
5
Fisheries
GROUNDFISH FISHERIES
Introduction
Fisheries conducted off Alaska’s extensive 7,500-mile coastline can be divided into three very large regions—Gulf of Alaska, Aleutian Islands, and eastern Bering Sea (see Figure 5.1). The Gulf of Alaska is subdivided into eastern, central, and western areas. These areas are further subdivided into smaller federal reporting areas and even finer state statistical areas. A region of international waters exists in the central Bering Sea, beyond 200 nautical miles (nm) from the U.S. and Russian coasts, which is often termed the “Donut Hole.” These designations differ from those used to describe the distribution of Steller sea lions.
Groundfish fisheries are of particular interest with respect to Steller sea lions. These fisheries are the main focus of this chapter because groundfish species comprise a large fraction of the sea lion diet (see Chapter 4). Groundfish include many fish species that are primarily distributed on or near the seafloor. The primary Alaskan groundfish species of commercial interest include walleye pollock (Theragra chalcogramma); Pacific cod (Gadus macrocephalus); Atka mackerel (Pleurogrammus monopterygius); sablefish (Anoplopoma fimbria); rockfish, especially Pacific ocean perch (Sebastes alutus) and several other rockfish species; and yellowfin sole (Pleuronectes aspera), and other flatfishes, including Pacific halibut (Hippoglossus stenolepis), which are not considered groundfish in the reporting of catch
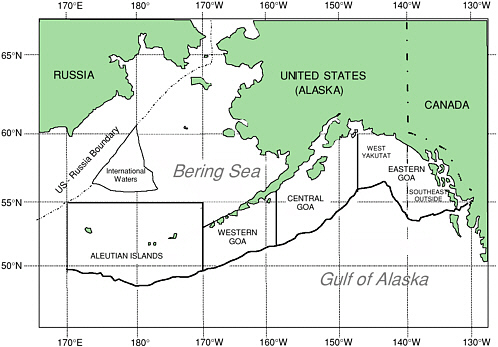
FIGURE 5.1 This map shows the principal federal regulatory areas for groundfish fisheries off the coast of Alaska extending to 200 nm offshore (solid wavy lines). The region in the central Bering Sea labeled “International Waters” is commonly referred to as the Donut Hole.
SOURCE: 50 CFR 679, Figure 14. Available from www.fakr.noaa.gov/rr/figures.htm, Sablefish Regulatory Areas and Districts.
statistics. Currently, pollock, Atka mackerel, and flatfish (except Greenland halibut, Reinhardtius hippoglossoides, which are caught mostly by longline) are harvested nearly entirely by trawl gear. Depending on the species, rockfish are taken largely by trawls, with smaller amounts taken on longline and jig gear. Pacific cod are harvested mostly by trawls and to a lesser extent by longlines, pots, and jig gear. Pacific halibut and sablefish are taken by longlines. Species abundance and the magnitude of the fisheries vary spatially. For instance, the Bering Sea accounts for more than 90% of Alaskan pollock harvests, whereas Atka mackerel are taken almost exclusively in the Aleutian Islands. For Pacific cod, the largest fishery takes place in the Bering Sea, although significant fisheries occur in the Gulf of Alaska and Aleutian Islands as well.
Given the diversity and complexity of commercial fisheries in the vast area covered by the Gulf of Alaska, Aleutian Islands, and Bering Sea, it is not possible to completely describe each fishery in this report. Instead, the
focus is on particular aspects of fisheries that may be most relevant to considerations of the decline in sea lions. In the rest of this chapter, the strategy is first to review the history of the groundfish fishery with respect to Steller sea lion trends and, second, to describe the status of groundfish stocks and exploitation rates used to prescribe catch quotas under federal fishery management plans. Third, the spatial distribution of fisheries is considered, particularly with respect to the localized depletion hypothesis. Then, a brief overview is provided of some fisheries managed by the state of Alaska. Finally, management measures aimed at reducing potential competition between fisheries and Steller sea lions are summarized.
Fishery History with Reference to Steller Sea Lion Trends
Alverson et al. (1964), Alverson (1992), Kruse et al. (2000), Rigby et al. (1995), and the National Marine Fisheries Service (2002) reviewed the history of groundfish fishing in Alaska. A brief overview is provided here. Although the focus of this chapter is the groundfish fishery, it is important to note that other species were very important in the early development of commercial fisheries in Alaska. In the Gulf of Alaska, Russian, Japanese, and domestic shrimp trawl fisheries developed in the early 1960s. By 1972, this fishery was fully Americanized. Domestic landings peaked in the mid-1970s, and shrimp stocks collapsed in the early 1980s. A Japanese trawl fishery developed in the Bering Sea in the later 1950s, peaked in the early 1960s, and ended in the later 1970s. Attempts to develop a domestic shrimp fishery in the Bering Sea failed, owing to depletion of shrimp resources. The histories of fisheries for salmon, crab, and herring are reviewed later in this chapter.
Gulf of Alaska
Groundfish fisheries have been conducted off the Alaskan coast for nearly 150 years. In the 1860s a domestic schooner-based dory fishery using hand lines began for Pacific cod at various locations in the Gulf of Alaska (Bean, 1887). In the 1880s a longline fishery, conducted by U.S. fishermen, was developed for Pacific halibut and then sablefish in the inside waters of southeastern Alaska (Rigby et al., 1995). The sablefish fishery remained relatively unimportant until the mid-1930s, but halibut were depleted on some fishing grounds by 1916, leading to conservation concerns. In the 1960s Japanese longline fisheries developed for Pacific halibut, cod, and sablefish. Effort increased in the 1970s, and Korean vessels also participated in longline fisheries in the early 1970s.
Russian and, to a lesser extent, Japanese trawl vessels heavily exploited Pacific ocean perch (a slope rockfish species) in the early 1960s
and 1970s, with a peak catch of 350,000 metric tons (mt) in 1965 (see Figure 5.2). This heavy exploitation occurred during a period when Steller sea lion populations were considered healthy and Pacific ocean perch rarely appeared in the sea lion diet. After a period of pulse fishing, Pacific ocean perch were overfished in the Gulf of Alaska, and Russian fleets moved south to exploit rockfish stocks off Oregon and Washington. High trawl bycatch, accounting for one-third of the total domestic longline halibut harvest in 1965, caused concern among domestic fishermen. Groundfish landings grew steadily through the 1970s and into the 1980s.
With the passage of the Magnuson Fishery Conservation and Management Act of 1976, the U.S. declared management authority over the Exclusive Economic Zone (EEZ) to 200 nm offshore, and the total allowable level of foreign fishing was limited to that portion of the optimum yield that was not expected to be harvested by domestic vessels. Americanization was promoted by a “Fish and Chips” amendment to the Magnuson Fishery Conservation and Management Act in 1980, which significantly raised foreign fishing fees and tied foreign fishing privileges to commitments to purchase products by the developing U.S. industry. The years 1978-1990 were a transitional period in which a number of joint foreign-
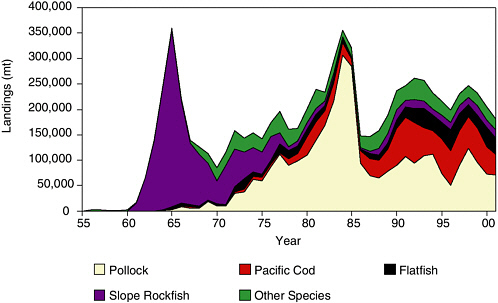
FIGURE 5.2 Groundfish landings (metric tons) from the Gulf of Alaska, 1956-2001.
SOURCE: Graph prepared from data in Table 5, North Pacific Fishery Management Council (2002).
U.S. ventures were established whereby domestic fishermen operated catcher vessels that delivered to foreign processing motherships. A prime example was the pollock fishery in Shelikof Strait, a large body of water between Kodiak Island and the Alaska Peninsula in the central Gulf of Alaska (see Figure 2.1). After spawning aggregations were discovered there, a large joint-venture trawl fishery developed from 1981 to 1985 with primary emphasis on roe harvest and surimi production. Landings from this fishery alone peaked at 300,000 mt in 1984, when pollock accounted for 86% of all groundfish landings in the Gulf of Alaska. Pollock biomass declined significantly during 1982-1986, and landings were cut sharply starting in 1986. The pollock fishery became fully domestic in 1988, and all foreign and joint-venture fisheries were phased out of Alaska by 1991.
Shelikof Strait lies in the central Gulf of Alaska portion of the sea lion distribution. Although index counts of nonpup Steller sea lions declined 23% during 1976-1985 when the Shelikof Strait fishery was most intense, sea lions index counts for this region declined most sharply (63%) during 1985-1990, a period of reduced pollock harvest (landings averaged 79,500 mt annually during 1986-1990). Potential connections between this fishery and declines of Steller sea lions are discussed later in this chapter and also in the context of the “Human-Related Steller Sea Lion Kills” section in Chapter 6.
From 1990 to 2001, Steller sea lions continued to decline in the Gulf of Alaska, albeit at a reduced rate. During this same period, groundfish landings were relatively stable and averaged 229,000 mt, of which pollock averaged 91,000 mt, Pacific cod averaged 64,000 mt, flatfish averaged 30,000 mt, and the remainder was comprised mainly of sablefish and rockfishes. Perhaps the most significant fishery trend during this period was a doubling of flatfish landings, largely due to increased global demand and new foreign purchasing arrangements for domestically caught groundfish.
Most groundfish fisheries off Alaska are managed by the National Marine Fisheries Service (NMFS) under federal fishery management plans adopted by the North Pacific Fishery Management Council (NPFMC). For most groundfish fisheries in territorial waters (0 to 3 nm), state regulations parallel federal management actions. In some other instances, Alaska’s Department of Fish & Game establishes separate catch quotas and other regulations for groundfish fisheries in state waters under management plans adopted by the Alaska Board of Fisheries. The state has lead management authority over lingcod (Ophiodon elongatus), black rockfish (Sebastes melanops), and blue rockfish (S. mystinus) throughout all waters of the territorial sea and the EEZ. Although these state-managed groundfish fisheries are important locally, they account for a small frac-
tion of the total groundfish landings. In 2000 the largest state-managed fisheries were conducted for Pacific cod, and they accounted for 5% (12,265 mt) of the total (244,193 mt) cod landings off Alaska (Kristin Mabry, Alaska Department of Fish & Game, Division of Commercial Fisheries, Juneau, personal communication, 2001).
Aleutian Islands
As in the Gulf of Alaska, the foreign trawl fishery depleted stocks of Pacific ocean perch in the Aleutian Islands in the 1960s and 1970s (see Figure 5.3). Landings of this species peaked at 115,000 mt in 1965. In the late 1970s a relatively small fishery developed for pollock, primarily in the eastern Aleutian Islands (Fritz, 1993). During 1980-1988 the fishery averaged 54,000 mt, peaked at 156,000 mt in 1989, and declined steadily from 74,000 mt in 1990 to 24,000 mt in 1998. In the late 1990s this fishery was increasingly constrained by regulations to avoid possible adverse effects on Steller sea lions. As a precautionary measure, the pollock fishery in the Aleutian Islands has been halted since the 1998 season.
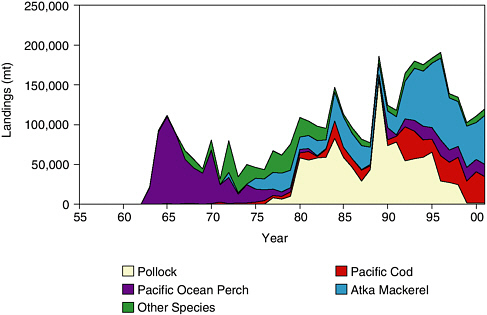
FIGURE 5.3 Groundfish landings from the Aleutian Islands, 1962-2001.
SOURCE: Graph prepared from data in Table 3, North Pacific Fishery Management Council (1999).
A foreign fishery developed for Atka mackerel in the 1970s with mean annual landings of 13,000 mt during 1972-1979. Joint-venture fisheries dominated the 1980s with average landings of 24,000 mt, but in 1990 U.S. domestic fishermen took over. Atka mackerel became the most important groundfish fishery in the Aleutian Islands in the 1990s as landings increased from 47,000 mt in 1992 to 103,000 mt in 1996 and averaged 55,000 mt during 1997-2000 (see Figure 5.3). The fishery is conducted in scattered locations, but most catches are taken from the central Aleutian Islands (Fritz, 1993).
In the 1960s, Japanese longline fisheries developed for Pacific halibut, cod, and sablefish. Effort increased in the 1970s, and Korean vessels also participated in longline fisheries in the early 1970s.
It is difficult to closely match periods of fishery development with periods of sharp declines of Steller sea lions in the Aleutian Islands. Declines of Steller sea lions appeared to begin in the eastern Aleutian Islands where nonpup index counts declined 61% during 1977-1985. Indeed, a significant pollock fishery first developed there during 1980-1985. However, pollock landings peaked in 1989 and continued at 1980-1985 levels through 1995, but sea lion index counts for this region increased 55% during 1989-1996. Likewise, sea lion counts in the central Aleutian Islands declined the sharpest (67%) during 1985-1989, and the Atka mackerel fishery has occurred in this area since the late 1970s (Fritz, 1993), but landings did not increase markedly until the 1990s.
During 1990-2001, groundfish landings in the Aleutian Islands averaged 145,000 mt per year. Average annual landings were dominated by Atka mackerel (58,000 mt), pollock (39,000 mt), cod (27,000 mt), and Pacific ocean perch (14,000 mt).
Bering Sea
Japanese vessels conducted exploratory groundfish fisheries in the Bering Sea in 1929 and 1931. A fishery was conducted in 1933-1937; yellowfin sole was the primary target species. After a hiatus during World War II, the Japanese resumed a yellowfin sole fishery in 1954 (see Figure 5.4). A Russian fleet began fishing the eastern Bering Sea in 1958, and by 1960 combined total landings of yellowfin sole peaked at 500,000 mt annually. This fishery subsequently declined sharply by 1963, likely due to stock depletion.
In the early 1970s, foreign fleets of predominantly Japanese, Russian, and Korean vessels participated in Bering Sea groundfish trawl fisheries. About 450 foreign fishing vessels operated in the eastern Bering Sea in 1971. Reported foreign landings of pollock, flatfish, rockfish, and cod peaked at 2.1 million mt in 1972, of which 85% were pollock. Subse-
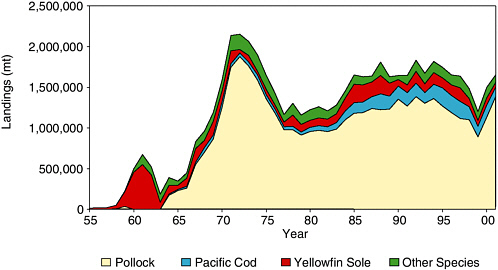
FIGURE 5.4 Groundfish landings from the Bering Sea, 1955-2001.
SOURCE: Graph prepared from data in Table 2, North Pacific Fishery Management Council (1999).
quently, total groundfish harvests declined, presumably due to reduced abundance.
The fishery recovered during the 1980s and landings have remained fairly constant since then. In the past decade (1990-2001), groundfish landings from the Bering Sea averaged 1.6 million mt, of which pollock accounted for 76% of the landings. Pacific cod accounted for 10%; yellowfin sole accounted for 7%; and the remainder included other flatfish, sablefish, and rockfish.
In the mid-1980s to early 1990s, a large pulse fishery for pollock developed in the Donut Hole in the central Bering Sea and a much smaller fishery developed near Bogoslof Island (north of the eastern Aleutian Islands) beyond the eastern Bering Sea shelf (see Figure 5.5). The Bogoslof fishery peaked at 377,000 mt in 1987, whereas the Donut Hole fishery peaked at 1.4 million mt in 1989. The latter was an intensive, essentially unregulated fishery with vessels from many foreign nations. Owing to severe depletion of pollock, most nations had stopped fishing by the time an international agreement was signed that closed the Donut Hole to fishing in 1993. Because of concerns about relationships between pollock near Bogoslof Island and those in the Aleutian Basin, the domestic Bogoslof Island fishery was closed in 1991.
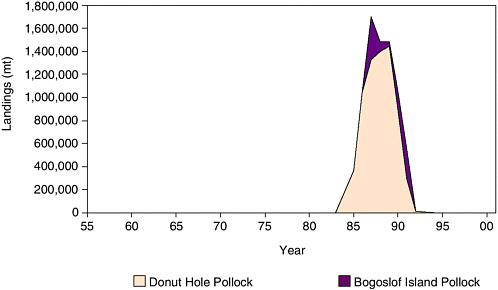
FIGURE 5.5 Pollock landings (metric tons) from the Bogoslof Island area and Donut Hole region in the central Bering Sea, 1955-2001.
SOURCE: Graph prepared from data in Table 1.12, Ianelli et al. (2001).
Numbers of Fishing Vessels
The size and number of fishing vessels in addition to landings are important because they are the platforms by which humans interact with Steller sea lions and their prey. Northern Economics, Inc., and EDAW, Inc. (2002) reviewed fishing vessel participation in the federally managed groundfish fisheries of the Gulf of Alaska, Aleutian Islands, and Bering Sea for 1992-2000. During this period, the number of catcher-processor vessels (i.e., vessels capable of both catching and processing fish) declined 31% from 130 in 1992 to 90 in 2000 (see Figure 5.6a). Likewise, the number of groundfish catcher vessels (i.e., vessels that catch but do not process fish) declined 24% from 1,658 in 1992 to 1,261 in 2000. Most vessels that exited the groundfish fisheries were fixed-gear (nontrawl) vessels less than 60 feet in length. Significant numbers of vessels made small groundfish landings but are not included in these statistics. For instance, during 1992-2000, of the 4,403 vessels that landed at least some groundfish, 2,376 landed at least 70 tons of groundfish per year. Total landings from the remaining 2,037 vessels accounted for just 0.1% of the total volume of groundfish landings. Most of these vessels targeted other species but delivered some groundfish as bycatch.
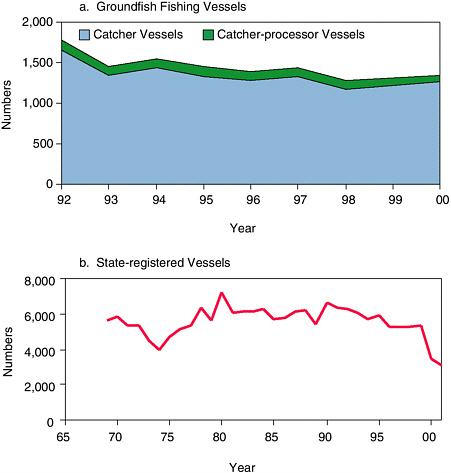
FIGURE 5.6 Number of vessels participating in (a) federal groundfish and (b) state fisheries in Alaska. (a) Catcher vessels and catcher-processor vessels that made significant landings (>70 tons annually per vessel) in the groundfish fisheries of the Gulf of Alaska, Aleutian Islands, and Bering Sea, 1992-2000 and (b) fishing vessels (all fisheries) registered with the state of Alaska in the region occupied by the western stock of Steller sea lions, which excludes southeastern Alaska and regions north of Bristol Bay, 1969-2001. Vessels must register with the state to fish for salmon, herring, crabs and other invertebrates, and groundfish (state waters only). Data for 2001 are preliminary. Numbers of state-registered vessels may be overestimated because vessel codes are unedited and may include errant identification numbers. Vessels participating in the halibut longline fishery are not included, and groundfish vessels fishing only in federal waters are not included unless they opt to register with the state.
SOURCES: Data for (a) from Northern Economics, Inc., and EDAW, Inc. (2002), Table 1.2-1, available at www.fakr.noaa.gov/npfmc/NorthernEconomics/NorthernEconomics.htm. Data for (b) provided by Susan Shirley, Alaska Department of Fish & Game, Juneau, personal communication, 2002.
Vessel size and type are other considerations in evaluating the effects of fishing vessels on the marine environment. Most catcher vessels can be divided into nine classes (Northern Economics, Inc., and EDAW, 2002). The participation of catcher vessels in groundfish fisheries off Alaska in 2000 was 590 (54.7%) nontrawl vessels of size 33 to 59 feet, 158 (14.7%) pot vessels greater than 59 feet, 72 (6.7%) longline vessels greater than 59 feet, 69 (6.4%) nontrawl vessels less than 33 feet, 46 (4.3%) trawl vessels less than 60 feet, 45 (4.2%) Bering Sea pollock trawlers of size 60 to 124 feet, 38 (3.5%) trawl vessels greater than 59 feet that are not eligible under the American Fisheries Act (AFA), 30 (2.8%) trawl vessels greater than 59 feet that are AFA eligible, and 30 (2.8%) trawl vessels greater than 124 feet.
Comparable time series of groundfish vessels have not been assembled prior to the 1990s owing to a lack of standardized data collection from foreign fishing fleets and lack of U.S. jurisdiction to 200 miles offshore prior to the Magnuson Fishery Conservation and Management Act of 1976. The state of Alaska has maintained data on fishing vessels since 1969. Vessels must register with the state to fish for salmon, herring, crabs, and other invertebrates in state and federal waters and groundfish (state waters only) or to enter state waters to deliver groundfish at an Alaskan port. Vessels participating solely in federally managed groundfish fisheries and delivering to offshore processors are not required to register with the state, and these vessels are underrepresented in the database. Participation was summarized for state-registered vessels for the region occupied by the western stock of Steller sea lions (see Figure 5.6b). Aside from a dip in the number of state-registered fishing vessels in the mid-1970s, participation has fluctuated around 6,000 vessels through the 1980s. As with the groundfish vessels, the number of state-registered fishing vessels declined 53% from 6,630 in 1991 to 3,108 in 2001. Counts of fishing vessels do not appear to reveal any striking patterns concurrent with Steller sea lion declines; however, the committee did not analyze trends at finer spatial scales or for particular fisheries.
Groundfish Stock Status and Harvest Rates
Harvest rates have varied over time with changes in policy, improvements in stock assessments, and increased understanding of fish stock productivity (DiCosimo, 1998). For groundfish fisheries under the purview of the NPFMC, acceptable biological catch (ABC) and overfishing levels are prescribed under a six-tier system depending on the level of information available about stock status. Many fish stocks in the Gulf of Alaska, Aleutian Islands, and Bering Sea fall into tier 3 in which the ABC is set based on F40%, the theoretical rate of fishing mortality that reduces spawning stock biomass per recruit to 40% of the pristine (estimated
unfished) level. The theoretical steady-state spawning stock biomass resulting from an F40% harvest strategy is termed B40%. For modeled fish populations with mortality, growth, and recruitment parameters typical of groundfish such as pollock and cod, an F35% strategy provides for high yields with low probability of reducing the fish stocks to dangerously low levels (Clark, 1991; Mace, 1994), and F40% was chosen to be more conservative by providing a buffer against uncertainty and management error. The actual fraction of fish biomass removed under an F40% harvest rate depends on life history characteristics that vary by species and stock as well as the variability in year-class strength and the fraction of immature fish not included in the spawning biomass calculations.
After considering estimates of ABC and other biological and economic information, the NPFMC sets a total allowable catch (TAC) to regulate commercial harvests for the year. For the Bering Sea the sum of all TACs must be no greater than 2 million mt; if this cap would be exceeded, then TACs for individual species (often pollock) are reduced so that the aggregate TAC stays within the prescribed limit of 2 million mt. Within the season, when the TAC for a particular species is reached (based on removals by the directed fishery and bycatch from other fisheries), that species becomes prohibited. Moreover, if continued fishing for other species creates a risk of overfishing (from bycatch), those other directed fisheries also could be suspended for the year.
Witherell et al. (2000) provide an example of the catch specification process for pollock in the eastern Bering Sea. Based on stock assessments conducted in 1999, projected biomass of age 3+ pollock (i.e., weight of all pollock age 3 and older; younger fish are not fully vulnerable to survey gear and cannot be accurately assessed) was 7.7 million mt. Although ABC was calculated to be 1.2 million mt using the F40% strategy, fishery scientists recommended a more conservative harvest strategy owing to uncertainties in recruitment and other information. Thus, the NPFMC lowered the ABC to 1,139,000 mt and set TAC equal to ABC. In the Aleutian Islands a similar process was undertaken, although less information about that stock prevents use of the F40% strategy. Instead, harvest rates are taken to be 75% of the natural mortality rate (M), resulting in an ABC of 23,800 mt for 2000. However, rather than setting TAC equal to ABC, TAC was set equal to zero, owing to low stock abundance and concern for Steller sea lions. For similar reasons, no fishing (TAC = 0) was allowed in the Bogoslof area.
The F40% harvest strategy has been a source of some confusion, regarding its effect on the fish biomass and prey availability to Steller sea lions. It is not a fishing rate that depletes the entire fish stock to 40% of the level that would occur in the absence of fishing. Rather, it is a rate that, in theory, would reduce spawning stock biomass per recruit to 40% of unfished levels on average. In the case of the latest pollock stock assess-
ment for the eastern Bering Sea (Ianelli et al., 2001), spawning biomass (i.e., the metric against which the F40% rate is applied) was projected to be 3 million mt at the time of spawning (spring) in 2002. On the other hand, age 3+ biomass was estimated to be 9.8 million mt at the start of 2002. The 6.8-million-mt difference between the two biomass estimates is immature fish; only a small fraction of age 2 pollock are mature, and 100% maturity is not reached until age 10 (Ianelli et al., 2001). The biomasses of age 1 and 2 pollock are not included in either estimate because of gear selectivity. As sea lions consume both juvenile and adult pollock (Merrick and Calkins, 1996), an F40% strategy does not deplete the entire pollock stock or even the portion consumed by sea lions to 40% of unfished levels. In fact, in the case of eastern Bering Sea pollock, the projected spawning biomass for 2002 is well above estimates of B40% at 2.6 million mt and Bmsy(biomass corresponding to maximum sustainable yield) at 2.1 million mt.
Historical exploitation rates applied to fish stocks can be approximated from estimates of exploitable fish biomass and catch. For most Alaskan groundfish stocks, historical changes in fish biomass are reconstructed using age-structured analytical models, such as the Stock Synthesis Analysis (Methot, 2000), whereby historical data from assessment surveys, catch sampling programs, and landings records are assimilated with life history statistics in a model estimation framework. Time series of biomass and historical catches were collected from the recent stock assessment conducted in 2001 (North Pacific Fishery Management Council, 1999, 2002). For expediency the proportionate exploitation rate (i.e., the fraction of the exploitable biomass removed by the fishery) was estimated by simply dividing the catch into the annual exploitable biomass estimate. Only pollock, cod, and Atka mackerel were considered here because of their apparent importance to the Steller sea lion diet.
In the Gulf of Alaska, estimated biomass of pollock of age 2 and greater increased from 431,000 mt in 1961 to 4.1 million mt in 1981 and declined in the 1990s to levels typical of the 1960s and early 1970s (see Figure 5.7a). Dynamics of this stock are largely attributed to a series of three strong year classes in the early 1970s followed by five consecutive strong year classes in the mid- to late 1970s (Dorn et al., 2001). Since then the frequency of strong year classes has diminished. Likewise, four strong year classes during 1976-1980 (Thompson et al., 2001) led to increases in cod biomass during the late 1970s and early 1980s, although it has declined since 1990 due to below-average year classes (see Figure 5.7b). New survey data suggest that juvenile abundance is near record-high levels owing to one or two strong year classes, and the expectation is that the stock will increase over the next few years as these fish grow and mature (Dorn et al., 2001). Exploitation rates for both pollock and cod fluctuated between 5 and 15% during the 1980s and 1990s (see Figures 5.7c and d).
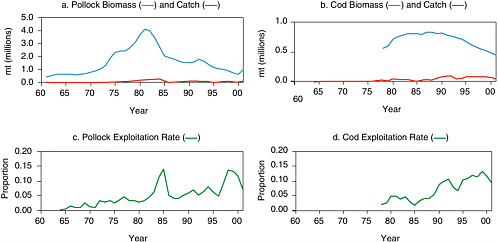
FIGURE 5.7 Gulf of Alaska groundfish exploitation, 1961-2001. (a) Estimated biomass of age 2+ walleye pollock and commercial catch, and (b) estimated biomass of age 3+ Pacific cod and commercial catch. Catch and biomass estimates are based on stock assessments conducted during 2001. Exploitation rate was approximated as estimated biomass divided by reported catch for (c) pollock and (d) cod.
SOURCES: Biomass data for pollock from Table 1.16, Dorn et al. (2001); data also available NMFS Alaska Fisheries Science Center website www.refm.noaa.gov/stocks/specs/Data%20Tables.htm. Catch data from Table 5, North Pacific Fishery Management Council(2002). Cod data from Table 2.24, Thompson et al., (2001); also available at the NMFS Alaska Fisheries Science Center website (see above).
In the Aleutian Islands, biomass estimates of Atka mackerel are highly variable because this species is patchily distributed on rough, rocky bottoms that are difficult to sample with trawl survey gear (Lowe et al., 2001). The species occurs primarily in the Aleutian Islands, but the 1994 and 1997 stock assessments located a significant biomass at a single survey station in the southern Bering Sea. Two strong year classes in 1977 and 1988 were largely responsible for an increase in biomass in the late 1970s and again in the early 1990s (see Figure 5.8a). During 1977-2001, the exploitation rate averaged 6.5%, with a peak of 14.2% in 1996 (see Figure 5.8b).
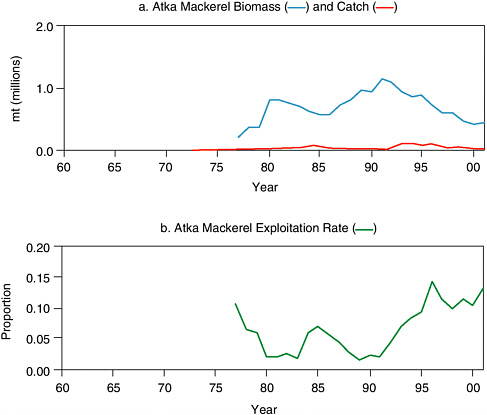
FIGURE 5.8 Aleutian Islands and Bering Sea Atka mackerel exploitation, 1977-2001. The top graph (a) shows estimated age 3+ biomass and commercial catch. Catch and biomass estimates are based on stock assessments conducted during 2001. Exploitation rate was approximated as estimated biomass divided by reported catch.
SOURCES: Biomass data from Table 13.9, Lowe et al. (2001); data also available at the NMFS Alaska Fisheries Science Center website www.refm.noaa.gov/stocks/specs/Data%20Tables.htm. Catch data from Table 3, North Pacific Fishery Management Council (1999).
The eastern Bering Sea supports by far the largest pollock fishery in Alaska, so trends in biomass and exploitation rate are of particular interest with respect to sea lions. Estimated biomass of pollock averaged 3.9 million mt during 1964-1980, increased more than 2.5-fold during the early 1980s, and averaged 10.4 million mt during 1983-2001 (see Figure 5.9a). Strong year classes in 1978, 1982, 1984, 1989, 1992, and 1996 are associated with subsequent periods of high abundance (Ianelli et al., 2001). Exploitation rates averaged 12.3% during 1980-2002 (see Figure 5.9c). Estimates of exploitation rate are questionable for the early years of this fishery because of uncertainty in the biomass estimates. For example, the exploitation rate in 1974 was approximately 50% based on the stock assessment conducted in 2001 and 94% based on the stock assessment conducted in 2000 (Ianelli et al., 2001). For the most recent 3 years, biomass estimates are also uncertain because young fish are only partially recruited to the fishery and are difficult to survey.
Estimated biomass of Pacific cod in the eastern Bering Sea and Aleutian Islands (note: most catch is taken in the Bering Sea) increased by an order of magnitude during the late 1970s to early 1980s and has been declining steadily since 1987, with a fairly steady increase in exploitation rate until 1997 (see Figures 5.9b and d). The biomass increase was associated with four strong year classes during 1977-1982, and the subsequent decline is associated with nine weak year classes during 1986-1998.
Spatial Distribution of Harvests
Fishing effort is unevenly distributed off the coast of Alaska (Fritz, 1993; Fritz et al., 1998; National Research Council, 2002). The concentration of fishing effort in some areas has focused concern on the potential for fishing to cause localized depletion of prey species for Steller sea lions.
With the shift from foreign to domestic groundfish fisheries in the 1980s, the fraction of pollock, cod, and Atka mackerel landings in Steller sea lion critical habitat increased (see Figure 5.10). Critical habitat is defined by the 20-nm radii around sea lion rookeries and important haulouts, and three aquatic foraging areas in Shelikof Strait, Bogoslof Island area, and Sequam Pass. This shift in effort was confounded by increases in fish biomass in the 1980s, and it is difficult to evaluate potential effects on local prey densities from these data alone.
Fritz (1993) compiled maps of trawl locations from observer data collected aboard foreign, joint venture, and domestic fisheries for pollock in 1977-1992. It is difficult to accurately summarize the temporal and spatial variability in trawl locations in the eastern Bering Sea because of large variability by year and season. During 1977-1980, much of the observed hauls occurred along the continental shelf break, with some scattered
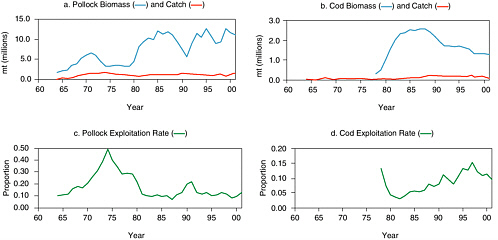
FIGURE 5.9 Bering Sea groundfish exploitation, 1964-2001. (a) Estimated biomass of age 3+ walleye pollock commercial catch, and (b) estimated biomass of age 3+ Pacific cod and commercial catch. Catch and biomass estimates are based on stock assessments conducted during 2001. Exploitation rate was approximated as estimated biomass divided by reported catch for (c) pollock and (d) Pacific cod.
SOURCES: Pollock biomass data from Ianelli et al. (2001). For cod, biomass data from Table 2.24, Thompson and Dorn (2001); data also available at the NMFS Alaska Fisheries Science Center website www.refm.noaa.gov/stocks/specs/Data%20Tables.htm. Pollock and cod catch data from Table 2, North Pacific Fishery Management Council (1999).
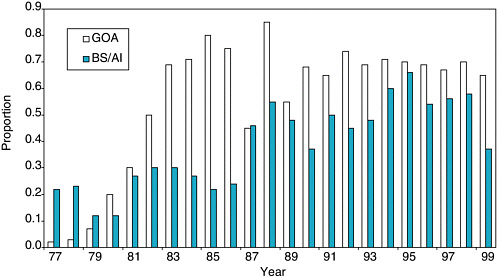
FIGURE 5.10 Proportion of pollock, Pacific cod, and Atka mackerel catches in Steller sea lion critical habitat for the Gulf of Alaska (GOA) and Bering Sea and Aleutian Islands (BS/AI) areas, during 1977-1999.
SOURCE: Data from National Marine Fisheries Service (2000).
effort on the continental shelf. During the early 1980s, effort expanded off the shelf in the Aleutian Basin from January to April, and effort expanded onto the continental shelf from May to December. From the mid-1980s to early 1990s, the geographic distribution of trawl locations was quite variable. For example, from January to April 1985 effort was concentrated mostly on the southeastern Bering Sea shelf in a relatively small area north of Unimak Island. From January to April 1986 nearly all effort occurred in the Aleutian Basin but shifted from May to August 1986 to the shelf north of 56° N with most effort on the northwestern portion of the shelf north of 58° N. After the closure of the Bogoslof Island fishery in 1992, most of the roe season (January to March) has been concentrated north and west of Unimak Island (Ianelli et al., 2001). Oceanographic factors, such as temperature, appear to influence the geographic distribution of both juvenile and adult pollock on the eastern Bering Sea shelf (Swartzman et al., 1994; Ianelli et al., 2001). In summary, survey and fishery data suggest a very high level of movement by pollock in the eastern Bering Sea.
In the Gulf of Alaska, observers documented several major shifts in the distribution of the pollock fishery during the late 1970s to early 1990s (Fritz, 1993). During 1977-1981, most pollock trawling occurred during
May to December along the continental shelf break. The distribution of observed trawl locations from September to December 1978 (see Figure 5.11a) typifies this pattern. In January to April 1982 an intense roe fishery developed in Shelikof Strait. In the same year, trawl effort continued and intensified along the shelf break during May through December. This new pattern continued until 1984, when the Shelikof Strait roe fishery became most intense (see Figure 5.11b), and fishing effort in May to December 1984 encompassed both the shelf break and portions of the outer continental shelf, particularly off Kodiak Island and the Alaska Peninsula (see Figure 5.11c). Both the winter roe fishery in Shelikof Strait and the non-roe fishery on the outer shelf and shelf break diminished substantially in intensity by 1986. This drop resulted from a combination of declining abundance and reduction in harvest rate (see Figure 5.7a, c). During 1990-1992, Shelikof Strait no longer dominated trawl locations during January to April to the same degree as during 1982-1986 (see Figure 5.11d), and during the rest of the year fishing effort was patchily distributed over the continental shelf in the central and western Gulf of Alaska with some concentrations in gullies off Kodiak Island (see Figure 5.11e).
The pollock roe fishery was intense for 5 years (1981-1985) in Shelikof Strait, and this area appears to be an important foraging area for Steller sea lions. It is uncertain whether these major shifts in fish removals adversely affected the number and density of pollock schools and the foraging success of Steller sea lions in the region. However, as described more fully under “Human-Related Steller Sea Lion Kills” in Chapter 6, thousands of sea lions were captured by this fishery (Perez and Loughlin, 1991) and many more were shot by fishermen to protect their catches and gear as nets were towed near the surface for delivery to motherships for processing. Summer bottom-trawl survey data from 1984 to 1996 show that there was no reduction in stations with high concentrations of pollock in the area (Shima et al., 2002). Interestingly, as the stock declined through the 1990s, the geographic distribution expanded, albeit at low densities, into deeper (200 to 300 m) waters than observed in the 1980s. This appears counter to expectations that pollock distributions would contract with reduced abundance as a density-dependent response consistent with MacCall’s (1990) basin model. The spatial expansion of pollock into deeper waters when abundance declined may simply reflect an expansion of suitable habitat as ocean temperatures increased following the 1976-1977 regime shift.
In recent years a few studies have been conducted to examine the potential for localized depletion of sea lion prey by fishery removals. An ongoing study is attempting to determine whether commercial fishing adversely affects the distribution and abundance of pollock and other
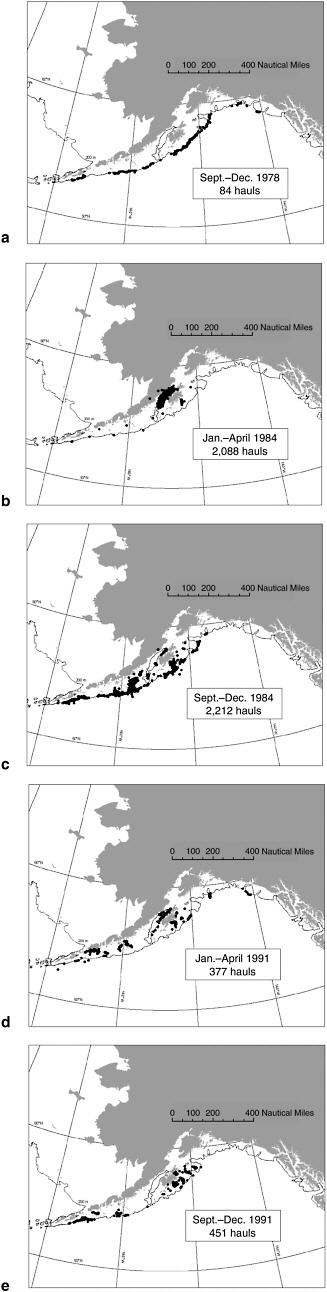
FIGURE 5.11 Locations of observed pollock trawl hauls in the foreign and joint-venture fishery in the Gulf of Alaska in (a) September-December 1978, (b) January-April 1984, (c) September-December 1984, and in the domestic fishery during (d) January-April 1991, and (e) September-December 1991.
SOURCE: From Fritz (1993); (a) p. 97, (b) p. 113, (c) p. 115, (d) p. 128, (e) p. 130.
forage fish over short spatiotemporal scales (Wilson et al., 2002). Two areas were studied off Kodiak, one open to trawling (Barnabas trough) and one closed (Chiniak trough). Hydroacoustic surveys were conducted before and during the fishery in 2001. No significant differences in the biomass of juvenile or adult pollock were detected in response to commercial fishing in which 27 vessels harvested 2,850 mt of pollock from Barnabas trough. Moreover, analysis of pollock school dynamics (e.g.,
shape, vertical distribution) indicated no significant differences between prefishing and fishing periods. In summary, there appears to be no significant link between this trawl fishery and the availability of pollock to Steller sea lions in a relatively small area on short timescales. However, these results are preliminary and reflect only one investigation into the localized depletion issue.
Other Steller sea lion prey species may be more susceptible to localized depletion. Evidence of seasonal, localized depletion of Atka mackerel by the trawl fishery in 1992-1995 has been reported by Fritz (1999). This fishery is concentrated in relatively few fishing areas (Fritz, 1993). In eight localized fisheries in the Aleutian Islands and Gulf of Alaska, Leslie regression analyses showed a sharp reduction in fishery catch per unit effort in some years and areas (Fritz, 1999). In areas with statistically significant declines, estimates of localized harvest rates were 55 to 91%. Fritz (2001) found significant depletion again at some sites in 1996 and 1997. In the Aleutian Islands, catch per unit effort returned to initial levels by the start of the fishery the following year, suggesting that the depletion was temporary. A more persistent depletion was observed from 1993 to 1994 at the Gulf of Alaska sites, but length-frequency distributions suggested that immigration of small fish contributed to the population. Preliminary results from a subsequent study indicate little movement of tagged adults in summer within 40 days of release, whereas a small proportion moved between areas within 64 days of release (Fritz et al., 2001). Because of concerns about localized depletion of Atka mackerel in areas important to foraging Steller sea lions, the NPFMC adopted regulations in June 1998 to disperse this fishery both temporally and spatially (Lowe et al., 2001). Now, the TAC is equally split into two fishing seasons—January 1 to April 15 (A season) and September 1 to November 1 (B season). A 4-year schedule was implemented to reduce the percentage of harvest in sea lion critical habitat in the central and western Aleutian Islands from 80% (1999) to 40% (2002). No similar reduction plan was adopted for the eastern subarea, but 20-nm trawl exclusion zones around Seguam and Agligadak rookeries, previously in effect for pollock trawling only, are now in effect year-round for the Atka mackerel fishery. Once this implementation schedule is completed, it would be prudent to conduct a follow-up to Fritz’s (1999; Fritz et al., 2001) study to evaluate the efficacy of these management actions that were intended to eliminate apparent localized depletion. A similar study of localized depletion on walleye pollock in the Bering Sea is currently in progress, and results are not yet available (Brian Battaile, University of Alaska, Fairbanks, personal communication, June 27, 2002).
Summary of Potential Effects of the Groundfish Fishery on Steller Sea Lions
Evidence linking the decline of the western stock of Steller sea lions to the development of commercial groundfish fisheries is equivocal. There is much uncertainty about the effects of fishing on fish prey on spatial and temporal scales associated with foraging sea lions due to a lack of directed research. With few exceptions, little is known about the effects of fishing on fish school dynamics and sea lion foraging behavior. By necessity, the committee focused on available information, which primarily includes historical regional trends of Steller sea lions, fish stock biomass, and fishery removals, and a few studies of fish spatial distributions and potential localized depletion by fisheries. From this review the following general conclusions are drawn.
There appears to be some evidence for overfishing of halibut in some areas of the Gulf of Alaska in the early 1900s: Pacific ocean perch in the Gulf of Alaska and Aleutian Islands in the 1960s and 1970s, yellowfin sole in the eastern Bering Sea in the 1960s, and walleye pollock in the central Bering Sea in the late 1980s. However, in each case the extent of overfishing cannot be quantified owing to the lack of assessment surveys on these fish stocks during those years. Depletion of pollock by a multinational fishery in the Donut Hole could have directly reduced prey availability to some Steller sea lions.
Although satellite telemetry data and at-sea observations (National Marine Fisheries Service, 2000) indicate that most sea lions stay on the continental shelf, some animals have been observed in the central Bering Sea. The diet of Steller sea lions in the Aleutian Islands is dominated by Atka mackerel, but depletion of pollock could further reduce diet diversity, which appears to be inversely correlated to the rate of decline (Merrick et al., 1997). Aside from pollock, other instances of overfishing occurred prior to the rapid decline of the sea lions in the 1980s and involved fish species that are less important components of the sea lion diet (see Chapter 4). For these earlier overfishing events to have played a significant role in sea lion declines, the effects must have been indirect and lagged over decades. Such potential ecosystem effects are evaluated in Chapter 6.
The intense Shelikof Strait pollock fishery in the early to mid-1980s may be implicated in declines in sea lions in the central Gulf of Alaska during 1976-1985. Undoubtedly, trawl bycatch and shooting of sea lions contributed to the decline in this region in these years, as discussed under “Human-Related Steller Sea Lion Kills” in Chapter 6. However, aspects of the pollock stock biomass and fishery history do not fit well with the decline of Steller sea lions in the central Gulf of Alaska in the context of a
fishery competition hypothesis. For instance, pollock biomass increased during the mid-1970s to early 1980s (see Figure 5.7a), when the sea lion decline appears to have started (see Chapter 3). Counts of sea lions in this region declined most sharply during 1985-1990, after this fishery was substantially reduced, but there may have been a lagged effect of the fishery.
Pollock biomass declined from the early 1980s through the 1990s in concert with sea lion declines in this region. Yet pollock biomass in the late 1990s was similar to biomass levels in the 1960s to early 1970s when sea lion populations were larger. Biomass of another important gadid prey, Pacific cod, was high in the 1980s and began a decade of decline around 1990 (see Figure 5.7b). It is difficult to evaluate whether localized depletion occurred with these changes in fish biomass. Survey stations with high densities of pollock persisted during the pollock decline of the mid-1980s and 1990s, at least in summer when the assessments were conducted (Shima et al., 2002). Depletion may have occurred during winter fisheries but subsequent seasonal movements of fish prior to the summer survey may have masked this effect. However, preliminary results from an ongoing experimental study off Kodiak Island suggest that localized depletion may not occur within timescales of days to weeks during the fishing season (Wilson et al., 2002), at least under the conditions of that experiment.
The best-supported case for localized depletion has been made for Atka mackerel in the 1990s. Catches and exploitation rates increased in the 1990s. Also, Atka mackerel are vulnerable to localized depletion because of their schooling behavior and apparently limited movement. The sharpest decline in the central Aleutian Islands, where Atka mackerel are fished, occurred in 1985-1989, a period of low exploitation rates. Unfortunately, no information on localized depletion was developed during these years. In the 1990s the recovery of catch rates to prefishery levels by the start of the next fishing season suggests that within-season localized depletion is the concern, not overfishing.
In general, the fishery competition hypothesis is not supported by comparisons of trends in Steller sea lion populations and fish biomass in the Gulf of Alaska, Aleutian Islands, and eastern Bering Sea. The 1970s was a decade of high sea lion abundance despite relatively low groundfish abundance. The 1980s was a decade of high groundfish biomass not-withstanding sharp declines in Steller sea lions. Biomass of eastern Bering Sea pollock, the largest of all fisheries, declined to intermediate levels during 1987-1991 but rebounded by 1993 and has since remained at or near all-time record high levels of abundance. Likewise, Atka mackerel biomass increased in the 1980s and peaked in the early 1990s, a period of sharp declines in the sea lion population in the Aleutian Islands. Most of
these shifts in groundfish biomass are related to changes in recruitment success, which appear to be largely driven by environmental conditions, rather than effects of fishing, in the range of observed fish biomass (e.g., Hollowed and Wooster, 1992, 1995).
An evaluation of groundfish harvest policy indicates that contemporary groundfish fisheries in the Gulf of Alaska, Aleutian Islands, and Bering Sea are generally managed conservatively based on stock assessments and reference points for target and overfishing rates. None of the groundfish stocks in the Alaska region are currently considered to be overfished (National Marine Fisheries Service, 2002). Errors in assessment are of concern, potentially leading to higher-than-intended harvest rates in some years, but even with this caveat, historical exploitation rates appear to be lower than overfishing rates with few exceptions over the past several decades. Accordingly, as discussed later in this chapter, sea lion protective measures have focused less on the overall rate of harvest and more on temporal and spatial apportionments of catches so as to mitigate the potential for localized effects by the groundfish fishery.
STATE-MANAGED FISHERIES
Overview
In addition to the groundfish fisheries, the state of Alaska manages other fisheries, including salmon, herring, crab, shrimp, scallops, and other invertebrates. Federal fishery management plans provide frame-works for state management authority for scallop and salmon fisheries statewide and for crabs in the Bering Sea and Aleutian Islands region. For other species the state manages fisheries within territorial and EEZ waters without the auspices of federal fishery management plans. Brief summaries are provided below of the historical and current herring, crab, and salmon fisheries managed by the state of Alaska following the more detailed review provided by Kruse et al. (2000).
Herring Fisheries
Fisheries for herring are considered here because of their relative importance in the diet of Steller sea lions. Herring are one of the top 10 prey of sea lions, based on frequency of occurrence, although they generally occur in 10% or less of stomachs (Pitcher, 1981) or scats (National Marine Fisheries Service, 1999). Their contribution to the diet varies both seasonally and regionally. Thomas and Thorne (2001) observed aggregations of Steller sea lions feeding on herring schools at night in late winter in Prince William Sound. Fishery managers with the Alaska Department
of Fish & Game commonly utilize the co-occurrence of sea lions and whales as indications that herring are approaching nearshore waters for spawning in spring (Kruse et al., 2000). The junk food hypothesis posits that sea lion nutrition was compromised by a regime shift in the late 1970s, which resulted in decreased abundance of herring and other fatty forage species and increased abundance of pollock and other ground-fishes of lesser nutritional value (Alverson, 1992; Rosen and Trites, 2000).
Alaska’s commercial herring industry began in 1878, and catch records are available dating back to 1915 (see Figure 5.12). Starting in the late 1800s, herring were rendered for oil at reduction plants. The fishery developed in southeastern Alaska, and this region dominated the catches in 1930, when it accounted for roughly 75,000 mt of the total 80,000 mt reported for Alaska. As herring stocks in southeastern Alaska declined,
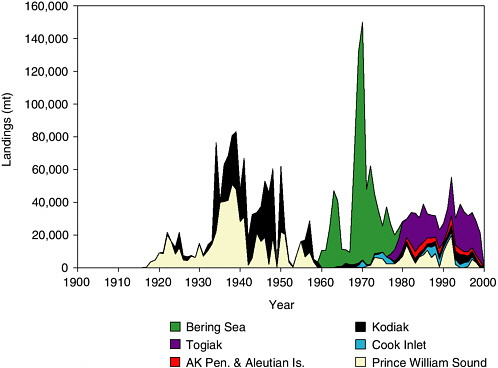
FIGURE 5.12 Historical herring catches from the Gulf of Alaska (excluding southeastern Alaska), Aleutian Islands, and Bering Sea. Catches include those from reduction fisheries prior to 1960, offshore foreign trawl fisheries in the Bering Sea during 1960-1980, and recent sac roe fisheries since 1970. AK Pen & Aleutian Is. includes catch data from Dutch Harbor (Aleutian Islands), and North and South Peninsula (Alaska Peninsula).
SOURCE: Data from Fritz Funk, Alaska Department of Fish & Game, Juneau, personal communication (2002).
the fishery shifted to Prince William Sound and Kodiak areas. Overall, the reduction fishery peaked at 142,000 mt in 1934. Catches generally declined but were highly variable through 1960, when they were discontinued. A large foreign fishery took place in the Bering Sea from 1960 to 1980. The Russians developed a trawl fishery in the late 1960s, and the Japanese developed trawl and gillnet fisheries in the early to mid-1960s. Catches peaked at over 140,000 mt in 1970. The contemporary fishery is conducted on prespawning fish to harvest their valuable roe for Asian markets. Harvests from the roe fisheries were relatively stable from 1980 to the mid-1990s (see Figure 5.12). In the late 1990s, catches declined sharply, with reductions in herring biomass, particularly in Prince William Sound, Cook Inlet, and Kodiak areas. Herring biomass and catches have remained relatively stable in southeastern Alaska and the eastern Bering Sea.
Herring are harvested in 25 distinct fisheries, almost all of which are tied to specific herring spawning locations. Contemporary fisheries are conducted by gillnet and purse seines. Fisheries are managed with catch quotas typically based on a harvest rate of 20% of estimated spawning biomass. Fishery thresholds, stock levels below which no fishing occurs, are commonly established at 25% of estimated unfished biomass. Fisheries have been closed in Prince William Sound since 1998 and lower Cook Inlet since 1999, because herring abundance has been at or below threshold.
Whether fisheries have depleted herring populations to the detriment of Steller sea lions remains an open question (see Chapter 6). The recent declines in herring abundance in Prince William Sound and Cook Inlet in the late 1990s occurred well after the declines in sea lions, but these declines are worrisome owing to the apparent importance of herring and squids in the diet of sea lions in Prince William Sound (Pitcher, 1981). Conversely, the large reduction fishery in the Gulf of Alaska peaked in 1934 and ended in 1960—well before the steep sea lion declines in the 1980s. Bering Sea herring stocks declined sharply in the late 1960s to mid-1970s associated with a large offshore trawl fishery and senescence of strong 1957-1958 year classes (Wespestad, 1991; Wespestad and Gunderson, 1991). Associated with recruitment of strong 1977-1978 year classes and termination of the trawl fishery, Bering Sea herring biomass recovered to average levels in the early to mid-1980s (Wespestad, 1991; Funk and Rowell, 1995), a period of decline of Steller sea lions in the Aleutian Islands. Since the late 1980s, herring biomass slowly declined (Funk and Rowell, 1995).
Crab Fisheries
Crabs are not preyed on by Steller sea lions. However, as described in Chapter 6, sea lions are known to puncture inflated crab pot floats, prior
to the introduction of solid-core Styrofoam “sea lion” buoys to prevent lost gear. In the 1960s and 1970s, anecdotal reports indicated that it was not uncommon for crab fishermen to shoot sea lions both in defense of their gear and for use as crab bait (Ed Opheim, former cod and salmon fisherman, Kodiak, personal communication, September, 2001). Because many of these fisheries are now closed due to reduced crab abundance, the historical development of Alaskan crab fisheries is of interest regarding a potential pulse of sea lion mortality associated with shootings by crab fishermen. Here, we only consider crab fisheries conducted in the range of the western stock of Steller sea lions. In southeastern Alaska, a portion of the region occupied by the eastern stock of Steller sea lions, there are small, but relatively stable, fisheries for red king, golden king, Tanner, and Dungeness crabs. Also, a small red king crab fishery occurs in Norton Sound.
The Japanese pioneered Alaskan crab fisheries with the tanglenet fishery in the 1930s. After a hiatus in the 1940s, the fishery resumed in the eastern Bering Sea in the early 1950s and the Russians entered the fishery in the late 1950s. The Japanese also caught crabs with pot gear in the late 1960s and early 1970s. These foreign crab fisheries continued until the early 1970s.
Domestic red king crab (Paralithodes camtschaticus) fisheries developed in Alaska in the mid-1950s to early 1960s (see Figure 5.13a). The Kodiak fishery peaked at 45,000 mt in 1965. Smaller fisheries were prosecuted in lower Cook Inlet and the Aleutian Islands. As red king crab fisheries declined through the 1970s in the Gulf of Alaska, a large fishery developed in Bristol Bay in the eastern Bering Sea. The Bristol Bay fishery peaked at 60,000 mt in 1980. This fishery plummeted in the early 1980s and was closed for a few years owing to depressed stock conditions. All red king crab fisheries have been closed since 1983, except for Bristol Bay and two small fisheries in southeastern Alaska and Norton Sound. Small (annual landings <5,000 mt) blue king crab (Paralithodes platypus) fisheries took place around the Pribilof Islands and St. Matthew Island in the eastern Bering Sea during the mid-1960s to late 1990s.
Japanese fishermen developed a blue king crab fishery off the Pribilof Islands in 1965, and U.S. domestic fishermen entered the fishery in 1973 (see Figure 5.13b). Domestic fishermen developed a similar fishery off St. Matthew Island in 1977. Landings peaked at 5,000 mt by 110 vessels from the Pribilof area in 1980-1981 and 4,300 mt by 164 vessels from the St. Matthew area in 1983. Both fisheries have been closed since 1999 owing to low abundance. Following the decline of red king crab fisheries, new fisheries developed for other species, such as the golden king crab (Lithodes aequispinus; see Figure 5.13c). The largest golden king crab fishery developed in the Aleutian Islands in the 1980s. Harvest from the Adak Island
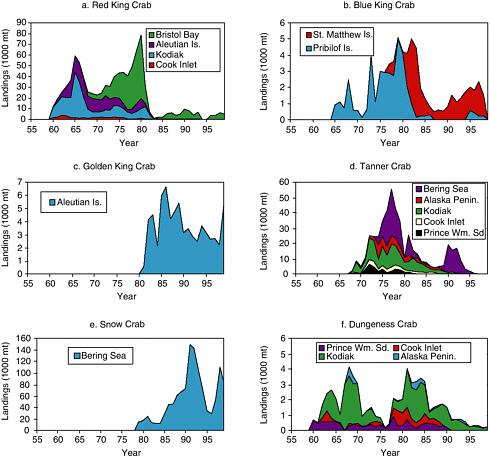
FIGURE 5.13 Historical landings from commercial fisheries for (a) red king crab, (b) blue king crab, (c) golden king crab, (d) Tanner crab, (e) snow crab, and (f) Dungeness crab from major crab fisheries conducted in the region occupied by the western stock of Steller sea lions. For Tanner crab, the Alaska Peninsula includes two fishing districts: Chignik and South Peninsula. Not shown are commercial crab fisheries in southeastern Alaska and relatively small crab fisheries for golden king crab in Prince William Sound, hair crab in the Bering Sea, and red king crab in Norton Sound.
SOURCE: Data from Shareef Siddeek, Alaska Department of Fish & Game, Juneau, personal communication (2002).
area peaked at 5,800 mt by 62 vessels in the 1981-1982 season and 900 mt by 17 vessels in 1995-1996 from the Dutch Harbor region. A small golden king crab fishery developed in Prince William Sound, peaking at just 67 mt in 1982-1983.
Fisheries developed for Tanner crab (Chionoecetes bairdi) in the late 1960s and early 1970s (see Figure 5.13d). The largest fishery in the Gulf of
Alaska occurred off Kodiak Island, which peaked at 13,000 to 15,000 mt in the mid- to late 1970s. The largest fishery occurred in the eastern Bering Sea, where 30,000 mt were landed in 1977 and 15,000 to 20,000 mt were caught in the early 1990s. Most Tanner crab stocks collapsed in the Gulf of Alaska in the 1980s. The eastern Bering Sea stock declined in the early 1980s but rebounded with several strong year classes that were recruited to the fishery in the late 1980s and early 1990s. In the late 1990s this stock declined again, and the fishery has been closed since 1997.
A large fishery for snow crab (C. opilio) developed in the late 1980s (see Figure 5.13e). This stock has gone through two large swings in abundance, with peak landings of >150,000 mt in 1992 and 1993, a sharp drop in landing to ˜30,000 mt in 1996 and 1997, and a second peak of 110,000 mt in 1999. More recently, the stock has declined sharply once again.
Fisheries developed for Dungeness crabs (Cancer magister) in the 1960s (see Figure 5.13f). Important fisheries occur in southeastern Alaska and Yakutat (east of 144° W). The Kodiak fishery is the largest in the central and western Gulf of Alaska. Many Dungeness crab fisheries show a cycle in landings, similar to stocks along the U.S. Pacific northwest coast. Dungeness crab stocks in Prince William Sound and Cook Inlet have not recovered from declines in the 1980s.
Small fisheries exist for other crab species as well. Korean hair crab (Erimacrus isenbeckii) harvests from the Bering Sea peaked at 1,100 mt in 1980. Effort ranged from 2 to 99 vessels, with less than 10 vessels in most seasons. Since the 1990s, even smaller fisheries developed for other species, including grooved Tanner crab (Chionoecetes tanneri), triangle Tanner crab (C. angulatus), and scarlet king crab (Lithodes couesi).
In the early years of most Alaskan crab fisheries, fishing seasons lasted much of the year. However, with increasing vessel participation and reductions in catch quotas as stocks became depleted, fishing seasons became shortened. For instance, a red king crab fishery in Bristol Bay landed 3,700 mt in just a 4-day fishing season during October 16-10, 2000. With the exception of Dungeness and golden king crab fisheries, most Alaskan crab fisheries occur in winter.
It is difficult to assess the potential role of shooting by crab fishermen on the decline of the western stock of Steller sea lions owing to a lack of data on the numbers of shot animals. Inferences must be drawn from landings data. The largest red king crab fisheries in the Gulf of Alaska and Aleutian Islands occurred in the 1960s and 1970s, before the steep decline of Steller sea lions. Large red king crab fisheries, primarily in Bristol Bay, persisted through 1982, when the sea lion decline started in the Aleutian Islands. Most red king crab fisheries have been closed since 1983. On the other hand, Tanner crab fisheries persisted into the late 1980s, and fisheries for golden king crab (mostly Aleutian Islands) and snow crab (Bering
Sea) developed in the 1980s during the sharpest period of sea lion declines. Dungeness crab fisheries (particularly in Kodiak) underwent two periods of increased landings, once in the 1960s and 1970s when sea lion abundance was high and once in the early 1980s when sea lions declined.
Salmon Fisheries
Fisheries for salmon are considered here because salmon are among the top 10 most frequently observed prey species in the diet of Steller sea lions (Hoover, 1988; National Marine Fisheries Service, 2000) and because of historical conflicts between sea lions and salmon fisheries (see Chapter 6). The apparent importance of salmon in the diet of sea lions depends on area, season, and data collection methods. On average, from the 1950s to the 1980s salmon were found in 5.6 to 7.1% of the stomachs of the eastern stock of Steller sea lions and just 1.1 to 1.8% of those collected from the western stock (National Marine Fisheries Service, 2000, see Figure 4.5). Pitcher (1981) found salmon in an average of 3.9% of 250 sea lion stomachs sampled during 1975-1978, but their presence was virtually confined to spring and summer, with salmon in only 1% of stomachs sampled during October to March. On the other hand, salmon were present in 12% (winter) to 25% (summer) of scat samples collected in the 1990s (National Marine Fisheries Service, 2000, see Figure 4.6).
Five salmon species reside in Alaska: pink (Oncorhynchus gorbuscha), sockeye (O. nerka), chum (O. keta), coho (O. kisutch), and chinook (O. tshawytscha). Salmon spawn in fresh water. Embryos overwinter in bottom substrate, and fry emerge in spring. Young salmon enter saltwater after 0 to 3 years of freshwater residence, depending on species. After extensive ocean migrations for 1 to 4 years, depending on species, maturing salmon return to natal freshwater systems for spawning.
Salmon have been an important food source to Alaska natives for thousands of years. The first small salmon fishery began in Kodiak in 1785 to provide dried salmon to Russian fur traders, and the Russian American Company exported small amounts of salted salmon in the early 1800s. However, large-scale development of Alaskan salmon fisheries followed construction of the first salmon cannery in Alaska in 1878 (see Figure 5.14).
In the early 1950s, Japan developed a large high-seas salmon fishery in the North Pacific using drift gillnets. A mothership fishery peaked at 16 motherships and 506 catcher boats in 1956. Catches from this mothership fishery and a land-based fishery declined through the 1970s, and in 1988 the Japanese drift gillnet fishery ended in the U.S. EEZ and part of the Bering Sea.
Record-high commercial catches occurred in the 1980s and 1990s (see
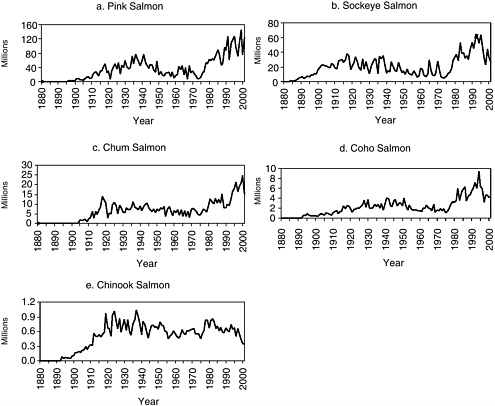
FIGURE 5.14 Historical commercial catches (millions of fish) of (a) pink, (b) sockeye, (c) chum, (d) coho, and (e) chinook salmon in Alaska. Data for 2001 are preliminary.
SOURCE: Data from Herman Savikko, Alaska Department of Fish & Game, Juneau, personal communication (2002).
Figure 5.14). However, there are exceptions, most notably chum and chinook salmon in western Alaska in recent years (Kruse, 1998). In the 1990s, on average, catches were dominated by pink salmon (60.0%), followed by sockeye (27.6%), chum (8.6%), coho (3.4%), and chinook salmon (0.4%). Most pink salmon are caught in southeastern Alaska and Prince William Sound, with significant landings off Kodiak Island. By far the largest sockeye salmon fishery occurs in Bristol Bay, but significant catches are taken from southeastern Alaska to the Alaska Peninsula. The largest fisheries for coho, chum, and chinook salmon generally occur in south-eastern Alaska, but smaller fisheries exist throughout the state. Most salmon are caught by gillnet, purse seines, and troll fisheries. Fisheries are generally managed using sequences of openings and closures in attempts to allow an optimal number of adult fish to return for spawning.
As with other fisheries, it is difficult to draw definitive conclusions about potential linkages between salmon fisheries and Steller sea lion declines. Salmon catches increased in the 1980s, a period of sharpest sea lion declines. However, these increased catches were associated with large increases in salmon runs and they continued until the late 1990s, so it is not possible to associate increased catches with reduced availability of salmon to sea lions as prey. Increases in Alaska salmon production are associated with the late 1970s climate regime shift (Beamish and Bouillon, 1993; Hare and Francis, 1994). An earlier period of reduced salmon abundance corresponds to an alternate climate regime in the late 1940s to late 1970s, during which sea lion populations were much healthier. Some lethal sea lion interactions with salmon fishing gear and fishermen have been documented (see Chapter 6), and apparently some salmon fisheries continue to occur in close proximity to sea lion haulouts (Kruse et al., 2001), at least providing the opportunity for disturbance. The issue of sea lion shootings by fishermen is considered in Chapter 6.
MANAGEMENT MEASURES TO MITIGATE POTENTIAL ADVERSE FISHING EFFECTS
A number of management measures have been implemented in attempts to mitigate potential adverse effects of fishing on Steller sea lions since they were first listed as threatened under the Endangered Species Act in 1990 (North Pacific Fishery Management Council, 1999, 2002; National Marine Fisheries Service, 2001). Additional measures were imposed after the western stock was listed as endangered in 1997. The following timeline provides a brief overview of many of these actions.
1990
-
Shooting firearms at sea lions or within 100 yards of sea lions is prohibited. Previously, it was legal to shoot at or near sea lions, and thousands of animals are believed to have died this way.
-
The number of sea lions that could be killed incidental to fishing is reduced from 1,350 to 675 animals.
-
Three-mile no-entry zones are established around major rookeries west of longitude 144° W.
-
The Bering Sea pollock fishery is divided into roe (or A season, January to March), and non-roe (or B season, usually September to October) fishing seasons to spread the harvest out over time.
-
An ending date for the B season is set at November 1 to prevent compression of pollock fisheries and to decrease the chance of localized depletion of prey for Steller sea lions.
-
Roestripping is banned statewide, partly due to concerns that discards attract sea lions to the fishing grounds.
1991
-
Trawl closure areas are implemented to reduce disturbance of feeding sea lions within 10 nm of 27 rookeries.
-
Some trawl closures are extended to 20 nm during the pollock A season.
-
The domestic Bogoslof Island fishery ends due to resource conservation concerns not solely related to sea lions.
1992
-
Trawl closure areas are implemented to reduce disturbance of feeding sea lions within 10 nm of 37 rookeries.
-
Trawl closures are expanded to 20 nm around five rookeries.
1993
-
The Atka mackerel harvest quota is apportioned among several small subareas in attempts to prevent localized depletion.
1998
-
An improved retention and utilization program takes effect, which bans the discard of pollock and Pacific cod in all fisheries off Alaska, regardless of gear type or fishery. Discards are only permitted at times required to avoid exceeding the TAC.
-
The Atka mackerel fishery is further dispersed both temporally and spatially. The TAC was equally split into two fishing seasons— January 1 to April 15 (A season) and September 1 to November 1 (B season). A 4-year schedule is implemented to reduce the percentage of harvest in sea lion critical habitat in the central and western Aleutian Islands from 80% (1999) to 40% (2002). No similar reduction plan is adopted for the eastern subarea, but 20-nm trawl exclusion zones around Seguam and Agligadak rookeries, previously in effect for pollock trawling only, are now in effect year-round for the Atka mackerel fishery.
-
In the Gulf of Alaska, full retention is required for Pacific cod.
-
Fishing is banned for forage fishes, such as capelin, sand lance, and others in efforts to prevent the development of new fisheries that
-
could compete with higher trophic levels, such as groundfish, seabirds, and marine mammals.
1999
-
Trip limits are imposed for the pollock fishery in the Gulf of Alaska.
-
Pollock fishing in the Aleutian Islands is prohibited.
-
Pollock catches in critical habitat areas are restricted.
-
Four fishing seasons are created for pollock in attempts to further spread the harvest out over time.
2000
-
More rookeries are added to the trawl closures.
-
Several new measures are instituted to spread out the catch of Pacific cod throughout the year.
Obviously, the potential need for additional fishery restrictions depends on the chances that fishing adversely affects Steller sea lions and the degree to which these effects may have been mitigated by measures already implemented. Because there have been frequent changes in sea lion protective measures since 1990, it is very difficult to evaluate the impacts of individual management measures on fish distribution, Steller sea lions, and the fishery.



































