
Evolutionary Foundations of Human Prosocial Sentiments
![]()
JOAN B. SILK*†‡ AND BAILEY R. HOUSE*
A growing body of evidence shows that humans are remarkably altruistic primates. Food sharing and division of labor play an important role in all human societies, and cooperation extends beyond the bounds of close kinship and networks of reciprocating partners. In humans, altruism is motivated at least in part by empathy and concern for the welfare of others. Although altruistic behavior is well documented in other primates, the range of altruistic behaviors in other primate species, including the great apes, is much more limited than it is in humans. Moreover, when altruism does occur among other primates, it is typically limited to familiar group members—close kin, mates, and reciprocating partners. This suggests that there may be fundamental differences in the social preferences that motivate altruism across the primate order, and there is currently considerable interest in how we came to be such unusual apes. A body of experimental studies designed to examine the phylogenetic range of prosocial sentiments and behavior is beginning to shed some light on this issue. In experimental settings, chimpanzees and tamarins do not consistently take advantage of opportunities to deliver food rewards to others, although capuchins and marmosets do deliver food rewards to others in similar kinds of tasks. Although chimpanzees do not satisfy experimental criteria for prosociality in food delivery tasks, they help others complete tasks to obtain a goal. Differences in performance across
___________________
*Department of Anthropology and †Center for Society and Genetics, University of California, Los Angeles, CA 90095. ‡To whom correspondence should be addressed. E-mail: jsilk@anthro.ucla.edu.
species and differences in performance across tasks are not yet fully understood and raise new questions for further study.
It is not from the benevolence of the butcher, the brewer, or the baker that we expect our dinner, but from their regard to their own interest.
Adam Smith, The Wealth of Nations
How selfish soever man may be supposed, there are evidently some principles in his nature, which interest him in the fortune of others, and render their happiness necessary to him, though he derives nothing from it except the pleasure of seeing it.
Adam Smith, The Theory of Moral Sentiments
As Adam Smith pointed out more than 250 years ago, humans often act out of self-interest but also feel concern for the welfare of others. These sentiments come into conflict when selfish behavior produces negative impacts on others and when concern for others leads to altruistic behavior that reduces one’s own welfare. For evolutionary biologists, selfishness is a straightforward consequence of selective forces that favor behaviors that enhance individual fitness. Natural selection is not expected to favor indiscriminate altruism, because altruists bear the costs of the altruistic behaviors that they perform; this reduces their relative fitness. Altruism can only evolve if altruists confer benefits selectively on others who carry the same altruistic alleles. Kin selection (Hamilton, 1964a) and reciprocal altruism (Trivers, 1971; Axelrod and Hamilton, 1981) both rely on this principle. Selection can favor altruism to close relatives, because recent common descent provides a reliable cue of genetic similarity. In the case of reciprocity, past behavior of other group members provides a cue about whether they carry alleles that lead to altruistic behavior. These processes can generate biases in favor of kin and reciprocating partners but not a general predisposition to behave altruistically to others.
Altruism is also paradoxical for many economists. Selfishness is the expected outcome when, as is often assumed, utility functions only include personal consumption. However, as Adam Smith realized, human behavior deviates from the expected behavior for self-interested actors. Experimental studies in behavioral economics that are designed to bring conflicts between self-interest and altruism into sharp relief show that people value their own welfare but also value the welfare of others (Henrich et al., 2004; Fehr and Schmidt, 2006). This body of work provides insight about some of the dimensions of our altruistic social preferences.
DIMENSIONS OF ALTRUISTIC SOCIAL
PREFERENCES IN HUMANS
Generosity
In the Dictator Game, subjects are allowed to distribute an endowment between themselves and another player (Camerer and Thaler, 1995). The units of the endowment may take the form of cash or monetary equivalents that will be converted to cash at the end of the experiment. One player, the proposer, is given the opportunity to allocate any amount of his endowment to a second player, the recipient. In the standard form of the game, the proposer’s offer is relayed to the recipient anonymously; the two players never meet and never interact again. This eliminates reputational benefits or expectations based on reciprocity. A selfish player would keep the full endowment; an altruistic player would allocate some fraction of the endowment to the recipient. Typically, proposers allocate 20–30% of the endowment to the other player (Camerer and Thaler, 1995), indicating that they value the welfare of others but not as highly as they value their own welfare.
Trust
In the Trust Game, two players are given endowments. Player 1 can allocate any amount of her endowment, e, to Player 2; the experimenter will triple the allocation, and the full amount will be delivered to Player 2. Then, Player 2 is given the opportunity to make an allocation to Player 1. Player 2 can keep all of the money or send some money back to Player 1. If Player 1 sends her whole endowment to Player 2, then Player 2 would receive 3e. This would be added to Player 2’s initial endowment, e, and equal 4e. If Player 2 sends back one-half, both would get 2e, which is double their initial endowment. In contrast, if Player 1 sends Player 2 only one-half of the original endowment, then Player 2 will end up with only 2.5e (0.5e × 3 + e). If Player 1 trusts Player 2 to repay her, then it is best to send Player 2 the full amount. However, if Player 1 expects Player 2 to defect, then it is best to send nothing. If Player 2 is selfish, she would keep the whole amount; any money sent back to Player 1 is a form of altruism. In fact, the majority of people who take the role of Player 2 do send back money, and the amount that they send is proportional to the amount that they have received (Fehr and Fischbacher, 2003).
Punitive Sentiments
The Ultimatum Game (Camerer and Thaler, 1995) adds a second step to the Dictator Game. The proposer is given an endowment and makes
an allocation. Now, the recipient decides whether to accept or reject the proposer’s offer. If the recipient accepts the offer, each player gets the designated amount; if the recipient rejects the offer, neither one gets any money. The recipient has little material incentive to reject any nonzero offer, because this will result in a loss of income. Rejections constitute a form of altruistic punishment, because the recipient suffers a cost to punish the proposer; because the players will never interact again, this cannot be a strategy for improving the recipient’s payoffs in the future. The Ultimatum Game has now been played by thousands of people in dozens of countries all over the world. Recipients typically reject offers of less than 20%, and the size of the initial endowment has surprisingly little impact on rejections (Hoffman et al., 1996; Camerer, 2003; Henrich et al., 2006, 2010a).
In the Third-Party Punishment Game, a third party is given the opportunity to impose sanctions on the proposer in a Dictator Game (Fehr and Fischbacher, 2004). As in the standard Dictator Game, the proposer receives an endowment and can transfer any fraction of the endowment to a receiver. In this game, however, a third player is given an endowment and informed of the proposer’s allocation decision. The third player can spend one unit to reduce the proposer’s payoff by three units but cannot have any effect on the recipient’s payoff. A majority of subjects imposed sanctions on proposers who made offers of less than one-half of the endowment, and those that offered much less than one-half were punished more severely than those that made offers closer to one-half. It is noteworthy that people are willing to incur costs to punish others for making low offers, although they have not been directly harmed themselves (Fehr and Fischbacher, 2004).
Fairness
Proposers’ offers in the Dictator Game, recipients’ behavior in the Ultimatum Game, and responses of third parties in the Third-Party Punishment Game all suggest that people have a strong preference for equitable outcomes. Although people are more sensitive to inequities that disadvantage themselves than inequities that benefit themselves, a substantial majority of people are willing to reduce their own payoffs to produce more equitable outcomes for others (Fehr and Schmidt, 1999). Interestingly, people are less bothered by inequitable outcomes that are the product of chance events, such as a coin flip, than inequitable outcomes that are the result of deliberate human action (Blount, 1995; Camerer and Thaler, 1995).
LIMITS ON ALTRUISTIC SOCIAL PREFERENCES IN HUMANS
The evidence for generosity, trust, punitive sentiments, and fairness does not mean that all humans are indiscriminate altruists. Like many other animals, humans show strong nepotistic biases (Flinn et al., 2007; Sear and Mace, 2008; Hrdy, 2009) and also develop long-term relationships with reciprocating partners (Gurven, 2006; Allen-Arave et al., 2008). There is also substantial individual variation in social preferences. For example, in public goods games that continue across multiple rounds, a substantial fraction of subjects are contingent cooperators (Fischbacher et al., 2001; Fehr and Fischbacher, 2003). Conditional cooperators follow cooperative norms as long as other group members cooperate but stop cooperating if others defect. In the presence of contingent cooperators, a small number of selfish, uncooperative individuals can precipitate the collapse of group-level cooperation. Sanctions that make it costly to defect help prevent this from happening. In addition, humans show strong parochial biases, which favor group members over outsiders (Shinada et al., 2004; Bernhard et al., 2006).
CRITIQUES OF INTERPRETATION OF
BEHAVIORAL ECONOMICS GAMES
In behavioral economics games, players are paired with strangers in one-shot games to eliminate egoistic motives for altruism, including the opportunity for reciprocity and reputational benefits. Some researchers question the validity of these conditions by claiming that our psychology was designed for a world in which we lived in small groups of close kin and reciprocating partners. In such settings, there may be little opportunity for anonymous, one-shot interactions (Hagen and Hammerstein, 2006; Burnham and Hare, 2007). As a result, participants in experiments may find it hard to believe that their behavior is actually anonymous, and subjects may be influenced by subtle cues that influence their perceptions of being observed and the salience of reputational cues. Such cues matter, because our psychology is “exquisitely sensitive to cues that are (or were, under ancestral conditions) informative with respect to the likely profitability of co-operation in a given situation” (Haley and Fessler, 2005). According to this argument, people behave altruistically in behavioral economics experiments, because they are motivated to enhance others’ perception of their value as a cooperative partner. Thus, altruistic behavior is motivated by self-interest not other-regarding preferences.
This claim is partially supported by evidence that levels of contributions increase when subjects are exposed to subtle cues of being watched. For example, a pair of eyes on a sign that instructed users of a university coffee room to pay for their drinks produced more revenue than neutral
images (Bateson et al., 2006). A pair of eyes had a similar effect on littering in a university cafeteria (Ernest-Jones et al., 2010). Players in a public goods game that were faced with an image of an anthropomorphic robot on the computer screen contributed more than players faced with a blank screen (Burnham and Hare, 2007). Haley and Fessler (2005) found that contributions in an anonymous Dictator Game were higher when the computer monitor displayed a pair of stylized eyes than when it displayed plain text. The effects of eyes in the Dictator Game have been replicated (Rigdon et al., 2009; Mifune et al., 2010; Oda et al., 2011) and seem to be a function of the expectation of future benefits, not fear of punishment (Oda et al., 2011).
However, cues of being watched do not have the same effects in all games. Fehr and Schneider (2010) found that the stylized eyes that Haley and Fessler (2005) used had no effect on the amount that Player 1 transferred in an anonymous trust game. In contrast, when players were told that their partners would be informed about the amount that they had transferred in previous rounds (but not their identity), transfer amounts doubled. Thus, people were strongly influenced by explicit reputational information but not by the kinds of subtle cues that might have suggested that they were being watched.
If subtle cues of being watched affect cooperative behavior, then the actual presence of others ought to amplify cooperative behavior. To assess this, Lamba and Mace (2010) conducted a series of Ultimatum Games in which they manipulated the degree of anonymity that subjects experienced. In one condition, subjects played an anonymous double-blind game alone in a room (Anonymous/Private). In a second condition, subjects played an anonymous double-blind game in a room with other subjects (Anonymous/Public). In the third condition, subjects’ offers were announced to all participants who were together in the same room (Public/Public). Proposers’ offers did not differ in the Anonymous/Private and Anonymous/ Public conditions, suggesting that players were confident that their offers were anonymous and were not affected by subtle cues of being watched by others. In contrast, knowledge that their offers would be made public significantly increased offers in the Public/Public condition, and this effect was enhanced when proposers were acquainted with others in the room when the experiment was conducted.
At this point, there is no consensus about the importance of subtle cues of being watched on decision making in behavioral economic games or the impact of such cues on prosocial behavior in more naturalistic settings. It is not yet clear whether differences in the effectiveness of cues of being watched depend on the game being played or details of the experimental procedures. In contrast, there is abundant evidence that explicit
reputational information has clear and unambiguous effects on prosocial behavior across games and settings.
MOTIVES UNDERLYING HUMAN ALTRUISM
Behavioral economic experiments are designed to elicit preferences that guide choices about payoff outcomes, but they do not provide direct information about the psychological mechanisms that produce these preferences. This is important, because behaviors that have similar outcomes can be the product of very different mechanisms. Human altruism might be motivated by empathy and concern for the welfare of others. Alternatively, altruism might be prompted by more selfish, egoistic concerns, such as improving one’s reputation for generosity. If people are motivated by empathy and concern for the welfare of others, their ultimate goal is to provide benefits to others, and any benefits that individuals accrue are incidental by-products. However, if people are motivated by egoistic motives, then the benefits that they deliver to others may be incidental to their primary goals. People might be motivated to help others, because helping brings rewards to themselves (including reputational benefits or future material gains), prevents punishment (including material sanctions), or reduces aversive arousal that comes from observing others in need.
Batson (1991, 2011) has conducted a long series of experiments that were designed to assess the relative importance of egoistic and empa-thetic motives in altruistic predispositions [reviewed in Batson (1991) and (2011)]. For example, to assess the hypothesis that altruism is prompted by the desire to reduce aversive arousal, Batson (1991, 2011) conducted a series of experiments in which subjects observed a worker receiving electric shocks and were told that they could help by volunteering to take the shocks themselves. (In reality, the workers were confederates, and no electric shocks were administered to anyone.) The experimenters manipulated empathetic responses and how easy it was for subjects to avoid the aversive stimulus: In the easy condition, subjects would not see the worker being shocked after they made their decision, but in the difficult condition, subjects would continue to watch the worker being shocked. If altruism is the product of egoistic motives, then subjects should help more when escape is difficult than when escape is easy under the high-empathy condition. However, if altruism is the product of altruistic motives, then subjects should be equally likely to help under both conditions. Subjects generally conformed to the latter pattern, suggesting that they were motivated to help for altruistic, not egoistic, reasons. The results of the full series of experiments and related experimental work by other investigators (Piliavin and Charng, 1990) are consistent with the hypothesis that
altruistic behavior is shaped by empathic concern for the welfare of others, and it is not motivated entirely by self-interest or reputational concerns.
PHYLOGENETIC FOUNDATIONS OF HUMAN ALTRUISM
Social relationships play an important role in the daily lives of both human and nonhuman primates. Social bonds seem to enhance the ability to cope with chronic stressors, such as low social status, or acute stressors, such as the recent loss of preferred partners or immediate risk of infanticide [reviewed in Cheney and Seyfarth (2009). In humans, social support is correlated with better physical and mental health as well as lower mortality risks (Thorsteinsson and James, 1999; Cacioppo et al., 2000; Taylor et al., 2000; Kendler et al., 2005). Similar kinds of findings are accumulating for a range of nonhuman species, including rodents (Weidt et al., 2008; Yee et al., 2008), dolphins (Frère et al., 2010), wild horses (E. Z. Cameron et al., 2009), female baboons (Silk et al., 2003a, 2009, 2010b), and male macaques (Schülke et al., 2010).
There is good evidence that, like humans, monkeys and apes form strong and lasting ties, particularly with close kin (Silk, 2009) and reciprocating partners (Cheney, Chapter 15, this volume), and close social bonds are the foundation for cooperation in nonhuman primate groups (Mitani, 2009; Silk et al., 2010a). Like humans, nonhuman primates also have strong in-group biases. Responses to strangers and members of neighboring groups range from passive avoidance to active hostility (Crofoot and Wrangham, 2010). However, despite these intriguing parallels in the patterns of cooperation and the correlates of social bonds among humans and other primates, there are also important differences in the scope of cooperation. In most primate species, there is no sexual division of labor and little active food sharing. Primates do not cooperate with members of other groups in collective activities, such as warfare, territorial defense, or trade.
DIMENSIONS OF ALTRUISTIC SOCIAL
PREFERENCES IN NONHUMAN PRIMATES
Differences in the scope and pattern of cooperation between humans and other primates may be reflected in differences in the nature of their social preferences. Researchers have recently begun to explore the dimensions of altruistic social preferences in nonhuman primates in systematic ways using the same kinds of tools that behavioral economists have used to assess human social preferences. These experimental methods are useful, because the difficulties of identifying the motives underlying altruism are compounded when we extend the analysis to other species
(Silk, 2007a). A chimpanzee who has just caught a colobus monkey might allow another male to take part of his kill, because he feels empathy for the other’s hunger, because he can forestall the other’s efforts to take the entire carcass by force, or because he received meat from the other male on the previous day. These sorts of ambiguities have prompted a series of experiments that are designed to determine whether other primates have preferences for outcomes that benefit others, a sense of fairness, and punitive sentiments.
GENEROSITY AND PROSOCIAL BEHAVIOR IN CHIMPANZEES
The Prosocial Test is similar to a discrete Dictator Game. Proposers are presented with a choice between two options (Silk et al., 2005). One option delivers a food reward to the proposer and nothing to another individual in an adjacent enclosure. For convenience, this is referred to as the 1/0 option (the reward for the proposer is given on the left side of the slash and the reward for the recipient is given on the right side of the slash). The other option delivers a food reward to the proposer and an identical reward to the other individual (the 1/1 option).
Chimpanzees have prepotent biases for larger number of rewards, regardless of their distribution (Boysen and Berntson, 1995; Uher and Call, 2008). This means that proposers might prefer the prosocial option (1/1) over the selfish option (1/0), because they have biases in favor of larger numbers of rewards. Therefore, a nonsocial control condition was included in which no recipient was present to receive rewards.
Actors’ choices in the Prosocial Test provide insights about their social preferences. If individuals are concerned about the welfare of others, they will choose the 1/1 option over the 1/0 option. Moreover, if they are motivated by concern for the welfare of others and not by prepotent biases for a larger number of rewards, their bias in favor of the prosocial option will be stronger when another individual is present (test condition) than when the actor is alone (nonsocial control condition). Alternatively, individuals might view potential recipients as rivals or competitors for a fixed quantity of rewards, and they might be motivated to deprive them of resources. If so, they will choose the 1/0 option over the 1/1 option, and their bias in favor of this selfish option will be stronger in the test condition than in the control condition. Finally, if actors are indifferent to the welfare of others, they will choose at random, and their choices in the control and test conditions will not differ. The major advantage of the Prosocial Test over the Dictator Game is that it costs the proposer nothing to confer a prosocial outcome, meaning that any positive altruistic tendencies should manifest clearly.
The Prosocial Test and several closely related variants have now been conducted with chimpanzees from several different populations using a number of different experimental apparatuses. Jensen et al. (2006) conducted three related experiments. In one, the payoff distribution was 1/1 vs. 1/0; in the second experiment, the actor could provide rewards to the recipient but obtained nothing for herself (0/1 vs. 0/0), and in the third experiment, the actor could prevent the recipient from obtaining rewards. In all three experiments, there was no difference between the test condition and the nonsocial control condition. Yamamoto and Tanaka (2010) trained chimpanzees to associate one colored button with a 1/1 payoff and a differently colored button with 1/0 payoff. The chimpanzees were as likely to choose the 1/1 option in the test condition as in the nonsocial control condition. Vonk et al. (2008) conducted two experiments in which actors could deliver food rewards to themselves and others with independent but identical actions. In both of these studies, the actors were as likely to deliver rewards in the nonsocial control condition as in the social test condition.
Brosnan et al. (2009) and Yamamoto and Tanaka (2010) allowed participants to switch roles within trials, and therefore the proposer in one round became the recipient in the next round. This manipulation had no impact on the level of prosocial responses in either study. The rate of prosocial responses in the iterated Prosocial Test was the same as the rate of prosocial responses in one-shot versions of the Prosocial Test conducted with the same animals.
Thus, in the Prosocial Test, chimpanzees consistently act as if they are indifferent to the welfare of other individuals. This set of findings has been both surprising and controversial, because chimpanzees cooperate in a wide range of contexts, share food in the wild (Muller and Mitani, 2005), collaborate effectively in mutualistic tasks in the laboratory (Melis et al., 2006a,b), and seem to be helpful in other experimental paradigms. Before we turn to experiments in which chimpanzees show helpful behavior, we consider a number of explanations that have been proposed to explain chimpanzees’ behavior in the prosocial task.
It is possible that proposers did not differentiate between the test and control conditions in these experiments, because they did not understand how the experimental apparatuses worked. However, subjects’ understanding of the experimental apparatuses was explicitly tested in several studies (Jensen et al., 2006; Brosnan et al., 2009; Yamamoto and Tanaka, 2010). Alternatively, the chimpanzees might have found it difficult to track the distribution of food items when rewards were delivered to themselves and their partners simultaneously (Warneken and Tomasello, 2009). However, when Jensen et al. (2006) tested chimpanzees with a payoff distribution that did not provide any rewards for the actor, they found
that overall response rates dropped substantially, and proposers still did not distinguish between the test and nonsocial control conditions. In the experiments conducted by Vonk et al. (2008), proposers could deliver food rewards to themselves and others with separate actions. Proposers almost always obtained their own rewards first. After they had obtained rewards for themselves, they sometimes delivered the other reward. However, as the experiment progressed, they were less likely to deliver rewards to the other enclosure in both test and control conditions. Thus, subjects in these experiments became less generous (and no more discriminating), as they gained more familiarity with the test apparatus.
Although chimpanzees seem to have some understanding of others’ desires and intentions in competitive situations (Hare et al., 2000, 2001; Kaminski et al., 2008), proposers may have been unaware of their partners’ desires for rewards in the prosocial test (Warneken and Tomasello, 2009). However, analyses of recipients’ begging gestures in two experiments cast doubt on this possibility (Vonk et al., 2008). Recipients that made begging gestures consistently directed them to the option that contained food for themselves, but begging had no consistent impact on proposers’ responses.
It is also possible that proposers did not choose the 1/1 option more often, because chimpanzees often compete over access to food in the wild, and prosocial preferences are muted in the presence of food (Warneken and Tomasello, 2009). However, if chimpanzees view food as a limited, zero-sum resource, they would be expected to show a strong preference for the 1/0 over the 1/1 option. This was not seen in any of the studies.
HELPFUL RESPONSES OF CHIMPANZEES IN
OTHER EXPERIMENTAL SETTINGS
The conclusions derived from the prosocial test with chimpanzees conflict with results derived from experimental paradigms in which one individual is given the opportunity to help another individual obtain a goal. The first of these studies was conducted by Warneken and Tomasello (2006) with three young chimpanzees that were paired with their human caretaker in several different task situations. In each task situation, there was one version in which help was needed (test), and a second very similar version in which no help was needed (no-need control). The chimpanzees responded positively to caretakers’ requests for help in several tasks that involved retrieving out-of-reach objects and consistently differentiated between the control and test conditions. However, they did not meet this criterion for a number of other kinds of tasks. Warneken and Tomasello (2006) suggest that this may have been because they did not grasp what the recipient needed.
The three young chimpanzees that Warneken and Tomasello (2006) tested had a close relationship with their caretakers and had been rewarded for accommodating behavior in the past (Warneken et al., 2007). To determine whether helpful behavior would extend to unfamiliar humans, Warneken et al. (2007) performed a second set of experiments. In these experiments, two experimenters struggled over a stick, and then, the victor placed the stick out of the loser’s reach. In the test condition, the loser stretched out his arm and reached to the stick, and in the control condition, the loser looked at the object but did not reach for it. The chimpanzees were significantly more likely to retrieve the stick in the test condition than in the control condition. In a second experiment, the cost of helping was increased, because the chimpanzees had to climb 2.5 m into an overhead compartment to retrieve the stick. The chimpanzees were actually more likely to retrieve the stick in this experiment than in the first experiment, and they did not distinguish between the test and control conditions. Warneken et al. (2007) speculated that this was “likely due to a carryover effect from experiment 1 in which subjects had possibly learned that the experimenter wanted the object.”
Chimpanzees’ willingness to help humans complete certain tasks is mirrored in their behavior to conspecifics trying to complete a task. Warneken et al. (2007) conducted a third set of experiments in which one chimpanzee was given an opportunity to help another chimpanzee gain access to a locked room. In this experiment, there were four adjacent rooms. The doors to two rooms were fastened by chains held in place by pegs. The actor was confined to one room and could reach a peg that released one of the two doors, but he could not enter either of the locked rooms. The recipient could not release either of the doors but could enter one of the rooms that the actor could unlock. Recipients were motivated to gain access to the locked room, because it contained food rewards, but these rewards were not visible to the actors. In the test condition, food rewards were placed in the room that the actor could unlock and the recipient could enter; in the control condition, food rewards were placed in the room that the recipient could not enter. Actors were significantly more likely to remove the peg and release the door in the test condition than in the control condition.
Chimpanzees also provide tools that others need to complete food-related tasks (Yamamoto et al., 2009). In this experiment, two chimpanzees were placed in adjacent enclosures, and each was presented with a food task that required a particular tool (stick or straw). In the baseline condition, each chimpanzee was given the appropriate tool. In the test condition, the chimpanzee that needed the stick was given the straw, and the chimpanzee that needed the straw was given the stick. The chimpanzees were significantly more likely to transfer tools when they were given the
wrong tools than when they were given the right tools, and tool transfers generally occurred in response to explicit requests.
As noted earlier, there has been speculation that results in the Prosocial Test might reflect chimpanzees’ reluctance to provide others with food or their lack of understanding of others’ needs and desires. To test these two hypotheses, Melis et al. (2011) constructed an apparatus in which one chimpanzee could release a peg and cause a reward to roll down a ramp to the recipient in a separate enclosure. In one set of trials, the reward was connected to a chain so that the recipient could pull the reward to them after it was released. In another set of trials, the food rolled directly down the ramp and into their enclosure, and there was no chain for recipients to pull. These conditions were meant to encourage active and passive responses by recipients, respectively, but this manipulation was not entirely effective; therefore, analyses were based on the recipients’ level of activity across the reach and no-reach conditions. To evaluate whether helpful responses were inhibited when food rewards were present, some trials were conducted with food rewards, and other trials were conducted with tokens, which recipients could trade for food rewards out of the actor’s sight. The chimpanzees were significantly more likely to release the peg when food rewards were present than when tokens were present, and they were more likely to release the peg when recipients responded actively than when they responded passively. Melis et al. (2011) concluded that “the main finding of the present study is that recipients’ signaling is necessary to elicit helping behaviour.”
However, this conclusion is inconsistent with results derived from another study of helping behavior in chimpanzees. In this experiment, conducted by Greenberg et al. (2010), two chimpanzees had to pull on a rope to move a sliding platform along a set of parallel tracks. The apparatus was designed so that it could be baited to dispense rewards at the beginning, middle, or end position of each of the tracks. In the baseline condition, both chimpanzees obtained rewards when the platform was pulled to the middle condition. In one of two altruism conditions, one chimpanzee obtained a reward when the platform was pulled to the middle position, and the other chimpanzee obtained a reward when the platform was pulled to the end position. In the other altruism condition, one chimpanzee obtained a reward when the platform was pulled to the end position, but the reward for the other chimpanzee was simply placed on the apparatus. The participants were significantly more likely to pull the platform to the end position in the two altruism conditions than in the baseline condition, suggesting that the chimpanzees were willing to continue pulling even after they had obtained their own reward. In this setting, direct solicitations or requests for help were rarely observed.
RECONCILING RESULTS FROM EXPERIMENTAL STUDIES
OF PROSOCIAL BEHAVIOR IN CHIMPANZEES
In the Prosocial Test, chimpanzees are as likely to choose the prosocial option when another individual is present as when they are alone, and therefore, they do not meet the experimental criterion for prosocial preferences. In several helping tasks, chimpanzees are more likely to provide help when it is needed than when it is not needed and thus, satisfy the criterion for prosocial behavior within these protocols. A number of factors have been invoked in an effort to determine why chimpanzees pass some tests but fail others, including cognitive demands of the task, actors’ preoccupation with their own rewards, competitive attitudes to food, limited understanding of what others want, and salience of others’ requests. However, none of these explanations seem to be consistent with the full body of evidence.
The focus on whether chimpanzees pass or fail various experimental tests has obscured an important feature of the data. Across the full range of experiments and test treatments, chimpanzees behave prosocially to their partners about one-half of the time (median = 0.51, interquartile range = 0.23). On average, the rates of prosocial responses tend to be higher in the Prosocial Tests than in the helping tasks (Wilcoxon rank sum test, z = -1.913, P = 0.0558, n1 = 10, n2 = 12) (Fig. 16.1).
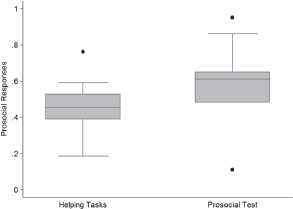
FIGURE 16.1 Rate of prosocial responses. The median and interquartile range of prosocial response rates in the test condition of all studies of prosocial behavior are plotted here. For studies that reported response rates for different categories of actor–recipient pairings or different categories of behavior of recipient, all values are included.
PROSOCIAL BEHAVIOR IN OTHER PRIMATES
If ecological factors shape predispositions about the fitness benefits of cooperation, then prosocial preferences in food distribution tasks might be expected to emerge in species that share food more willingly than chimpanzees do, such as cooperatively breeding marmosets and tamarins as well as capuchins. Marmosets and tamarins are cooperative breeders, and adults and immature helpers often provision younger individuals with food (Brown et al., 2004). In the laboratory, capuchins are remarkably tolerant of others’ efforts to take portions of their food (de Waal, 1997a, 2000).
Burkart et al. (2007) conducted a modified version of the prosocial test with marmosets. These animals are significantly more likely to choose 0/1 over 0/0 when others are present than when they are alone. This pattern held for male donors paired with adult female and immature recipients and for adult females paired with immature recipients, but it did not hold for subadult females paired with immature recipients. The scope of prosocial responses roughly parallels food-sharing patterns in naturalistic settings.
Tests for prosocial behavior in tamarins, which also breed cooperatively, have produced mixed results. Cronin et al. (2009) paired tamarins with their long-term mates and offered proposers a choice between 1/0 and 1/1 in one experiment and a choice between 0/0 and 0/1 in a second experiment. Proposers’ choices were not influenced by the presence or absence of their mates. In another study of tamarins, the proposer was placed in the middle of three adjacent enclosures (Stevens, 2010). The proposer was able to pull one handle that brought a tray within reach of itself and the occupant of an adjacent enclosure (with a one-to-three payoff structure) or a second handle that brought a tray to within reach of itself and an empty enclosure (also with a one-to-three payoff). Proposers nearly always pulled to obtain food for themselves when the adjacent enclosures were empty, but the rate of pulling declined sharply when they were unable to obtain food for themselves. In trials in which they could deliver food to an empty cage or their mates, they chose at random.
However, in a third experiment, tamarins showed more positive responses to their partners (Cronin et al., 2010). In this experiment, tama-rins could deliver food rewards to their mates (but not themselves) after their mates had delivered food to them (reciprocity condition) or had not delivered food to them (no reciprocity condition). Their behavior to their mates was compared with their behavior when the adjacent cage was empty (nonsocial control). The tamarins were significantly more likely to deliver food rewards to their mates in the reciprocity condition than in the no-reciprocity condition. Moreover, they were significantly more likely to deliver rewards to their partners in the reciprocity condition than in the nonsocial control condition. Cronin et al. (2010) argue that the tamarins’
behavior is evidence for prosocial preferences and not contingent reciprocity, because the effects only emerged in the last one-third of the 5-minute trials. However, this leaves open the question of why the tamarins did not provide rewards to their partners in the no-reciprocity condition.
There have also been several studies of prosocial preferences in capuchins. Lakshminarayanan and Santos (2008) offered capuchins a choice between two options. In some test trials, the proposer received a low-quality reward (L) and could deliver a low- or high-quality (H) reward to the recipient (L/L vs. L/H). In other test trials, the proposer received a high-quality reward and could deliver a low- or high-quality reward to the recipient (H/L vs. H/H). Behavior in test trials was compared with control trials in which no recipient was present and control trials in which no recipient was present and the proposer had access to the other compartment. Overall, the capuchins were significantly more likely to choose the option that delivered the high-value reward when another monkey was present than when they were alone (and could not obtain the reward themselves). However, there was considerable variability across subjects, and the magnitude of the difference between the test and control conditions was small. Four of seven subjects showed a preference for the prosocial option when they chose between H/L and H/H, whereas three of seven subjects showed a preference for the prosocial option when they chose between L/L and L/H.
de Waal et al. (2008) trained capuchin monkeys to associate tokens with particular payoffs for themselves and another individual in a separate enclosure, and they monitored their choices of tokens across a series of trials. If monkeys chose one token, the experimenter delivered one reward to them and another reward to the other individual (1/1 payoff); if they chose the other token, the experimenter delivered one reward to them and nothing to the other individual (1/0 payoff). This experiment did not include a nonsocial treatment to control for the effects of prepotent biases for larger numbers of rewards. Monkeys were increasingly likely to choose the token associated with the 1/1 payoff as the experiment progressed, and all statistical analyses were limited to the last one-third of the experimental trials. In these trials, capuchins chose the 1/1 option significantly more often than expected by chance when they were paired with kin or nonkin, but their choices dropped to chance levels when they were paired with monkeys from another group. The monkeys’ preferences for the prosocial option also declined to chance levels when the 1/1 option provided a more highly valued reward to the recipient than the proposer. This experiment also included a condition in which visual contact between the proposer and recipient was blocked by an opaque partition. The partition had a small window, and therefore, the proposer knew that another individual was present and the identity of that individual. When the
partition was in place, monkeys behaved antisocially—they chose the 1/1 option significantly less often than would be expected by chance.
In the third study on capuchins, Takimoto et al. (2010) manipulated the relative rank of the proposer and recipient and the relative value of rewards delivered to both parties. Recipients were either the alpha male or the lowest ranking female in the group. In some trials, proposers received rewards of medium value (M) and could deliver rewards of high (H) or low (L) value to recipients (M/H vs. M/L). In other trials, proposers received rewards of high value and could deliver rewards of high or low value (H/H vs. H/L). Proposers’ behaviors were compared when recipients were present or absent. Overall, proposers were more likely to choose generous options when they were paired with the subordinate recipient than when they were alone. However, this effect disappeared when they were paired with the dominant recipient.
Takimoto et al. (2010) also conducted a set of trials in which proposers could not see recipients during trials. When they were paired with a dominant recipient, they were significantly less likely to choose the generous option in the recipient-present condition than in the no-recipient condition. In contrast, when proposers were paired with the subordinate recipient, they did not differentiate between the two conditions.
The results from these three studies suggest that capuchin monkeys have preferences for outcomes that benefit other group members. However, their preferences for generous outcomes disappear or are reversed when proposers and recipients cannot see each other. de Waal et al. (2008) suggest that this is because capuchins derive rewards from “seeing the partner receive or consume food.” However, this does not explain why the proposers chose the 1/1 option significantly less often than expected by chance when visual access was blocked (Barnes et al., 2008; de Waal et al., 2008). It is possible that actors have preferences for antisocial outcomes, but antisocial preferences are suppressed when others can see them and can potentially take punitive action.
There has been only one instrumental helping task conducted with monkeys. The experiment was designed to replicate the experiments in which a human experimenter reaches for an inaccessible object (Warneken et al., 2007; Yamamoto et al., 2009). Unlike the chimpanzees, the capuchins did not consistently distinguish between the test and control conditions (Barnes et al., 2008). The capuchins were strongly motivated by the availability of rewards for themselves but not by the opportunity to provide help to others.
PUNISHMENT
Although primates and other animals often use aggression to manipulate the behavior of others to their own advantage (Clutton-Brock and Parker, 1995), evidence for third-party punishment is scarce (Jensen, 2010). Jensen et al. (2007a) adapted the Ultimatum Game for chimpanzees to examine their propensity to impose punishment on conspecifics. In this case, one chimpanzee that played the role of the proposer was able to choose between two set distributions of rewards by pulling one of two rods. To accept the proposer’s offer, the responder pulled another rod that brought the food rewards to within reach of both individuals, allowing each to claim their respective rewards. If the responder did not pull the rod, neither one got any food. The chimpanzees were offered a series of different options across trials. One option in every trial provided eight pieces of food to the proposer and two pieces to the responder (eight-to-two payoff). The other option provided a distribution of five to five, eight to two, or ten to zero. Proposers strongly preferred offers that benefited themselves (e.g., eight to two over five to five), but responders rarely rejected any nonzero offers. Moreover, responders showed little evidence of arousal in any of the trials.
FAIRNESS
Research on fairness and inequity aversion in primates was initiated by Brosnan and de Waal (2003), who trained tufted capuchins to exchange tokens for food rewards. The monkeys consistently offered experimenters tokens in exchange for small pieces of cucumbers, but some individuals refused to complete exchanges after they saw other group members receive more highly valued rewards in exchange for tokens or saw other group members obtain more highly valued rewards without exchanging tokens.
Brosnan and de Waal (2003) suggested that their findings were evidence of inequity aversion, although critics pointed out that the monkeys increased the extent of inequity by refusing to complete exchanges (Henrich, 2004a). Brosnan and de Waal (2003) acknowledged this point and argued that monkeys are averse to inequities that disadvantage themselves but not inequities that favor themselves (Brosnan and de Waal, 2004). Others argued that monkeys’ responses might have been prompted by frustration at seeing more preferred foods that were inaccessible to themselves (Dubreuil et al., 2006), frustration at being offered less preferred foods after seeing more desirable foods (Roma et al., 2006), or violation of expectation and loss aversion (Chen and Santos, 2006).
At this point, more than a dozen experimental studies of inequity aversion have been conducted in monkeys and apes. S. F. Brosnan et al.
(2010) found that negative responses to inequity were consistently found when subjects were required to perform a task to obtain food, whereas no responses to inequity were found when subjects were able to obtain food without performing a task. However, this generalization does not fit at least one study of great apes that used a token exchange task modeled on the task that S. F. Brosnan et al. (2010) used. Bräuer et al. (2009) found no evidence for inequity aversion among orangutans, bonobos, or chimpanzees.
CONCLUSIONS
Altruism and mutualistic cooperation play important roles in the lives of nonhuman primates, but there are important differences in the scope of altruistic behavior between humans and other primates. In other primates, altruism is strongly biased in favor of kin and reciprocating partners, and it is never extended to strangers. Primates use aggression to deter competitors and rivals, but there is no compelling evidence of third-party punishment. Unlike humans, nonhuman primates show no aversion to inequitable distributions of resources that favor themselves.
It is important to continue efforts to chart the size and dimensions of the gap between humans and other primates if we want to understand the evolutionary forces that have shaped human social preferences. Evidence that closely related primates, particularly great apes, have altruistic social preferences would suggest that our social preferences were built on a set of ancestral motivations that facilitated altruism to kin and reciprocating partners, mutualistic activities with group members, punitive behavior to competitors, antagonistic attitudes to strangers, and concern for reputational status. At the same time, evidence that closely related primates lack the kinds of altruistic social preferences that characterize modern humans would suggest that emergent forces, possibly including cultural group selection (Richerson and Boyd, 2005), demands of raising slow-growing offspring (Hrdy, 2005b, 2007; Jaeggi et al., 2010), knowledge-intensive human foraging strategy (Kaplan et al., 2000, 2003), or risks associated with uncertain hunting returns (Winterhalder, 1986), have transformed us in consequential ways and given rise to important differences in the scope and scale of cooperation, our capacity for empathy and compassion, the development of moral sentiments, and the willingness to enforce culturally specified social norms.
We believe that well-designed experimental studies of social preferences in other primates provide important insights about the nature of social preferences. Such studies should be designed to test hypotheses that are grounded in evolutionary theory and our knowledge of the natural history, social organization, and cognitive capacities of our study subjects.
It would be desirable to establish collaborative efforts to standardize methods and procedures and to replicate experiments in different test populations and species. Such studies would be useful for several reasons. First, they would strengthen comparative analyses by providing a richer database to test functional hypotheses about the factors that contribute to variation in prosocial preferences across species. Second, by using standardized methods, we can generate larger samples of behavior within species. This would generate more robust characterizations of behavior and enable us to begin assessing the extent of intraspecific variation in behavioral responses.




















