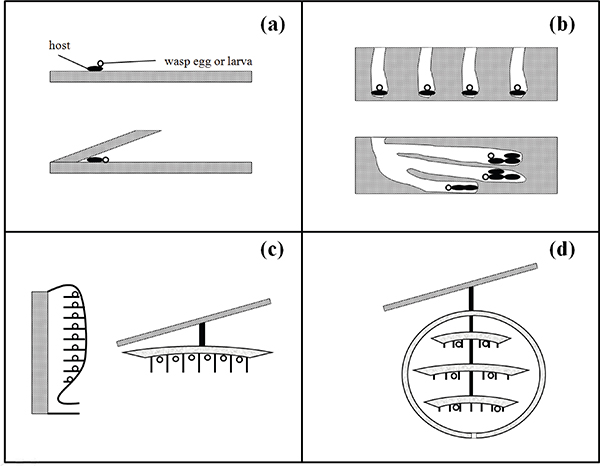
FIGURE 12-3 Schematic diagrams of the nests or prey sequestration behavior of wasps in various evolutionary stages of sociality, all of which alter selection on lifespan (Wenzel, 1991).
NOTES: (a) Parasitoid wasps. Host as incubator of parasitoid larvae (short lifespans with ranges from 14-60 days). Examples: spider wasps, Notocyphus; cockroach wasp, Ampulex; (b) Nest provides a protected microenvironment for female and brood, thus different selective factors operate; selection reduces senescence rates and increases life span (intermediate lifespan ranges from 30-60 days). Example: sand nesting wasps, Haploneurion; Ammophila; (c) Extensive parental care increases life span to 1 year; larval and pupal mortality reduced to near zero; this fosters reduction in birth rate; allows females to invest more resource for their own maintenance and for rearing of their offspring (lifespans range 60-365 days). Examples: progressive provisioning Gorytes spp. and paper wasps, Polistes; (d) Colony and queen concepts. Female’s life further prolonged and overlaps with progeny which remain in nest to form extended family (helpers); colony rather than the individuals within it begins to become the unit of selection with the fate of the queen inextricably tied to fate of colony; a new level of individualism emerges (colony) and thus life of queen becomes adaptive to the long-term colony needs (lifespan range: 180-1,000 days). Example: advanced eusocial wasps including yellow jacket, Vespula spp.
SOURCE: Carey (2001).
females occasionally care for the brood of another female. Extensive overlap of generations occurs because of extended lifespans and because a clear division of labor and caste systems begins to emerge. A single queen, usually the original foundress, is much more long lived than the workers and monopolizes oviposition (i.e., queen control). As long as the colony remains small and the lifespans of reproductive individuals and helpers are not significantly different, then helpers may be expected to resist evolutionary specialization as workers because this would reduce or eliminate their chances to replace their mother as the sole reproductive female (Alexander, 1974).
Social Insects: Biodemographic Principles
The main principles of longevity extension in social insects, as derived from the wasp evolutionary model include the following (Carey, 2001).
Generation Overlap Is a Prerequisite for the Evolution of Incipient Sociality
One of the prerequisites for the evolution of eusociality in insects is generation overlap—the parents must live long enough to still be alive when their offspring emerge as adults. Evans (1958) noted that an important factor explaining the lack of significant social evolution in sphecid wasps is the relatively short female lifespan, so that the adult lives of mothers and daughters rarely overlap.
Progressive Provisioning Evolved after the Evolution of Extended Longevity
The assumption is that progressive provisioning arose from delayed provisioning. There is no strong evidence that adult longevity depends on progressive rearing but that extended adult longevity is an ancestral trait and that progressive provisioning evolved later (Evans, 1958). This concept is important because it suggests that extended longevity serves as a necessary precondition for the evolution of incipient sociality. Thus subsocial behavior may be an outcome of extended longevity rather than vice versa.
Sociality Provides Insurance-based Survival Advantages
In many facultatively eusocial insects, offspring need continuous care during development, but the life expectancy of adult caregivers is shorter than the developmental period. When a lone foundress dies, her partly reared brood is usually doomed. Helpers in a tropical hover wasp (Liostenogaster flavolineata) have an insurance-based advantage over lone foundresses because after a helper dies, most of the brood that she has partly reared will be brought to maturity by surviving nest-mates (Queller,
1989). The consequences of early mortality differ between solitary reproducers and helpers in several Polistes spp.—the former that die early will fail to bring any young to independence, while helpers that die at the same age may have made substantial contributions.
Queen-Worker Longevity Differentials Diverge at Advanced Stages of Eusociality
As eusocial colonies become large and long-lasting, mothers become increasingly specialized as reproductives, and offspring as workers and soldiers (Johnson and Linksvayer, 2010). As this happens, daughters have fewer opportunities to become replacement reproductives, both because the mother lives longer and because so many other individuals are available to replace the mother as well. Thus the identity of a queen’s replacement becomes a sweepstakes, with each individual having a chance of, perhaps, one in fifty thousand or one in a million. One consequence of increased longevity as a preadaptation to sociality is that the stage is set for the evolution of large colonies (with even longer-lived queens) that have certain advantages over smaller ones, including the ability to organize labor more efficiently, integrate colony activities, defend the nest more aggressively, and exert homeostatic control over physical conditions within the nest. The fitness to the individual worker and the queen is subordinated to the fitness of the colony (i.e., emergence of the super-organism).
SOCIAL INSECTS: MOLECULAR MODELS OF AGING
Operational Framework
We consider the two basic conceptual frameworks for understanding aging in all organisms including social insects—accumulated damage and adaptive senescence (Finch, 1990). The first assumes that molecular damage of various sorts accumulates in the adult organism (who may or may not be post-reproductive) until such time as it impairs function and causes or contributes to death (Muller et al., 2007). Molecular damage is thought to be mainly caused by reactive oxygen species (ROS) that are the result of normal mitochondrial function. Adaptive senescence simply assumes that aging-related decreases in function are caused by genetic changes that occur as a result of a developmental program that has been shaped by natural selection. This is common sense in that it is often the case that relatively closely related organisms have widely different longevity patterns. Hence, lifespan in some sense must be the result of selection for an optimal longevity.
Of course, ultimately these two hypotheses are not mutually exclusive, as an optimal longevity (and a genetic program designed to achieve it)
can be the result of regulating the rate of damage produced by ROS (or other processes) such that they only lead to death at the appropriate age. Generating telomeres of a suitable length would be an example, as would be producing sufficient age-specific levels of antioxidants. We will frame much of our review of the molecular work on aging in social insects such that it either falls under one category or another, with the assumption that ultimately the two are complementary.
Extreme Longevity
Social Insects as Models
Social insects are good models for the study of extreme longevity and the underlying mechanisms for many reasons (Amdam, 2011; Roach and Carey, 2014). First, across species there is wide variation, with some species having lifespans typical for insects (on the order of weeks to months) and some having lifespans on the order of several decades (for queens). Second, within the same species, it is often the case that both patterns are apparent. Workers often have typical lifespans of a matter of weeks to a couple of years, while queens in the same nest can live for an order of magnitude longer. Given that genetic background can be a confounding variable in studies of the molecular basis of aging (Helfand and Rogina, 2003b), having the ability to study radically different aging patterns encoded by the same genome is highly beneficial. Of course, it is also possible to use the comparative method across species to identify genes, and networks that are associated with aging processes, as is done in many other taxa.
Social insects also decouple a fundamental relationship thought to be associated with aging in other species, namely that patterns of senescence are thought to be selected to change strongly before and after reproduction. This is because once an organism has reproduced, there should be little selective force for maintenance of somatic functions (Carey et al., 1992; Vaupel et al., 1998). In the social insects, however, because the queen produces eggs throughout her life (and has no post-reproductive life), this is not the case. Workers can also show atypical patterns of development with respect to hormonal state, reproductive status, and age. Genes that show positive relationships with aging in other insects, such as vitellogenin (the yolk protein), can show the opposite pattern in social insects such as the honey bee (Amdam et al., 2007). These patterns allow an experimenter to decouple the effects of reproduction from aging, something that is impossible in many taxa. In addition, humans, of course, are also social creatures who contribute to the fitness of their offspring, and grandchildren, throughout life, meaning that selection for long life is expected for humans as well. In this way, humans are atypical organisms, like the social insects,
with respect to aging and reproduction. Animal models that share the pattern of decoupling reproduction and senescence might therefore be the best models for the study of human longevity.
With respect to mechanistic studies of extreme longevity in social insect queens, one would like to know by what mechanism queens are able to prolong their lifespan far beyond the norm for insects. With respect to molecular damage, is it by generating large amounts of antioxidants, by slowing down metabolism, or by increased ability to repair damage? Do they continue to upregulate their immune systems for later into life than workers? We review briefly what research has been done, which supports all of these mechanisms (with notable exceptions, such as in the role played by ROS production).
Research has shown in model systems that upregulation of antioxidants causes longer than normal lifespan (Tatar et al., 2001; Helfand and Rogina, 2003a). Parker et al. (2004) tested the hypothesis in ants using the antioxidant gene Cu-Zn superoxide dismutase 1 (SOD1). Contrary to predictions, long-lived queens had lower expression of SOD1 than shorter-lived males. Queens and workers, further, did not differ in expression of this gene. Corona et al. (2005) also found no evidence for higher expression of antioxidants in long-lived honey bee queens relative to short-lived workers. They did find preliminary evidence for variation in ROS production, given their finding of caste-specific expression patterns for key mitochondrial metabolism genes. In sum, these studies support the hypothesis that long lifespan need not be associated with high expression of antioxidants. Decreased production of ROS rather seems a more plausible mechanism for these cases of longevity variation, as has been shown in a number of other systems (Muller et al., 2007).
ROS and Longevity
A variant of the ROS damage hypothesis has recently attracted much attention (Hulbert et al., 2007). This is the hypothesis that the fatty acid composition of cell membranes (ratio of saturated to unsaturated fats) is a major determinant of the rate of oxidative damage to membranes. Further, there are strong correlations between the Peroxidative index (PI) index (a measure of the susceptibility of membranes to peroxidative damage) and longevity across many groups of animals (vertebrates and invertebrates). Haddad et al. (2007) tested the potential for this hypothesis to explain lifespan variation between honey bee queens and workers. They found that the PI index of queens is lower than that of workers (a low index indicates low susceptibility to peroxidative damage). They also found that newly emerged workers have the lowest indexes for workers (still higher than for queens), but that the PI index increases with age. They hypothesized that
the increase in the PI index of workers is the result of pollen consumption, something that the queen is protected against given that she is fed a special nurse bee derived secretion throughout her life. They also found that the difference in PI index between queens and workers is sufficiently large to potentially explain the difference in longevity between these castes. To date, such research is preliminary, in that only correlations have been shown. Although no experimental cause-and-effect studies have been conducted, the data are intriguing and warrant more attention.
Thus far, we have focused on variation between workers and queens in longevity; however, there are also cases of strong variation within the workers on lifespan. In honey bees, the summer bees live about 6 weeks on average, while in the winter bees live up to 6 months (Johnson and Linksvayer, 2010). Essentially, in the summer, the colony is focused on rapid growth and food accumulation and the way they maximize this is by using a system of division of labor (mentioned previously). In the winter, however, the focus switches from growth to survivorship and the physiological specializations of the workers (to be nurses or foragers, for example) disappear. Instead, all the bees go into one physiological condition that is similar to the nurse bee phase (high vitellogenin titer and low juvenile hormone). Much work has taken advantage of this summer versus winter bee phenomenon to study aging processes (Seehuus et al., 2006a; Munch and Amdam, 2010). We will cover one case here related to oxidative damage and then return to the topic of worker aging in later sections.
Foraging honey bees have the highest metabolic rates of any animal measured (Vance et al., 2009). This is not surprising giving that their flight muscle alone makes up a significant portion of their total biomass. Foraging honey bees spend most of their day busily shuttling back and forth from the hive to the field (i.e., loads of nectar or pollen). Williams et al. (2008) took advantage of this phenomenon, and the fact that nurse bees only take one short flight a day, to look at the interaction between ROS production and senescence in honey bees. As expected, flight caused an increase in the production of Hsp70, which is correlated with oxidative damage. Older foragers, however, did not increase their antioxidant capacity after flight (presumably to respond to this damage), while young foragers did (to a modest, but significant extent). Tests for oxidization damage (based on recording protein carbonyl content) after flight (and in young and old foragers in general) did not show oxidative damage in either group. The authors interpreted these results as supporting the notion that ROS production in the flight muscles contributes to functional senescence in foragers; however, it is unclear that a cause-and-effect connection was made, as no damage was detected and older foragers had high antioxidant capacity even if they had no ability to increase it further. Nevertheless, it is the case that high ROS production in forager honey bees is a good context
to explore the mechanisms by which organisms mitigate the effects of high ROS production.
Adaptive Senescence
Adaptive Aging
Worker honey bees have been the focus of much work on adaptive senescence. In short, honey bee workers in the spring and summer have a complex system of division of labor in which workers exhibit different physiological states (castes) that appear to vary in their rates of senescence. There are four castes that occur in a developmental sequence: newly emerged bees, nurses, food storers, and then finally foragers. The nursing phase is most like the queen honey bee in terms of having high vitellogenin stores (and high nutrition levels in general) and low juvenile hormone (JH) titer. Further, bees that are kept in the nursing phase, by manipulating the nest such that they never transition to the next phase, can live for many months, while foragers, in contrast, only live for 2-3 weeks (Dukas and Visscher, 1994; Remolina et al., 2007). After a couple of weeks of foraging, for example, bees display obvious signs of damage (torn wings, damaged flight muscles, etc). There is also some evidence that this decline in foragers may be adaptive (i.e., the result of patterns of gene expression that turn off various pathways important for repairing damage; this is likely associated with reallocation of resources to other more critical functions). Finally, winter bees live throughout the whole winter (up to 6 months) and have a physiology superficially similar to that of nurses. Taken together, these patterns of aging in worker honey bees have been the subject of considerable work. However, several key experiments have not been conducted. Here we review some of the key studies thus far and point out the areas still needing work.
Aging Stasis
In honey bees, an appropriate number of workers in each caste is optimal for a given colony-level growth rate. This is because the different castes work together such that work is either passed from one group to another (foragers collect food, but younger food processors process it), or the work rate of one group is dependent in some more general manner on the activities of another group. The balance of workers in each developmental phase is maintained by workers either speeding up or slowly down their development in response to the colony-level ratios of workers current in each caste. Of chief experimental importance, development can be sped up or slowed down via various experimental techniques. It can be increased, for example, when a large of number of workers in the next phase dies, or it can be
slowed down when new workers are not produced to take the place of older bees (Huang and Robinson, 1992, 1996). Although it does not happen in natural nests, the aging processes can even be frozen by manipulating the ratios of bees in the nest (and the amount of brood in the nest) such that the ratio of workers in each phase does not change (Johnson, 2010). This has been done by removing newly emerged bees (to prevent replacement of nurses) and by adding open brood requiring care. The result is that many bees never seem to leave the nursing phase. Haydak (1963) was able to produce nurses 138 days old in this way. This is equivalent in some sense to freezing the aging of a vertebrate such that it remains a juvenile indefinitely.
Although the biology underlying this frozen development is derived, it is not unique. A similar process occurs in sex-changing fish. In some of these species, individuals all begin as females and a transition to being male only occurs in the fish that is the largest in the group. Hence, most individuals never make the switch. In the case of bees, because their aging is controlled by mechanisms similar to those that control sex in sex-changing fish (Munday et al., 2006), it is thus possible to ensure that some bees never make the transition to some developmental phases. This allows for studying the effects of extreme aging in the context of a body that is working to maintain an optimal functional state. In other words, adaptive senescence should not occur in bees frozen in the nursing phase, but senescence due to unrepairable damage (caused by ROS, for example) should nevertheless occur. Hence, it provides a context to study aging due to molecular damage controlling for the effects of adaptive senescence.
Aging Reversal
Bees that have entered the last of the developmental phases that characterize adult development, the foraging caste, can be induced to revert to the nursing phase by simply removing all the young bees from the nest (Huang and Robinson, 1996). When this manipulation is done, there is a rapid decrease in the JH titer of all the foragers in the nest and then some of the foragers acquire the ability to feed brood. Essentially, they reactivate their brood-feeding glands, and they seem to increase production of Vg, which is required for production of brood food (Amdam et al., 2005). This process is thought to be mediated by chemical signals sent by the foragers to the younger bees (and likely vice versa) that inform the members of the nest as to whether the correct ratio of bees in the different phases is present for optimal colony performance (Huang and Robinson, 1996). The fact that bees can revert from the old phase to the young phase has attracted considerable attention from aging researchers, as it is a natural example of repairing the damaging effects of senescence (Amdam et al., 2005; Amdam and Page, 2005; Seehuus et al., 2006a). It should be possible to learn
about molecular mechanisms (genetic and epigenetic) for the prevention of senescence and for the repair of aging damage by documenting what genes turn on in foragers that have reverted to nurses (Herb et al., 2012; Locket et al., 2012). There is thus much to learn from studying this highly derived pattern of aging in bees. However, as for extending the longevity of nurses, there are still some key experiments missing.
Missing Experiments
The most derived (as distinct from ancestral) patterns of aging are (1) bees can be kept in the nursing phase perhaps indefinitely, and (2) foragers can be induced to revert to the nursing phase, recovering some aspects of nursing physiology (perhaps including increased longevity). With respect to both phenomena, key experiments have not been conducted. Hence, although there is great promise in the study of these two phenomena, there is also some need for caution.
Although it is known that nurses can be kept in the nursing state for a very long period of time, it is also known that young and old nurses have many differences in their physiology (Wegener et al., 2009). Remolina et al. (2007) also found that overage nurses do experience senescence, in that they are less able to handle physiological stresses, such as heat and oxidative damage (relative to younger nurses). It is thus possible that overage nurses are a mosaic of young and old phenotypes. The fact that they live for so long is indicative of anti-aging processes, but the fact that they are more susceptible to physiological stress suggests that the freezing of development is not complete. Much work will be needed to tease apart what aspects of aging are frozen and what proceed at a normal rate in overage nurses.
Although it has been shown that foragers can revert to the nursing phase, and many studies have taken advantage of this phenomenon to study developmental polyphenism, there are several key studies that have not been conducted that must be performed before the utility of this phenomenon for the study of aging can be determined. First, studies showing aging reversal have not controlled for the age of the foragers that revert. It is known that senescence does not start until late in a forager’s career. It is possible (and even likely) that the bees that revert from foraging to nursing are only the youngest foragers (and conversely the older foragers might not have this ability). Further, since the young foragers may not have begun the process of senescence, it is not clear that they have reversed the aging process. In contrast, they may simply have changed physiological and behavioral state without having to repair or retard senescence. Studies that control for age of the foragers must therefore be performed. These studies should ensure that only old foragers are present in the nest, and that it is these foragers that revert to the nursing phase.
Second, although it has been shown that foragers can reactivate their brood-feeding glands and change their JH titer to that more characteristic of the younger phase, it has not been shown that this leads to greater longevity. What has been shown, essentially, is that foragers can become nurses, but given that the reverted nurses appear to be rather poor nurses, it is possible that this is a temporary solution for the colony (that is, the reverted foragers will not last long and must be replaced by a new batch of young nurses). Hence, studies must be conducted that show that bees that revert from foraging to nursing can then be induced to stay in the nursing phase for an extended period of time. This will show that the bees reacquire not only the ability to perform nursing tasks, but also the physiological capacity to extend their lifespan for as long as is necessary for the colony. Given that ants that start out as workers and then switch to reproductives can increase their physiological capacity (Schneider et al., 2011) suggests that such a capacity is possible.
Senescence and Immunity: Health in Older Workers
Disposable Castes: Downregulation of Immunity in the Honey Bee?
Foragers in social insect colonies have long been considered the disposable caste. This is because the act of foraging is risky and is associated with a high mortality rate relative to those workers that remain in the nest. Hence, if mortality rates are high, it might make sense to strip the foragers’ bodies of all the resources that are not necessary and to store those resources in the nest (either in food stockpiles or in the bodies of younger workers), the result being that if foragers die, there is little cost to the colony. There is evidence to support this notion from several species of ants and bees (Toth et al., 2005; Robinson et al., 2009). Ant and bee foragers, for example, have much lower protein and fat titers than younger workers.
A corollary to the disposable caste idea is that the immune system of workers should be adaptively downregulated in foragers relative to younger bees (Amdam et al., 2005; Seehuus et al., 2006a). In this case, by shutting down the immune system, which is costly to maintain, foragers cost the colony less. Of course, decreased immune function is characteristic of senescence across most organisms (Ames et al., 1993; Sansoni et al., 2008). If the same process of functional decline is observed in forager bees, then this system would be ideal for studying age-related declines in immunity. This is because it is simple to record when a bee begins foraging and, hence, begins to senesce. Precise measurements of the rate of decay of immune function (in its various components) could then be made. Further, because foragers can revert to nurses, it may also be the case that they reactivate their immune systems, allowing one to observe what pathways are turned
back on to recover immune function. Much work has been done on this phenomenon of immune decline in foraging bees (Schmid et al., 2008; Laughton et al., 2011; Helft et al., 2012; Gätschenberger et al., 2013; Jefferson et al., 2013). Early work supported the basic hypothesis outlined above, but more recent work has questioned this simple perspective.
Re-Evaluation of Support for Adaptive Downregulation of Immunity
Early work supported the notion that foragers downregulate their immune system. In support is the long-known fact that nurses have high Vg titers and low JH titers, and foragers have the opposite pattern. Amdam et al. (2004) showed that Vg may act as an antioxidant and protect bees from oxidative damage (Seehuus et al., 2006b). Further, it was shown that foragers have low levels of functional hemocytes, while nurses have higher levels (Amdam et al., 2005). When foragers were forced to revert to nurses, for example, they recovered a higher level of functional hemocytes (and showed a moderate increase in Vg titer). All told, these results supported the notion that foragers actively downregulate immune function. The basic hypothesis could perhaps be expressed as the immune machinery is turned off at the transition to foraging, and then a process of senescence erodes the functional properties of the system.
In these first studies, the circulating level of functional hemocytes was taken as a proxy for the functional properties of the entire insect immune system. The insect immune system is complex, however. In broad strokes, there are humoral and cellular-based defense mechanisms (Lavine and Strand, 2002; Lemaitre and Hoffmann, 2007). The humoral system includes the production of anti-microbial peptides and ROS, while the cellular system is involved in phagocytocis, nodulation, and encapsulation of large foreign bodies. Both nodulation and encapsulation are often followed by melanization, which is associated with the phenoloxidase system (PO). The PO system can also apparently work in the absence of hemocytes (Lemaitre and Hoffmann, 2007). Insects also have an RNAi-based system for defense against viruses. Essentially, the insect immune system is complex with many different components, many of which overlap in function. Hence, it is not clear that a decline in one component (hemocytes of one type) can be a reliable indicator of either overall immune function, or even a particular component of immunity such as the totality of cell-based immune responses.
Work conducted since the original studies by the Amdam lab have generally supported the notion that immunity is not downregulated in foragers. Instead, it appears the immune system is re-modulated such that it retains functionality, but with a different mixture of immune system components playing key roles. It may even be the case that the immune system of
foragers is upregulated (Bull et al., 2012), which would not be surprising given that the foragers are the bees that are exposed the most to pathogens. Supporting this new prospect are a number of studies. With respect to the humoral response, Jefferson et al. (2013) found no decline in the production of antimicrobial peptides with age in honey bees. This barrier line of defense does not apparently decline with age, and this suggests that one component of the immune system (number of functional hemocytes) is not a good proxy for the whole immune system.
With respect to the number of functional hemocytes, Wilson-Rich (2008) found no decline in the number of functional hemocytes with age in honey bees. However, this study misidentified newly emerged bees as nurses, so it is possible that nurses do show higher levels of hemocytes than foragers. This study also found that foragers can encapsulate foreign bodies as well as larva, pupae, and newly emerged bees. Schmid et al. (2008), however, did find that hemocytes decline with age, but not for the reasons initially supposed. They tested whether the decline in hemocytes is specific to the worker caste or whether it also occurs in queens and drones (males). They found that the decline in hemocytes is found in all three castes. This suggests that this decline is not a result of adaptive senescence in workers alone, but is generally true independent of task or even sex. Further, they found that overage nurses also have depressed hemocyte counts, suggesting that aging, independent of functional role, contributes to the loss of hemocytes.
The function of the PO system is to facilitate melanization of pathogens (Lemaitre and Hoffmann, 2007). It is thought that high PO activity can compensate for overall hemocyte losses. In support of this, in the same study in which Schmid et al. (2008) found a decline in hemocytes with age (independent of caste), they found no decrease in PO activity between ages. Laughton et al. (2011), in a similar study, found higher levels of PO in foragers, relative to nurses. Helft et al. (2012) also found no difference in PO expression between young and old ants. This information, taken together with the results of Wilson-Rich (2008) that show that foragers can encapsulate as well as nurses, suggests that the decline in hemocytes found in several studies may not be associated with a decline in immune function. Other components of the immune system are retained, and perhaps are even amplified, to compensate for the loss of functional hemocytes. What is clear based on this mini-review is that the relationship between age and immune system functional senescence is quite complex in social insects, and it will require considerable work to disentangle the many confounding variables.
Lessons to Be Learned
Two lessons can be learned from the ongoing story concerning functional senescence of immunity in honey bees. First, simple interpretation
of gene expression differences alone is prone to error if functional studies are not also conducted (Bull et al., 2012; Gätschenberger et al., 2013). Second, there has been considerable oversimplification of the honey bee’s complex system of developmental polyphenism (correlations have been interpreted as causes). With respect to the first principle, Gätschenberger et al. (2013) found that although winter bees are unable to turn on the genetic machinery to cause the formation of nodules for bacterial defense, they are nevertheless equally effective at antibacterial defense as summer bees. This means tests that look for an immune response by looking for increases in expression of genes thought to be central to the process may miss an alternative mechanism supporting the same function. Likewise, Bull et al. (2012) conducted studies in which they challenged bees with bacteria and checked for the ability to increase expression of AMPs (a common bioassay for immune competence). They found that nurses respond to bacterial challenge by increasing expression of many key antimicrobial genes, while foragers do not show a gene expression change. In functional studies, however, foragers survived longer post-infection with a bacterial pathogen than did nurses. It appears that nurses respond strongly with a change in gene expression because their immune system is largely turned off at the point of challenge. Foragers, in contrast, have an already maximally activated immune system: that is, they have already produced much of the required protein and do not need to make more. It is likely the lag time in production of proteins for nurses (relative to foragers that have them already made) that is responsible for the difference in survivorship between nurses and foragers. Hence, the lack of an ability to respond with a rise in gene expression is actually indicative of superior function in this case. The conventional view has been that organisms without the ability to mount a response to challenge (assumed to be based on a rise in expression of key antimicrobial genes) are functionally deficient. Hence, it can be quite problematic to infer function from studies of gene expression alone.
The initial studies by Amdam and others that suggested that Vg is the linchpin underlying functional senescence in honey bees oversimplify the biology of developmental polyphenism in bees. Essentially, high JH in foragers and low Vg (and the opposite in nurses) are two out of thousands of differences between nurses and foragers. Although many correlations were shown suggesting that Vg was a key player in immunity, there were no cause-and-effect manipulations that clearly tie Vg to immunity. A critical test would be to test for various immune functions in drones, since these bees have very low Vg titers at all ages. Gätschenberger et al. (2012) did this and found that drones have competent immune systems in spite of very low Vg. Hence, in general, the developmental polyphenism of nurses and foragers is complex and care must be taken not to overinterpret the significance of differences in the expression of particular individual molecular actors.
RESEARCH OPPORTUNITIES
Because all individuals within societies are interconnected through mechanisms such as social support, social influences, social engagement, individual-to-individual contact, and access to resources, their health and longevity are also interconnected. There is no group of organisms where this is more evident than in the social insects. It follows that social insects are virtually untapped resources for use in answering basic questions regarding the influence of hierarchy and connectedness on health and longevity. Indeed, many behaviors co-evolved in the context of both individual and colony health.
We see at least five areas that hold promise for the use of social insects in future research on the biodemography of health, aging and longevity.
- Integration of colony-, organismal-, genetic,- and molecular-level studies on health, aging, and longevity. The hierarchical organization of insect societies from individuals to the superorganism of the colony provides unprecedented opportunities for research. Bee biologist Rob Page and his co-workers consider social evolution a vertical process from the molecular level through the colony using behavioral and anatomical analyses, physiology, genetic mapping, and gene knockdowns (Hunt et al., 2007; Page, 2013). They map out the phenotypic and genetic architectures of food storage and foraging behavior and show how they are linked through broad epistasis and pleiotropy affecting a reproductive regulatory network that influences foraging behavior, a major determinant of honey bee health and longevity.
- Intergenerational transfer, health, and longevity. In most models of life history theory, reproduction is treated as a purely demographic matter—i.e., birth is viewed as a single event independent of parental care. However, for many social species, continuing transfers to offspring are centrally important for survival, growth, and eventual reproductive success (Duarte et al., 2011). The theory offered by demographer Ronald Lee (Lee, 2003) strikingly shows that only the transfer effect shapes mortality, explaining both post-reproductive survival and why juvenile mortality declines with age. Since this theory also has deep links to kin selection and inclusive fitness, use of social insects to test hypotheses related to parental care and intergenerational transfer of resources has great potential for future research.
- Interdisciplinary research at the individual level. As Charbonneau and his colleagues note (Charbonneau et al., 2013), emerging technologies are facilitating the way for social insect researchers to
-
gather and analyze data at the individual level (digital tracking). This development is thus fostering interdisciplinary collaboration among population biologists, computer scientists, engineers, and mathematicians, thus creating a synergism in research. There are exciting new possibilities for gerontologists and biodemographers to engage in this exciting development and use social insect models to gain new insight into questions concerning health and longevity in social systems that are relevant to humans.
- Comparative biodemography of social insects. Because of the diversity of life histories and the evolutionary gradations of sociality from subsocial to eusocial within each of the four major groups of social insects (ants, bees, wasps, termites), the potential for comparative studies of concern with evolutionary and biodemographic studies of health, aging, and longevity is vast. Comparative studies have the potential to shed light on the evolutionary interconnectedness of health-related levels of defense including (a) individual defenses, (b) pair-wise defenses such as grooming, (c) colony defenses such as task differentiation, (d) minimizing the entry of infectious agents, and (e) use of environmental (e.g., resins) in colony shielding (Evans and Spivak, 2010). In addition, comparative studies may reveal how selection promotes genes that are most efficient in their ability to produce disease resistance phenotypes, gene products such as proteins and cells that play vital roles in physiological immunity, new groups of individuals that mount collective defenses (i.e., social immunity), and population-level scale disease dynamics (Wilson-Rich et al., 2009).
- Honey bee health. One of the burgeoning fields in social insect ecology involves studies of the determinants of individual- and communal (colony)-level health, respective concepts that are akin to clinical (medical) and public (communal) health in human societies. Even though the history of health studies in social insects and particularly in honey bees extends back well over a century with investigations focused mostly on various brood diseases (e.g., foul brood) or parasites (e.g., Varroa mite), the modern incarnation of health research in social insects emerged with arrival of colony-collapse disorder (CCD) in the mid-2000s, an unsolved decline in bees from parts of the United States, Europe, and Asia (Evans and Schwarz, 2011). Although the causes of CCD have yet to be determined, the efforts have led to the creation of a new framework to study health in social insects at multiple levels, ranging from the immunity of individuals to the defenses at the level of the colony (i.e., social immunity).
REFERENCES
Alexander, R.D. (1974). The evolution of social behavior. Annual Review of Ecology and Systematics, 5, 325-383.
Amdam, G.V. (2011). Social context, stress, and plasticity of aging. Aging Cell, 10(1), 18-27.
Amdam, G.V., Aase, A., Seehuus, S.C., Fondrk, M.K., Norberg, K., and Hartfelder, K. (2005). Social reversal of immunosenescence in honey bee workers. Experimental Gerontology, 40(12), 939-947.
Amdam, G.V., Nilsen, K.-A., Norberg, K., Fondrk, M.K., and Hartfelder, K. (2007). Variation in endocrine signaling underlies variation in social life history. American Naturalist, 170(1), 37-46.
Amdam, G.V., and Page, R.P. (2005). Intergenerational transfers may have decoupled physiological and chronological age in a eusocial insect. Ageing Research Reviews, 4(3), 398-408.
Amdam, G.V., Simoes, Z.L.P., Hagen, A., Norberg, K., Schroder, K., and Mikkelsen, O. (2004). Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Experimental Gerontology, 39(5), 767-773.
Ames, B.N., Shigenaga, M.K., and Hagen, T.M. (1993). Oxidants, antioxidants, and the degenerative diseases of aging. Proceedings of the National Academy of Sciences of the United States of America, 90, 7915-7922.
Barry, A.-M., and Yuill, C. (2012). Understanding the Sociology of Health (3rd ed.). London, UK: SAGE.
Bonner, J.T. (1988). The Evolution of Complexity. Princeton, NJ: Princeton University Press.
Bull, J.C., Ryabov, E.V., Prince, G., Mead, A., Zhang, C.J., and Baxter, L.A. (2012). A strong immune response in young adult honeybees masks their increased susceptibility to infection compared to older bees. PLOS Pathogens, 8(12).
Carey, J.R. (2001). Demographic mechanisms for the evolution of long life in social insects. Experimental Gerontology, 36, 713-722.
Carey, J.R., Liedo, P., Orozco, D., and Vaupel, J.W. (1992). Slowing of mortality rates at older ages in large medfly cohorts. Science, 258, 457-461.
Charbonneau, D., Blonder, B., and Dornhaus, A. (2013). Social insects: A model system for network dynamics. In P. Holm and J. Saramaki (Eds.), Temporal Networks (pp. 217-244). Berlin, Germany: Springer-Verlag.
Corona, M., Hughes, K.A., Weaver, D.B., and Robinson, G.E. (2005). Gene expression patterns associated with queen honey bee longevity. Mechanisms of Ageing and Development, 126(11), 1230-1238.
Couzin, J. (2009). Friendship as a health factor. Science, 323, 454-457.
Duarte, A., Weissing, F.J., Pen, I., and Keller, L. (2011). An evolutionary perspective on self-organized division of labor in social insects. Annual Review of Ecology, Evolution, and Systematics, 42(1), 91-110.
Dukas, R., and Visscher, P.K. (1994). Lifetime learning by foraging honey bees. Animal Behaviour, 48, 1007-1012.
Edwards, R. (1980). Social Wasps. East Grinstead, UK: Rentokil, Ltd.
Evans, H.E. (1958). The evolution of social life in wasps. Proceedings of the 10th International Congress of Entomology, 2, 449-457.
Evans, J.D., and Schwarz, R.S. (2011). Bees brought to their knees: Microbes affecting honey bee health. Trends in Microbiology, 19(12), 614-620.
Evans, J.D., and Spivak, M. (2010). Socialized medicine: Individual and communal disease barriers in honey bees. Journal of Invertebrate Pathology, 103, S62-S72.
Finch, C.E. (1990). Longevity, Senescence, and the Genome. Chicago, IL: The University of Chicago Press.
Gätschenberger, H., Gimple, O., Tautz, J., and Beier, H. (2012). Honey bee drones maintain humoral immune competence throughout all life stages in the absence of vitellogenin production. Journal of Experimental Biology, 215(8), 1313-1322.
Gätschenberger, H., Azzami, K., Tautz, J., and Beier, H. (2013). Antibacterial immune competence of honey bees (Apis mellifera) is adapted to different life stages and environmental risks. PLOS ONE, 8(6).
Haddad, L.S., Kelbert, L., and Hulbert, A.J. (2007). Extended longevity of queen honey bees compared to workers is associated with peroxidation-resistant membranes. Experimental Gerontology, 42(7), 601-609.
Haydak, M.H. (1963). Age of nurse bees and brood rearing. Minnesota Agricultural Experiment Station, 5122.
Helfand, S.L., and Rogina, B. (2003a). Genetics of aging in the fruit fly, Drosophila melanogaster. Annual Review of Genetics, 37, 329-348.
Helfand, S.L., and Rogina, B. (2003b). Molecular genetics of aging in the fly: Is this the end of the beginning? Bioessays, 25(2), 134-141.
Helft, F., Tirard, C., and Doums, C. (2012). Effects of division of labour on immunity in workers of the ant Cataglyphis cursor. Insectes Sociaux, 59(3), 333-340.
Herb, B.R., Wolschin, F., Hansen, K.D., Aryee, M.J., Langmead, B., Irizarry, R., Amdam, G.V., and Feinberg, A.P. (2012). Reversible switching between epigenetic states in honeybee behavioral subcastes. Nature Neuroscience, 15, 1371-1373.
Hölldobler, B., and E. O. Wilson. (1990). The Ants. Cambridge, MA: The Belknap Press of Harvard University Press.
Huang, Z.Y., and Robinson, G.E. (1992). Honeybee colony integration. Worker worker interactions mediate hormonally regulated plasticity in division-of-labor. Proceedings of the National Academy of Sciences of the United States of America, 39, 147-158.
Huang, Z.Y., and Robinson, G.E. (1996). Regulation of honey bee division of labor by colony age demography. Behavioral Ecology and Sociobiology, 39(3), 147-158.
Hulbert, A.J., Pamplona, R., Buffenstein, R., and Buttemer, W.A. (2007). Life and death: Metabolic rate, membrane composition, and life span of animals. Physiological Reviews, 87(4), 1175-1213.
Hunt, J.H. (1991). Nourishment and the evoluton of the social Vespidae. In K.G. Ross and R.W. Matthews (Eds.), The Social Biology of Wasps (pp. 426-450). Ithaca, NY: Comstock.
Hunt, J.H., and Amdam, G. V. (2005). Bivoltinism as an antecedent to eusociality in the paper wasp genus Polistes. Science, 308(5719), 264-267.
Hunt, G., Amdam, G., Schlipalius, D., Emore, C., Sardesai, N., and Williams, C. (2007). Behavioral genomics of honeybee foraging and nest defense. Naturwissenschaften, 94(4), 247-267.
Jeanne, R.L. (1975). The adaptiveness of social wasp nest architecture. Quarterly Review of Biology, 50, 267-287.
Jefferson, J.M., Dolstad, H.A., Sivalingam, M.D., and Snow, J.W. (2013). Barrier immune effectors are maintained during transition from nurse to forager in the honey bee. PLOS ONE, 8(1).
Johnson, B.R. (2010). Division of labor in honeybees: Form, function, and proximate mechanisms. Behavioral Ecology and Sociobiology, 64(3), 305-316.
Johnson, B.R., and Linksvayer, T.A. (2010). Deconstructing the superorganisms: Social physiology, ground plans and socioeconomics. Quarterly Review of Biology, 85, 57-79.
Keller, L. (1998). Queen lifespan and colony characteristics in ants and termites. Insectes Sociaux, 45, 235-246.
Keller, L., and Genoud, M. (1997). Extraordinary lifespans in ants: A test of evolutionary theories of ageing. Nature, 389, 958-960.
Laughton, A.M., Boots, M., and Siva-Jothy, M.T. (2011). The ontogeny of immunity in the honey bee, Apis mellifera L. following an immune challenge. Journal of Insect Physiology, 57(7), 1023-1032.
Lavine, M.D., and Strand, M.R. (2002). Insect hemocytes and their role in immunity. Insect Biochemistry and Molecular Biology, 32(10), 1295-1309.
Lee, R.D. (2003). Rethinking the evolutionary theory of aging: Transfers, not births, shape senescence in social species. Proceedings of the National Academy of Sciences of the United States of America, 100, 9637-9642.
Lemaitre, B., and Hoffmann, J. (2007). The host defense of Drosophila melanogaster. Annual Review of Immunology, 25, 697-743.
Lockett, G.A., Kucharski, R., and Maleszka, R. (2012) DNA methylation changes elicited by social stimuli in the brains of worker honey bees. Genes Brain and Behavior, 11(2), 235-242.
Marmot, M. (2004). The Status Syndrome. New York: Times Books.
Muller, F.L., Lustgarten, M.S., Jang, Y., Richardson, A., and Van Remmen, H. (2007). Trends in oxidative aging theories. Free Radical Biology and Medicine, 43(4), 477-503.
Munch, D., and Amdam, G.V. (2010). The curious case of aging plasticity in honey bees. FEBS Letters, 584(12), 2496-2503.
Munday, P.L., Buston, P.M., and Warner, R.R. (2006). Diversity and flexibility of sex-change strategies in animals. Trends in Ecology & Evolution, 21(2), 89-95.
Page, R.E.J. (2013). The Spirit of the Hive: Mechanisms of Social Evolution. Cambridge, MA: Harvard University Press.
Parker, J.D., Parker, K.M., Sohal, B.H., Sohal, R.S., and Keller, L. (2004). Decreased expression of Cu-Zn superoxide dismutase 1 in ants with extreme lifespan. Proceedings of the National Academy of Sciences of the United States of America, 101(10), 3486-3489.
Purves, W.K., Sadava, D., and Orians, G.H. (2004). Life: The Science of Biology (7th ed.). New York: W.H. Freeman.
Queller, D.C. (1989). The evolution of eusociality: Reproductive head starts of workers. Proceedings of the National Academy of Sciences of the United States of America, 86(9), 3224-3226.
Remolina, S.C., Hafez, D.M., Robinson, G.E., and Hughes, K.A. (2007). Senescence in the worker honey bee Apis Mellifera. Journal of Insect Physiology, 53(10), 1027-1033.
Roach, D.A., and Carey, J.R. (2014). The population biology of aging. Annual Review of Ecology, Evolution and Systematics, 45.
Robinson, E.J.H., Richardson, T.O., Sendova-Franks, A.B., Feinerman, O., and Franks, N.R. (2009). Radio tagging reveals the roles of corpulence, experience and social information in ant decision making. Behavioral Ecology and Sociobiology, 63(5), 627-636.
Sansoni, P., Vescovini, R., Fagnoni, F., Biasini, C., Zanni, F., and Zanlari, L. (2008). The immune system in extreme longevity. Experimental Gerontology, 43(2), 61-65.
Schmid, M.R., Brockmann, A., Pirk, C.W.W., Stanley, D.W., and Tautz, J. (2008). Adult honeybees (Apis mellifera L.) abandon hemocytic, but not phenoloxidase-based immunity. Journal of Insect Physiology, 54(2), 439-444.
Schneider, S.A., Schrader, C., Wagner, A.E., Boesch-Saadatmandi, C., Liebig, J., and Rimbach, G. (2011). Stress resistance and longevity are not directly linked to levels of enzymatic antioxidants in the ponerine ant Harpegnathos saltator. PLOS ONE, 6(1).
Seehuus, S.C., Krekling, T., and Amdam, G.V. (2006a). Cellular senescence in honey bee brain is largely independent of chronological age. Experimental Gerontology, 41(11), 1117-1125.
Seehuus, S.C., Norberg, K., Gimsa, U., Krekling, T., and Amdam, G.V. (2006b). Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proceedings of the National Academy of Sciences of the United States of America, 103(4), 962-967.
Seeley, T.D. (1995). The Wisdom of the Hive: The Social Physiology of Honey Bee Colonies. London, UK: Harvard University Press.
Starr, S.K. (1991). The nest as the locus of social life. In K.G. Ross and R.W. Matthews (Eds.), The Social Biology of Wasps (pp. 520-539). Ithaca, NY: Comstock.
Tatar, M., Kopelman, A., Epstein, D., Tu, M.P., Yin, C.M., and Garofalo, R.S. (2001). A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science, 292(5514), 107-110.
Toth, A.L., Kantarovich, S., Meisel, A.F., and Robinson, G.E. (2005). Nutritional status influences socially regulated foraging ontogeny in honey bees. Journal of Experimental Biology, 208(24), 4641-4649.
Vance, J.T., Williams, J.B., Elekonich, M.M., and Roberts, S.P. (2009). The effects of age and behavioral development on honey bee (Apis mellifera) flight performance. Journal of Experimental Biology, 212(16), 2604-2611.
Vaupel, J.W., Carey, J.R., Christensen, K., Johnson, T.E., Yashin, A.I., and Holm, N.V. (1998). Biodemographic trajectories of longevity. Science, 280(5365), 855-860.
Waite, L., and Das, A. (2010). Familes, social life, and well-being at older ages. Demography, 47, S87-S109.
Waite, L.J., and Lehrer, E.L. (2003). The benefits from marriage and religion in the United States: A comparative analysis. Population and Development Review, 29, 255-275.
Wegener, J., Huang, Z.Y., Lorenz, M.W., and Bienefeld, K. (2009). Regulation of hypo-pharyngeal gland activity and oogenesis in honey bee (Apis mellifera) workers. Journal of Insect Physiology, 55(8), 716-725.
Wenzel, J.W. (1991). Evolution of nest architecture. In K.G. Ross and R.W. Matthews (Eds.), The Social Biology of Wasps (pp. 480-519). Ithaca, NY: Comstock.
Williams, J.B., Roberts, S.P., and Elekonich, M.M. (2008). Age and natural metabolically-intensive behavior affect oxidative stress and antioxidant mechanisms. Experimental Gerontology, 43(6), 538-549.
Wilson-Rich, N., Dres, S.T., and Starks, P.T. (2008). The ontogeny of immunity: Development of innate immune strength in the honey bee (Apis mellifera). Journal of Insect Physiology, 54(10-11), 1392-1399.
Wilson-Rich, N., Spivak, M., Fefferman, N.H., and Starks, P.T. (2009). Genetic, individual, and group facilitation of disease resistance in insect societies. Annual Review of Entomology, 54, 405-423.
Wilson, E.O. (1971). The Insect Societies. Cambridge, MA: The Belknap Press of Harvard University Press.
Wilson, E.O. (1985). The sociogenesis of insect colonies. Science, 228, 1489-1495.


























