
4
Comparative Phylogeography of Oceanic Archipelagos: Hotspots for Inferences of Evolutionary Process
KERRY L. SHAW*‡ AND ROSEMARY G. GILLESPIE†
Remote island archipelagos offer superb opportunities to study the evolution of community assembly because of their relatively young and simple communities where speciation contributes to the origin and evolution of community structure. There is great potential for common phylogeographic patterns among remote archipelagos that originate through hotspot volcanism, particularly when the islands formed are spatially isolated and linearly arranged. The progression rule is characterized by a phylogeographic concordance between island age and lineage age in a species radiation. Progression is most likely to arise when a species radiation begins on an older island before the emergence of younger islands of a hotspot archipelago. In the simplest form of progression, colonization of younger islands as they emerge and offer appropriate habitat, is coincident with cladogenesis. In this chapter, we review recent discoveries of the progression rule on seven hotspot archipelagos. We then discuss advantages that progression offers to the study of community assembly, and insights that community dynamics may offer toward understanding the evolution of progression. We describe results from two compelling cases of progression where the mosaic genome may offer insights into contrasting demographic histories that shed light on mechanisms of speciation and progression on remote archipelagos.
__________________
* Department of Neurobiology and Behavior, Cornell University, Ithaca, NY 14853; and †Environmental Science Policy and Management, University of California, Berkeley, CA 94720. ‡To whom correspondence should be addressed. Email: KLS4@cornell.edu.
Evolutionists are drawn to the study of island biotas for their unique species (Otte, 1994; Rubinoff and Haines, 2005; Grant and Grant, 2008; Bowen et al., Chapter 1, this volume; Yoder et al., Chapter 12, this volume), as refuges of extant, “relict” organisms (Buckley et al., 2009; Wood et al., 2015), and most famously, for their displays of adaptive radiation (Losos and Ricklefs, 2009; Gillespie, 2015). Likewise, ecologists have long recognized the value of islands as microcosms of the processes of community assembly (MacArthur and Wilson, 1967; Warren et al., 2015). In recent years, there has been growing interest in combining these elements to study the evolution of community assembly, with particular focus on islands within (and beyond) the “radiation zone” (MacArthur and Wilson, 1967), where in situ speciation can be a major contributor to the origin of ecological communities (Heaney, 2007; Rominger et al., 2016). Under such circumstances, rules for community assembly can be illuminated by comparative phylogeographic approaches, revealing common evolutionary histories of codistributed, endemic taxa both within and between island archipelagos.
Remote island archipelagos offer relatively simple arenas for the evolutionary dynamics of community assembly because they are generally small in size and are often characterized by spatial isolation beyond the probable dispersal range of most organisms (Gillespie and Baldwin, 2009; Rosindell and Phillimore, 2011). As G. G. Simpson (1940) hypothesized, the probability of colonization should decline with increasing remoteness of an island from a mainland source pool, which he aptly named “sweepstakes” dispersal. Moreover, remote island archipelagos are veritable specks of land in a wide ocean world, again reducing the probability of colonization. Evidence of these hypothesized filters to colonization shows that remote islands frequently harbor disharmonious biotas (where there is an imbalance of taxonomic representation compared with mainland source pools). Moreover, the degree of disharmony increases with increasing distance from a probable source pool offering additional evidence of these spatial sieves and the potential for a relatively simpler community assembly (Gillespie and Roderick, 2002). Together with small size and extreme isolation, the assemblage of biotas may be further reduced in remote oceanic archipelagos because they are generally formed without life due to the nature of their geological origins (Gillespie et al., 2012). In contrast, species and community evolution in island systems in close proximity to continental species pools (e.g., continental “fragment” islands) (Gillespie and Roderick, 2002) can be extraordinarily complex (Steppan et al., 2003) in particular because they are often formed with a full complement of species and have more frequent connections with source pools.
Along with the effects of small size, spatial isolation, and dispersal origins on community assembly of remote islands, common phylogeographic
patterns may further enhance the “laboratory” for study of the evolution of community assembly. The evidence of disharmony, and potential for geographically diverse source pool origins (Gillespie and Roderick, 2002; Steppan et al., 2003; de Queiroz, 2005; Keeley and Funk, 2011; Gillespie et al., 2012), might lead us to expect somewhat haphazard phylogeographic patterns in taxa distributed across multiple remote archipelagos (Gillespie et al., 2012), for example, as appears to be the case for the spider genus Tetragnatha across Polynesia (Gillespie, 2002). However, once a lineage has initially established within a remote archipelago, it is largely cut off from its source population (qualified, of course, on the dispersal ability of the taxon). Thereafter, colonization among the constituent islands is considerably more predictable, and geological history at this more restricted geographic scale inspires hypotheses of common phylogeographic patterns among codistributed taxa (Funk and Wagner, 1995b; Avise, 2000).
Many remote archipelagos are volcanic in origin and sometimes exist in a linear age progression. These “hotspot” archipelagos are formed by molten lava rising from relatively fixed spots on the sea floor. Plate tectonic drift creates a conveyor belt motion that carries newly formed islands away from the hotspot location in a consistent direction at a steady rate, the mechanism that generates the linear age progression (Wilson, 1963; Clouard and Bonneville, 2005). These geological origins create a geographical and chronological context for hypotheses of phylogeographic congruence among codistributed taxa that track the ages of the islands, termed the “progression rule” (Figs. 4.1 and 4.2).
Common evolutionary histories of taxa, as well as deviations, can have a profound impact on our understanding of community assembly in the radiation zone. In this paper, we examine the progression rule to gain insight into the process of community assembly. In the following sections, we first present a brief overview of the progression rule from exemplar remote oceanic archipelagos. We then consider a simple question: can the progression rule be explained on the basis of island hot spot theory alone? We evaluate recent studies of progression from remote oceanic islands, highlighting two cases where contrasting phylogeographic histories are implicated within the same species radiations. The potential for distinct genetic histories held within the same demographic taxon, a phenomenon now widely recognized (Seehausen et al., 2014), illuminates the link between ecological community assembly and evolutionary assembly in isolated archipelagos.
THE PROGRESSION RULE IN COMPARATIVE ISLAND PHYLOGEOGRAPHY
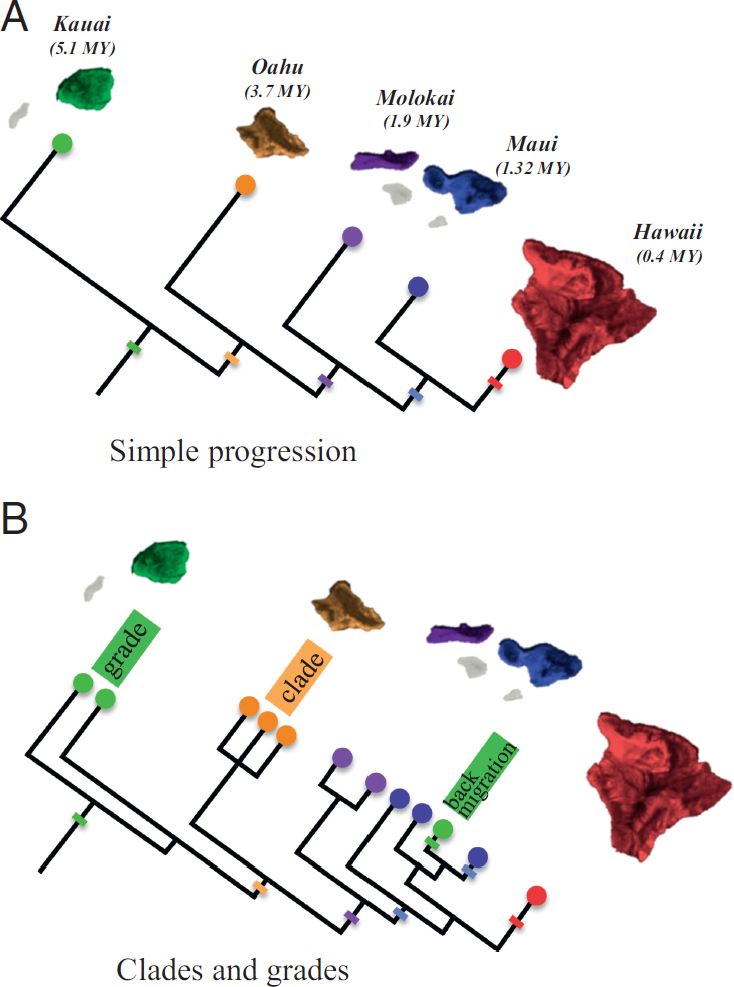
In the context of islands, a progression rule (or pattern) refers to a phenomenon of phylogeographic concordance with island age, whereby older lineages map to older islands within an archipelago, and younger lineages map to progressively younger islands in that system (Fig. 4.1A) (Funk and Wagner, 1995b). Progression is hypothesized to result from the
early colonization by a lineage when older islands existed but younger islands had not yet formed. Subsequent colonization occurs as new islands form. In his seminal work on phylogenetic theory, Hennig (1966) proposed the progression rule as a consequence of a general speciation mechanism, wherein species ranges consist of an ancestral, central portion within an older geographic area, and a derived, peripheral portion in a younger geographic area. Progression becomes evident in the phylogeny once speciation occurs; the ancestral, central portion splits from the derived, peripheral portion of the ancestral species. Thus, the ancestral lineage is concordant with older area, whereas the derived lineage is concordant with newer geological areas (Fig. 4.1A). Hennig’s (1966) model would not predict a progression to the extent of present-day discussions, unless the next new habitat that forms is spatially closer to the newest of the previous habitats.
Oceanic hotspot archipelagos offer, at a minimum, the starting conditions where progression could begin. Indeed, at least superficially, progression is not difficult to explain, as colonists of younger islands are most likely to come from spatially proximate, older islands. However, there is ample variation among taxonomic lineages in both the degree and form of progression (see below). In general, progression is more likely to evolve with (i) increasing spatial isolation of the archipelago, (ii) increasing spatial linearity of the islands, and (iii) enough dispersal to ensure colonization, such that when new habitat arises after the emergence of a new island, propagules are spatially poised to colonize from the next oldest habitat.
Variation in the starting conditions for progression may arise for a variety of reasons (we treat the persistence of progression below). For example, some archipelagos are less isolated from potential source areas (relative to the dispersal ability of the organisms) than others, resulting in repeated colonizations by the same lineages, producing conditions for either biotic turnover or anagenesis, rather than the within-archipelago cladogenesis associated with the progression pattern (Rosindell and Phillimore, 2011). Effectively, too much dispersal prevents the development of a progression pattern. In addition, although less likely, some organisms may be dispersal limited, and while habitat is available, they do not disperse to it predictably. In addition, some archipelagos do not show a linear spatial arrangement concordant with age, resulting in potential within archipelago colonizations from islands of mixed age. This nonlinear age arrangement would interfere with the older to younger colonization and ensuing phylogeographic pattern characteristic of progression. Likewise, we would not predict a progression for taxa arriving after the current major islands came into existence.
Progression can be complex; Funk and Wagner (1995a) discuss a variety of reasons for this complexity, providing a general framework
for interpreting more complex patterns. A strict progression pattern is the simplest, where all cladogenic events arise coincident with migration to newly arisen volcanoes along a linear chronological island sequence (Fig. 4.1A). Funk and Wagner (1995a) discuss this pattern with reference to Hawaiian endemics, where it is rare for taxa to conform exactly to this simple manifestation of the progression rule, although some taxa do illustrate the progression perfectly (Magnacca and Price, 2015).
Another more common pattern shows older to younger island colonization followed by some degree of within-island speciation (a pattern of progressive clades; Fig. 4.1B) (Funk and Wagner, 1995a). This progression pattern would arise when the interisland colonists from an older island migrate early in the history of the extant, older island clade. Progressive clades can give way to progressive grades (Fig. 4.1B), where a formerly monophyletic clade within an older island becomes paraphyletic when a subsequent colonization to a younger island occurs by a terminal taxon within that clade. In addition to these basic progression histories, instances of backmigration (migrations from younger to older islands) may also occur (Fig. 4.1B).
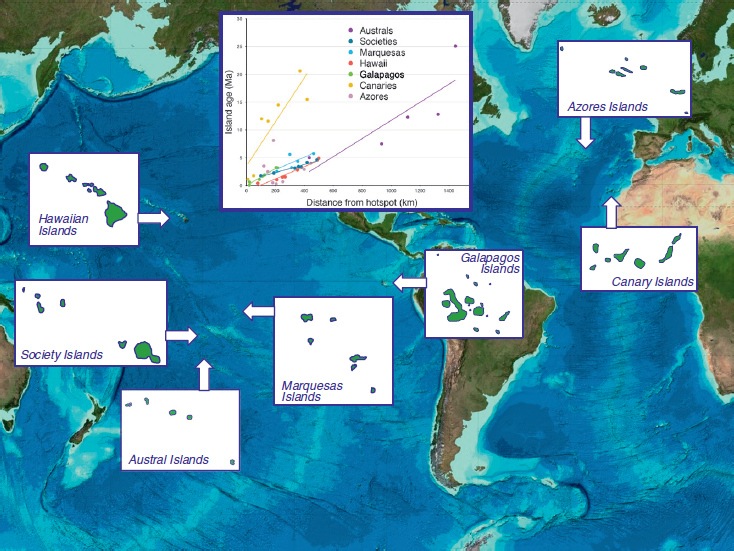
Armed with some understanding of the complexities of the progression pattern, we now examine progression in some well-recognized hotspot archipelagoes, with particular focus on the Pacific Ocean (Fig. 4.2).
Hawaiian Islands
Progression has been established most famously as a general pattern in the Hawaiian Archipelago (Fig. 4.2), where the formation of new, colonizable habitats does follow such a chronological sequence (Funk and Wagner, 1995b). Hawai’i’s indigenous biota is highly disharmonious, and chance colonization has undoubtedly played a large part in the initial establishment of the native diversity that we see today. However, within the Hawaiian Islands, a taxonomically broad expression of the progression rule is evident, including examples from plants [e.g., Hawaiian silverswords (Baldwin and Robichaux, 1995); Schiedea (Nepokroeff et al., 2003); Psychotria (Ree and Smith, 2008); and lobeliads (Givnish et al., 2009)], insects [e.g., Megalagrion damselflies (Jordan et al., 2003); Laupala crickets (Mendelson and Shaw, 2005); Banza katydids (Shapiro et al., 2006); Hyposmocoma moths (Haines et al., 2014); and picture-winged Drosophila (Bonacum et al., 2005; Magnacca and Price, 2015)], land snails [e.g., succineid species (Rundell et al., 2004)], spiders [e.g., Orsonwelles (Hormiga et al., 2003)], and birds (VanderWerf et al., 2010), among many others. Funk and Wagner (1995b) documented progression in 18 of 25 endemic lineages. More recent reviews (Roderick and Gillespie, 1998; Cowie and Holland, 2008) have further summarized this impressive phylogeographic result.
Austral Islands
The Austral Archipelago (Fig. 4.2) is considered geologically continuous with the Cook Islands (located to the northwest), which together were formed from repeated episodes of volcanism at several sites (Dickinson, 1998; Bonneville et al., 2002). Like the other Pacific hotspots, they are sequentially ordered from southeast to northwest by increasing age, although there has been secondary volcanic activity in the older islands, making them older than other Pacific hotspots (Clouard and Bonneville, 2005) (Fig. 4.2, Inset; and Table S11). Among independent lineages of spiders, in particular the crab spider Misumenops rapaensis (Garb and Gillespie, 2006), the orb web spider Tangaroa tahitiensis (Uloboridae) (Gillespie et al., 2008), and Rhyncogonus weevils (Claridge et al., 2016), a similar pattern of sequential colonization of islands is emerging, with large genetic distances between island populations. Thus, despite the modest extent and topography of the Austral islands and the widespread and generalist nature of their taxa, all studies to date show strong support for a progression rule in this archipelago.
Society Islands
Age progression within the Society Islands is in good agreement with the fixed hotspot hypothesis (Clouard and Bonneville, 2005), although the islands are considerably smaller in size than those of the Hawaiian chain (Fig. 4.2 and Table S1). However, there is considerable topographic diversity. Among insects, blackflies in the genus Simulium (Joy and Conn, 2001; Craig, 2003) show no evidence of a progression rule, which may be because of extinctions of habitat-specialized species on the older islands, due to loss of habitat. Likewise, among weevils in the genus Rhyncogonus, there appear to have been multiple independent colonizations of the island of Tahiti from neighboring island chains and no evidence of progression from older to younger islands (Claridge, 2006; Claridge et al., 2016). A similar conclusion results from phylogenetic analyses of Polynesian reed-warblers (genus Acrocephalus; Cibois et al., 2011). One reason that has been suggested is that there are ancient islands in close proximity to the Societies that may have served as a source of propagules (Gillespie et al., 2008).
__________________
1 Supporting information for this chapter, which includes Table S1, is available online at http://www.pnas.org/lookup/suppl/doi:10.1073/pnas.1601078113/-/DCSupplemental.
Marquesas Islands
The chronological arrangement of the Marquesas Islands is not strictly regular (Clouard and Bonneville, 2005). Nevertheless, among birds, an approximate progression from older to younger islands is found in the genus Pomarea (Cibois et al., 2004). Among spiders, the endemic taxa fall into a northern and southern lineage (Gillespie, 2003), consistent with the progression, although without strong phylogenetic support. A similar pattern of northern and southern lineages is found in Rhyncogonus weevils (Claridge, 2006) and partulid land snails of the genus Samoana (Johnson et al., 2000).
Galapagos Islands
The Galapagos Islands, although more geographically clustered than other Pacific hotspots (White et al., 1993), are still arranged chronologically (Fig. 4.2). This spatial arrangement implies a potential diversification sequence from southeast to northwest, paralleling the geological formation of the archipelago’s islands (Poulakakis et al., 2008). Some lineages in the Galapagos show a very clear progression from older to younger islands (Parent et al., 2008), well illustrated by the Galapagos giant tortoise (Geochelone nigra), in which both the species-level phylogeographic pattern based on mtDNA data and the pattern of lineage sorting suggest diversification in parallel with the island geological formation (Caccone et al., 2002; Beheregaray et al., 2004). One of the two lineages of Galapagos lava lizards (Microlophus) (Wright, 1983; Lopez et al., 1992) has also diversified in concert with the geological formation of island clusters of similar age (Kizirian et al., 2004). Among marine iguanas (Amblyrhynchus cristatus), mtDNA shows population differentiation concordant with geographical isolation of populations across the archipelago, a result largely in agreement with nuclear microsatellite data (Steinfartz et al., 2007). Among birds, Galapagos mocking birds also appear to follow the progression rule (Arbogast et al., 2006), although Darwin’s finches show limited evidence of diversification closely associated with the geological formation of the islands (Grant and Grant, 2008). A progression pattern has been inferred for Galapagos bulimulid land snails (Parent and Crespi, 2009). In contrast, insects including Galapagos flightless weevils (Galapaganus) (Sequeira et al., 2008) and the microlepidopteran genus Galagete (Schmitz et al., 2007) do not follow the progression rule. Evidently, many Galapagos terrestrial faunal groups follow the progression rule, the major exceptions coming from more vagile lineages (Galapagos finches, Galagete lepidopterans, and Galapaganus weevils) (Parent et al., 2008). Interestingly, in no situation is adaptive radiation associated with progression.
Canary Islands
Phylogeographically, the Canary Islands are the most thoroughly studied of all of the Atlantic island groups (Juan et al., 2000). In this archipelago, the islands farther from the mainland are younger, and those closer to the mainland (Fuerteventura and Lanzarote) are older (Fig. 4.2, Inset). Compared with the Pacific volcanoes, the archipelago is much older and moves much more slowly. Older islands closer to the continent are drier, as well as lower in elevation; thus, Fuerteventura and Lanzarote do not contain any wet forest habitats. The progression rule appears to be a common pattern of colonization shown by several groups of organisms (Juan et al., 2000; Sanmartín et al., 2008; Faria et al., 2016). However, species may, at least sometimes, also disperse from younger to older islands, even when these islands are occupied by close relatives (Juan et al., 2000). Among spiders, a progression rule has been documented for Loxosceles (Planas and Ribera, 2014) and Dysdera (Arnedo et al., 2001; Macías-Hernández et al., 2008), whereas Pholcus and Spermophorides probably colonized the older Fuerteventura and Lanzarote from the younger Gran Canaria (López-Mercader, 2005; Dimitrov et al., 2008). The mixed support for a progression across the Canary Islands presents the possibility of an alternative explantion for the origin of the endemics of the oldest islands, which could be the result of secondary replacement of its original fauna by new colonists better adapted to increasingly arid conditions (Cardoso et al., 2010). Older islands in the Canaries may show loss of old resident species due to aridification and orographic simplification; in some lineages, this may have been compensated by colonization and subsequent diversification of new, better-adapted organisms that could take advantage of empty niches and new opportunities.
Azores Islands
The Azores have also been used for examination of the role of island progression in dictating biogeographic patterns (Carvalho et al., 2015). However, the islands are located over a complex microplate rather than a single hotspot. Nevertheless, there is some evidence for a directional mode of dispersal from older to younger islands [plants (Díaz-Pérez et al., 2008; Rumeu et al., 2011), arthropods (Amorim et al., 2012; Parmakelis et al., 2015)], although a simple progression rule is not common [yet Van Riel (2005) presents intriguing evidence from gastropods]. Much of the current evidence consists of age estimates of haplotypes/alleles rather than robustly structured cladogenetic branching between islands. This pattern may be in part due to young radiations, since much of the landmass in the Azores is younger than 1 Ma. Moreover, the islands have suffered extensive habitat modification due to more recent volcanic activity
and deforestation of native forests over many centuries of occupation (Carvalho et al., 2015).
PERSISTENCE OF THE PROGRESSION PATTERN AND THE EVOLUTION OF COMMUNITY ASSEMBLY
As with most evolutionary processes, community assembly in the radiation zone happens more slowly than can be observed. Progression patterns on remote oceanic archipelagos are valuable in that they suggest a temporal frame of reference among extant communities as well as comparative insights into past events. For example, a progression pattern is typically interpreted as evidence of an historical path of colonization. A phylogenetic estimate that allies early branches with older islands and more distal branches with younger islands suggests that a lineage has been established within a given archipelago because the earliest islands were habitable by that taxon. Subsequent colonizations occur coincident with the emergence of younger islands, initiating new communities at progressively younger times. In archipelagos that meet conditions for progression, codistributed taxa with common phylogeographic histories therefore can be studied as part of their communities on specific islands in the sequence, representing slices of time in the history of the archipelago. Thus, a progression pattern provides snapshots of communities through time, for as long as the oldest islands have been supporting such communities.
The history of community assembly over evolutionary time also may be informed by the geographical polarity evident in the progression pattern of a lineage. In an archipelago with a linear spatial and chronological sequence, like those often present in hotspot regions, a simple progression pattern is coincident with a unidirectional path of colonization from older to younger islands. A possible explanation for this is that a regular supply of propagules on older islands maintains the constant potential for colonization as new habitat emerges with the origin of a new island. This dynamic suggests that once new habitat is available (on progressively younger islands), colonists from spatially proximate, older habitat have the potential to arrive, establish, and for many such radiations, diverge.
Biotic Resistance on an Evolutionary Timescale
Although a progression rule may be a valuable tool to understand the evolution of community assembly, there is also the possibility that community assembly may help us understand the development of the progression rule. One of the more prominent features of the progression pattern, and oceanic colonization pathways in general, is that they are largely unidirectional (Waters et al., 2013). In hotspot regions, the chronological
sequence of the archipelago landscape promotes a progression pattern because a probable source of colonists is available to establish the next stepping stone in the series once the next island habitat emerges. Thus, the initial progression arising from older to younger island colonizations seems relatively easy to explain.
On the other hand, dispersal vectors (wind, ocean currents. or biological vehicles) (Gillespie et al., 2012) are not expected to promote a unidirectional migration route (e.g., in Hawai’i, complex wind currents have the potential to bring propagules in a variety of directions, and tropical storm tracks, in particular, at times flow in a reverse direction to island age). Indeed, radiations that evidently postdate the Hawaiian Archipelago formation [e.g., the spider genus Havaika (Arnedo and Gillespie, 2006); the plant genus Tetramolopium (Lowrey, 1995); some Hawaiian birds (Fleischer and McIntosh, 2001)] show colonization routes at odds with the chronosequence, which suggests that there is no physical feature dictating an older to younger colonization route after the islands exist.
Although an early progression pattern arising from the initial colonization opportunities in hot spot archipelagos might often be expected, the unpredictable nature of dispersal might be expected to erase the pattern over evolutionary time. Drawing on ideas dating back to MacArthur and Wilson (1967), it has been argued that an apparent unidirectional pathway of colonization toward younger islands, and by extension the progression rule, may be bolstered by niche preemption, a type of priority effect (Gillespie and Roderick, 2002; Fukami et al., 2007; Gillespie and Baldwin, 2009; Waters et al., 2013; Fukami, 2015).
Priority effects, which arise from the impact that species have on one another within a community, depend on the order of arrival in the community (Fukami, 2015). By this process, once the first colonist (a founder) establishes in a newly available habitat, it soon monopolizes resources in the critical dimensions of the niche and blocks subsequent propagules from establishing. Key to the concept of priority effect is order of arrival. Although priority effects can be positive or negative (Fukami, 2015), this manifestation of the priority effect is negative, yielding advantage to the first colonist to arrive: once a niche has been filled, it is more difficult for ecologically similar individuals to enter (MacArthur and Wilson, 1967).
Priority effects would have an impact on phylogeographic patterns potentially in two ways. First, and most dramatically, patterns that suggest older to younger island colonization would unfold over evolutionary time as a hotspot archipelago forms in a spatiotemporal sequence. On each successively younger island, initial (early) colonists would arrive from the most proximate older island, establish in an empty habitat, and increase in population size and density, that is, preempting the relevant niche. At any point in the future, ecologically similar propagules from younger
islands backcolonizing to older islands would be second to arrive (late invaders), suffer a disadvantage, and fail to establish due to arriving in the preempted niche (Gillespie and Roderick, 2002). Thus, the ecological consequences of niche preemption by the early colonists is that backmigrations are discouraged from establishing. Second, niche preemption may further accentuate the initial older to younger island progression pattern by bolstering the genetic legacy of the early colonists via a “founder takes all” dynamic, whereby repeated older to younger island colonizations are likewise thwarted, again due to niche preemption (Waters et al., 2013). Both of these scenarios suggest that ecological dynamics and species interactions are important in giving rise to progression. In other words, to explain features of a progression rule pattern, particularly for a species radiation where all available habitat has been colonized (suggesting propagules are not rare), but speciation has occurred in concert with interisland colonization (suggesting that gene flow has terminated), priority effects may be critical.
Moreover, the ability of new colonists to enter a community is frequently thought to decline as the diversity of species already in the community increases; the established species are considered to dictate biotic resistance against further colonization or invasion (Chapin et al., 2000; Kennedy et al., 2002; Hooper et al., 2005). Thus, biotic resistance results from a complementarity effect; the larger the number of species in a community, the more resource used, in turn creating more community resistance to new invaders. Diversity can also generate biotic resistance through a selection effect under which the most effective competitors, such as those most similar to focal invaders, are more likely to be present in more diverse mixes (Chapin et al., 2000; Loreau and Hector, 2001). Priority effects can play a synergistic role in each of these dynamics. Curiously, although the consequences of priority effects are well understood over evolutionary timescales, the opposite appears to be true at ecological timescales.
Biotic Turnover on an Ecological Timescale
MacArthur and Wilson (1967) proposed that species richness on islands is the result of a dynamic balance between stochastic immigrations and extinctions. Their thinking was a radical shift from most community assembly theory with rejection of a deterministic equilibrium driven by species interactions. Today, the proximity to the mainland, the size of the mainland species pool, and the area of the island are hypothesized to be the three main factors explaining species richness on islands. The equilibrium theory of island biogeography assumes a dynamic equilibrium with continual species turnover, because of coloni-
zation and extinction events that constantly modify community composition, a proposition subsequently developed in metapopulation ecology (Hanski, 1999). Thus, biotic turnover prevents genetic differentiation from accumulating, and thereby continually erases the development of a progression pattern.
How Do We Reconcile the Concepts of Biotic Turnover with Biotic Resistance?
Based on many (perhaps most) models of ecological community assembly, biotic resistance seems unlikely. So why, then, is biotic resistance so apparent on an evolutionary timescale (Strauss et al., 2006)? Put another way, how does ecological turnover (and associated ecological processes) give way to the biotic resistance and community “lockup” dynamics that might sustain a progression pattern over evolutionary timescales?
Possible hypotheses to explain community lockup dynamics that maintain progression patterns might be as follows. (i) Species and/or communities might be impervious to subsequent invasion due to “high-density blocking” (Waters et al., 2013). This hypothesis posits that, although additional colonizations occur (e.g., through backmigration), numerical dominance of the early, resident species deters establishment or dilutes the genetic contributions of late colonists with similar niche requirements. A prediction of this hypothesis is that the more similar late colonists are to resident species, the less likely they are to successfully invade. A genetic manifestation of this idea is that the distinct alleles of late invaders that manage to hybridize (admix) with residents have a high likelihood of extinction, thereby obscuring repeated- and backcolonization events. (ii) Late invaders suffer a mating disadvantage, or mate with residents but have no genetic impact, because they either fail to reproduce or their offspring have low fitness. (iii.a) Early invaders have adapted to conditions specific to each island, so that late invaders are always inferior competitors, or (iii.b) early colonizers undergo ecological release, resulting in extensive niche overlap among members of the early community; over time, and as the environment becomes more stable, they may specialize, placing subsequent colonizers at a disadvantage. Alternatively, (iv) after some initial turnover, the combined island communities lock in at a state where “by chance” no island can be invaded. Although it is difficult to separate these possible explanations (and they may not be mutually exclusive), genomic data promise an opportunity at least to test whether priority effects are real, and if so, what consequences would arise for genetic admixture and introgression.
THE PROGRESSION RULE AND THE MOSAIC GENOME
To address the lockup hypothesis will require a multidisciplinary approach. However, the first explanation suggested above, that late invaders hybridize with species in residence and their genes admix into the resident population, falls in the domain of comparative phylogeography. This hypothesis is predicated on the idea of a mosaic genome where specific regions can have separate historical identities (Baum and Shaw, 1995; Maddison, 1995; Shaw, 2001; Wu, 2001; Seehausen et al., 2014), some reflecting a history of progression, and others reflecting additional demographic complexities (such as backmigrations). Teasing apart complex histories within the same species lineage requires separate analyses of distinct hereditary units. Whether sufficient data exist to mark these separate histories is an empirical issue, but to be sure, if we combine data across data partitions that represent distinctive histories, we will lose insights into these complexities.
In a recent phylogeographic context, it is possible for different gene partitions to reveal conflicting but true histories that manifest more than one of these progression patterns within the same lineage. Such situations are extremely valuable because they suggest a multidimensional insight into past demography of the lineage.
A potential example comes from the Hawaiian cricket genus Laupala, a morphologically cryptic group of 38 flightless, single-island endemic, species (Otte, 1994). Multiple datasets reveal a compelling case for progressive clades and grades in Laupala, the most resolved of which emerges from a nuclear (presumably) phylogeny based on amplified fragment length polymorphisms (AFLPs; Mendelson and Shaw, 2005) (Fig. 4.3A). The phylogeny is well supported, is concordant with the species taxonomy, and is consistent with a less-resolved nuclear sequence phylogeny (Shaw, 2002). The area cladogram shows that the group began on Kaua‘i (or an older island) and split into two species groups once on the younger O‘ahu. Subsequent diversification occurred via fine-scale progression to increasingly younger islands, colonizing first either Moloka‘i (absent in one group) or West Maui, then East Maui, and, last, Hawai’i Island. Thus, the nuclear phylogeny suggests a unidirectional pattern with rare interisland migrations.
However, the unidirectional pattern of colonization is contradicted by the mtDNA phylogeny (Fig. 4.3B). Although there is a general pattern of progression in the mtDNA area cladogram as well, there is extensive conflict between an mtDNA tree and both the nuclear DNA trees (Shaw, 2002). Importantly, the phylogeographic patterns in the mtDNA include two potential backmigrations between Hawai’i and Maui and one additional backmigration between Maui and Moloka‘i. These backmigrations are between neighboring islands, a trend shared by other taxa where back-

migrations occur over a pattern of progression (Funk and Wagner, 1995a). In short, the mtDNA evidence suggests that there has been considerably more interisland movement than is revealed by the nuclear partitions.
A recent study of the flightless, species-rich weevil Laparocerus (Faria et al., 2016) from the Canary and Madeira Islands comes to a similar conclusion. The mtDNA and nuclear gene trees sampled from Laparocerus show conflicting topologies, similar to Laupala, along with a progression from older to younger islands evident in both data partitions. Like Laupala, the nuclear phylogeographic history is simpler, whereas the mtDNA phylogeographic history captures considerably more interisland movement. Although it is unclear in the case of Laupala, the nuclear data show evidence of subsequent admixture in cases of multiple colonization in Laparocerus. Importantly, in both systems, the patterns of mtDNA variation do not conform to expectations of incomplete lineage sorting, and instead appear to be informative about interisland colonization patterns.
Recent phylogeographic studies of Hawaiian planthoppers and spiders using next-generation sequencing approaches are starting to reveal similar patterns of higher-than-expected movement, at least between the younger islands. Together, the data suggest that there is some movement between islands, at least among younger islands (Roderick et al., 2012). The promise of these new genetic data technologies is that the heterogeneity of progression patterns can be fully investigated, and potentially lead to an understanding of the causes of such heterogeneous genetic signatures.
DISCUSSION
The progression rule is one of the most pervasive phylogeographic patterns yet documented, at least for the well-studied remote archipelago of Hawai’i. There is considerable evidence accumulating from two other well-studied archipelagos, the Canary Islands (Juan et al., 2000) and the Galapagos (Parent et al., 2008; Faria et al., 2016), despite the complicating factors of spatial nonlinearity and greater proximity to continents. Additional evidence is accumulating for progression in other remote archipelagos as well (summarized here for the Australs and the Marquesas) (Cibois et al., 2011). Evidence for progression, thus far, is mixed in the Azores, presumably due to the geological complexity of the hotspot (Carvalho et al., 2015), and the Society Islands, perhaps due to the close proximity to much older islands, meaning that there were other islands that served as a source of colonists, or a “bridge” between other archipelagos to the younger islands of the Society chain.
Progression is a feature that emerges over evolutionary time and can only develop once a community transitions from biotic turnover (mani-
fest on an ecological timescale) to biotic resistance (on an evolutionary timescale). To a large degree, the transition from biotic turnover to biotic resistance mirrors the transition from population genetic cohesion to the genetic differentiation that characterizes speciation. In species formation, eventually, a nascent species closes genetic connections with its past (represented by its sister species) and persists as an independent evolutionary entity. Likewise, in the transition from ecological turnover to biotic resistance, a new community forms. Whereas formerly, the community was in a dynamic state of revision due to immigration and extinction (under some theories), eventually, the community resists repeated immigration of colonists. The speciation process occurs within the context of this community transition, and embodies the zone of radiation.
The geographic and chronological settings of remote, hotspot archipelagos offer conditions under which progression patterns can arise. However, the ecological features of the organisms involved play an important role in the subsequent patterns of diversification, and the rate at which they evolve. Key are the rates of dispersal and likelihood of colonization and establishment (Buckley et al., 2013). Obviously, if little to no dispersal occurred, a progression pattern would be unlikely to arise as the species would likely remain in the ancestral habitat and not radiate (e.g., see potential examples reviewed in Keeley and Funk, 2011). Likewise, if dispersal and subsequent gene flow facilitated panmixia, neither speciation nor progression would be expected to arise because biotic turnover would prevail. However, if dispersal were infrequent but predictable (the likely condition for the majority of taxa), colonization would occur as new island habitats become available. Gene flow would be reduced, and depending on its magnitude, would not continue to homogenize gene frequencies between founder and source.
Once the founder population, now the resident species, established and has filled the available niche, priority effects could come into play by a number of possible mechanisms: (i) The community might be resistant to subsequent invasion due to the monopolization of resources by the numerically dominant resident species; the more similar late colonists are to resident species, the less likely they are to successfully invade. (ii) Late colonists may suffer incompatible mating encounters with the numerically dominant resident species, leading to a failure to reproduce. (iii) The resident species may have adapted to local conditions, giving it a competitive edge over later colonists. The last mechanism involves natural selection and competitive exclusion, whereas numerical dominance drives the first, and may contribute to the second interaction. These possible mechanisms could, in theory, be tested with appropriate experiments.
Depending on the degree of differentiation, the late colonist may nonetheless hybridize with the resident species and its genes may diffuse
into the resident species gene pool, leaving some trace of its attempt at establishment. As discussed in the two examples described earlier, mosaic histories contained within the genome of a given lineage may harbor evidence regarding how a progression pattern evolves in this intermediate dispersal domain. It seems unlikely that species radiations manifesting a progression have occurred in the clean manner that a phylogeny might suggest. Even in the most straightforward cases, such as the patterns of progressive clades and grades in the AFLP phylogeographic pattern in Laupala crickets, additional data cautions us from concluding that interisland migrations do not occur. More efforts are needed to examine contrasting evidence of progression patterns as they may provide insights into how the pattern develops. If we can understand the development of progression, we may begin to understand how ecological turnover (where progression would not arise) gives way to biotic resistance (where progression could arise).
As elucidated by Fukami (2015), priority effects by niche preemption are most likely to occur when two species show highly overlapping resource use, the first colonist has a high impact on the overlapping niche dimensions, and when the growth rate of the late colonist is heavily dependent on the environment. Thus, we might expect that ecological shifts create opportunities for late colonists to escape the impact of priority effects as a result of exploring new niche dimensions. Highly dynamic adaptive radiations that appear to violate the progression rule via backmigration, such as some branches of the Hawaiian picture-winged Drosophila radiation (Magnacca and Price, 2015), provide an opportunity to test this idea. Perhaps herein lies the explanation for the finding that adaptive radiations in the Galapagos were not found to adhere to the progression rule (Parent et al., 2008). Likewise, species that have more plastic attributes to their resource use might have a heightened immunity to priority effects, which would act to discourage the development of a progression pattern in the lineage. Better characterization of ecological traits, reproductive behaviors, divergence times, and genetic admixture among lineages of island radiations should allow for more rigorous evaluation of priority effects on the development of progression within a phylogenetic context.
The study of community assembly in and beyond the radiation zone is exciting in part because it brings together two fairly disparate disciplines: the ecological study of community structure and the evolutionary study of the origin of species. The presence of progression, and its more nuanced manifestation revealed by the mosaic genome, is fortunate for the study of community assembly because the pattern provides a temporal framework for both ecological and evolutionary studies of communities and their interrelationship. The key importance of the progression pattern is that multiple lineages are establishing and assembling, interacting and adapt-
ing, over a similar time frame that plays out over extended evolutionary time. Thus, we can measure ecological metrics at different time slices of the community assembly process to find how properties (species diversity, abundance, body size distributions, trophic interactions) change over time (Loreau et al., 2002; Harmon et al., 2009), and how the origin of new species affects these properties. Moreover, as genomic data become available across multiple lineages that appear to follow a progression in a given system (Gillespie, 2015), we are gaining insight into how taxa differ in mode, rates, and patterns of establishment and diversification (Rominger et al., 2016). Integrating multidimensional datasets across stages of the progression will allow us to understand how interactions develop and evolve and the importance of such interactions in dictating properties of stability, turnover, and the evolution of biotic resistance in a community. Effectively, with multiple lineages being formed over the same time frame, we can examine the feedback between ecology and evolution and hence generate insights into the processes involved in the formation and loss of biodiversity.
ACKNOWLEDGMENTS
We thank Francisco Ayala, John Avise, and Brian Bowen for the invitation to participate in the symposium and special issue. We thank two reviewers for comments that helped improve the manuscript. We also thank our students and colleagues of the Dimensions of Hawaiian Biodiversity team for many stimulating discussions. This work was supported by National Science Foundation (NSF) Division of Environmental Biology Grants 1241060 (to K.L.S.) and 1241253 (to R.G.G.).




















