
2
The Environmental Setting
THE PHYSICAL ENVIRONMENT
To place the decreased abundance of Steller sea lions (Eumetopias jubatus) in context with changes in the sea lion’s environment, this chapter describes salient features of the physical and biological regimes of the North Pacific. Steller sea lions in the endangered western stock live throughout the Gulf of Alaska (west of ~144° W), Aleutian Islands, and eastern Bering Sea (Figure 2.1). Climate variability and oceanic responses to climate affect biota, including sea lions, through a number of pathways and mechanisms (Francis et al., 1998; Schumacher and Alexander, 1999; Schumacher et al., in press). Because of severe limitations in data and understanding, researchers face a number of challenges when attempting to identify cause and effect relationships between climatic-environmental changes and changes in marine species such as Steller sea lions (see Benson and Trites, 2002, for a comprehensive review). In general, changes in the physical environment may affect the abundance and types of prey available to Steller sea lions through nutrient-phytoplankton-zooplankton dynamics and responses of higher-trophic level biota such as forage fish and groundfish. Physical perturbations, particularly those in upper-ocean temperature, could affect Steller sea lions by changing the distribution and abundance of sea lion prey, competitors, or predators.
The physical environment in the western range of the Steller sea lions is profoundly influenced by atmospheric phenomena (e.g., wind stress, heat flux, precipitation). Frequent fall and winter storms (low-pressure
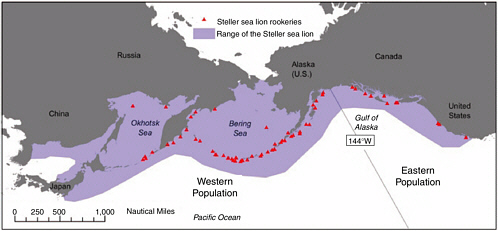
FIGURE 2.1 The range (in purple ) and rookeries (in red) of Steller sea lions. 144° W defines the boundary between the eastern and western populations.
SOURCE: National Marine Fisheries Service, Alaska Fisheries Science Center ( www.afsc.noaa.gov/stellers/range.htm ).
systems) passing across the Aleutians and midlatitudes of the North Pacific produce the climatological Aleutian Low pressure cell. Averaged over one to several months, the Aleutian Low pressure pattern exhibits strong intraseasonal, interannual, and interdecadal variability (Trenberth, 1990; Trenberth and Hurrell, 1994). The intensity of the fall/winter Aleutian Low is correlated with El Niño-Southern Oscillation variations (primarily on a 2- to 7-year timescale) and also varies as an element of the Pacific Decadal Oscillation (PDO) at timescales as long as 50 to 70 years (Minobe, 1997). The Aleutian Low varies in association with the Arctic Oscillation over a broad spectrum of timescales (interannual to interdecadal; Overland et al., 1999).
In the North Pacific and Bering Sea, 20th-century climatic regimes persisted for periods ranging from 10 to 30 years (Francis and Hare, 1994; Ware, 1995; Mantua et al., 1997; Minobe, 1997). Tree-ring reconstructions for coastal Alaska and North Pacific surface temperatures provide evidence of multidecadal climatic regime shifts over the past three centuries, though the amplitude of this variability was weak in the mid-1980s (e.g., Ingraham et al., 1998; D’Arrigo et al., 2001; Gedalof and Smith, 2001). Studies of tree-ring records (Ingraham et al., 1998) and nitrogen isotopes in salmon remains (Finney et al., 2000) have identified periodic changes in the climate and biota at least as far back as the 1500s. The PDO represents one aspect of a regime shift, which is manifested in the patterns of sea surface temperature throughout the Pacific Ocean north of 20° N. Recent regime shifts occurred in 1976-1977, 1989, and likely in the late 1990s (see Figure 2.2). Despite strong year-to-year climate variability in both the North Pacific and Bering Sea, there remains compelling evidence for coherent interdecadal variations in the large marine ecosystems of the North Pacific and Bering Sea (at least as indicated by collections of fishery and ecosystem survey records from 1965 to 1997). Hare and Mantua (2000) speculate that aspects of North Pacific and Bering Sea ecosystems have responded nonlinearly to “noisy” external forces like those associated with strong year-to-year climate variations that are ubiquitous in midlatitudes.
In subarctic regions, sea ice is an important feature that links atmospheric changes to the ocean. Air-sea heat exchange and wind stress dictate the formation, transport, and persistence of sea ice. In the Gulf of Alaska and the Aleutian Islands, sea ice forms in small areas: the greatest coverage occurs in Cook Inlet (Wilson and Overland, 1986) and plays a minor role in local ecosystem dynamics. In the eastern Bering Sea, variations in sea ice extent, time of advance, and subsequent disappearance either by melting in place or being blown away are striking physical phenomena (Schumacher and Stabeno, 1998). The strongest signal occurs at interannual periods, although changes also occur during regime shifts
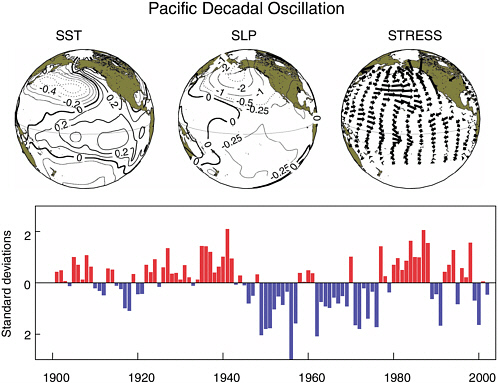
FIGURE 2.2 Anomalous climate conditions associated with warm phases of the Pacific Decadal Oscillation (PDO). Sea surface temperature (SST) values are given in degrees Celsius, sea level pressure (SLP) is in millibars, and surface wind stress direction and intensity are indicated by arrows where the longest wind vector represents a pseudostress of 10 m2/s2. The lower graph presents November-March average values of the PDO index during the 20th century. Actual anomaly values for a given year at a given location are obtained by multiplying the climate anomaly by the associated index value (Reprinted from Mantua and Hare, 2002, with kind permission of Kluwer Academic Publishers).
(Brodeur et al., 1999; Stabeno and Overland, 2001). The changes in sea ice characteristics have profound impacts on biota (Schumacher et al., in press). A bloom of phytoplankton is associated with the spring sea ice and accounts for 10 to 65% of the total annual primary production (Niebauer et al., 1995). In years when sea ice is either not present or retreats before adequate light is available (early March), the conventional spring bloom is delayed until late spring (Stabeno et al., 2001). The newly described oscillating control hypothesis (OCH) relates ecosystem changes to changes in atmospheric and oceanic features of the southeastern Bering Sea (Hunt et al., 2002). In essence, the OCH states that during cold physical
regimes the ecosystem is driven in a bottom-up manner, whereas during warm regimes it is in a top-down mode. Under the OCH, cold regimes are characterized by late retreat of sea ice and low water column temperatures. At low temperature, survival of fish eggs is reduced and there is less zooplankton prey for larval fish, resulting in reduced recruitment to adult populations of pollock and other fishes.
Bottom-up processes generally dictate the flow of energy through the ecosystem during a cold regime. Because low water column temperatures can change the distribution of some forage fish species, the OCH explains why Steller sea lions, other pinnipeds, and piscivorous seabirds may thrive even under cold conditions if forage fish become more available as prey. Warm regimes occur during years when sea ice is either absent or retreats before there is sufficient sunlight to initiate an under-ice bloom. In this case, the spring bloom occurs later, water column temperatures are warmer, and higher zooplankton production provides prey for larval and juvenile fish. This should favor recruitment and strong year classes of pollock and other piscivorous fish.
There are years when the observed regime conditions (e.g., 1976, cold with low recruitment; 1996, warm with high recruitment) and estimated pollock production fit the outcomes predicted by the OCH; however, there are also years that do not fit this pattern (e.g., 1987, warm with low recruitment; 1992, cold with high recruitment). The following scenario explains how the OCH could affect the abundance of fish stocks through top-down control by predators. Adult pollock eat age 1 and younger fish and together with predation by other fish (e.g., arrowtooth flounder and Pacific cod) may depress the size of new year classes (Livingston and Methot, 1998; Livingston et al., 1999).
When there is a sequence of warm regime years, recruitment will be above average and the populations of adult predatory fish will increase to a point where the control of future year-class strength will be mainly a top-down process. In the Gulf of Alaska, for example, expansion of the arrowtooth flounder population correlates with a severe decline in the pollock population (Bailey, 2000). If predation on forage fish increases, zooplankton will become available for other populations (e.g., jellyfish, salmon, baleen whales). Farther up the food web, the reduced abundance of forage fish could affect the feeding success and survival of pinnipeds and piscivorous seabirds.
Although decadal timescale or regime shifts have been observed in the Bering Sea (e.g., Stabeno et al., 2001; Minobe, 2002), much of the variance is in the annual cycle (Royer et al., 2001). The OCH addresses the impact of annual changes in the following manner. If a cold year occurs during a warm regime, the existing stock of predatory adult fish will maintain top-down control of prey fish, reducing recruitment of the next
year class. During a cold regime, stocks of piscivorous predators (e.g., pollock) will be relatively low; hence, recruitment will mostly depend on bottom-up processes. One or two warm years could yield strong year classes of piscivores that could then switch the system from bottom-up to top-down control by predation. Variations in forage fish and pollock year class strength could also occur within a regime if predators become redistributed.
In addition to the effects on bottom-up forcing of energy flow in the pelagic ecosystem, ice and its attendant cold pool of water directly influence distributions of higher-trophic-level biota such as marine mammals by providing habitat (Ohtani and Azumaya, 1995; Wyllie-Echevarria and Wooster, 1998; Brodeur et al., 1999).
The coastline of the Gulf of Alaska is marked by a nearly continuous mountain barrier that results in topographic uplift high winds and substantial precipitation (Wilson and Overland, 1986; Royer, 1998; Royer et al., 2001; Stabeno et al., in press). The continental shelf relevant to the western stock of Steller sea lions is generally about 150 km wide with many troughs and underwater canyons. Unlike the shallow eastern Bering Sea shelf, in the Gulf of Alaska depths greater than 150 m often occur close to the coast. Numerous islands provide habitat for the Steller sea lion. The Alaska Current flows westward and becomes a strong boundary current, the Alaskan Stream off Kodiak Island (Reed and Schumacher, 1986). The dominant current over the shelf is the Alaskan Coastal Current (ACC), which flows generally westward along the coastline (Schumacher and Reed 1986; Stabeno et al., 1995; Royer, 1998; Stabeno et al., in press) and is generated by a combination of wind-driven convergence along the coast and freshwater runoff. The ACC is a continuous feature westward to Unimak Pass, where it flows northward to the Bering Sea shelf (Reed and Schumacher, 1986). The transport of nutrients from the oceanic waters occurs in the Shelikof Strait with the mixing of fresh water and seawater as the ACC flows seaward (Reed et al., 1987). Other mechanisms that renew nutrients on this shelf include flow along the deep troughs, oceanic eddies, and an Ekman flux (Stabeno et al., in press). The latter process results from the mean winds generating upwelling in the central North Pacific Ocean that is then driven toward the Alaskan coastline in the upper or Ekman layer of the water column.
The Aleutian Islands form a porous boundary between the North Pacific Ocean and the Bering Sea, with a narrow continental shelf. Strong tidal currents mix the northward-flowing waters, which can then provide nutrients to the euphotic zone. Adjacent to the Aleutian Islands, the dominant current to the south is the Alaskan Stream, which provides relatively fresh surface and warm subsurface waters to the Bering Sea. The flow northward through Amchitka and Amukta Pass is primarily
responsible for supplying the Aleutian North Slope Current (Reed and Stabeno, 1999), which flows eastward along the northern side of the Aleutian Islands and provides the main source of the Bering Slope Current. The importance of these currents to chemical and planktonic material is threefold. First, they provide transport from oceanic to shelf waters, a feature potentially important for pollock that spawn near Bogoslof Island. Second, their eddies can temporarily provide a habitat that favors survival of larvae (Schumacher and Stabeno, 1994). Third, their relatively warm temperature characteristics are potentially important to fish stocks by increasing egg maturation rates and thus reducing time for predation on that life history stage (Reed, 1995).
The continental shelf of the southeastern Bering Sea is broad (~500 km) and relatively shallow (<180 m) with canyons incising the shelf break. Compared to the Gulf of Alaska, the bathymetry of this shelf is featureless. Unlike the Gulf or Aleutian Islands, circulation over the shelf proper is generally sluggish. There is a moderate flow toward the northwest over the outer shelf in the vicinity of the 100-m isobath, and a weaker feature exists around the perimeter of the eastern shelf flowing east and then northward in the general vicinity of the 50-m isobath (Schumacher and Stabeno, 1998). Tidal currents mix the waters of the coastal domain and create a bottom mixed layer over the remainder of the shelf. Low-frequency currents around some of the islands and in submerged canyons are generated as water masses encounter submerged topographic features (Schumacher and Stabeno, 1998; Kowalik, 1999; Stabeno et al., 1999).
THE BIOLOGICAL ENVIRONMENT
It is now generally recognized that ecosystems can shift between different states and that these shifts often involve gradual changes in a system until a threshold is reached and reorganization of the ecosystem occurs (Francis and Hare, 1994; Anderson and Piatt, 1999; Scheffer et al., 2001). Recent studies indicate that high-latitude Arctic ecosystems may be particularly vulnerable to these regime shifts, with these transitions potentially happening every few decades, especially when they are associated with decreases in sea ice extent (Aagaard and Carmack, 1989; Manabe et al., 1991; Lynch et al., 1995; Hu et al., 1999). The allocation of carbon fixation to respiration, storage, and export, as mediated by coupling of pelagic and benthic carbon cycles, is of major interest (Walsh et al., 1989; Walsh, 1995; Rivkin and Legendre, 2001). Although hemispheric climate variability has been tied to regime shifts, threshold responses in the regional Gulf of Alaska and/or Bering Sea biological ecosystem are more elusive yet critical to understanding the environmental factors potentially acting on the decline of Steller sea lion populations in the region.
Large-scale climate and ecosystem changes have been documented in the North Pacific and Gulf of Alaska (Francis et al., 1998; Hare and Mantua, 2000) and the Bering Sea (Mantua et al., 1997; Minobe, 1997; Scheffer et al., 2001) in 1925, 1947, 1977, and 1989 (Beamish et al., 1999; Overland et al., 1999). It has been suggested that these changes in biological and physical variables have ecosystem-wide consequences as seen in changes of food web structure and carbon cycling (Beamish et al., 1999; Hare and Mantua, 2000). These shifts have had widespread and important impacts on commercial fisheries, sea birds, marine mammals, and Alaska natives who harvest these resources (National Research Council, 1996; Callaway et al., 1998; Criddle et al., 1998).
Changes in North Pacific and Gulf of Alaska Biology
Oceanic and atmospheric interactions in the North Pacific and Gulf of Alaska have a direct influence on water column productivity. Intensification of regional winds is believed to enhance primary productivity by increasing the divergence of surface water in the center of the Alaska Gyre, enhancing the upwelling of nutrient-rich water. Surface water warming and enhanced stability would keep the phytoplankton in more favorable light conditions in the euphotic zone, thus potentially boosting primary production when light is the limiting factor. Copepod biomass also increased after the 1976-1977 regime shift, compared to the 1950s to mid-1970s, likely due to an increase of the Alaska Gyre and an intensification of the Aleutian Low (Brodeur and Ware, 1992).
Historical records tracking aspects of Pacific marine ecosystems suggest a strong association between the PDO variability and Pacific salmon production (Beamish and Bouillon, 1993; Beamish et al., 1999; Hare et al., 1999), Pacific sea birds (Vandenbosch, 2000), Alaska groundfish and zooplankton production in the central and eastern North Pacific (Francis et al., 1998; Hollowed et al., 1998), and Gulf of Alaska marine species assemblages (Anderson and Piatt, 1999), to name just a few (see Figure 2.3). The small-mesh Gulf of Alaska trawl survey data analyzed by Anderson and Piatt show an apparent major redistribution in the relative abundance of species closely associated with the 1977 regime shift (see Figure 2.4).
As the relatively cool era of the early 1970s switched to a relatively warm era (in Gulf of Alaska sea surface temperatures and ocean bottom temperatures and wintertime coastal Alaska air temperatures), data on catch per unit effort indicate that shrimp and small pelagic fish became scarce concurrently with a dramatic increase in the biomass of gadids and flatfish (Ronholt et al., 1978; Alverson, 1992; Anderson and Piatt, 1999; Mueter and Norcross, 2000).
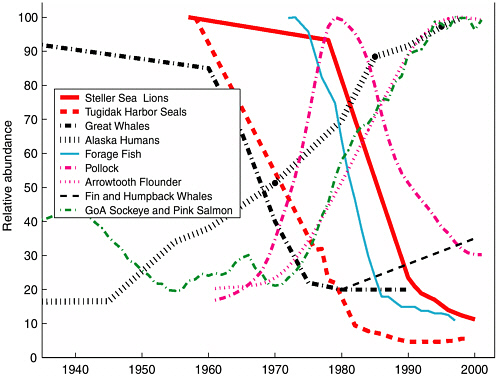
FIGURE 2.3 Gulf of Alaska ecosystem trends: 1945-2000. Schematic diagram indicates the relative abundance of select marine mammals, fish, and shellfish in the Gulf of Alaska. To clarify multiyear trends a 10-year running average was applied to groundfish, forage fish, and Tugidak harbor seal molt count data.
SOURCES: Data for blue, fin, sei, sperm, and humpback whales (key = Great Whales) came from National Research Council (1996). Fin and humpback whale population trends were updated with data from Robyn Angliss and Sue Moore from NMFS National Marine Mammal Lab, Seattle, WA, personal communication. Data for groundfish (pollock and arrowtooth flounder) came from NMFS 2001 stock assessments (www.refm.noaa.gov/docs/). Steller sea lion data came from NMFS stock assessments (www.refm.noaa.gov/docs/). Human population came from Alaska census data (www.labor.state.ak.us/research/popover.htm). The 1958 data point for Tugidak harbor seal beach molt counts came from Douglas DeMaster (National Marine Fisheries Service, Marine Mammal Lab, Seattle, personal communication, 2002). All other data for harbor seals came from L.A. Jemison and G.W. Pendleton, Harbor Seal Population Trends and Factors Influencing Counts on Tugidak Island, Alaska, an unpublished manuscript provided by Bob Small of the Alaska Department of Fish & Game, November, 2002. Data for Gulf of Alaska forage fish catch per unit effort was estimated from the NMFS’s small-mesh trawl survey data (kg/km) for all shrimps, capelin, smelts, sandfish, herring, and juvenile pollock <20 cm (www.fakr.noaa.gov/trawl/index.htm). The catch data for the 10-year running average of central Gulf of Alaska sockeye and pink salmon were taken from Hare et al. (1999). Sockeye and pink account for up to 90% of the total annual salmon landings in the region.
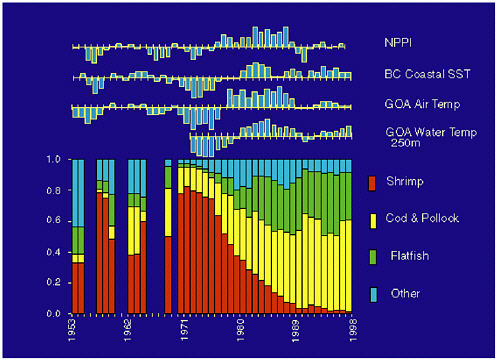
FIGURE 2.4 Proportional species distribution in small-mesh trawl catches in the Gulf of Alaska between 1953 and 1997 relative to climate indices. Climate data expressed as normalized anomalies. NPPI, North Pacific Pressure Index; BC SST, British Columbia sea surface temperature from lighthouse stations; trends were smoothed by taking 3-year running averages. SOURCE: Reproduced from Anderson and Piatt (1999).
Changes in Bering Sea Biology
The Bering Sea also reflects large-scale circulation and regime shifts, although not necessarily the timing and extent of changes observed in the Gulf of Alaska and vice versa. There was an observed cooling era in the Bering Sea from 1965 to 1978, the main 1977-1978 regime shift, and a warming period from 1979 to 1989 (Wooster and Hollowed, 1995). Niebauer (1998), using sea ice as an indicator of Bering Sea condition, found a significant decrease in ice cover from 1977 into the late 1980s. Recent studies indicate a trend toward earlier retreat of ice in the spring during the 1990s (Stabeno and Overland, 2001), which may have a dramatic impact on ice edge production and associated ecosystem response in the spring (Hunt et al., 2002). Coincidently, over the past 30 years, the Bering Sea has exhibited substantial changes in both pelagic and benthic community structure (National Research Council, 1996). Major environmental
changes in the Bering Sea include increased water temperature, possibly influencing the observed increase in zooplankton biomass (Brodeur and Ware, 1992; Sugimoto and Tadokoro, 1997; Brodeur et al., 1999, 2002) invasion by warm-water organisms and a 10-fold increase in gelatinous zooplankton between 1979 and 1997 (Brodeur et al., 1999), and more frequent blooms of coccolithophorid phytoplankton, which can cause substantial mortality of endotherm predators (Baduini et al., 2001; Lovvorn et al., 2001). Various marine birds and mammals have declined precipitously (see Figure 2.5), with food web changes suggested as the likely cause (Springer, 1998, 2001).
Similar ecosystem-level change has been observed in the northern Bering Sea, with indications of benthic biomass changes south of St. Lawrence Island, in the Chirikov Basin between St. Lawrence Island and Bering Strait, and in the southern Chukchi Sea (Grebmeier and Cooper, 2002). Studies begun in the mid-1980s have shown declines in the biomass (Sirenko and Koltun, 1992; Grebmeier, 1993; Grebmeier and Dunton, 2000) and mean sizes of the dominant bivalves in the area (Grebmeier and Cooper, 2002). Sediment respiration rates, which indicate carbon loading to the seafloor, have also declined since the late 1980s. Seasonal patterns of sediment chlorophyll concentrations show that deposition of carbon in this area is closely tied to the ice-edge spring bloom (Cooper et al., 2002); thus, any change in the timing of ice retreat during the late winter/early spring will likely have a major impact on ecosystem structure. Although commercial fishing may have played a role in the trophic changes in the southern Bering Sea, there is little commercial fishing on the northern shelf, where changes in benthic faunal populations and declines in dominant fauna have occurred, with cascading effects on higher trophic levels (Grebmeier and Dunton, 2000; Grebmeier and Cooper, 2002; Lovvorn et al., in press).
Studies by Wyllie-Echevarria and Wooster (1998) have related the extent and duration of the cold pool on the middle shelf of the Bering Sea with the location of fish stocks. The cold pool (<2° C) occurs subsurface (50 to 100 m) and is remnant winter water that is maintained through the summer. Only after the 1977 regime shift (and reduced ice extent and duration) were pollock larvae observed in the northern Bering Sea, with pollock larvae observed north of the Bering Strait in the Chukchi Sea in 1988. Only Arctic cod (Boreogadus saida) occur regularly in the cold pool, whereas the subarctic walleye pollock are variable, moving to the outer continental shelf in cold years (when the cold pool expands) but dispersing across the middle domain in warmer years (when the cold pool is smaller). Recent stock assessments (North Pacific Fishery Management Council, 2001b), based largely on annual bottom trawl surveys of the eastern Bering Sea shelf, are generally consistent with the regime shift
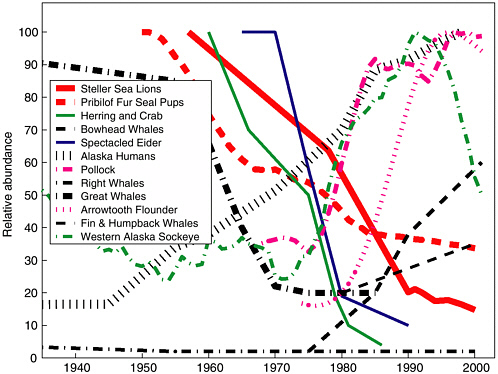
FIGURE 2.5 Bering Sea ecosystem trends: 1945-2001. Schematic diagram indicates the relative abundance of select marine mammals, fish, and shellfish from 1945 to 2001 in the Bering Sea/Aleutian Islands (BSAI). To clarify multiyear trends a 10-year running average was applied to groundfish, forage fish, fur seal pups, and western Alaska sockeye salmon.
SOURCES: Data for blue, fin, and sei and humpback whales (key = Great Whales), and right and bowhead whales are taken from National Research Council (1996). Data from 1996-2002 on bowhead, fin, and humpback whale population trend data came from Robyn Angliss, National Oceanic and Atmospheric Administration, Marine Mammal Lab in Seattle, personal communication, 2002. Data for the northern fur seal annual pup counts on the Pribilofs came from Anne York, National Marine Fisheries Service, Marine Mammal Lab, Seattle, personal communication, 2002. Spectacled eider (Somateria fischeri) and herring and crab (except Opilio) data were estimated from National Research Council (1996, Figure 6.8). Steller sea lion, pollock, and arrowtooth flounder data came from National Marine Fisheries Service 2001 stock assessments (www.refm.noaa.gov/docs/). Sockeye salmon catch data are taken from Hare et al. (1999). Sockeye account for up to 75% of the total annual salmon landings in this region. Human population came from Alaska census data (www.labor.state.ak.us/research/pop/popover.htm).
patterns revealed by Anderson and Piatt (1999) for the Gulf of Alaska. In particular, many demersal fish species (e.g., flatfishes, pollock, cod) increased in the late 1970s to early 1980s in conjunction with declines in some crab species (Zheng and Kruse, 2000), together with anecdotal evidence of declines in shrimp and forage fishes.













