
Cooperation and Competition in a Cliff-Dwelling People
![]()
In animals that breed cooperatively, adult individuals will sometimes delay reproduction to act as helpers at the nest who raise young that are not their genetic offspring. It has been proposed that humans are also a cooperatively breeding species because older daughters, grandmothers, and other kin and nonkin may provide significant childcare. Through a prospective cohort study of children’s (n = 1,700) growth and survival in the Dogon of Mali, I show that cooperative breeding theory is a poor fit to the family dynamics of this population. Rather than helping each other, siblings competed for resources, producing a tradeoff between the number of maternal siblings and growth and survival. It did not take a village to raise a child; children fared the same in nuclear as in extended families. Of critical importance was the degree of polygyny, which created conflicts associated with asymmetries in genetic relatedness. The risk of death was higher and the rate of growth was slower in polygynous than monogamous families. The hazard of death for Dogon children was twofold higher if the resident paternal grandmother was alive rather than dead. This finding may reflect the frailty of elderly grandmothers who become net consumers rather than net producers in this resource-poor society. Mothers were of overwhelming importance for child survival and could not be substituted by any category of kin or nonkin. The idea of cooperative breeding taken from animal studies is a poor fit to the complexity and diversity of kin interactions in humans.
___________________
Department of Anthropology and Research Center for Group Dynamics, Institute for Social Research, University of Michigan, Ann Arbor, MI 48109. E-mail: bis@umich.edu.
It has been reported that humans are a cooperatively breeding species that depends on individuals other than the mother and father for the successful rearing of offspring (Hrdy, 2005a, 2009). These individuals are known as alloparents and may be siblings, aunts, uncles, grandparents, and other kin or nonkin. One of the most thorough studies took place among the Maya of the Yucatan, Mexico. Hardworking girl-farmers paid back to their parents 94% of their cumulative consumption costs before leaving home to marry, and boys repaid 80%, enabling mothers to have more closely spaced births (Kramer, 2005a,b). In other species, alloparents seem to enhance their inclusive fitness by helping (Emlen, 1997a), but it is not yet clear whether this is also the case for Mayan children. In a study on the fishing atoll of Ifaluk in Micronesia, women (n = 7) whose two firstborn children were daughters had a mean of nine surviving offspring, and women (n = 11) who bore two sons first had a mean of five surviving offspring (Turke, 1988). This difference of four offspring was attributed to the role of elder daughters as helpers at the nest and was an enormous effect size for such a small sample (n = 18). Given that the analysis was bivariate, there is a strong possibility that the results reflect confounding variables.
In 19th century Finland, the survival of the maternal grandmother was correlated with improved grandoffspring survival (Lahdenperä et al., 2004). This study controlled for occupation (e.g., farmer, priest, or landless laborer) but did not distinguish among farmers by the size of their holdings; hence, the phenotypic correlation between grandmater-nal survival and grandoffspring survival may be caused by variation in wealth. A recent review (Sear and Mace, 2008) concluded that cooperative breeding was prevalent in traditional farming populations that had high fertility and high mortality, but only 6 of 17 studies controlled for wealth. A meta-analysis showed that the positive association between grandparental and grandchild survival was found only for the maternal and not the paternal side, although in these farming populations, the children were more likely to live with their paternal grandparents (Strassmann and Garrard, 2011).
Even in forager populations, the data on grandparental investment are mixed. In the Hadza of Tanzania, foraging grandmothers worked longer hours gathering tubers than they did in their prime, and children who received food from a grandmother or great aunt had higher nutritional status than children who were without alloparents (Hawkes et al., 1997, 1998). In the !Kung, having four surviving grandparents was not associated with improved grandchild survival or nutritional status (Draper and Howell, 2005).
Here, I use evolutionary social theory to explore the family dynamics of the Dogon of Mali, West Africa. My underlying premise is that
cooperative breeding is a facultative response to particular socioecological environments and that there is no strong evolutionary or empirical argument for viewing cooperative breeding as the species typical or evolved pattern in humans. In exploring Dogon social dynamics, I will focus on (i) siblings, (ii) extended families, and (iii) grandparents and grandchildren to test predictions from the hypothesis that humans are cooperative breeders against predictions from the following evolutionary hypotheses: kin selection (Hamilton, 1964a), life history theory (Stearns, 1992), parent–offspring conflict (Trivers, 1974), and local resource competition (Clark, 1978). My research design is a prospective cohort study of the growth and survival of 1,700 children (Methods).
ETHNOGRAPHIC BACKGROUND
The Dogon are traditional agriculturalists whose staple crop is pearl millet (Pennisetum glaucum). They have been the subject of a 25-year longitudinal study of human evolutionary biology that I initiated in 1986 and that is presently ongoing. The Dogon retain many of the features that have characterized humans over our evolutionary past, including the absence of contraception, polygyny, preservation of the indigenous religion, a subsistence economy, life in a tight-knit web of close and more distant kin, and high mortality levels (Strassmann, 1992, 2000). Marital residence is patrilocal (wives take up residence with the husband’s family), and descent is patrilineal (father to son). Women who are married to the same man are never sisters, and closely related women are not allowed to marry into the same patrilineage, a custom that disfavors female kinship bonds (Strassmann, 2003).
The Dogon are one of the most traditional people of Africa, and their cliffside villages have been designated a United Nations Educational, Scientific, and Cultural Organization (UNESCO) World Heritage site. On a seasonal basis, the area attracts tourists, but the revenues that are generated do not trickle down to the population. Infrastructure is sparse (electricity is absent and latrines are rare) in this area of Mali, which is a country that had a per capita income of $470 in 2006 (US Department of State, 2010). The United States Agency for International Development made condoms and low-cost hormonal contraceptives available in 2010; however, the population remains pronatalist, and the demand for these products in rural areas is nonexistent. The modal fertility per woman per lifetime is 10 live births (Strassmann and Gillespie, 2002).
RESULTS AND DISCUSSION
Siblings
The cooperative breeding hypothesis predicts that (i) older siblings help to rear younger siblings and (ii) the productive labor of children enables mothers to have more closely spaced births (Kramer, 2005a,b). It points to the beneficial impact that siblings have on each other during the juvenile period. By contrast, life history theory emphasizes the finite nature of parental resources and the tradeoff between offspring number and offspring quality. Kin selection theory (Hamilton, 1964a) predicts that siblings will help each other when rB > C in Hamilton’s rule. The theory of parent–offspring conflict (Trivers, 1974) and parental manipulation (Alexander, 1974) or social dominance theory (Emlen, 1997a) point to the divergent genetic interests of parents and offspring and the ability of parents, who are older and more powerful, to manipulate offspring, making them serve the genetic interests of parents.
How do the Dogon data stack up against these theoretical expectations? In a previous study of 176 children who were followed for 8 years, the addition of one extra child (age 0–10 years) to the extended family increased the odds of child death by 26% (Strassmann, 2000). In the present study (Methods), child growth (measured annually) decreased linearly as the number of maternal siblings increased (Fig. 14.1). Thus,
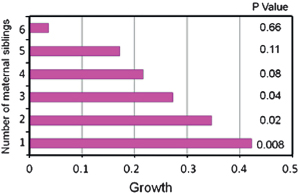
FIGURE 14.1 Child growth by number of maternal siblings. Growth is measured as the annual change in z score for height for age. The reference is seven maternal siblings. The other variables controlled include child’s age, year of study, sex, age by sex interaction, village of residence, survival of the paternal grandparents, sex by survival of the father’s mother interaction, sex by survival of the mother’s mother interaction, birth order (mother’s offspring), standardized wealth rank, and mother’s marital rank (n = 572 and the ages of the children were 41–98 months).
children were stunted in direct proportion to maternal family size. Sibling competition can also be seen in data on female lifetime reproductive success, which showed a tradeoff between the number of offspring born and the proportion that was successfully reared (Strassmann and Gillespie, 2002). Reproductive success reached a maximum of 4.1 offspring who survived to age 10 years at 10.5 maternal live births, and the 95% confidence limits around the maximum were 3.4–4.8 live births (Strassmann and Gillespie, 2002). Because of child mortality, the reproductive success of women who had 10 or 11 live births was not significantly greater than that of women who had 6 live births (Strassmann and Gillespie, 2002). In support of life history theory, the prevailing effect of siblings on each other was competition and not cooperation.
Behavioral scan data on childcare and work performed in the agricultural fields and the village show differences by sex and age (Fig. 14.2). Girls aged 5–9 years were observed doing childcare in the fields as often as adult women of reproductive age (Fig. 14.2A). Boys did relatively little childcare (Fig. 14.2A and B). Children of both sexes also performed non-childcare chores in the fields and the village (Fig. 14.2C and D). Children might prefer to play, but they are constrained by social norms, especially parental expectations and occasional scolding. Apparently, reprimands are also needed in the Maya (Kramer, 2005b). In the Efe, the services of fostered orphan boys help to offset the costs of their sustenance (Ivey, 2000; Ivey et al., 2005). Dogon children depend on parents for nutrition and survival in a harsh environment; their labor is obligatory, and I propose that it reflects parental manipulation (Alexander, 1974). Children’s work is also cooperative; however, it is not directly analogous to cooperative breeding in birds and other mammals because the latter refers to the postponement of personal reproduction caused by constraints on independent breeding opportunities (Emlen, 1997a). Human children who work and tend siblings are constrained not only by their economic insufficiency but also by their subordinance to parents as well as their reproductive immaturity.
In the context of parental coercion, Dogon children can become alloparents who habitually carry heavy infants on their backs. Sometimes, a Dogon allomother is an orphan, and the infant that she carries is the younger of two twins whose prospects for survival are poor. The obligation to take care of siblings, especially after the birth of twins, was cited by children as a reason for being unable to stay in school. If there is parent–offspring conflict over allomothering, then the reproductive success of adults who served as alloparents in childhood should be lower than that of individuals who were comparatively free of such responsibilities. Future research on cooperative breeding in humans should test this prediction to assess the role played by parental coercion.
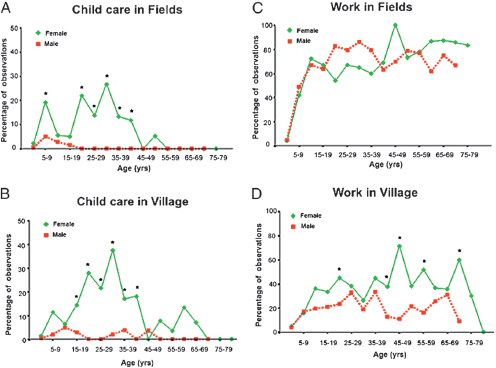
FIGURE 14.2 Childcare and work in the fields and village by sex and age. Solid lines females; dashed lines males. *P < 0.05 for the sex difference. (A) Child care in fields. At age 5–9 years, and 20–44 years females did significantly more child care than males. At other ages the difference was not statistically significant. (B) Child care in village. At age 15–44 years females did significantly more child care than males. (C) Work in fields. At no age was there a sex difference in the percentage of work observations over total observations for males versus females in the fields. Child care is excluded from the definition of “work.” (D) Work in village. From age 10 years onward, females generally did more nonchild care work in the village than did males with significant differences in five age groups (asterisks).
Work–Eat Groups
The work–eat group (WEG) is the functional family unit or economic group, and it is composed of the people who work together and eat from the same harvest during the millet growing season (Strassmann and Warner, 1998; Strassmann, 2003). The cooperative breeding hypothesis predicts that WEG members participate in alloparental care, especially sisters and grandmothers (Hrdy, 2009). I investigated the popular adage that it takes a village to raise a child (Hrdy, 2009), which implies diffuse and shared responsibility for children across closer and distant
kin or even nonkin. Specifically, I asked if children had better survival in extended families than nuclear families. If extended families are preferable, then children should survive better when there are more married adults in the WEG. Kin selection theory does not make a prediction about WEG size per se, but it does predict increased conflict in WEGs that have more asymmetries in genetic relatedness, such as in polygy-nous WEGs. Kin selection theory (Hamilton, 1964a) also predicts a linear relationship between the child’s relatedness to the WEG boss and his or her survival.
I tested these predictions using longitudinal data on the hazard of death in the first 5 years of life (Methods) and found that the number of married adults in the WEG was not significantly associated with child mortality (Table 14.1). I infer that, in the Dogon, extended families are not at any advantage or disadvantage compared with nuclear families. Interestingly, the dependency ratio (number of children less than 10 years of age divided by the number of married adults) was also not a significant predictor of the risk of death. The one person in the WEG
TABLE 14.1 Predictors of Death by Age 5 Years (n = 3,000 person-years and n = 165 deaths)
| Variable | p | Hazard Ratio | 95% Confidence Limits |
| Mothera | 0.0113 | 0.242 | 0.081-0.726 |
| Fathera | 0.6899 | 1.511 | 0.199-11.471 |
| Year 1b | 0.0002 | 2.316 | 1.495-3.589 |
| Year 2b | <0.0001 | 2.500 | 1.644-3.802 |
| Malec | 0.0595 | 0.740 | 0.541-1.012 |
| WEG boss | |||
| Father's fatherd | 0.9647 | 1.011 | 0.610-1.677 |
| Mother's fatherd | 0.1425 | 0.535 | 0.232-1.234 |
| Father's brotherd | 0.5266 | 1.234 | 0.644-2.364 |
| Father's father's brotherd | 0.1349 | 1.719 | 0.845-3.497 |
| Unrelatedd | 0.4339 | 0.553 | 0.126-2.436 |
| Miscellaneousd | 0.2434 | 0.646 | 0.309-1.347 |
| Grandparents | |||
| Father's mothera | 0.0007 | 1.857 | 1.305-2.693 |
| Father's fathera | 0.0485 | 1.556 | 1.003-2.415 |
| Mother's mothera | 0.7668 | 0.949 | 0.672-1.341 |
| Mother's fathera | 0.6570 | 1.075 | 0.781-1.479 |
| Number of married adults in WEG | 0.5651 | 0.988 | 0.948-1.030 |
Notes: This model includes the number of married adults in the WEG. A hazard ratio greater than 1.0 means an increased risk of death, and a hazard ratio less than 1.0 means a decreaseed risk of death. Because child's age was the time axis in the analysis, it was not used as a predictor in the model.
a Reference is dead. b Reference is year 3. c Reference is female. d Reference is father.
TABLE 14.2 Predictors of Death by Age 5 Years (n = 2,933 person-years and n = 161 deaths)
| Variable | p | Hazard Ratio | 95% Confidence Limits |
| Mothera | 0.0097 | 0.230 | 0.076-0.701 |
| Fathera | 0.7093 | 1.468 | 0.195-11.045 |
| Mother's age | 0.3458 | 0.987 | 0.962-1.014 |
| Year 1b | 0.0001 | 2.420 | 1.533-3.822 |
| Year 2a | <0.0001 | 2.718 | 1.756-4.206 |
| Malec | 0.0805 | 0.750 | 0.543-1.036 |
| Wealth | 0.0532 | 1.134 | 0.998-1.287 |
| WEG polygynyd | 0.0196 | — | — |
| Village 9e | 0.3659 | — | — |
| Polygyny x village 9f | 0.0626 | — | — |
| WEG boss | |||
| Father's fatherg | 0.8824 | 1.038 | 0.631-1.709 |
| Mother's fatherg | 0.0899 | 0.473 | 0.199-1.124 |
| Father's brotherg | 0.3400 | 1.378 | 0.713-2.662 |
| Father's father's brotherg | 0.0424 | 1.960 | 1.023-3.755 |
| Unrelatedg | 0.5591 | 0.633 | 0.136-2.937 |
| Miscellaneousg | 0.3540 | 0.689 | 0.313-1.516 |
| Grandparents | |||
| Father's mothera | 0.0009 | 1.926 | 1.309-2.832 |
| Father's fathera | 0.0649 | 1.544 | 0.974-2.450 |
| Mother's mothera | 0.6917 | 0.931 | 0.653-1.327 |
| Mother's fathera | 0.6366 | 1.083 | 0.778-1.508 |
Notes: A hazard ratio greater than 1.0 means an increased risk of death, and a hazard ratio less than 1.0 means a decreased risk of death. Because child's age was the time axis in the analysis, it was not used as a predictor in the model. There was no significant interaction between offspring sex and grandparental survival.
aReference is dead. bReference is year 3. cReference is female. dThis variable shows the effect of polygyny on the hazard of death in villages 1-8. eReference is villages 1-8. fthis variable shows the interaction of polygyny and village. gReference is father.
who emerged as overwhelmingly important for child survival was the child’s own mother (Table 14.2). Children whose mothers were alive faced a hazard of death that was 77% lower than that of children whose mothers were dead (P < 0.01). In the Dogon, it is mothers alone who are critical for getting children past the early-life bottleneck in survival. The survival of the father was not a significant predictor of the hazard of death for Dogon children in early childhood (P = 0.71) (Table 14.2).
The hazard of death was more than twofold greater in year 1 (P = 0.0001) and year 2 (P < 0.0001) of the study than in year 3 (the reference year). In year 3, a major drought decreased insectborne illnesses such as malaria. The hazard of death for boys was 25% lower than for girls, a difference that was not quite statistically significant (P = 0.08). WEG wealth was a rank variable from 1 (rich) to 6 (poor). A change in wealth of one rank (from richer to poorer)
increased the hazard of death by 13% (P = 0.05) (Table 14.2). I defined WEG polygyny as the ratio of married women to married men in the WEG. There was an interaction between WEG polygyny and the child’s village of residence (P = 0.06). In villages 1–8, the hazard of death was significantly higher in polygynous WEGs (P = 0.02). Village 9 was exceptionally large and wealthy, and only in this village did polygyny have no adverse impact (Table 14.2). In regard to child survival, it was the polygyny index for the WEG as a whole that mattered and not the marital status (sole wife or first, second, or third wife) of the child’s own mother.
To explore the mechanisms that underlie the decrease in child survival under polygyny in the more traditional villages (1–8), I compared the growth of Dogon children to the World Health Organization’s healthy international reference population (World Health Organization, 2005). During their first 18 months, the children (n = 474 observations) fell behind the international reference population at the rate of ~1 SD per year, which is indicative of severe stunting. In the age group 19–40 months, most of the children (n = 432 observations) were continuing to fall behind the reference population, but the rate of stunting had slowed down and was now less than 0.1 SD per year. From 41 to 98 months (n = 572 observations), the children were experiencing catch-up growth, and the mean change in z score for height for age between successive years was positive. Controlling for other significant predictors, the rate of stunting was significantly lower for the children of sole wives than the children of first wives and intermediate for the children of second, third, and fourth wives (Fig. 14.3). It is unclear why mother’s marital status mattered for children’s growth, whereas their survival was only impacted by the polygyny of the WEG as a whole. However, it is clear that one needs to examine fam-
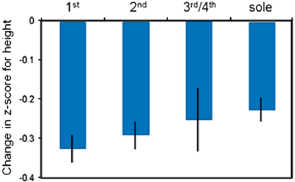
FIGURE 14.3 Child growth by mother’s marital rank. Children of first wives were falling behind the healthy reference population at a faster rate than children of sole wives.
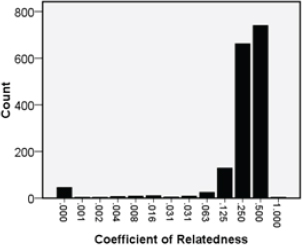
FIGURE 14.4 The number of children by their coefficient of relatedness to their work–eat group (WEG) boss.
ily structure, including such features as polygyny, to understand the social dynamics and patterns of parental and alloparental investment in families.
A major feature of WEG structure is the genetic relatedness among the various family members. The coefficient of relatedness of the children in the prospective cohort study to the boss of their WEGs was quite variable (Fig. 14.4). Most commonly, the WEG boss was the child’s father (r = 0.5) or grandparent (usually the paternal grandfather; r = 0.25), but sometimes, the WEG boss was the paternal great uncle (r = 0.125) or rarely, an unrelated man (r = 0) who was married to a female relative. Based on kin selection theory (Hamilton, 1964a), I predicted a linear relationship between the child’s relatedness to the WEG boss and his or her survival. Instead, there was no significant difference in child survival when the WEG boss was the child’s father, paternal grandfather, maternal grandfather, or father’s brother (Fig. 14.5 and Table 14.2). Evidently, children whose coefficient of relatedness to the WEG boss was 0.25 (uncles and grandfathers) fared as well as children whose coefficient of relatedness to the WEG boss was 0.50 (fathers). However, if the WEG boss was the child’s paternal great uncle (related to the child by 0.125), then the hazard of death by age 5 years was twofold higher than if the WEG boss was the child’s father (P = 0.04) (Table 14.2).
A possible explanation for these results is that the grandfather is symmetrically related to all of his grandchildren, and he may treat them equally, whereas a paternal great uncle has descendants who are much more closely related to him (0.5 vs. 0.125) and who are better targets for his investment according to Hamilton’s rule (Hamilton, 1964a). Grandfathers are only one-half as related to grandchildren as fathers are to children, but the disadvan-
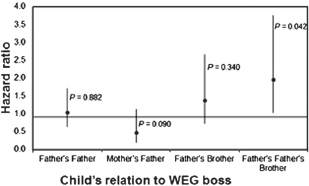
FIGURE 14.5 The hazard of death by age 5 years in relation to the child’s relatedness to the WEG boss (n = 2,933 person-years and n = 161 deaths). The child’s father as WEG boss is the reference category. A hazard ratio greater than 1.0 means an increased risk of death if the WEG boss is the indicated person instead of the child’s father. A hazard ratio less than 1.0 means a decreased risk of death if the WEG boss is the indicated person instead of the child’s father. Error bars are 95% confidence limits. Table 14.2 shows the other variables that were controlled.
tage of having a grandfather as WEG boss may be offset by the advantage of having a WEG boss who is older and who commands more authority in this gerontocratic society. Some fields are inherited father to son, but others are owned by the patrilineage under a system of gerontocratic control that gives the eldest WEG boss the choice parcels and the most junior WEG boss the worst parcels (Bouju, 1984). The finding that runs counter to kin selection theory is the lack of difference in survival between children whose father was the WEG boss vs. children whose WEG boss was the paternal uncle (father’s brother). This result echoes the finding that the death of the father did not jeopardize child survival.
WEGs are seasonal and do not hold together for the entire year. After the millet has been harvested, WEGs often break up for the onion gardening season. Each person waters their own individual onion garden, and a collaborative effort is needed only for pounding the bulbs and stems. The onion mash is rolled into balls that are spread out in the sun to dry. The desiccated onion balls are easier to store and transport than the fresh bulbs, and they are an export commodity that is shipped to Bamako and Abidjan. People invariably said that they preferred the onion work because there is no problem of free riders: every married adult (male or female) reaps the reward for their own efforts. Children work in the onion gardens as well, but their yield belongs to their fathers.
Over time, larger WEGs tend to fracture into multiple smaller WEGs. Kin selection theory (Hamilton, 1964a) predicts that WEG fission
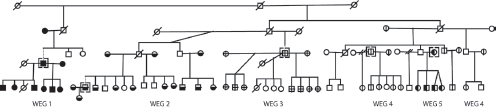
FIGURE 14.6 WEG dissolution over time. In 2000, there were only two WEGs as the members of WEGs 2–5 worked together. By 2010, there were five WEGs. This family segment illustrates the typical pattern wherein half brothers separate after the death of the patriarch, followed ultimately by the separation of full brothers. White icons indicate persons who lived outside the patrilineage in 2010. WEG bosses are indicated in brackets. Slashes through icons indicate that the person was deceased in 2010.
occurs along kinship lines. I tested this hypothesis using data on WEG stability from pedigree data for 29 Dogon patrilineages in nine villages. These patrilineal pedigrees are based on genealogical interviews of 1,285 adult males. Fig. 14.6 shows three generations of descendants of a single Dogon man in one of the patrilineages (the full pedigree has 11 generations and is too large to reproduce here). At the time of the census in the year 2000, the patriarch’s son by one wife had split off from his two sons by his other wife, producing a small WEG (Fig. 14.6, WEG1) and a large WEG containing everybody else. By the year 2010, the large WEG had further split into four separate WEGs (WEGs 2–5). The typical pattern is that when the patriarch (grandfather) is alive, he requires his sons and their wives and children to work together. After he dies, there is a brief grace period, and then, the half brothers separate from each other. More time elapses, and then, even the full brothers often split apart (Strassmann, 2003). This pattern is consistent with kin selection theory (Hamilton, 1964a), which predicts greater discord among half than full brothers. It also shows the coercive role of the grandfather in eliciting cooperation from his sons.
Because postmarital residence is patrilocal, women are not genetically related to anyone in the WEG except their own offspring. In interviews (n = 113 women and n = 77 men), women invariably disliked working with their husband’s extended family and preferred smaller WEGs. Women were said to instigate WEG fissioning by quarreling with their sisters-in-law. Tensions between cowives extend to their children and often lead to the rupture of the WEG in the next generation (Strassmann, 2003). Although men usually assigned women the blame, it is unlikely that WEG fissioning is entirely caused by the women. A
common pattern was for a younger brother who had monetary savings from working in the city to split off from his older brother so as to keep control of his wealth. Brothers are first-degree relatives (r = 0.5), but Hamilton’s rule takes into consideration not only relatedness but also costs and benefits. Although WEGs are cooperative units, they are unstable and experience internal conflicts of interest. Kin selection theory predicts important aspects of WEG dynamics such as the conflict among cowives and sisters-in-law and the circumstances that trigger WEG fissioning. The person in the WEG who is most important for child survival is the child’s own mother, a finding also seen in other demographic datasets (Sear and Mace, 2008) as well as in primate species that are not cooperative breeders.
Grandparents and Grandoffspring
The hypothesis that humans are cooperative breeders has focused, in particular, on grandparental investment (Hawkes et al., 1997, 1998; Beise, 2005; Gibson and Mace, 2005; Hawkes and Jones, 2005; Hrdy, 2005a, 2009; Leonetti et al., 2005; Hill and Hurtado, 2009; Coall and Hertwig, 2010; Kaptijn et al., 2010; Strassmann and Garrard, 2011). The Dogon data provide an opportunity to examine the survival status of children in relation to the survival status of the four kinds of grandparents (father’s mother, father’s father, mother’s mother, and mother’s father). In the presence of controls for other predictors of child mortality, the hazard of death for Dogon children was twofold higher if the father’s mother was alive rather than dead (P = 0.0009) and 54% higher if the father’s father was alive (P = 0.06) (Fig. 14.7 and Table 14.2). The hazard of death was not influenced by the survival status of the maternal grandparents.
Given that residence is with the husband’s and not the wife’s parents, I predicted that older paternal grandparents become a drain on family resources and consume more than they produce. In testing this prediction, I used the age of the father as a proxy for the age of his parents. Children whose father’s were older (and who presumably had older paternal grandmothers) were at the greatest survival disadvantage if the paternal grandmother was living (Table 14.3). The Dogon seem to take this pattern into account, because they kick the paternal grandmother out of the WEG after the death of her husband (Strassmann, 2000, 2003). In Dogon society, an old woman whose husband is deceased is perceived as a liability rather than as a valuable alloparent. Old women work physically harder than old men (Fig. 14.2D), but the old men have a valuable asset: They wield power.
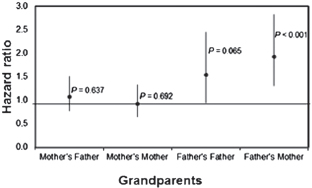
FIGURE 14.7 The hazard of death by age 5 years in relation to the survival status of the child’s grandparents (dead is the reference category; n = 2,933 person-years and n = 161 deaths). A hazard ratio greater than 1.0 means an increased risk of death if the grandparent is alive (rather than dead). Error bars are 95% confidence limits. Table 14.2 shows the other variables that were controlled. The hazard of death was twofold greater if the father’s mother was alive.
The old ladies must return to their natal patrilineages and work to support themselves with a small parcel of land that they cultivate alone or with the help of a daughter’s daughter (Strassmann, 2003). A woman’s two firstborn offspring are raised by their maternal grandparents, so boys sometimes also live matrilocally. In the case of boys, this arrangement is usually temporary, and sons eventually go to their fathers. I investigated the growth in height of these matrilocal children (n = 107) in relation to both their sex and the survival status of their four grandparents. The data are adjusted for other significant predictors of growth. In each year of the study, the family structure variables were used to predict the change in a child’s height-for-age z score from one year to the next (Methods). Girls tended to grow at a faster rate if their maternal grandmother was dead rather than alive (Fig. 14.8), presumably on account of the work that girls perform. Little girls primarily fetch water, but as they grow older, they take on such tasks as weeding gardens, pounding millet, and helping to gather firewood. Boys performed less work for their maternal grandmothers, and their growth rates were unaffected by their maternal grandmothers’ survival status (Fig. 14.8).
The relationship between growth and grandparental survival status in the children who lived patrilocally is shown in Fig. 14.9. Because there were more children who lived patrilocally (Fig. 14.10), it was possible to disaggregate the data into three age groups. In the youngest children (ages 0–18 months; n = 474), the survival status of the father’s father,
TABLE 14.3 Predictors of Death by Age 5 Years (n = 2,762 person-years and n = 155 deaths)
| Variable | P | Hazard Ratio | 95% Confidence Limits |
| Mothera | 0.0050 | 0.206 | 0.068-0.620 |
| Fathera | 0.6597 | 1.546 | 0.222-10.755 |
| Father's age | 0.0696 | — | — |
| Father's age x father's mother | 0.0116 | — | — |
| Year 1b | 0.0007 | 2.214 | 1.395-3.513 |
| Year 2b | <0.0001 | 2.484 | 1.603-3.851 |
| Malec | 0.1535 | 0.786 | 0.565-1.094 |
| Wealth | 0.0616 | 1.128 | 0.964-1.281 |
| WEG polygynyd | 0.0510 | — | — |
| Village 9e | 0.4676 | — | — |
| Polygyny x village 9f | 0.0934 | — | — |
| WEG boss | |||
| Father's fatherg | 0.6160 | 1.140 | 0.683-1.904 |
| Mother's fatherg | 0.5642 | 0.727 | 0.245-2.152 |
| Father's brotherg | 0.4564 | 1.294 | 0.657-2.549 |
| Father's father's brotherg | 0.0274 | 2.107 | 1.087-4.086 |
| Unrelatedg | 0.5884 | 0.554 | 0.065-4.699 |
| Miscellaneousg | 0.6710 | 0.832 | 0.357-1.941 |
| Grandparents | |||
| Father's mothera | 0.1026 | — | — |
| Father's fathera | 0.1316 | 1.443 | 0.869-2.324 |
| Mother's mothera | 0.9408 | 0.987 | 0.691-1.409 |
| Mother's fathera | 0.5745 | 1.100 | 0.788-1.537 |
Notes: The significant interaction of father's age and survival status of father's mother shows that older paternal grandmothers are more harmful than younger paternal grandmothers. Mother's age was omitted because it is collinear with father's age.
aReference is dead. bReference is vear 3. cRererence is temale. dIhis vanable shows the effect of polygyny in villages 1-8 in terms of the ratio of married women to married men in the WEG. eReference is villages 1-8.fthis variable shows the interaction of polygyny and village. gReference is father.
mother’s mother, and mother’s father was unrelated to grandchild growth rates (Fig. 14.9A). However, the survival status of the father’s mother affected girls and boys differently. Girls grew better than boys if the father’s mother was alive (P = 0.05). Boys grew better if the father’s mother was dead rather than alive (P = 0.04) (Fig. 14.9A).
In the next age group (ages 19–40 months; n = 432), the major finding was that boys were catching up in height to the international reference population if the father’s mother was dead but continuing to fall further behind if she was alive (P = 0.001) (Fig. 14.9B). Additionally, when the father’s mother was dead, boys grew at a rate that was significantly better than that of girls (P = 0.0003). In this age group, girls
were continuing to fall behind the reference population, regardless of the survival status of the paternal grandmother (Fig. 14.9B).
In the oldest age group (ages 41–98 months; n = 559), the children were experiencing catch-up growth across the board (Fig. 14.9C). The catch-up growth of girls was greater if the father’s mother was alive rather than dead (P = 0.003). Moreover, if the father’s mother was alive, the catch-up growth of girls was greater than that of boys (P = 0.0002) (Fig. 14.9C). Interestingly, girls grew at a faster rate if their mother’s mother was dead rather than alive (P = 0.003), echoing the results for the girls who lived matrilocally. When the mother’s mother was dead, girls also grew faster than boys (P = 0.0004) (Fig. 14.9C).
Interactions between sex, grandparental survival, and child survival or growth have also been found in other human populations (Jamison et al., 2002; Gibson and Mace, 2005; Sear, 2008). Rather than being a spurious result, it seems that Dogon paternal grandmothers may prefer girls over boys. A possible evolutionary explanation is local resource competition (Clark, 1978). Girls marry out of the patrilineage, whereas boys remain home to compete against each other for resources, and therefore, it may make sense to produce fewer boys. In no age group did the survival status (alive or dead) of grandfathers make a difference for growth. It could be that grandfathers are not sufficiently involved in the care and feeding of young children for their presence to make a difference.
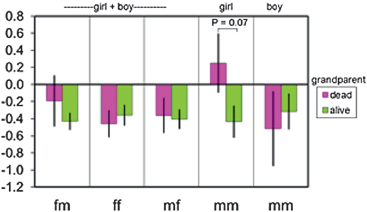
FIGURE 14.8 Growth (change in z score for height for age) in children who lived matrilocally (n = 107; ages 0–98 months). Other significant predictors of growth were controlled (child’s age centered on the mean, year of study, sex, mother’s age at child’s birth). fm, father’s mother; ff; father’s father; mf, mother’s father; mm, mother’s mother.
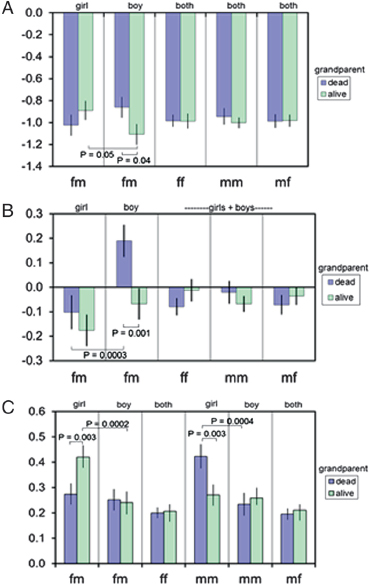
FIGURE 14.9 Growth (change in z score for height for age) in children who lived patrilocally in relation to sex and the survival status of the child’s grandparents, fm, father’s mother; ff, father’s father; mm, mother’s mother; mf, mother’s father. (A) Age group 0–18 months (n = 473). (B) Age group 19–40 months (n = 432). (C) Age group 41–98 months (n = 559). The analysis controlled for child’s age, child’s age squared, year, sex, age by sex interaction, nursing vs. weaned, village of residence, birth order (one to seven), number of maternal siblings (one to seven), standardized wealth rank, mother’s marital status (fiancée, polygyny, or monogamy), and nursing by mother’s marital status interaction.
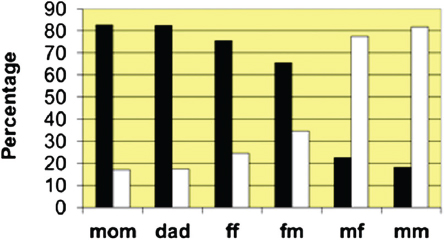
FIGURE 14.10 The percentage of children who lived in the same village as each kind of relative (dead relatives were excluded from the analysis). Black bars represent the children that lived with the relative, and white bars represent the children that did not live with the relative. ff, father’s father; fm, father’s mother; mf, mother’s father; mm, mother’s mother.
Maternal grandmothers had an adverse impact on the growth of girls, regardless of whether the girls lived matrilocally or patrilocally. A possible explanation is that growth is slowed by the energetically demanding work that girls perform for their maternal grandmothers. In the literature on grandmothering, it is usually assumed that the most beneficial grandmother is the maternal one (Hawkes et al., 1997, 1998). The data for the growth of Dogon girls point in the opposite direction.
CONCLUSION
If the term cooperative breeding is restricted to human populations in which adult offspring delay or forego reproduction to act as helpers, then the Dogon would not qualify because girls usually initiate reproduction by about age 19 years and boys by age 25 years. It would potentially apply, however, to 19th and early 20th century European farming communities in Ireland and elsewhere that had an unusually high percentage of individuals who postponed reproduction to their thirties or who were permanently celibate. Ireland was akin to a saturated habitat in which opportunities to reproduce were restricted by the availability of farms. It has been suggested that unmarried siblings often emigrated or acted as helpers (Strassmann and Clarke, 1998).
The Dogon pattern is very different. Rather than disqualify the Dogon as cooperative breeders a priori because they do not delay repro-
duction, I searched for evidence for cooperative breeding behavior in sibling groups, in families, and on the part of grandparents. Siblings competed with each other, producing a tradeoff between the number of maternal siblings and growth and survival. To understand why children work or care for younger siblings, parental coercion is a possibility that needs further investigation. It did not take a village to raise a child; instead, children in nuclear families did as well as children in extended families. The hazard of child mortality to age 5 years was twofold higher if the paternal grandmother was alive rather than dead. In the first 5 years of life, mothers were essential for child survival and could not be replaced by other kinds of kin. These results do not align with the expectations of cooperative breeding theory. Kin selection theory was more helpful for understanding the dialectic between cooperative and competitive interactions in the Dogon, especially when conjoined with parent–offspring conflict and life history theory. The predicted linear relationship between child survival and relatedness of the child to the WEG boss was not supported. However, kin selection theory was helpful for understanding the dynamics of polygyny and WEG instability over time.
METHODS
Subjects
The study population was an entire cohort of 1,700 children in nine Dogon villages who were enrolled between 1998 and 2000. These children have been followed prospectively since their initial enrollment to the present date (2011). The criterion for enrollment was age ? 5 years on May 1, 1998, or born during the first 2 years of follow-up study. Subject cooperation for participation in the cohort study was >99.9%. Participation in the anthropometric measurements (height or supine length in centimeters and weight in kilograms) was 91% (1998), 83% (1999), 85% (2000), 81% (2004), 79% (2007), and 85% (2010). The primary reason for nonparticipation was that the child was living outside the study area that year. Approval to conduct this study was obtained from the University of Michigan Internal Review Board, the Malian government, local authorities, and village chiefs and elders. Informed consent (adults) or assent (children) was obtained from the subjects.
WEG Structure
An annual door to door census was conducted of the 652 WEGs in the nine villages. This census asked the identity of the head of the
family (WEG boss) and each WEG member’s name, parents’ names, sex, year of birth, marital status [unmarried, fiancée (tanganu), or married (tanga)], spouses’ names, spousal rank order (sole, first, second, third, or fourth wife), wife type [arranged (ya bire), nonarranged (ya kezu), or levirate: married to the deceased husband’s brother (ya pani)], and religion (indigenous, Muslim, Catholic, Protestant, or free thinker/ agnostic). From these data, other family structure variables were calculated to reflect the social niche of the child (e.g., number of maternal and paternal siblings, ratio of married women to married men in the WEG, and total number of children aged 10 years and younger). Data were also gathered on the survival status of the children’s parents and grandparents (dead or alive) at the time the children were born and at the time of the interview.
WEG Wealth
Forty-one informants who lived in the nine villages independently ranked the relative wealth of each WEG in their natal village, producing strong agreement among informants [Cronbach’s ?: C = 0.89 ± 0.06 (1999) and C = 0.93 ± 0.03 (2000)]. After doing their individual rankings, the informants also met together to produce a consensus ranking.
Children’s Survival
Because most child mortality in the Dogon occurs by age 5 years (US Department of State, 2010), I was primarily interested in survival in early childhood. Therefore, in studying the relationship between family structure and child survival, I used the field data from the years 1998–2001. The statistical analysis employs a Cox proportional hazards model to analyze survival as a function of time-varying covariates (Cox, 1972; Allison, 1995). The covariates measured in a given year served as predictors of survival from that year to the next (e.g., covariates measured in 1998 were used to predict survival to 1999). After the initial year (1998), I had 3 years of follow-up (1999, 2000, and 2001). The dependent variable in the statistical model was the duration of survival time for each child across all years that he or she was in the study between 1998 and 2001. If a child survived to the end of a given year, I calculated her survival time as the child’s age at the end of the year minus her age at the start of the year. If she died during the year, her survival time was her age at death minus her age at the start of the year. If she was lost to follow-up, her survival time was calculated as her age at midyear minus her age at the start of the year. If she was born and died between annual censuses, her survival time was her age at death. The survival analyses were carried
out using Proc Phreg in the statistical software SAS 9.2 (SAS Institute, Inc., 2008), which allowed for control for multiple observations of the same child. The statistical models included a random effect for mother.
Behavioral Data
From 1986 to 1988, an instantaneous behavioral scan was conducted during daylight hours in the fields and the initial study village in all months of the year. The date, time, identification, and activity of each person were recorded (n = 5,097 observations).
Children’s Growth
I calculated height-for-age z scores in relation to the international World Health Organization (WHO) standard that is appropriate for breastfed children in developing countries (World Health Organization, 2005). These z scores quantify the distance (measured in SDs) of a given child’s height relative to the mean for the reference population. I calculated growth as the change in height-for-age z scores. The covariates in a given year were used to predict the change in the child’s height-for-age z score from that year to the next. The statistical analysis used a linear mixed model (Littell et al., 1996) that (i) used a repeated measures design to take into account the autocorrelation in the growth of a given child between years and (ii) included the mother of the child as a random effect to take into account the correlation among maternal siblings. The analysis was carried out using Proc Mixed in the statistical software SAS 9.2 (SAS Institute, Inc., 2008).
WEG Fissioning
Males (n = 1,218) belonging to 29 patrilineages in 10 villages provided genealogical information for their paternal and maternal ancestry as far back as they could remember. From the oral histories, I made patrilineal pedigrees in the program Progeny (Progeny Software, LLC, 2007) that have a depth of up to 11 generations from the youngest generation to the common ancestor. These pedigrees contain data on WEG composition and changes over time.
ACKNOWLEDGMENTS
I thank the Dogon for their generous participation in this research, including my Dogon field assistants for their valuable help. I am grateful to the Malian government for permission to conduct this study and
the following undergraduate and Master degree-level field assistants: Sylvie Moulin, Thomas Stevenson, Betsy Abrams, Sera Young, Brendan Hug, Lindsey Boynton, Joy Rowe, Riley Bove, Anne Lyne, Jean Baptiste, David Plate, Joanna Rogers, Robin Martz, Christine Sauve, and Russell Herrold. I thank Kathy Welch for statistical advice and Claudius Vincenz, Laura Betzig, and two anonymous reviewers for helpful comments. This research was supported by the Louis S. B. Leakey Foundation, National Science Foundation Grants SBR 9727229 and BCS 1029056, the American Philosophical Society, and National Institutes of Health Grant 09-PAF00653.






















