
12
Choline
SUMMARY
Choline functions as a precursor for acetylcholine, phospholipids, and the methyl donor betaine. The primary criterion used to estimate the Adequate Intake (AI) for choline is the prevention of liver damage as assessed by measuring serum alanine aminotransferase levels. The AI for adults is 550 mg/day of choline for men and 425 mg/day for women. There are no nationally representative estimates of the intake of choline from food or food supplements. Choline in the diet is available as free choline or is bound as esters such as phosphocholine, glycerophosphocholine, sphingomyelin, or phosphatidylcholine. The critical adverse effect from high intake of choline is hypotension, with corroborative evidence on cholinergic side effects (e.g., sweating and diarrhea) and fishy body odor. The Tolerable Upper Intake Level (UL) for adults is 3.5 g/day.
BACKGROUND INFORMATION
Choline is a dietary component that is important for the structural integrity of cell membranes, methyl metabolism, cholinergic neurotransmission, transmembrane signaling, and lipid and cholesterol transport and metabolism. Human cells grown in culture have an absolute requirement for choline (Eagle, 1955). When cells are deprived of choline, they die by apoptosis (Albright et al., 1996; Cui et al., 1996; Holmes-McNary et al., 1997; James et al., 1997; Shin et
al., 1997; Zeisel et al., 1997). There is an endogenous pathway for the de novo biosynthesis of the choline moiety via the sequential methylation of phosphatidylethanolamine using S-adenosylmethionine as the methyl donor (Bremer and Greenberg, 1961) (see Figure 12-1). Thus, the demand for dietary choline is modified by metabolic methyl-exchange relationships between choline and three nutrients: methionine, folate, and vitamin B12 (lipotropes) (Zeisel and Blusztajn, 1994).
With this type of nutrient interdependence, designation of the essential nature of a nutrient depends on showing that de novo synthesis rates are not adequate to meet the demand for the nutrient when the other nutrients are available in amounts sufficient to sustain normal growth and function. Healthy men with normal folate and vitamin B12 status fed a choline-deficient diet have diminished plasma choline and phosphatidylcholine concentrations and develop liver damage (Zeisel et al., 1991). For these individuals, de novo synthesis of choline was not adequate to meet the demand for
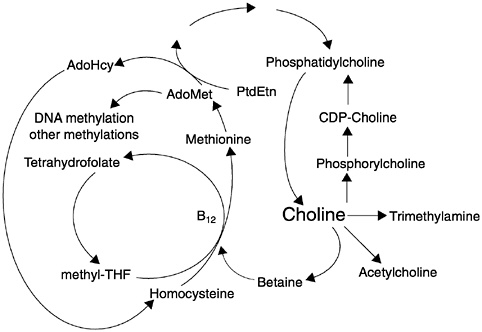
FIGURE 12-1 Choline, folate, and methionine metabolism are closely interrelated. AdoHcy = S-adenosylhomocysteine, AdoMet = S-adenosylmethionine, B12 = vitamin B12, CDP-Choline = cytidine diphosphocholine, PtdEtn = phosphatidylethanolamine, THF = tetrahydrofolate. Reprinted with permission, from Zeisel and Blusztajn (1994). Copyright 1994 by Annual Reviews.
the nutrient. Information about women, infants, children, and older adults is not sufficient to know whether choline is needed in the diet of these groups.
Function
Choline can be acetylated, phosphorylated, oxidized, or hydrolyzed. Several comprehensive reviews of the metabolism and functions of choline have been published (Kuksis and Mookerjea, 1978; Zeisel, 1981; Zeisel and Blusztajn, 1994).
Choline accelerates the synthesis and release of acetylcholine, an important neurotransmitter involved in memory storage, muscle control, and many other functions (Cohen and Wurtman, 1975; Haubrich et al., 1974; Wecker, 1986). It is also a precursor for the synthesis of (1) phospholipids, including phosphatidylcholine (a membrane constituent important for the structure and function of membranes), for intracellular signaling (Exton, 1994; Zeisel, 1993) and hepatic export of very low-density lipoproteins (Yao and Vance, 1988, 1989); (2) sphingomyelin (another membrane constituent) for structural and signaling functions (Hannun, 1994); and (3) platelet activating factor, a potent messenger molecule (Frenkel et al., 1996). Choline is a precursor for the formation of the methyl donor betaine. Betaine is also required by renal glomerular cells, which use betaine and glycerophosphocholine as organic osmolytes to adapt to osmotic stress (Bauernschmitt and Kinne, 1993; Burg, 1995; Garcia-Perez and Burg, 1991; Grossman and Hebert, 1989).
Physiology of Absorption, Metabolism, and Excretion
Dietary choline is absorbed from the lumen of the small intestine via transporter proteins in the enterocyte (Herzberg and Lerner, 1973; Herzberg et al., 1971; Kuczler et al., 1977; Sheard and Zeisel, 1986). Before choline can be absorbed from the gut, some is metabolized by bacteria to form betaine (which may be absorbed and used as a methyl donor) and methylamines (which are not methyl donors) (Zeisel et al., 1983). No other component of the diet has been identified as competing with choline for transport by intestinal carriers. Choline is found in foods as free choline and as esterified forms such as phosphocholine, glycerophosphocholine, sphingomyelin, and phosphatidylcholine. Lecithin is a phosphatidylcholine-rich fraction prepared during commercial purification of phospholipids, and this term is often used interchangeably with phosphatidylcholine. Lecithin is often added to foods as an emulsifying agent.
Pancreatic enzymes can liberate choline from dietary phosphatidylcholine, phosphocholine, and glycerophosphocholine (Zeisel and Blusztajn, 1994). The free choline that is formed enters the portal circulation of the liver (Le Kim and Betzing, 1976) whereas phosphatidylcholine may enter via lymph in chylomicrons.
All tissues accumulate choline by diffusion and mediated transport (Zeisel, 1981). A specific carrier mechanism transports free choline across the blood-brain barrier at a rate that is proportional to the serum choline concentration. In the neonate this choline transporter has an especially high capacity (Cornford and Cornford, 1986). The rate at which the liver takes up choline is sufficient to explain the rapid disappearance of choline injected systemically (Zeisel et al., 1980c). The kidney also accumulates choline (Acara and Rennick, 1973). Some of this choline appears in the urine unchanged but most is oxidized within the kidney to form betaine (Rennick et al., 1977).
In the predominant pathway for phosphatidylcholine biosynthesis, choline is phosphorylated, converted to cytidine diphosphocholine, and then converted to phosphatidylcholine (Kennedy and Weiss, 1956; Vance, 1990) (Figure 12-1). In an alternative pathway, phosphatidylethanolamine is sequentially methylated to form phosphatidylcholine by the enzyme phosphatidylethanolamine-N-methyltransferase with S-adenosylmethionine as the methyl donor (Bremer and Greenberg, 1961; Vance and Ridgway, 1988). This is the major (perhaps only) pathway for de novo synthesis of the choline moiety in adult mammals. It is most active in the liver but has been identified in many other tissues (Blusztajn et al., 1979; Crews et al., 1981; Yang et al., 1988). Best estimates of in vivo activity of this enzyme, based on in vitro data, are that 15 to 40 percent of the phosphatidylcholine present in the liver is derived from the phosphatidylethanolamine-N-methyltransferase pathway, with the remainder coming from the cytidine diphosphocholine pathway (Bjornstad and Bremer, 1966; Sundler and Akesson, 1975). No estimates are available as to the relative extent of choline obtained from cell turnover. Dietary intake of phosphatidylcholine is approximately 6 to 10 g/day (Zeisel et al., 1991).
A significant portion of choline is oxidized to form betaine in the liver and kidney (Bianchi and Azzone, 1964; Weinhold and Sanders, 1973). The methyl groups of betaine can be scavenged and reused in single-carbon metabolism (Finkelstein et al., 1982) (see “Nutrient-Nutrient Interactions”).
Clinical Effects of Inadequate Intake
Humans
Although choline is clearly essential to life, there is only one published study examining the effects of inadequate dietary intake in healthy men. That study reported decreased choline stores and liver damage (elevated alanine aminotransferase) when men were fed a choline-deficient diet containing adequate methionine, folate, and vitamin B12 for 3 weeks (Zeisel et al., 1991) (Figures 12-2 and 12-3). Another study, in which men were fed a choline- and methyl-deficient diet, reported decreased choline stores but did not report on liver function (Jacob et al., 1995). Individuals fed with total parenteral nutrition (TPN) solutions devoid of choline but adequate for methionine and folate develop fatty liver and liver damage as assessed by elevated alanine aminotransferase; in some individu
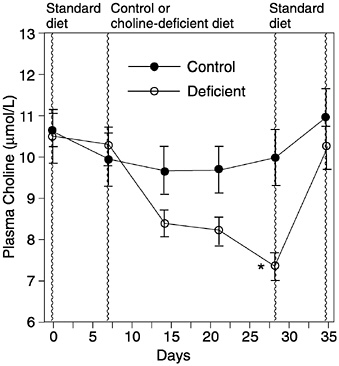
FIGURE 12-2 Plasma choline in healthy men ingesting a control (500 mg/day of choline) or choline-deficient (13 mg/day of choline) diet. *Difference from day 7 value: p < 0.01. Reprinted with permission, from Zeisel et al. (1991). Copyright 1991 by the Federation of American Societies for Experimental Biology.
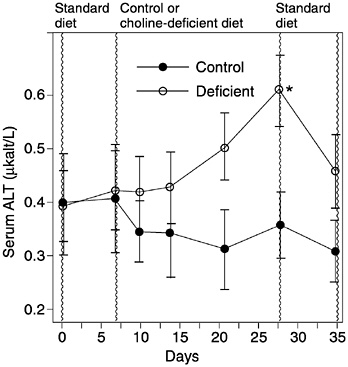
FIGURE 12-3 Serum alanine aminotransferase (ALT) activity in men ingesting a control or choline-deficient diet. Serum ALT was determined by using an automated spectrophotometric assay. Data are expressed as mean activity ± standard error of the mean. *Difference from day 7 value: p < 0.05. Reprinted with permission, from Zeisel et al. (1991). Copyright 1991 by the Federation of American Societies for Experimental Biology.
als, this is resolved when a source of dietary choline is provided (Buchman et al., 1992, 1993, 1995; Chawla et al., 1989; Shapira et al., 1986; Sheard et al., 1986). In a double-blind protocol, investigators administered lecithin (30 percent phosphatidylcholine) orally to patients receiving TPN twice daily for 6 weeks. At the end of this time, plasma choline had risen by more than 50 percent in the lecithin group whereas in the placebo group it had decreased by 25 percent. In the treated group, liver fat decreased by 30 percent (Buchman et al., 1992). In another small clinical study (Buchman et al., 1995), four patients who had low plasma concentrations of free choline after treatment with TPN (which contained no additional choline) were given 1 to 4 g/day of choline chloride for 6 weeks. During choline administration, plasma choline concentration
increased into the normal range but decreased back to baseline when choline supplementation was discontinued. Fatty liver was resolved completely during choline supplementation but steatosis (fatty liver) recurred in one patient after 10 weeks of return to choline-free TPN. The available data support the provisional conclusion that de novo synthesis of choline is not always sufficient to meet human requirements for choline.
Animals
Supporting animal studies (in many species, such as the baboon) also found that a choline-deficient diet resulted in decreased choline stores and liver dysfunction (Hoffbauer and Zaki, 1965; Sheard et al., 1986; Tayek et al., 1990; Yao and Vance, 1990). The following animals fed a choline-deficient diet may be susceptible to developing growth retardation, renal dysfunction and hemorrhage, or bone abnormalities: baboon (Hoffbauer and Zaki, 1965), chicken (Blair et al., 1973; Ketola and Nesheim, 1974), dog (Best and Huntsman, 1932; Hershey, 1931), guinea pig (Tani et al., 1967), hamster (Handler, 1949), pig (Blair and Newsome, 1985; Fairbanks and Krider, 1945), quail (Ketola and Young, 1973), rat (Newberne and Rogers, 1986), and trout (Ketola, 1976).
SELECTION OF INDICATORS FOR ESTIMATING THE REQUIREMENT FOR CHOLINE
Markers of Liver Dysfunction
The liver is damaged when humans consume an otherwise adequate diet that is deficient in choline, resulting in elevated alanine aminotransferase levels in blood (Burt et al., 1980; Tayek et al., 1990; Zeisel et al., 1991). Fatty infiltration of liver also occurs in choline deficiency but is difficult to use as a functional marker without special liver imaging techniques (Buchman et al., 1992).
Hepatic choline and choline metabolite concentrations have been shown to decrease during choline deficiency in the rat (Zeisel et al., 1989). Phosphocholine concentration in liver is highly correlated with dietary choline intake, decreasing to 10 to 20 percent of control values after 2 weeks on a diet sufficient in methionine, folate, and vitamin B12 but deficient in choline (Pomfret et al., 1990). Hepatic phosphocholine concentration was most sensitive to modest dietary choline deficiency, decreasing to 10 to 20 percent of control values after 2 weeks of a deficient diet (Pomfret et al., 1990). This
measurement is not easily undertaken in humans, although magnetic resonance spectroscopy does makes it possible (Cohen et al., 1995).
Plasma Concentrations
Plasma choline concentration varies in response to diet and is found in the water-soluble fraction as free choline (Buchman et al., 1993; Burt et al., 1980; Chawla et al., 1989; Sheard et al., 1986; Zeisel et al., 1991). It decreases approximately 30 percent in subjects fed a choline-deficient diet for 3 weeks (Zeisel et al., 1991). Plasma choline concentration can increase twofold after a meal high in choline content and three- or fourfold after a supplemental choline dose (Zeisel et al., 1980b). Fasting plasma choline concentrations vary from 7 to 20 µmol/L, with most subjects having concentrations of 10 µmol/L. The disadvantage of using plasma choline as a functional indicator is that these concentrations do not appear to decline below approximately 50 percent of normal, even when subjects fast for more than 1 week (Savendahl et al., 1997). Perhaps this is because membrane phospholipids, which are a large storage pool for choline, are hydrolyzed to maintain plasma choline concentration above this minimal level. Fasting plasma phosphatidylcholine concentrations (mostly as part of plasma lipoproteins) are approximately 1 to 1.5 mmol/L (Aquilonius et al., 1975; Zeisel et al., 1980b, 1991). Plasma phosphatidylcholine concentration also decreases in choline deficiency (Zeisel et al., 1991) but is also influenced by factors that change plasma lipoprotein levels.
Reduction of Risk of Chronic Disease
Dementia
Studies in rodents suggest that dietary intake of choline early in life can diminish the severity of memory deficits in aged animals (Bartus et al., 1980; Meek and Williams, 1997a, b, c). Most available human studies have used choline-containing compounds to treat rather than prevent the symptoms of dementia and therefore did not address whether dementias could be prevented. In the absence of food composition data, epidemiological studies on the association of choline intake with dementia are not available. More human studies are needed to determine whether dietary choline intake is useful in the prevention of dementia.
Cardiovascular Disease
The choline-containing phospholipid phosphatidylcholine (lecithin) has been used as a treatment to lower cholesterol concentrations because lecithin-cholesterol acyltransferase has an important role in the removal of cholesterol from tissues. In humans phosphatidylcholine ingestion is associated with a modest reduction in plasma cholesterol (Hirsch et al., 1978; Wood and Allison, 1982; Zeisel et al., 1991). In addition, choline or betaine treatment has been used to lower high plasma homocysteine concentrations (Anonymous, 1997; Dudman et al., 1987; Wendel and Bremer, 1984; Wilcken et al., 1983, 1985), and choline-deficient rodents have elevated plasma homocysteine concentrations (Varela-Moreiras et al., 1995) (see Chapter 8, “Vascular Disease”). Wendel and Bremer (1984) reported that betaine treatment was more effective than folate treatment in normalizing plasma homocysteine and methionine concentrations of a child with homocystinuria, a genetic disease caused by 5,10-methylenetetrahydrofolate reductase deficiency (choline is the precursor for betaine, which itself is found in sugar beets and wine). Therefore, dietary choline intake might be correlated with cardiovascular disease risk. More human studies are needed before conclusions can be drawn about whether dietary choline intake is useful in preventing cardiovascular disease.
Cancer
In rodents dietary choline deficiency is associated with increased incidence of liver cancer and increased sensitivity to carcinogenic chemicals (Newberne and Rogers, 1986). The mechanisms of the carcinogenic actions of choline deficiency are not known but may be mediated by changes in protein kinase C activity (da Costa et al., 1993, 1995). There are no human data; studies in humans are needed to assess the role of dietary choline in the prevention of cancer.
FACTORS AFFECTING THE CHOLINE REQUIREMENT
Nutrient-Nutrient Interactions
Any consideration of the requirements for choline and methionine needs to include the close interrelationships with other methyl donors. Choline, methionine, and folate metabolism interact at the point that homocysteine is converted to methionine.
Betaine-homocysteine methyltransferase catalyzes the methylation of homocysteine using betaine as the methyl donor (see Figure 12-1) (Finkelstein et al. 1982; Mudd and Poole, 1975; Wong and Thompson, 1972). In an alternative pathway, 5-methyltetrahydrofolate-homocysteine methyltransferase regenerates methionine by using a methyl group derived de novo from the single-carbon pool (Finkelstein et al., 1982, 1988). Methionine adenosyltransferase converts methionine to S-adenosylmethionine (the active methylating agent for many enzymatic methylations, including the methylation of phosphatidylethanolamine to form phosphatidylcholine [Ridgway and Vance, 1988]).
Perturbing the metabolism of one of the methyl donors reveals the intermingling of these metabolic pathways. Total hepatic folate content decreased by 31 to 40 percent in rats after 2 weeks on a choline-deficient diet (Selhub et al., 1991; Varela-Moreiras et al., 1995). This effect was reversed by refeeding choline (Varela-Moreiras et al., 1995). Rats fed diets deficient in both methionine and choline for 5 weeks had hepatic folate concentrations that were half of those present in controls (Home et al., 1989). Tetrahydrofolate deficiency in rats, induced by treatment with methotrexate (Barak and Kemmy, 1982; Barak et al., 1984; Freeman-Narrod et al., 1977; Pomfret et al., 1990; Svardal et al., 1988) or by dietary folate deficiency (Kim et al., 1994) resulted in diminished hepatic total choline, with the greatest decrease occurring in hepatic phosphocholine concentrations. During choline deficiency in rats, hepatic S-adenosylmethionine concentrations also decreased by as much as 50 percent (Barak et al., 1982; Poirier et al., 1977; Shivapurkar and Poirier, 1983; Zeisel et al., 1989). In rats choline deficiency for 2 weeks doubled plasma homocysteine levels (Varela-Moreiras et al., 1995). See Chapters 7 and 8 for more information on plasma homocysteine.
Gender
Males may have a higher choline requirement than do females. Female rats are less sensitive to choline deficiency than are male rats (Tessitore et al., 1995), perhaps because of females’ enhanced capacity to form the choline moiety de novo. Females rats have greater phosphatidylethanolamine-N-methyltransferase activity in liver than do males (Arvidson, 1968; Bjornstad and Bremer, 1966; Lyman et al., 1971). Estimates of the amount of increased activity vary between 10 (Lyman et al., 1971) and 50 percent (Bjornstad and Bremer, 1966). A woman’s capacity to form the choline moiety
de novo may decrease after menopause (Lindblad and Schersten, 1976), because estrogens increase hepatic phosphatidylethanolamine-N-methyltransferase activity in rats (Drouva et al., 1986; Young, 1971).
Exercise
Strenuous physical activity in trained athletes reduced the plasma choline concentration by approximately 40 percent, from 14.1 to 8.4 µmol/L (Conlay et al., 1986). A choline supplement given to marathon runners modestly enhanced performance (Sandage et al., 1992). In 10 top-level triathletes who were given either a placebo or lecithin at 0.2 g/kg body mass 1 hour before each type of exercise, plasma choline concentrations in all the triathletes decreased on average by 16.9 percent after the bicycle exercise when placebo was taken before the race but did not do so when lecithin was given (Von Allworden et al., 1993).
Bioavailability
No estimates are available for percentage absorption of the various forms of choline in humans. The water-soluble choline-derived compounds (choline, phosphocholine, and glycerophosphocholine) are absorbed via the portal circulation whereas the lipid-soluble compounds (phosphatidylcholine and sphingomyelin) present in foods are absorbed into lymph as chylomicrons via the thoracic duct. This results in differential delivery and kinetics of distribution to tissues (Cheng et al., 1996; Zeisel et al., 1980b).
FINDINGS BY LIFE STAGE AND GENDER GROUP
Data are not sufficient for deriving an Estimated Average Requirement (EAR) for choline. The two published studies in healthy humans used male subjects only and tested a single level of choline intake. For these reasons only an Adequate Intake (AI) can be estimated. This amount will be influenced by the availability of methionine and folate in the diet. It may be influenced by gender, pregnancy, lactation, and stage of development. Although AIs are set for choline, it may be that the choline requirement can be met by endogenous synthesis at some of these stages.
To date, all studies have used choline-free diets and compared them with choline-containing diets; no intermediate levels of defi-
ciency have been reported. Careful dose-response experiments are needed before an EAR can be derived.
Infants Ages 0 through 12 Months
Method Used to Set the AI
An AI is used as the goal for intake by infants.
Ages 0 through 6 Months. The AI reflects the observed mean intake of choline by infants consuming human milk. Thus the choline AI for young infants is based on mean intake data from infants fed human milk exclusively for their first 6 months and uses the choline concentration of milk produced by well-nourished mothers. Human milk contains 160 to 210 mg (1.5 to 2 mmol)/L of choline moiety delivered as choline, phosphocholine, glycerophosphocholine, phosphatidylcholine, and sphingomyelin (Holmes-McNary et al., 1996; Zeisel et al., 1986). The choline phospholipids sphingomyelin and phosphatidylcholine are part of the milk fat-globule membrane (Holmes-McNary et al., 1996; Zeisel et al., 1986).
Rat pups denied access to milk have lower serum choline concentrations than do their fed litter mates (Zeisel and Wurtman, 1981). Thus, milk intake contributes to the maintenance of high serum choline concentrations in the neonate. In the rat, supplemental choline is concentrated in the rat dam’s milk (Garner et al., 1995; Zeisel, 1987). In women consuming a low-choline diet, milk choline content is lower than that in women consuming a more adequate diet (Zeisel et al., 1982). Consumption of either a choline-deficient or choline-supplemented diet by lactating rat dams results in significant changes in the phosphocholine concentration of their milk (Holmes-McNary et al., 1996; Zeisel et al., 1986) .The concentration of total choline in human milk is 160 mg/L (1.5 mmol/L). For the mean volume of output of human milk of 0.78 L/day and the average choline content of 160 mg/L, the AI for choline is 125 mg/day (1.2 mmol/day) for infants ages 0 through 6 months. For the reference infant weight of 7 kg, this corresponds to an AI of 18 mg/kg of body weight/day (0.17 mmol/kg/day).
Ages 7 through 12 Months. If the reference body weight ratio method described in Chapter 2 to extrapolate from the AI for choline for infants ages 0 through 6 months is used, the AI for choline for the older infants would be 150 mg/day (1.4 mmol/day). The second method (see Chapter 2), extrapolating from the AI for adults, gives
an AI that is essentially the same as that from extrapolating from infants. There are no data estimating choline intake from foods for this age group.
Choline AI Summary, Ages 0 through 12 Months
|
AI for Infants |
||
|
0–6 months |
125 mg/day of choline |
≈18 mg/kg |
|
7–12 months |
150 mg/day of choline |
≈17 mg/kg |
Special Considerations
Although commercially available infant formulas and bovine milk both contain choline and choline-containing compounds (Holmes-McNary et al., 1996; Rohlfs et al., 1993; Zeisel et al., 1986), human milk has a significantly higher phosphocholine concentration (718 µmol/L) than does either cow milk or infant formulas. However, cow milk and cow-milk-derived infant formulas have the same glycerophosphocholine concentration as human milk (400 to 800 µmol/L) (Holmes-McNary et al., 1996) or higher (415 µmol/L) (Holmes-McNary et al., 1996). Soy-derived infant formulas have lower glycerophosphocholine concentration (115 µmol/L or less) (Holmes-McNary et al., 1996). Human milk phosphatidylcholine and sphingomyelin concentrations do not differ significantly from those in cow milk and cow-milk-derived infant formulas (200 µmol/ L) (Holmes-McNary et al., 1996). Soy-derived infant formulas contain more phosphatidylcholine than do either human milk or cowmilk-derived formulas but less sphingomyelin than human milk (Holmes-McNary et al., 1996). Unesterified choline concentration in mature human milk is 30 to 80 percent lower than in either cow milk or the infant formulas (Holmes-McNary et al., 1996). The relative bioavailability of choline, phosphocholine, and glycerophosphocholine is similar in a rat model (Cheng et al., 1996) but no information is available for humans. Thus, it is not known whether these differences in milk and formula composition are clinically relevant.
Children and Adolescents Ages 1 through 18 Years
Method Used to Set the AI
No direct data on choline were found on which to base an EAR or AI for children and adolescents. In the absence of additional infor-
mation, AIs for these age groups have been extrapolated from adult values by using the method described in Chapter 2.
Choline AI Summary, Ages 1 through 18 Years
|
AI for Children |
1–3 years |
200 mg/day of choline |
|
4–8 years |
250 mg/day of choline |
|
|
AI for Boys |
9–13 years |
375 mg/day of choline |
|
14–18 years |
550 mg/day of choline |
|
|
AI for Girls |
9–13 years |
375 mg/day of choline |
|
14–18 years |
400 mg/day of choline |
Adults Ages 19 Years and Older
Method Used to Set the AI
An intake level of 500 mg/day (4.8 mmol/day; approximately 7 mg/kg/day [0.7mmol/kg/day]) of choline base is the dose that prevented alanine aminotransferase abnormalities in healthy men (Zeisel et al., 1991). This estimate for an AI is uncertain because it is based on a single published study; it may need revision when other data become available. This estimate fits within the bracketing estimates derived from patients on total parenteral nutrition for whom approximately 2 mg/kg/day of choline moiety did not prevent a deficiency syndrome (Sheard et al., 1986) and 31 mg/kg/day of choline moiety restored normal choline status (Buchman et al., 1992, 1993). The amount estimated as adequate for men should be sufficient to prevent an increase in alanine aminotransferase but it resulted in a small decrease in plasma choline in the one study in which it was evaluated, which suggests that dietary intake normally might be slightly higher. Thus the AI is set at approximately 7 mg/ kg/day or, for the reference man weighing 76 kg, at 550 mg after rounding.
To arrive at an estimate for AI for women, it is assumed that data from men can be used even though women may use choline more efficiently (see “Gender”). No experimental attempts to make healthy women choline deficient have been reported. However, women on total parenteral nutrition were just as likely as were men to develop low plasma choline concentrations and fatty liver (Buchman et al., 1995).
No experimental data are available from which to calculate an AI for life stage groups other than adults as a whole.
Choline AI Summary, Ages 19 Years and Older
The AI for choline in all forms for men in all age groups is 550 mg and for women is 425 mg. It is not known whether women have the same requirement on a body weight basis as men, but this AI is likely to be adequate on the basis of the earlier discussion on gender. Although there is some evidence that transport across the blood-brain barrier is diminished in the elderly, which suggests the possibility of a higher requirement than for younger adults (Cohen et al., 1995), no adjustment has been made in the AI for the elderly.
|
AI for Men |
19–30 years |
550 mg/day of choline |
|
31–50 years |
550 mg/day of choline |
|
|
51–70 years |
550 mg/day of choline |
|
|
> 70 years |
550 mg/day of choline |
|
|
AI for Women |
19–30 years |
425 mg/day of choline |
|
31–50 years |
425 mg/day of choline |
|
|
51–70 years |
425 mg/day of choline |
|
|
> 70 years |
425 mg/day of choline |
Pregnancy
Evidence Considered in Setting the AI
The need for choline is probably higher for pregnant than for nonpregnant women on the basis of animal data. Pregnancy renders female rats as vulnerable to deficiency as males (Zeisel et al., 1995). During pregnancy in humans (Welsch 1978; Welsch et al., 1981), guinea pigs (Swiery and Yudilevich, 1985; Swiery et al., 1986; Yudilevich and Sweiry, 1985), and rats (Jorswieck, 1974) large amounts of choline are delivered to the fetus through the placenta. Transport of choline from mother to fetus depletes maternal stores of choline; the choline concentration of maternal liver fell from a mean of 130 µmol/L in adult nonpregnant rats to 38 µmol/L in late pregnancy (Gwee and Sim, 1978).
Choline availability during embryogenesis and perinatal development may be especially important. In rats fed adequate diets during pregnancy, postnatally, and at weaning, 1 mmol/day of extra dietary choline results in long-lasting enhancement of spatial memory
(Meek and Williams, 1997a, b, c), altered morphology of septal neurons (Loy et al., 1991; Williams et al., 1998), and enhanced hippocampal long-term potentiation (Pyapali et al., 1998) and cholinergic neurotransmission (Cermak et al., 1998; Holler et al., 1996). The two periods of sensitivity to extra choline occur during embryonic days 12 to 17 and postnatal days 16 to 30 (Loy et al., 1991; Meek et al., 1988, 1989).
In mammals the placenta transports choline to the fetus (Welsch, 1976); choline concentration in amniotic fluid is 10-fold greater than that in maternal blood (S.Zeisel, University of North Carolina School of Public Health, unpublished observations, 1997). At birth, humans and other mammals have plasma choline concentrations that are much higher than those in adults (Zeisel et al., 1980a). It is not known whether de novo synthesis of choline increases during pregnancy.
The AI for pregnant women is greater than that for the adult by the amount needed for the fetus and placenta. Through the use of published values for the choline concentration of various adult rat tissues (Pomfret et al., 1989) and with the assumption of a body organ weight percentage as estimated by Widdowson (1963) for the human fetus, the fetal choline content can be estimated as approximately 5 mmol/kg (520 mg/kg) fetal weight. Human placental tissue has been estimated to average 1.26 ± 0.24 mmol/kg (mean ± standard error) in a small sample (n = 7) (Welsch, 1976); a value of approximately 2 mmol of choline per kg of placental tissue should cover almost all pregnant women. If it is thus assumed that the average choline content of fetal and placental tissue combined is approximately 3 mmol/kg (312 mg/kg), that there is no extra synthesis during pregnancy, and that there is no contribution of choline by placental or fetal synthesis, the required dietary amount of choline for the 10 kg of tissue that comprises the fetus (3 kg) and organs of pregnancy (7 kg) is 30 mmol, or 3,000 mg (10 kg tissue × 312 mg), which is approximately 11 mg/day (10 µmol/day) of additional dietary choline throughout pregnancy. This amount would be achieved by increasing the AI (after rounding) to 450 mg/day of choline for pregnancy.
Choline AI Summary, Pregnancy
The increase in the AI to support pregnancy is based on the fetal and placental accumulation of choline.
|
AI for Pregnancy |
14–18 years |
450 mg/day of choline |
|
19–30 years |
450 mg/day of choline |
|
|
31–50 years |
450 mg/day of choline |
Lactation
Method Used to Set the AI
The need for choline is likely to be increased during lactation because a substantial amount of choline is secreted in human milk, and mechanisms for conserving maternal choline status have not been identified. Lactating rats are more sensitive to choline deficiency than are nonlactating rats (Zeisel et al., 1995).
The AI for women during the first 6 months of lactation should be increased above that in the nonpregnant, nonlactating woman to cover the choline that is transferred into milk. For the assumption of an average volume production of 0.78 L/day (see Chapter 2) and an average choline content of milk of 156 mg/L (1.5 mmol/L), this increase is 125 mg/day (1.2 mmol/day). This increase is based on an assumption of 100 percent efficiency. It is not known whether de novo synthesis of choline increases during lactation. Women who are breastfeeding older infants who are also eating solid foods may need slightly less because of a lower volume of milk production.
Choline AI Summary, Lactation
|
AI for Lactation |
14–18 years |
550 mg/day of choline |
|
19–30 years |
550 mg/day of choline |
|
|
31–50 years |
550 mg/day of choline |
INTAKE OF CHOLINE
Food Sources
Choline is widely distributed in foods, with most of it in the form of phosphatidylcholine in membranes. Foods that are especially rich in choline compounds are milk, liver, eggs, and peanuts. It is possible to consume a diet of normal foods that delivers 1 g/day of choline (Zeisel et al., 1980b). Lecithins added during food processing may increase the average daily per capita consumption of phosphatidylcholine by 1.5 mg/kg of body weight for adults (this corre-
sponds to 0.225 mg/kg of body weight of choline moiety) (SCOGS/ LSRO, 1979).
Dietary Intake
Choline intake is not reported in the Third National Health and Nutrition Examination Survey (Perloff et al., 1990), the Continuing Survey of Food Intake by Individuals (Perloff et al., 1990), or the Boston Nutritional Status Survey (Hartz et al., 1992), and the choline content of foods is not included in major nutrient databases. There are no reports on choline intake from Canada. Estimated average choline dietary intake in adults consuming a typical U.S. or Canadian diet (as free choline and the choline in phosphatidylcholine and other choline esters) is approximately 730 to 1,040 mg/day (7 to 10 mmol/day) (LSRO/FASEB, 1981; Zeisel, 1981). Calculations of dietary choline intake are based on estimates of the free choline and phosphatidylcholine content of foods (Engel, 1943; McIntire et al., 1944; Weihrauch and Son, 1983; Zeisel et al., 1986). Older assay procedures for choline were imprecise and did not always include glycerophosphocholine or phosphocholine content, making many of the available data unreliable. On the basis of a finding of decreased plasma choline and phosphatidylcholine concentrations when humans were switched from a diet of normal foods to a defined diet containing 500 mg/day of choline (Zeisel et al., 1991), the average dietary intake of choline probably exceeds this level in adults. Infant formulas contain approximately 240 mg/L (2.3 mmol/L) of choline in its various forms. (Holmes-McNary et al., 1996).
Intake from Supplements
Choline is available as a dietary supplement as choline chloride or choline bitartrate and as lecithin, which usually contains approximately 25 percent phosphatidylcholine or 3 to 4 percent choline by weight. In the treatment of neurological diseases, large doses (5 to 30 g) of choline and phosphatidylcholine have been administered to humans (LSRO/FASEB, 1981). There are no reliable estimates of the frequency of use or amount of these dietary supplements consumed by individuals in the United States and Canada.
TOLERABLE UPPER INTAKE LEVELS
Hazard Identification
Adverse Effects
Choline doses that are orders of magnitude greater than estimated intake from food have been associated with body odor, sweating, salivation, hypotension, and hepatotoxicity in humans (LSRO/ FASEB, 1975, 1981). There are no indications in the literature that excess choline intake produces any additional adverse effects in humans. The animal data provide supportive evidence for a low degree of toxicity of choline. However, some animal studies have indicated growth suppression at high intakes (LSRO/FASEB, 1975). Because of the large doses and routes of administration used (e.g., intravenous and intraperitoneal injection), they were considered not relevant to human intakes from food and supplements (Davis, 1944; Hodge, 1945; Sahu, 1989; Sahu et al., 1986).
Body Odor, Sweating, and Salivation. High doses of choline have been associated with fishy body odor, vomiting, salivation, sweating, and gastrointestinal effects (LSRO/FASEB, 1981). These symptoms were reported in patients with tardive dyskinesia and cerebellar ataxia treated with choline chloride at 150 and 220 mg/kg of body weight/day for 2 to 6 weeks (10 and 16 g/day, respectively) (Davis et al., 1975; Growdon et al., 1977b; Lawrence et al., 1980). Studies of the production of methylamines from ingested choline suggest that fishy odor would have been observed in healthy populations (Zeisel et al., 1983). Fishy body odor results from the excretion of excessive amounts of trimethylamine, a choline metabolite, as the result of bacterial action. Lecithin, a choline-containing phospholipid, does not present a risk of fishy body odor because it generates little methylamine because the bacterial enzyme cannot cleave the ester (Zeisel et al., 1983).
Hypotension. Oral administration of 10 g/day of choline chloride (which is equivalent to 7.5 g [72 mmol] of choline alone) had a slight hypotensive effect in humans (Boyd et al., 1977). Choline could be acting by increasing vagal tone to the heart or by dilating arterioles. Although added choline increases acetylcholine release from in vitro preparations of heart (Loffelholz, 1981), changes in cardiac rate have not been observed in healthy humans treated with choline.
Hepatotoxicity. Mild hepatotoxicity was reported in patients receiving choline magnesium trisalicylate (1,500 mg twice daily for 8 days) (Cersosimo and Matthews, 1987). There is also one reported case of severe hypersensitivity hepatitis with striking tissue and peripheral eosinophilia after ingestion of choline magnesium trisalicylate (Nadkarni et al., 1992). However, it is likely that hepatotoxicity was induced by salicylate rather than by choline (Cersosimo and Matthews, 1987). Humans with and without cirrhosis have been treated with large doses of choline chloride (6 g/day for 4 weeks) with no resultant liver toxicity (Chawla et al., 1989).
Nonspecific Toxicity. Tinnitus and pruritus have been reported in patients treated with doses of 3 g/day of choline magnesium trisalicylate for 6 weeks. These side effects were transient and probably caused by salicylate (Mody et al., 1983). The salicylate effect likely accounts for many of these observations, and the others are likely unusual anomalies, such as the one case of contact dermatitis reported after dermal exposure to choline chloride (Fischer, 1984).
Identification of Sensitive Subpopulations
Individuals with trimethylaminuria (fish odor syndrome), renal disease, liver disease, depression, and Parkinson’s disease may have increased susceptibility to the adverse effects of choline. Trimethylaminuria results from a rare genetic deficiency that causes excessive excretion of trimethylamine and, therefore, an increased risk of developing fishy body odor (Al-Waiz et al., 1988, 1989; Humbert et al., 1970; Shelley and Shelley, 1984). Individuals with renal or liver disease may have increased susceptibility because of increased levels of plasma choline (after ingestion of supplemental choline) compared with healthy individuals (Acara and Rennick, 1973; Acara et al., 1983; Chawla et al., 1989; Rennick et al., 1976). In rare cases, consumption of large amounts of choline has been associated with depression (Davis et al., 1979; Tamminga et al., 1976). Finally, mild and transient Parkinsonian signs (bradykinesia, tremor, and rigidity) were observed at high doses (12.7 g/day) of choline as a chloride in people with tardive dyskinesia (Gelenberg et al., 1979), which suggests that supplemental choline intake by Parkinsonian patients may exacerbate symptoms.
Summary
On the basis of considerations of causality, relevance, and the
quality and completeness of the database, hypotension was selected as the critical effect in deriving a Tolerable Upper Intake Level (UL); fishy body odor was selected as the secondary consideration.
Dose-Response Assessment
Adults
Data Selection. The data used to derive the UL for choline include a single case report of hypotension and several other studies involving cholinergic effects and fishy body odor after oral administration of large choline doses.
Identification of a no-observed-adverse-effect level (NOAEL) and a lowest-observed-adverse-effect level (LOAEL). There are no adequate data demonstrating a NOAEL for excess choline intake. A LOAEL of approximately 7.5 g/day of choline can be identified from evaluation of a pilot study that reported hypotension in seven patients treated with choline for Alzheimer senile dementia (Boyd et al., 1977) and reports of fishy body odor in individuals treated with choline for tardive dyskinesia and Huntington’s disease (Gelenberg et al., 1979; Growdon et al., 1977a, b; Lawrence et al., 1980). Boyd et al. (1977) treated seven older adult patients with 4 g/day of oral choline as choline chloride for 2 weeks followed by 2 weeks of choline at 7.5 g/day. At 4 g/day of choline, daily blood pressure recordings revealed no hypotension. In addition, there were no reports of nausea or diarrhea or other evidence of cholinergic effects at this dose level. At 7.5 g/day of choline, nausea, diarrhea, and a small decrease in blood pressure were reported in some patients. Other supportive data on cholinergic effects and fishy body odor after excess choline intake are summarized in Table 12-1.
Uncertainty Assessment. An uncertainty factor (UF) of 2 was selected because of the limited data regarding hypotension and the interindividual variation in response to cholinergic effects.
Derivation of a UL. A LOAEL of 7.5 g/day was divided by an UF of 2 to obtain a UL of 3.75 for adults, which was rounded down to 3.5 g/day.
Choline UL Summary, Adults
Because of the scarcity of data for any adult age group and
TABLE 12-1 Studies Reporting on Cholinergic Effects and Fishy Body Odor after Excess Choline Intake
|
Study |
No. of Subjects |
Dose |
Duration (wk) |
Adverse Effects |
|
Growdon et al., 1977aa |
20 |
2 |
Mild cholinergic toxicity: lacrimation, blurred vision, anorexia, and diarrhea. |
|
|
Growdon et al., 1977b |
10 |
8–20 g/dd |
2–17 |
Fishy body odor in all subjects; at 250–300 mg/ kg/d, produced lacrimation, anorexia, vomiting, and diarrhea. |
|
Gelenberg et al., 1979e |
5 |
8–19 g/dd |
6–8 |
100% with fishy body odor after several days; gastrointestinal irritation.f |
|
Lawrence et al., 1980g |
14 |
6 |
At 150 mg/kg/d: 5 of 14 with fishy body odor; 12 of 15 with nausea and diarrhea. |
|
|
a Study involved a double-blind, crossover protocol. b Choline was given as a chloride or bitartrate. c Doses were calculated from data in the report using a reference body weight of 61 kg. Depending on the body weights of the individuals in Lawrence et al. (1980) and Growdon et al. (1977a), the lowest-effect dose may be less than 7.5 g/d. d Choline was given as a chloride. e Nonblinded study; did not include a control group. f Mild, transient Parkinsonian signs (bradykinesia, tremor, and rigidity) were also reported. g Double-blind protocol; included control group. |
||||
because no specific physiological function might be expected to affect sensitivity to excess amounts of choline in older persons, no adjustments are proposed for the elderly.
|
UL for Adults |
19 years and older |
3.5 g/day of choline |
Other Life Stage Groups
For infants, the UL was judged not determinable because of lack of data concerning adverse effects in this age group and concern
about the infant’s ability to handle excess amounts. The only source of intake for infants should be from food or formula to prevent high levels of intake. There are no data to suggest that during pregnancy or lactation increased susceptibility to developing cholinergic effects or fishy body odor from excess choline intake would occur. Therefore, the UL of 3.5 g/day is also set for pregnant and lactating women. The UL of 3.5 g/day for adults was adjusted for children and adolescents on the basis of relative body weight as described in Chapter 3, with the use of reference weights from Chapter 1, Table 1-2. Values have been rounded down.
Choline UL Summary, Other Life Stage Groups
|
UL For Infants |
||
|
0–12 months |
Not possible to establish; source of intake should be formula and food only |
|
|
UL for Children |
1–3 years |
1 g/day of choline |
|
4–8 years |
1 g/day of choline |
|
|
9–13 years |
2 g/day of choline |
|
|
UL for Adolescents |
14–18 years |
3 g/day of choline |
|
UL for Pregnancy |
14–18 years |
3 g/day of choline |
|
19 years and older |
3.5 g/day of choline |
|
|
UL for Lactation |
14–18 years |
3 g/day of choline |
|
19 years and older |
3.5 g/day of choline |
|
Special Considerations
Individuals with the following conditions may be at risk of adverse effects with choline intakes at the UL: trimethylaminuria, renal disease, liver disease, depression, and Parkinson’s disease.
Intake Assessment
National surveys do not provide data on the dietary intake of choline. The UL applies to the weight of the choline moiety in the compound; for example, choline chloride contains more choline by weight than does choline bitartrate. Dietary supplements containing choline are available; however, reliable estimates of the
amount of these supplements consumed in the United States and Canada are unavailable.
Risk Characterization
Because there is no information from national surveys on choline intakes or on supplement usage, the risk of adverse effects within the United States or Canada can not be characterized.
RESEARCH RECOMMENDATIONS FOR CHOLINE
High Priority Recommendations
Sufficient human data are not available for determining whether choline is essential in the human diet, how much is required if it is essential, and the public health impact of poor choline nutriture. For this reason, research that could provide such human data is assigned the highest priority:
-
Examination of the effects of the use of graded levels of dietary intake of choline on parameters of health. This would include assessing plasma and tissue choline compounds and metabolites; plasma cholesterol and homocysteine concentrations; erythrocyte folate; and liver, renal, brain, and other organ function. To facilitate this process, food composition data are needed for choline, phosphocholine, glycerophosphocholine, sphingomyelin, phosphatidylcholine, and betaine and the analytic sensitivity and specificity of methods for analysis of food composition need to be validated.
-
Human studies on interrelationships among requirements for choline, methionine, folate, vitamin B6, and vitamin B12 to compare the homocysteine-lowering effects of combinations of these nutrients.
Other Research Areas
Two additional topics also merit attention:
-
The relative effectiveness of different choline-containing compounds in the diet in promoting health and determination of the sparing effect of endogenous synthesis of choline. It will be important to conduct studies on the bioavailability of choline and choline compounds and on the rate of de novo synthesis of choline in vivo.
-
Studies using increasing levels of dietary intake designed to assess toxicity for all organ systems, including heart, liver, brain and
-
kidney; fishy body odor; and possible growth suppression in children from observational data and as determined by experimental studies in animal models.
REFERENCES
Acara M, Rennick B. 1973. Regulation of plasma choline by the renal tubule: Bidirectional transport of choline. Am J Physiol 225:1123–1128.
Acara M, Rennick B, LaGraff S, Schroeder ET. 1983. Effect of renal transplantation on the levels of choline in the plasma of uremic humans. Nephron 35:241– 243.
Albright CD, Liu R, Bethea TC, da Costa KA, Salganik RI, Zeisel SH. 1996. Choline deficiency induces apoptosis in SV40-immortalized CWSV-1 rat hepatocytes in culture. FASEB J 10:510–516.
Al-Waiz M, Ayesh R, Mitchell SC, Idle JR, Smith RL. 1988. Trimethylaminuria (“fishodour syndrome”): A study of an affected family. Clin Sci 74:231–236.
Al-Waiz M, Ayesh R, Mitchell SC, Idle JR, Smith RL. 1989. Trimethylaminuria: The detection of carriers using a trimethylamine load test. J Inherit Metab Dis 12:80– 85.
Anonymous. 1997. Betaine for homocystinuria. Med Lett Drugs Ther 39:12.
Aquilonius SM, Ceder G, Lying-Tunell U, Malmlund HO, Schuberth J. 1975. The arteriovenous difference of choline across the brain of man. Brain Res 99:430– 433.
Arvidson GA. 1968. Biosynthesis of phosphatidylcholines in rat liver. Eur J Biochem 5:415–421.
Barak AJ, Kemmy RJ. 1982. Methotrexate effects on hepatic betaine levels in choline-supplemented and choline-deficient rats. Drug Nutr Interact 1:275–278.
Barak AJ, Tuma DJ, Beckenhauer HC. 1984. Methotrexate hepatotoxicity. J Am Coll Nutr 3:93–96.
Bartus RT, Dean RL, Goas JA, Lippa AS. 1980. Age-related changes in passive avoidance retention: Modulation with dietary choline. Science 209:301–303.
Bauernschmitt HG, Kinne RK. 1993. Metabolism of the “organic osmolyte” glycerophosphorylcholine in isolated rat inner medullary collecting duct cells. I. Pathways for synthesis and degradation. Biochim Biophys Acta 1148:331–341.
Best CH, Huntsman ME. 1932. The effects of the components of lecithine upon deposition of fat in the liver. J Physiol 75:405–412.
Bianchi G, Azzone GF. 1964. Oxidation of choline in rat liver mitochondria. J Biol Chem 239:3947–3955.
Bjornstad P, Bremer J. 1966. In vivo studies on pathways for the biosynthesis of lecithin in the rat. J Lipid Res 7:38–45.
Blair R, Newsome F. 1985. Involvement of water-soluble vitamins in diseases of swine. J Anim Sci 60:1508–1517.
Blair R, Whitehead CC, Bannister DW, Evans AJ. 1973. Involvement of diet in fatty liver and kidney syndrome in broiler chickens. Vet Rec 92:118–119.
Blusztajn JK, Zeisel SH, Wurtman RJ. 1979. Synthesis of lecithin (phosphatidylcholine) from phosphatidylethanolamine in bovine brain. Brain Res 179:319– 327.
Boyd WD, Graham-White J, Blackwood G, Glen I, McQueen J. 1977. Clinical effects of choline in Alzheimer senile dementia. Lancet 2:711.
Bremer J, Greenberg D. 1961. Methyl transfering enzyme system of microsomes in the biosynthesis of lecithin (phosphatidylcholine). Biochim Biophys Acta 46:205– 216.
Buchman AL, Dubin M, Jenden D, Moukarzel A, Roch MH, Rice K, Gornbein J, Ament ME, Eckhert CD. 1992. Lecithin increases plasma free choline and decreases hepatic steatosis in long-term total parenteral nutrition patients. Gastroenterology 102:1363–1370.
Buchman AL, Moukarzel A, Jenden DJ, Roch M, Rice K, Ament ME. 1993. Low plasma free choline is prevalent in patients receiving long term parenteral nutrition and is associated with hepatic aminotransferase abnormalities. Clin Nutr 12:33–37.
Buchman AL, Dubin M, Moukarzel A, Jenden D, Roch M, Rice K, Gornbein J, Ament M. 1995. Choline deficiency: A cause of hepatic steatosis during parenteral nutrition that can be reversed with intravenous choline supplementation. Hepatology 22:1399–1403.
Burg MB. 1995. Molecular basis of osmotic regulation. Am J Physiol 268:F983-F996.
Burt ME, Hanin I, Brennan MF. 1980. Choline deficiency associated with total parenteral nutrition. Lancet 2:638–639.
Cermak JM, Holler T, Jackson DA, Blusztajn JK. 1998. Prenatal availability of choline modifies development of the hippocampal cholinergic system. FASEB J 12:349–357.
Cersosimo RJ, Matthews SJ. 1987. Hepatotoxicity associated with choline magnesium trisalicylate: Case report and review of salicylate-induced hepatotoxicity. Drug Intell Clin Pharm 21:621–625.
Chawla RK, Wolf DC, Kutner MH, Bonkovsky HL. 1989. Choline may be an essential nutrient in malnourished patients with cirrhosis. Gastroenterology 97:1514– 1520.
Cheng W-L, Holmes-McNary MQ, Mar M-H, Lien EL, Zeisel SH. 1996. Bioavailability of choline and choline esters from milk in rat pups. J Nutr Biochem 7:457– 464.
Cohen BM, Renshaw PF, Stoll AL, Wurtman RJ, Yurgelun-Todd D, Babb SM. 1995. Decreased brain choline uptake in older adults. An in vivo proton magnetic resonance spectroscopy study. J Am Med Assoc 274:902–907.
Cohen EL, Wurtman RJ. 1975. Brain acetylcholine: Increase after systemic choline administration. Life Sci 16:1095–1102.
Conlay LA, Wurtman RJ, Blusztajn K, Coviella IL, Maher TJ, Evoniuk GE. 1986. Decreased plasma choline concentrations in marathon runners. N Engl J Med 315:892.
Cornford EM, Cornford ME. 1986. Nutrient transport and the blood-brain barrier in developing animals. Fed Proc 45:2065–2072.
Crews FT, Calderini G, Battistella A, Toffano G. 1981. Age-dependent changes in the methylation of rat brain phospholipids. Brain Res 229:256–259.
Cui Z, Houweling M, Chen MH, Record M, Chap H, Vance DE, Tercé F. 1996. A genetic defect in phosphatidylcholine biosynthesis triggers apoptosis in Chinese hamster ovary cells. J Biol Chem 271:14668–14671.
da Costa KA, Cochary EF, Blusztajn JK, Garner SC, Zeisel SH. 1993. Accumulation of 1,2-sn-diradylglycerol with increased membrane-associated protein kinase C may be the mechanism for spontaneous hepatocarcinogenesis in cholinedeficient rats. J Biol Chem 268:2100–2105.
da Costa KA, Garner SC, Chang J, Zeisel SH. 1995. Effects of prolonged (1 year) choline deficiency and subsequent re-feeding of choline on 1,2-sn-diradylglycerol, fatty acids and protein kinase C in rat liver. Carcinogenesis 16:327–334.
Davis JE. 1944. Depression of normal erythrocyte number by soybean lecithin or choline. Am J Physiol 142:65–67.
Davis KL, Berger PA, Hollister LE. 1975. Choline for tardive dyskinesia. N Engl J Med 293:152.
Davis KL, Hollister LE, Berger PA. 1979. Choline chloride in schizophrenia. Am J Psychiatry 136:1581–1584.
Drouva SV, LaPlante E, Leblanc P, Bechet JJ, Clauser H, Kordon C. 1986. Estradiol activates methylating enzyme (s) involved in the conversion of phosphatidylethanolamine to phosphatidylcholine in rat pituitary membranes. Endocrinology 119:2611–2622.
Dudman NP, Tyrrell PA, Wilcken DE. 1987. Homocysteinemia: Depressed plasma serine levels. Metabolism 36:198–201.
Eagle H. 1955. The minimum vitamin requirements of the L and HeLa cells in tissue culture, the production of specific vitamin deficiencies, and their cure. J Exp Med 102:595–600.
Engel RW. 1943. The choline content of animal and plant products. J Nutr 25:441– 446.
Exton JH. 1994. Phosphatidylcholine breakdown and signal transduction. Biochim Biophys Acta 1212:26–42.
Fairbanks BW, Krider JL. 1945. Significance of the B vitamins in swine nutrition. N Am Vet 26:18–23.
Finkelstein JD, Martin JJ, Harris BJ, Kyle WE. 1982. Regulation of the betaine content of rat liver. Arch Biochem Biophys 218:169–173.
Finkelstein JD, Martin JJ, Harris BJ. 1988. Methionine metabolism in mammals. The methionine-sparing effect of cystine. J Biol Chem 263:11750–11754.
Fischer T. 1984. Contact allergy to choline chloride. Contact Dermatitis 10:316–317.
Freeman-Narrod M, Narrod SA, Custer RP. 1977. Chronic toxicity of methotrexate in rats: Partial to complete protection of the liver by choline. J Natl Cancer Inst 59:1013–1017.
Frenkel R, Muguruma K, Johnston J. 1996. The biochemical role of platelet-activating factor in reproduction. Prog Lipid Res 35:155–168.
Garcia-Perez A, Burg MB. 1991. Role of organic osmolytes in adaptation of renal cells to high osmolality. J Membr Biol 119:1–13.
Garner SC, Mar MH, Zeisel SH. 1995. Choline distribution and metabolism in pregnant rats and fetuses are influenced by the choline content of the maternal diet. J Nutr 125:2851–2858.
Gelenberg AJ, Doller-Wojcik J, Growdon JH. 1979. Choline and lecithin in the treatment of tardive dyskinesia: Preliminary results from a pilot study. Am J Psychiatry 136:772–776.
Grossman EB, Hebert SC. 1989. Renal inner medullary choline dehydrogenase activity: Characterization and modulation. Am J Physiol 256:F107–F112.
Growdon JH, Cohen EL, Wurtman RJ. 1977a. Huntington’s disease: Clinical and chemical effects of choline administration. Ann Neurol 1:418–422.
Growdon JH, Hirsch MJ, Wurtman RJ, Wiener W. 1977b. Oral choline administration to patients with tardive dyskinesia. N Engl J Med 297:524–527.
Gwee MC, Sim MK. 1978. Free choline concentration and cephalin-N-methyltransferase activity in the maternal and foetal liver and placenta of pregnant rats. Clin Exp Pharmacol Physiol 5:649–653.
Handler P. 1949. Response of guinea pigs to diets deficient in choline. Proc Soc Exp Biol Med 70:70–73.
Hannun YA. 1994. The sphingomyelin cycle and the second messenger function of ceramide. J Biol Chem 269:3125–3128.
Hartz SC, Russell RM, Rosenberg IH. 1992. Nutrition in the Elderly. The Boston Nutritional Status Survey. London: Smith-Gordon.
Haubrich DR, Wedeking PW, Wang PF. 1974. Increase in tissue concentration of acetylcholine in guinea pigs in vivo induced by administration of choline. Life Sci 14:921–927.
Hershey JM. 1931. Substitution of lecithin for raw pancreas in the diet of depancreatized dog. Am J Physiol 93:657–658.
Herzberg GR, Lerner J. 1973. Intestinal absorption of choline in the chick. Biochim Biophys Acta 307:234–242.
Herzberg GR, Sheerin H, Lerner J. 1971. Cationic amino acid transport in chicken small intestine. Comp Biochem Physiol 40A:229–247.
Hirsch MJ, Growdon JH, Wurtman RJ. 1978. Relations between dietary choline or lecithin intake, serum choline levels, and various metabolic indices. Metabolism 27:953–960.
Hodge HC. 1945. Chronic oral toxicology of choline chloride in rats. Proc Exp Biol Med 58:212–215.
Hoffbauer FW, Zaki FG. 1965. Choline deficiency in baboon and rat compared. Arch Pathol 79:364–369.
Holler T, Cermak JM, Blusztajn JK. 1996. Dietary choline supplementation in pregnant rats increases hippocampal phospholipase D activity of the offspring. FASEB J 10:1653–1659.
Holmes-McNary MQ, Cheng WL, Mar MH, Fussell S, Zeisel SH. 1996. Choline and choline esters in human and rat milk and in infant formulas. Am J Clin Nutr 64:572–576.
Holmes-McNary MQ, Loy R, Mar MH, Albright CD, Zeisel SH. 1997. Apoptosis is induced by choline deficiency in fetal brain and in PC12 cells. Brain Res Dev Brain Res 101:9–16.
Horne DW, Cook RJ, Wagner C. 1989. Effect of dietary methyl group deficiency on folate metabolism in rats. J Nutr 119:618–621.
Humbert JA, Hammond KB, Hathaway WE. 1970. Trimethylaminuria: The fishodor syndrome. Lancet 2:770–771.
Jacob RA, Pianalto FS, Henning SM, Zhang JZ, Swendseid ME. 1995. In vivo methylation capacity is not impaired in healthy men during short-term dietary folate and methyl group restriction. J Nutr 125:1495–1502.
James ST, Miller BT, Basnakian AG, Pogribny IP, Pogribna M, Muskhelishvili L. 1997. Apoptosis and proliferation under conditions of deoxynucleotide pool imbalance in liver of folate/methyl deficient rats. Carcinogenesis 18:287–293.
Jorswieck I. 1974. Proceedings: Penetration of choline through rat placenta in vivo. Naunyn Schmiedebergs Arch Pharmakol 282:R42.
Kennedy EP, Weiss SB. 1956. The function of cytidine coenzymes in the biosynthesis of phospholipids. J Biol Chem 222:193–214.
Ketola HG. 1976. Choline metabolism and nutritional requirement of lake trout (Salvelinus namaycush). J Anim Sci 43:474–477.
Ketola HG, Nesheim MC. 1974. Influence of dietary protein and methionine levels on the requirement for choline by chickens. J Nutr 104:1484–1489.
Ketola HG, Young RJ. 1973. The need for dietary choline by young Japanese quail. Poult Sci 52:2362–2363.
Kim Y-I, Miller JW, da Costa K-A, Nadeau M, Smith D, Selhub J, Zeisel SH, Mason JB. 1994. Severe folate deficiency causes secondary depletion of choline and phosphocholine in rat liver. J Nutr 124:2197–2203.
Kuczler FJ, Nahrwold DL, Rose RC. 1977. Choline influx across the brush border of guinea pig jejunum. Biochim Biophys Acta 465:131–137.
Kuksis A, Mookerjea S. 1978. Choline. Nutr Rev 36:201–207.
Lawrence CM, Millac P, Stout GS, Ward JW. 1980. The use of choline chloride in ataxic disorders. J Neurol Neurosurg Psychiatry 43:452–454.
Le Kim D, Betzing H. 1976. Intestinal absorption of polyunsaturated phosphatidylcholine in the rat. Hoppe Seylers Z Physiol Chem 357:1321–1331.
Lindblad L, Schersten T. 1976. Incorporation rate in vitro of choline and methylmethionine into human hepatic lecithins. Scand J Gastroenterol 11:587–591.
Loffelholz K. 1981. Release of acetylcholine in the isolated heart. Am J Physiol 240:H431–H440.
Loy R, Heyer D, Williams CL, Meek WH. 1991. Choline-induced spatial memory facilitation correlates with altered distribution and morphology of septal neurons. Adv Exp Med Biol 295:373–382.
LSRO/FASEB (Life Sciences Research Office/Federation of American Societies for Experimental Biology). 1975. Evaluation of the Health Aspects of Choline Chloride and Choline Bitartrate as Food Ingredients. Report # PB-223 845/9. Washington, DC: Department of Health, Education and Welfare.
LSRO/FASEB (Life Sciences Research Office/Federation of American Societies for Experimental Biology). 1981. Effects of Consumption of Choline and Lecithin on Neurological and Cardiovascular Systems. Report # PB–82–133257. Bethesda, MD: LSRO/FASEB.
Lyman RL, Sheehan G, Tinoco J. 1971. Diet and 14CH3-methionine incorporation into liver phosphatidylcholine fractions of male and female rats. Can J Biochem 49:71–79.
McIntire JM, Schweigert BS, Elvehjem CA. 1944. The choline and pyridoxine content of meats. J Nutr 28:219–223.
Meek WH, Williams CL. 1997a. Characterization of the facilitative effects of perinatal choline supplementation on timing and temporal memory. Neuroreport 8:2831–2835.
Meek WH, Williams CL. 1997b. Perinatal choline supplementation increases the threshold for chunking in spatial memory. Neuroreport 8:3053–3059.
Meek WH, Williams CL. 1997c. Simultaneous temporal processing is sensitive to prenatal choline availability in mature and aged rats. Neuroreport 8:3045–3051.
Meek WH, Smith RA, Williams CL. 1988. Pre- and postnatal choline supplementation produces long-term facilitation of spatial memory. Dev Psychobiol 21:339– 353.
Meek WH, Smith RA, Williams CL. 1989. Organizational changes in cholinergic activity and enhanced visuospatial memory as a function of choline administered prenatally or postnatally or both. Behav Neurosci 103:1234–1241.
Mody GM, Naidoo PD, Singh TG. 1983. Clinical evaluation of choline magnesium trisalicylate in rheumatoid arthritis. S Afr Med J 64:195–196.
Mudd SH, Poole JR. 1975. Labile methyl balances for normal humans on various dietary regimens. Metabolism 24:721–735.
Nadkarni MM, Peller CA, Retig J. 1992. Eosinophilic hepatitis after ingestion of choline magnesium trisalicylate. Am J Gastroenterol 87:151–153.
Newberne PM, Rogers AE. 1986. Labile methyl groups and the promotion of cancer. Annu Rev Nutr 6:407–432.
Perloff BP, Rizek RL, Haytowitz DB, Reid PR. 1990. Dietary intake methodology. II. USDA’s Nutrient Data Base for Nationwide Dietary Intake Surveys. J Nutr 120:1530–1534.
Poirier LA, Grantham PH, Rogers AE. 1977. The effects of a marginally lipotropedeficient diet on the hepatic levels of S-adenosylmethionine and on the urinary metabolites of 2-acetylaminofluorene in rats. Cancer Res 37:744–748.
Pomfret EA, da Costa K-A, Schurman LL, Zeisel SH. 1989. Measurement of choline and choline metabolite concentrations using high-pressure liquid chromatography and gas chromatography-mass spectrometry. Analy Biochem 180:85–90.
Pomfret EA, da Costa K, Zeisel SH. 1990. Effects of choline deficiency and methotrexate treatment upon rat liver. J Nutr Biochem 1:533–541.
Pyapali GK, Turner DA, Williams CL, Meek WH, Swartzwelder HS. 1998. Prenatal dietary choline supplementation decreases the threshold for induction of long-term potentiation in young adult rats. J Neurophysiol 79:1790–1796.
Rennick B, Acara M, Hysert P, Mookerjee B. 1976. Choline loss during hemodialysis: Homeostatic control of plasma choline concentrations. Kidney Int 10:329– 335.
Rennick B, Acara M, Glor M. 1977. Relations of renal transport rate, transport maximum, and competitor potency for tetraethylammonium and choline. Am J Physiol 232:F443–F447.
Ridgway ND, Vance DE. 1988. Kinetic mechanism of phosphatidylethanolamine N-methyltransferase. J Biol Chem 263:16864–16871.
Rohlfs EM, Garner SC, Mar MH, Zeisel SH. 1993. Glycerophosphocholine and phosphocholine are the major choline metabolites in rat milk. J Nutr 123:1762– 1768.
Sahu AP. 1989. Effect of Choline and Mineral Fibres (Chrysotile Asbestos) on Guinea-pigs. Lyon, France: IARC Scientific Publications.
Sahu AP, Saxena AK, Singh KP, Shanker R. 1986. Effect of chronic choline administration in rats. Indian J Exp Biol 24:91–96.
Sandage BW, Sabounjian L, White R, Wurtman RJ. 1992. Choline citrate may enhance athletic performance. Physiologist 35:236.
Savendahl L, Mar M-H, Underwood LE, Zeisel SH. 1997. Prolonged fasting in humans results in diminished plasma choline concentrations but does not cause liver dysfunction. Am J Clin Nutr 66:622–625.
SCOGS/LSRO (Select Committee on GRAS Substances, Life Sciences Research Office). 1979. Evaluation of the Health Aspects of Lecithin as a Food Ingredient. Report # PB301405. Springfield, VA: National Technical Information Service.
Selhub J, Seyoum E, Pomfret EA, Zeisel SH. 1991. Effects of choline deficiency and methotrexate treatment upon liver folate content and distribution. Cancer Res 51:16–21.
Shapira G, Chawla RK, Berry CJ, Williams PJ, Roy RGB, Rudman D. 1986. Cysteine, tyrosine, choline and carnitine supplementation of patients on total parenteral nutrition. Nutr Int 2:334–339.
Sheard NF, Zeisel SH. 1986. An in vitro study of choline uptake by intestine from neonatal and adult rats. Pediatr Res 20:768–772.
Sheard NF, Tayek JA, Bistrian BR, Blackburn GL, Zeisel SH. 1986. Plasma choline concentration in humans fed parenterally. Am J Clin Nutr 43:219–224.
Shelley ED, Shelley WB. 1984. The fish odor syndrome. Trimethylaminuria. JAMA 251:253–255.
Shin OH, Mar MH, Albright CD, Citarella MT, daCosta KA, Zeisel SH. 1997. Methylgroup donors cannot prevent apoptotic death of rat hepatocytes induced by choline-deficiency. J Cell Biochem 64:196–208.
Shivapurkar N, Poirier LA. 1983. Tissue levels of S-adenosylmethionine and S-adenosylhomocysteine in rats fed methyl-deficient, amino acid-defined diets for one to five weeks. Carcinogenesis 4:1051–1057.
Sundler R, Akesson B. 1975. Regulation of phospholipid biosynthesis in isolated rat hepatocytes. Effect of different substrates. J Biol Chem 250:3359–3367.
Svardal AM, Ueland PM, Berge RK, Aarsland A, Aarsaether N, Lonning PE, Refsum H. 1988. Effect of methotrexate on homocysteine and other sulfur compounds in tissues of rats fed a normal or a defined, choline-deficient diet. Cancer Chemother Pharmacol 21:313–318.
Sweiry JH, Yudilevich DL. 1985. Characterization of choline transport at maternal and fetal interfaces of the perfused guinea-pig placenta. J Physiol 366:251–266.
Sweiry JH, Page KR, Dacke CG, Abramovich DR, Yudilevich DL. 1986. Evidence of saturable uptake mechanisms at maternal and fetal sides of the perfused human placenta by rapid paired-tracer dilution: Studies with calcium and choline . J Dev Physiol 8:435–445.
Tamminga CA, Smith RC, Chang S, Haraszti JS, Davis JM. 1976. Depression associated with oral choline. Lancet 2:905.
Tani H, Suzuki S, Kobayashi M, Kotake Y. 1967. The physiological role of choline in guinea pigs. J Nutr 92:317–324.
Tayek JA, Bistrian B, Sheard NF, Zeisel SH, Blackburn GL. 1990. Abnormal liver function in malnourished patients receiving total parenteral nutrition: A prospective randomized study. J Am Coll Nutr 9:76–83.
Tessitore L, Sesca E, Greco M, Pani P, Dianzani M. 1995. Sexually differentiated response to choline in choline deficiency and ethionine intoxication. Int J Exp Pathol 76:125–129.
Vance DE. 1990. Boehringer Mannheim Award lecture. Phosphatidylcholine metabolism: Masochistic enzymology, metabolic regulation, and lipoprotein assembly. Biochem Cell Biol 68:1151–1165.
Vance DE, Ridgway ND. 1988. The methylation of phosphatidylethanolamine. Prog Lipid Res 27:61–79.
Varela-Moreiras G, Ragel C, Perez de Miguelsanz J. 1995. Choline deficiency and methotrexate treatment induces marked but reversible changes in hepatic folate concentrations, serum homocysteine and DNA methylation rates in rats. J Am Coll Nutr 14:480–485.
Von Allworden HN, Horn S, Kahl J, Feldheim W. 1993. The influence of lecithin on plasma choline concentrations in triatheletes and adolescent runners during exercise. Eur J Appl Physiol 67:87–91.
Wecker L. 1986. Neurochemical effects of choline supplementation. Can J Physiol Pharmacol 64:329–333.
Weihrauch JL, Son Y-S. 1983. The phospholipid content of foods. J Am Oil Chem Soc 60:1971–1978.
Weinhold PA, Sanders R. 1973. The oxidation of choline by liver slices and mitochondria during liver development in the rat. Life Sci 13:621–629.
Welsch F. 1976. Studies on accumulation and metabolic fate of (N-Me3H) choline in human term placenta fragments. Biochem Pharmacol 25:1021–1030.
Welsch F. 1978. Choline metabolism in human term placenta—studies on de novo synthesis and the effects of some drugs on the metabolic fate of [N-methyl 3H] choline. Biochem Pharmacol 27:1251–1257.
Welsch F, Wenger WC, Stedman DB. 1981. Choline metabolism in placenta: Evidence for the biosynthesis of phosphatidylcholine in microsomes via the methylation pathway. Placenta 2:211–221.
Wendel U, Bremer H. 1984. Betaine in the treatment of homocystinuria due to 5,10-methylenetetrahydrofolate reductase deficiency. Eur J Pediatr 142:147–150.
Widdowson EM. 1963. Growth and composition of the fetus and newborn. In: Assali N, ed. Biology of Gestation, Vol. 2. New York: Academic Press. Pp. 1–51.
Wilcken DE, Wilcken B, Dudman NP, Tyrrell PA. 1983. Homocystinuria—the effects of betaine in the treatment of patients not responsive to pyridoxine. N Engl J Med 309:448–453.
Wilcken DE, Dudman NP, Tyrrell PA. 1985. Homocystinuria due to cystathionine β-synthase deficiency—the effects of betaine treatment in pyridoxine-responsive patients. Metabolism 34:1115–1121.
Williams CL, Meck WH, Heyer D, Loy R. 1998. Hypertrophy of basal forebrain neurons and enhanced visuospatial memory in perinatally cholinesupplemented rats. Brain Res 794:225–238.
Wong ER, Thompson W. 1972. Choline oxidation and labile methyl groups in normal and choline-deficient rat liver. Biochim Biophys Acta 260:259–271.
Wood JL, Allison RG. 1982. Effects of consumption of choline and lecithin on neurological and cardiovascular systems. Fed Proc 41:3015–3021.
Yang EK, Blusztajn JK, Pomfret EA, Zeisel SH. 1988. Rat and human mammary tissue can synthesize choline moiety via the methylation of phosphatidylethanolamine. Biochem J 256:821–828.
Yao ZM, Vance DE. 1988. The active synthesis of phosphatidylcholine is required for very low density lipoprotein secretion from rat hepatocytes. J Biol Chem 263:2998–3004.
Yao ZM, Vance DE. 1989. Head group specificity in the requirement of phosphatidylcholine biosynthesis for very low density lipoprotein secretion from cultured hepatocytes. J Biol Chem 264:11373–11380.
Yao ZM, Vance DE. 1990. Reduction in VLDL, but not HDL, in plasma of rats deficient in choline. Biochem Cell Biol 68:552–558.
Young DL. 1971. Estradiol- and testosterone-induced alterations in phosphatidylcholine and triglyceride synthesis in hepatic endoplasmic reticulum. J Lipid Res 12:590–595.
Yudilevich DL, Sweiry JH. 1985. Membrane carriers and receptors at maternal and fetal sides of the placenta by single circulation paired-tracer dilution: Evidence for a choline transport system. Contrib Gynecol Obstet 13:158–161.
Zeisel SH. 1981. Dietary choline: Biochemistry, physiology, and pharmacology. Annu Rev Nutr 1:95–121.
Zeisel SH. 1987. Choline availability in the neonate. In: Dowdall MJ, Hawthorne JN, eds. Cellular and Molecular Basis of Cholinergic Function. Chichester, England: Horwood. Pp. 709–719.
Zeisel SH. 1993. Choline phospholipids: Signal transduction and carcinogenesis. FASEB J 7:551–557.
Zeisel SH, Blusztajn JK. 1994. Choline and human nutrition. Annu Rev Nutr 14:269– 296.
Zeisel SH, Wurtman RJ. 1981. Developmental changes in rat blood choline concentration. Biochem J 198:565–570.
Zeisel SH, Epstein MF, Wurtman RJ. 1980a. Elevated choline concentration in neonatal plasma. Life Sci 26:1827–1831.
Zeisel SH, Growdon JH, Wurtman RJ, Magil SG, Logue M. 1980b. Normal plasma choline responses to ingested lecithin. Neurology 30:1226–1229.
Zeisel SH, Story DL, Wurtman RJ, Brunengraber H. 1980c. Uptake of free choline by isolated perfused rat liver. Proc Natl Acad Sci USA 77:4417–4419.
Zeisel SH, Stanbury JB, Wurtman RJ, Brigida M, Fierro BR. 1982. Choline content of mothers’ milk in Ecuador and Boston. N Engl J Med 306:175–176.
Zeisel SH, Wishnok JS, Blusztajn JK. 1983. Formation of methylamines from ingested choline and lecithin. J Pharmacol Exp Ther 225:320–324.
Zeisel SH, Char D, Sheard NF. 1986. Choline, phosphatidylcholine and sphingomyelin in human and bovine milk and infant formulas. J Nutr 116:50–58.
Zeisel SH, Zola T, daCosta K, Pomfret EA. 1989. Effect of choline deficiency on S-adenosylmethionine and methionine concentrations in rat liver. Biochem J 259:725–729.
Zeisel SH, daCosta K-A, Franklin PD, Alexander EA, Lamont JT, Sheard NF, Beiser A. 1991. Choline, an essential nutrient for humans. FASEB J 5:2093–2098.
Zeisel SH, Mar M-H, Zhou Z-W, da Costa K-A. 1995. Pregnancy and lactation are associated with diminished concentrations of choline and its metabolites in rat liver. J Nutr 125:3049–3054.
Zeisel SH, Albright CD, Shin O-H, Mar M-H, Salganik RI, da Costa K-A. 1997. Choline deficiency selects for resistance to p53-independent apoptosis and causes tumorigenic transformation of rat hepatocytes. Carcinogenesis 18:731– 738.

































